

Introduction
Brachial plexus root avulsion injuries present a complex problem for the upper extremity surgeon. Over 25 years ago, contralateral C7 root transfer was developed as a treatment option in these difficult cases.1 Since that time, multiple authors have confirmed the use of the C7 transfer procedure.2–5 However, there remains considerable variability in results, depending on factors such as age, time to reconstruction, length of grafts, and the specific target muscle group.5–7 In addition to these factors, it is unclear what role the central nervous system has in the variability of outcomes after peripheral nerve reconstruction, specifically with respect to cortical plasticity.
Previous studies have shown that changes occur in the cerebral cortex after peripheral nerve injury and repair.8,9 Moreover, these changes can be noninvasively observed using blood oxygen level–dependent functional magnetic resonance imaging (fMRI).9,10 Whereas some authors have investigated cortical changes that take place in response to the contralateral C7 nerve transfer procedure, these studies have relied on invasive testing, few time points, or no controls.11–14
Our aim in this study was to perform the C7 transfer procedure in a controlled, survival rat fMRI model to elucidate how both brachial plexus injury and this specific nerve transfer procedure affect the brain. We hypothesized that a survival rat model can be successfully employed and that a high degree of transhemispheric cortical plasticity occurs and continues to adapt over time after contralateral C7 nerve transfer. Moreover, these cortical changes parallel functional recovery.
Materials and Methods
All experiments were approved by the institution's Animal Care and Use Committee and performed in compliance with federal regulations.
Animal preparation
Anesthesia
We performed all surgeries under 2% isoflurane general anesthesia on 200-250-g Sprague-Dawley rats. On completion of the surgery, we transferred each rat to the MRI suite for scanning, during which the isoflurane was discontinued and anesthesia was maintained using subcutaneously administered dexmedetomidine at 100 to 200 μg/kg/h after a 20- to 40-μg bolus.15 Once scanning was complete, we reversed dexmedetomidine with atipamezole. We achieved pain control with 1 injection of 0.05 mg/kg buprenorphine subcutaneously before reversal and dissolved 25 mg tramadol in gelatin, administered daily for 7 days. During all procedures, we continuously monitored pulse oximetry and maintained heart rate and temperature using a thermostatic heating pad. During MRI scanning sessions, additional pulmonary monitoring included respiratory rate and end tidal gases.
Operation
We chose a complete avulsion model because this mirrors clinical indications for C7 nerve transfer (Fig. 1). In addition, this provides certainty that any cortical activation during stimulation is the result of the nerve repair and not intraplexus regeneration or interference from other healthy nerve roots.
Figure 1.
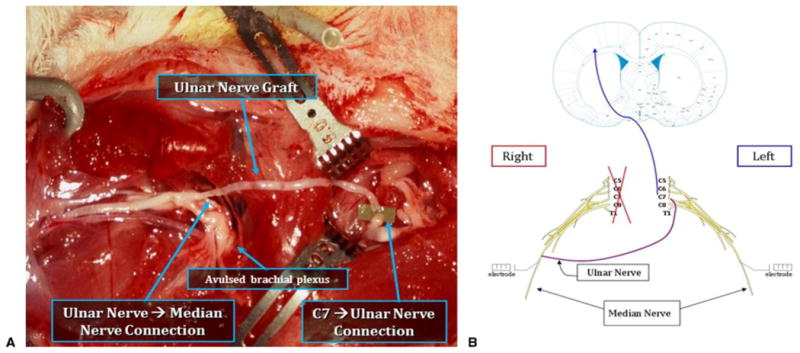
A Group III repair rat with right avulsion and contralateral left C7 nerve transfer. B Schematic illustrating neural pathway after C7 transfer. (Rat brain image adapted with permission from Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. 5th ed. Academic Press; 2004, copyright Elsevier.16)
All animals received 1 dose of 50 mg/kg cefazolin subcutaneously for perioperative antimicrobial prophylaxis. We then clipped the surgical site with an electric razor and prepared it with povidone-iodine. We made a longitudinal incision over the right chest and axilla. Within the axilla, we carried the incision down to the lateral border of the pectoralis major, where we identified the terminal branches of the brachial plexus. Using the operating microscope, we isolated the median nerve branch and atraumatically dissected it free of the surrounding tissue.
At this point, control rats underwent placement of implantable electrodes (MS303-8-A-SPC; Plastics One, Inc, Roanoke, VA) on bilateral median nerves 1 cm proximal to the elbow. Injury rats also underwent placement of implantable electrodes on bilateral median nerves. In addition, in injury rats, we split the right pectoralis major parallel to the muscle fibers and retracted the pectoralis minor superiorly to expose the brachial plexus. We then avulsed nerve roots C5-T1 from the spinal column using forceps; we excised a 1-cm portion of the avulsed roots to prevent intraplexus nerve regeneration. Repair rats also underwent placement of implantable electrodes on bilateral median nerves and right brachial plexus avulsion. After this, we incised the left chest and axilla, split the left pectoralis major, and retracted the pectoralis minor superiorly to expose the left brachial plexus. Next, we harvested the right ulnar nerve and used it as a nonvascularized reversed interposition graft between a division of the left C7 nerve root and the right median nerve. We performed end-to-end repairs using 3 interrupted 11-0 nylon sutures. In all groups, we closed the skin with interrupted 5-0 nylon suture.
The pedestal of the implantable electrodes was left exposed through the skin to allow for connection to the electrical nerve stimulator during MRI scanning. On completion of the scanning period, we closed the skin over the electrode pedestals using 5-0 nylon suture. For later time points, we made an incision over the electrode pedestal and achieved access for stimulation without violating tissues near the original sites of nerve surgery.
Functional MRI parameters, electrical stimulation method, and image analysis
We performed fMRI using a 9.4-T, small-bore animal scanner (AVANCE; Bruker, Billerica, MA) at 0, 3, and 5 months after surgery, to see the blood oxygen level–dependent response to nerve stimulation in the sensory regions of the brain (Fig. 2). We obtained rapid acquisition with relaxation enhancement anatomic images through 10 contiguous, 1-mm slices with slice 7 located directly over the anterior commissure. After obtaining accurate slice positioning, we employed the same rapid acquisition with relaxation enhancement sequence and scanning parameters to acquire 10 slices with the following acquisition parameters: relaxation time of 2,500 ms, echo time of 50.8 ms, average number of 2, field of view of 38.4 × 38.4 mm2, and matrix size of 256 × 256. After the anatomical scan, we took echo-planar imaging scans with the same setup and scanning parameters, except a relaxation time of 2,000 ms, echo time of 18.76 ms, single average, and matrix size of 96 × 96, with the same field of view of 38.4 × 38.4 mm2.
Figure 2.
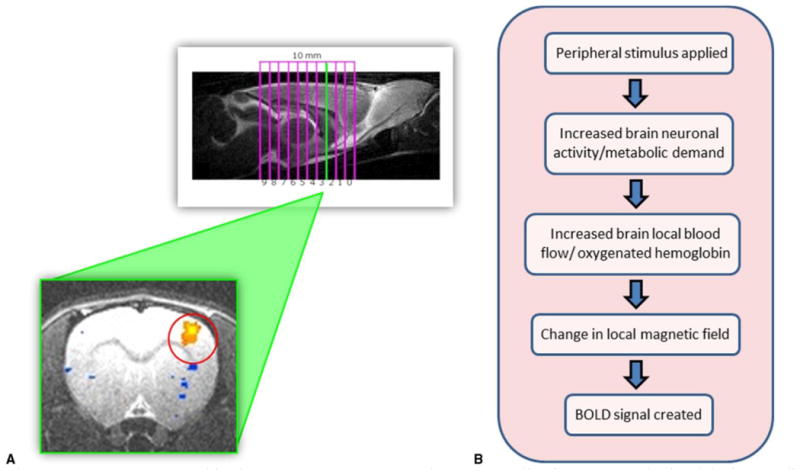
A Rats are placed in the 9.4-T MRI scanner and ten 1-mm slice images are obtained during median nerve stimulation. Positive blood oxygen level– dependent activation signals (orange) represent cortical areas associated with median nerve stimulation. B How blood oxygen level– dependent fMRI works.
During functional imaging, the implantable electrodes on the right and left median nerves were stimulated in sequential order. We achieved stimulation with a square wave electrical generator (S88; Grass Telefactor, Warrick, RI) and performed it at 0.5 mA at 10 Hz for 1 ms. The stimulation protocol for each nerve was 40 seconds of rest followed by 3 periods of 20 seconds on and 40 seconds off, for a total study period of 3 minutes 40 seconds. We chose these parameters based on earlier work from our laboratory.8,9
We registered echoplanar images to an ideal rapid acquisition with relaxation enhancement image using the Oxford Center for Functional Magnetic Resonance of the Brain's Linear Image Registration Tool program.16 We averaged data for each nerve and stimulation protocol and masked them using Analysis of Functional NeuroImages software.17 We determined activation with a P value threshold < .005. Image voxels were classified as active if the statistic was above threshold and counted as activated voxels (3dmaskave) through all slices. We determined voxel color coding by the amplitude of the fit coefficient. Regions of interest were drawn by consulting the activation map and cross-referencing it with a standard rat atlas.18 In this study, we selected bilateral primary somatosensory (S1), secondary somatosensory (S2), and motor (M1/M2) areas as primary regions of interest.
Functional testing
We performed functional testing on the animals concurrent with all fMRI scanning time points at 0, 3, and 5 months (Fig. 3). At time 0, however, we provided a delay of 2 weeks before testing to allow adequate recovery after the initial surgery. One member of the research team was dedicated to performing all functional testing, to exclude the potential for interobserver bias and variability.
Figure 3.

A, B Sensory function is assessed using a Von Frey Esthesiometer. A monofilament is applied to the forepaw through the floor of a wire cage with increasing force until a withdrawal reflex is initiated. C Motor function is assessed using a grip strength meter. The maximum force of pull before forepaw release is measured.
We assessed sensory function using a previously validated19 dynamic plantar Esthesiometer (Ugo Basile, Comerio, Italy). This instrument employs a monofilament applied to the paw to provide increasing force over time until the rat senses the monofilament and exhibits a withdrawal reflex. The amount of force to produce a withdrawal reflex is then recorded. We performed this 5 consecutive times for each rat forepaw in the cutaneous distribution of the median nerve. In groups II and III with the nerve injuries, a rat was determined to have “no response” if we observed no reactive muscle twitching after 5 consecutive attempts with the Esthesiometer. We made additional sensory observations with respect to autonomic sweat function. A qualitative evaluation of forepaw sweat function and nail grooming was performed. Parameters included whether the forepaw was moist and supple or dry and scaly, and whether the nails were adequately groomed.
We assessed motor function using a validated20 rat grip strength meter (Ugo Basile). With this device, a mounted bar is attached to a force meter. The rat is positioned on the bar and is then slowly withdrawn by the tail. Instinctively, the rat grips the bar and the maximum force generated before forepaw release is recorded. We performed this 5 consecutive times with each rat. We made additional motor observations with respect to volitional muscle twitching in the right volar forearm flexor muscles of injury and repair rats during grip strength testing.
Statistical analysis
We performed statistical analysis using unpaired t-test assuming equal variances when comparing 2 groups, and single-factor analysis of variance when comparing multiple groups.
Results
Functional MRI
Figure 4 presents representative image slices comparing all groups across time. At time 0 in control rats, median nerve stimulation elicited a positive blood oxygen level– dependent activation signal in the contralateral forelimb region of the primary somatosensory cortex (S1FL). Injury rats elicited a similar activation signal in the contralateral S1FL after left median nerve stimulation only; no signal was obtained with right median nerve stimulation. These patterns for both groups persisted over 3 and 5 months. During left median nerve stimulation at time 0, repair rats elicited diminished contralateral and weak ipsilateral S1FL activation signals. Over 3 and 5 months, the contralateral signal showed progressive improvement back to baseline, whereas the ipsilateral signal disappeared. At time 0, right median nerve stimulation elicited no signal in either hemicortex. At 3 months, we observed a weak signal in the ipsilateral S1FL and no signal in the contralateral S1FL. At 5 months, we observed a weak signal in the contralateral S1FL and no signal in the ipsilateral S1FL. Individual images of repair rats contained 3 different patterns of S1FL signal activation at 3 and 5 months, which were not appreciated in the group-averaged images (Fig. 5). These patterns included ipsilateral, bilateral, and contralateral S1FL activation, with a progression toward contralateral activation over time.
Figure 4.
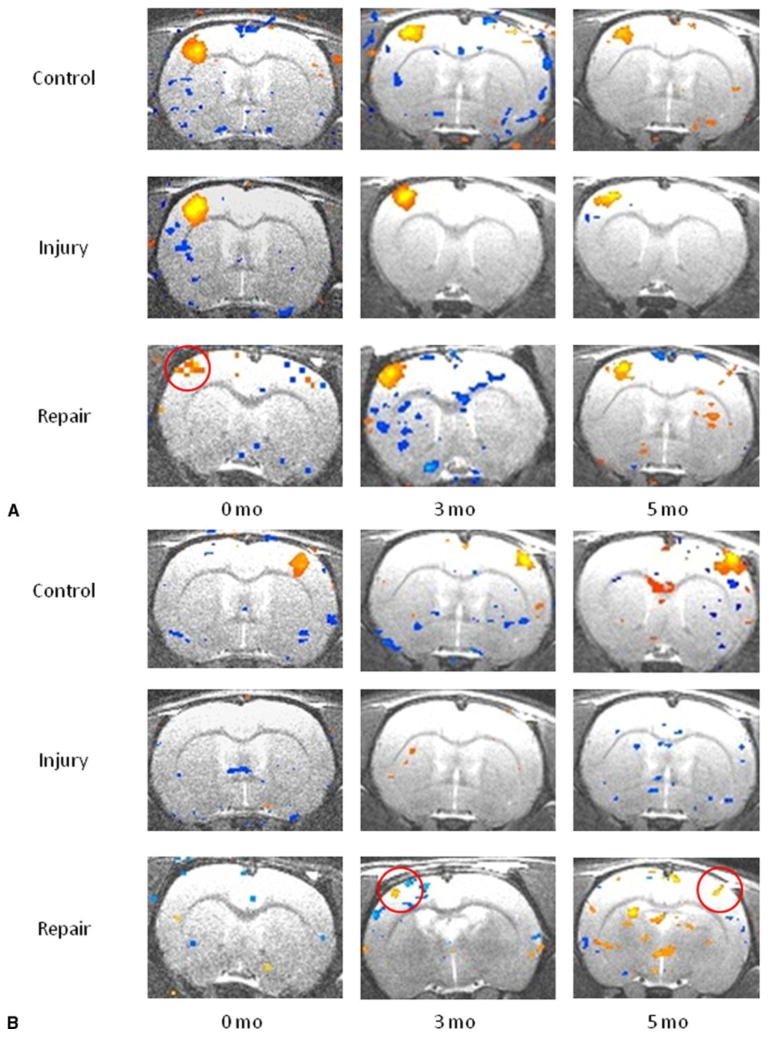
A Averaged blood oxygen level– dependent fMRI images in slice 3 across control, injury, and repair groups after left median nerve stimulation. Positive (orange) blood oxygen level– dependent activation signals are seen in the forelimb region of the primary somatosensory cortex (S1FL). Note diminished blood oxygen level– dependent signal in the repair group at 0 months associated with C7 donation, with recovery by 3 months (red circle). B Averaged blood oxygen level– dependent fMRI images in slice 3 across control, injury, and repair groups after right median nerve stimulation. Positive (orange) activation signals are again seen in S1FL. Note partial recovery of blood oxygen level– dependent signal in the repair group in the ipsilateral S1FL at 3 months and progression to the contralateral S1FL at 5 months.
Figure 5.
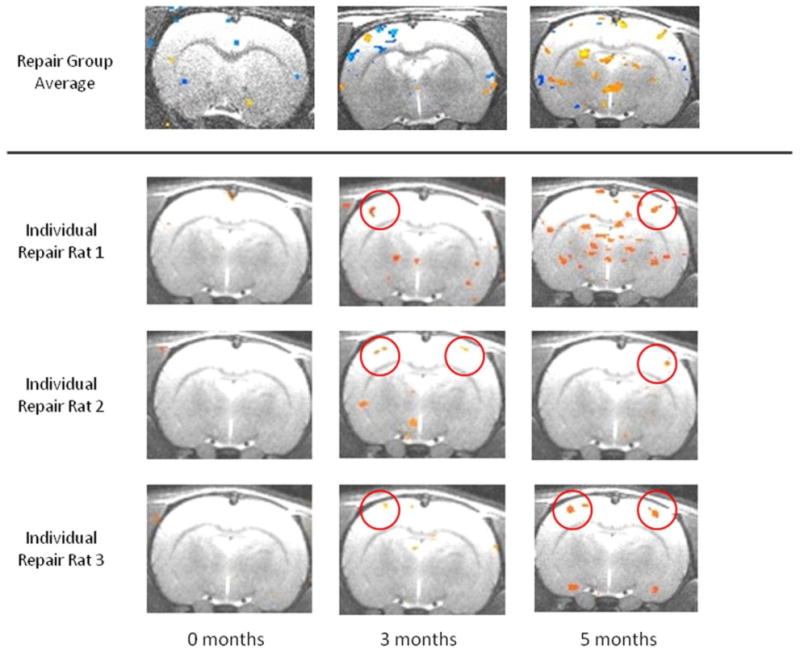
Averaged blood oxygen level– dependent fMRI images in group III repair rats and individual blood oxygen level– dependent fMRI images of 3 representative repair rats after right median nerve stimulation. Analysis of individual repair rats revealed 3 patterns of cortical activation over time. Cortical response tended to revert back to preinjury/repair contralateral S1FL activation.
Sensorimotor testing
Monofilament sensory testing of the left forelimb showed a progressive and statistically significant decrease in sensation in the median nerve forepaw distribution of all 3 groups over time (P < .002, P < .031, and P < .010). At time 0 (2 wk), sensory testing revealed a 21% increase in left forepaw sensitivity in the repair group compared with control rats (P < .053). By 3 months, however, there was no difference in left forepaw sensation between control and repair rats (P < .297). Meaningful monofilament sensory testing of the right forelimb in injury and repair rats was limited by the severity of the plexus injury.
Other indirect assessments of right forelimb sensory function included observations of autonomic sweating and self-grooming. At 3 months, all injury rats exhibited dry, scaly forepaws with poorly groomed claws; 5 of 8 repair rats exhibited moist, supple forepaws with adequately groomed claws. By 5 months, 3 of 8 injury rats and 7 of 8 repair rats exhibited moist, supple forepaws. Figure 6 shows a representative forepaw comparison.
Figure 6.

Autonomic sweat function evaluation. Representative images of control, injury, and repair rat forepaws are shown at 5 months. Control rats exhibited moist, supple forepaws with good nail grooming. Injury rats exhibited dry, scaly forepaws with poor nail grooming. Repair rats exhibited a return to moist, supple forepaws with good nail grooming over time.
Grip strength testing of the control rats showed an overall increase in forepaw strength with time (P < .001). There was no difference in left forepaw grip strength between injury and repair rats at 3 and 5 months (P < .210 and P < .800), which suggests no permanent motor deficit after C7 donation. We made qualitative observations in the right forepaw of injury and repair rats. A proportion of repair rats developed twitching of the right forearm flexor muscles during left forepaw gripping, indicative of some return of motor function. At 3 and 5 months, 3 and 7 of the repair rats, respectively, exhibited such twitching. None of the injury rats exhibited twitching at any time point.
Discussion
Brachial plexus injuries are devastating for patients and pose substantial challenges for surgeons. When nerve repair is precluded by root avulsion, nerve transfer procedures remain the primary option for nerve reconstruction. Although many variables can affect the success of repair,2,5,7 we sought to investigate the role of the central nervous system.
Multiple studies have demonstrated that remodeling of neural networks, termed brain plasticity, occurs after peripheral nerve injury and repair (for a broad overview, see Navarro et al21 and Lundborg22). In nerve transfer procedures, the brain is forced to adapt not only to nerve injury, but also to reassigned functions of donor nerves. We hypothesized that a substantial amount of interhemispheric cortical plasticity occurs after contralateral C7 nerve transfer.
Functional MRI has been widely employed to investigate the brain's response to task-specific use, injury, and intervention. The primary advantage of fMRI is that it is a noninvasive tool that can image the functioning brain and identify active regions associated with particular stimuli. Previous studies have used fMRI to detect the cortical changes after toe-to-thumb transfer23 and hand transplantation.6 Functional MRI has also been used to study the cortical response after peripheral nerve injury and repair.14,24 Moreover, the rat model has been used to study brachial plexus injury and repair.21,22,25,26 In the present study, we explored the cortical changes after contralateral C7 nerve transfer using fMRI in a rat model.
Although there is no standard quantitative measure of cortical plasticity, our results confirm that a substantial amount of transhemispheric cortical plasticity occurs over time after contralateral C7 nerve transfer. At time 0 after complete plexus avulsion, we observed an expected full loss of cortical activation during right median nerve stimulation. At 3 months, however, the neuraxis had been rerouted through the C7 donor nerve pathway, demonstrating ipsilateral cortical activation in S1FL. By 5 months, the cortical responses after contralateral C7 nerve transfer diverged into multiple activation patterns. Divergence in activation patterns may imply a divergence in individual functional recovery during this time. Given the small numbers of animals to compare in these pattern subgroups, we could not draw statistically significant conclusions. All of the activation patterns demonstrated a return of contralateral activation in S1FL over time. This suggests a predilection for the brain to restore the preinjury somatotopic representation of laterality over time. The reason why the brain seeks to restore this representation may be related to efficiency in preserving lateralized sensory networks.
The findings during right median nerve stimulation are in contrast to the results of Wang et al11 for sensory reorganization after contralateral C7 nerve transfer. In their study, they measured cortical activation by direct cortical stimulation of somatosensory evoked potentials and failed to find transhemispheric cortical reorganization in S1FL. Instead, the authors found refinements of the receptive field within the ipsilateral S1FL alone over 14 months. However, work by Pawela et al24 and Pelled et al27 demonstrated early transhemispheric sensory cortical reorganization after peripheral nerve injury without repair. Both conclude that extensive interhemispheric cortical plasticity occurs in the primary somatosensory cortex after peripheral nerve injury. This corroborates our findings of contralateral S1FL activation in the repair group over time. Moreover, multiple studies have shown transhemispheric reorganization of motor cortex after peripheral nerve injury28 and repair.12–14 Although the reasons for these conflicting results are unclear, certainly the work of Pawela et al and Pelled et al, and our results with healthy nerve stimulation, support our findings.
As seen in Figure 4A, left median nerve stimulation in the repair group elicited a weak ipsilateral signal that disappeared by 3 months, an unexpected finding of the study. Some authors24,27 have shown that after denervation, transient ipsilateral activation from the uninjured limb can occur in the first 2 weeks after injury. Only a few of the repair rats elicited this signal, and none of the injury rats did. It could be that transient plasticity occurred in all of the injury and repair rats, but only a few rats maintained this ipsilateral signal at time 0 of fMRI scanning (2 wk). Beyond this speculation, however, we do not have a conclusive explanation for these findings.
Both the intensity and the area of the activation signals diminished over time in all 3 groups, including controls. This may be the result of progressive scarring around the implanted electrodes. Scarring would lead to increased impedance and dampening of conducted electrical signals, with resultant decreased central neuronal activation.
Additional observations of sensorimotor function illustrate the qualitative correspondence between functional outcome and cortical adaptation. Our ability to detect accurate sensory thresholds to monofilament stimulation in the injured right forelimb was limited by the fact that the global palsy prevented elicitation of a motor withdrawal reflex. A lack of withdrawal may be misconstrued to be lack of sensation, when in fact it reflects lack of a coordinated functional motor response. For this reason, we assessed sensory function of the injured forelimb indirectly via observations of autonomic sweat function. Increasing recovery of sweat gland function over 3 and 5 months in the repair groups was qualitatively associated with reorganization and activation in the S1FL. With respect to donor limb sensation, Von Frey monofilament testing remained a useful tool and suggested a 21% improvement in sensitivity 2 weeks after C7 donation in the repair group. This paradoxical improvement is possibly related to early compensation and increased overall use of the uninjured limb after plexus avulsion, despite C7 donation. By 3 months, we saw no difference in sensation between control and repair groups. Moreover, grip strength testing of the C7 donor left limb in repair rats showed no difference compared with injury rats' healthy left limbs at 3 and 5 months. These results concur with clinical observations of slight morbidity and reliable recovery after C7 donation.3,5,29,30
Although a successful survival model using fMRI to detect cortical changes after contralateral C7 nerve transfer has been demonstrated, there are limitations to this study. We performed all repairs in group III immediately after injury, in contrast to clinical practice, in which there is often considerable delay between the time of injury and repair. Furthermore, we limited fMRI studies to the sensory cortex with the presumption that observed sensory plasticity implied concurrent motor plasticity. Assessing motor networks directly would require an awake rat brain, which is both more difficult and less controlled. Finally, we were unable to describe quantitative relationships between functional recovery and fMRI images.
The results of this study suggest many avenues of future research. First, are certain patterns of cortical reorganization associated with poor or improved outcomes, and can they be identified using fMRI? Second, can those patterns be modulated via pharmacologic or behavioral interventions? Third, what is the optimal timing of such interventions? Could it be during the time period of variability in cortical activation patterns (between 3 and 5 mo in rats)? Translational research in this area may be the next step in improving outcomes after peripheral nerve injury and repair.
References
- 1.Gu YD, Zhang GM, Chen DS, Yan JG, Cheng XM, Chen L. Seventh cervical nerve root transfer from contralateral healthy side for treatment of brachial plexus root avulsion. J Hand Surg Br. 1992;17(5):518–521. doi: 10.1016/s0266-7681(05)80235-9. [DOI] [PubMed] [Google Scholar]
- 2.Waikakul S, Orapin S, Vanadurongwan V. Clinical results of contralateral C7 root neurotization to the median nerve in brachial plexus injuries with total root avulsion. J Hand Surg Br. 1999;24(5):556–560. doi: 10.1054/jhsb.1999.0264. [DOI] [PubMed] [Google Scholar]
- 3.Songcharoen P, Wongtrakul S, Mahaisavariya B, Spinner RJ. Hemicontralateral C7 transfer to median nerve in the treatment of root avulsion brachial plexus injury. J Hand Surg Am. 2001;26(6):1058–1064. doi: 10.1053/jhsu.2001.27764. [DOI] [PubMed] [Google Scholar]
- 4.Chen L, Gu YD, Hu SN, Xu JG, Xu L, Fu Y. Contralateral C7 transfer for the treatment of brachial plexus root avulsions in children: a report of 12 cases. J Hand Surg Am. 2007;32(1):96–103. doi: 10.1016/j.jhsa.2006.05.013. [DOI] [PubMed] [Google Scholar]
- 5.Terzis JK, Kokkalis ZT. Selective contralateral C7 transfer in posttraumatic brachial plexus injuries: a report of 56 cases. Plast Reconstr Surg. 2009;123(3):927–938. doi: 10.1097/PRS.0b013e31819ba48a. [DOI] [PubMed] [Google Scholar]
- 6.Dubernard JM, Petruzzo R, Lanzetta M, et al. Functional results of the first human double-hand transplantation. Ann Surg. 2003;238(1):128–136. doi: 10.1097/01.SLA.0000078945.70869.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gu YD, Chen DS, Zhang GM, et al. Long-term functional results of contralateral C7 transfer. J Reconstr Microsurg. 1998;14(1):57–59. doi: 10.1055/s-2007-1006902. [DOI] [PubMed] [Google Scholar]
- 8.Cho YR, Jones SR, Pawela CP, et al. Cortical brain mapping of peripheral nerves using functional magnetic resonance imaging in a rodent model. J Reconstr Microsurg. 2008;24(8):551–557. doi: 10.1055/s-0028-1088231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cho YR, Pawela CP, Li R, et al. Refining the sensory and motor ratunculus of the rat upper extremity using fMRI and direct nerve stimulation. Magn Reson Med. 2007;58(5):901–909. doi: 10.1002/mrm.21408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chen W, Ogawa S. Principle of BOLD-functional MRI. In: Moonen CTW, Bandettini PA, editors. Functional MRI. Berlin: Spinger-Verlag; 1999. pp. 103–113. [Google Scholar]
- 11.Wang M, Li ZY, Xu WD, Hua XY, Xu JG, Gu YD. Sensory restoration in cortical level after a contralateral C7 nerve transfer to an injured arm in rats. Neurosurgery. 2010;67(1):136–143. doi: 10.1227/01.NEU.0000370603.45342.6B. [DOI] [PubMed] [Google Scholar]
- 12.Zuo CT, Hua XY, Guan YH, Xu WD, Xu JG, Gu YD. Long-range plasticity between intact hemispheres after contralateral cervical nerve transfer in humans. J Neurosurg. 2010;113(1):133–140. doi: 10.3171/2010.1.JNS09448. [DOI] [PubMed] [Google Scholar]
- 13.Jiang SU, Li ZY, Hua XY, Xu WD, Xu JG, Gu YD. Reorganization in motor cortex after brachial plexus avulsion injury and repair with the contralateral C7 root transfer in rats. Microsurgery. 2010;30(4):314–320. doi: 10.1002/micr.20747. [DOI] [PubMed] [Google Scholar]
- 14.Beaulieu JY, Blustajn J, Teboul F, et al. Cerebral plasticity in crossed C7 grafts of the brachial plexus: an fMRI study. Microsurgery. 2006;26(4):303–310. doi: 10.1002/micr.20243. [DOI] [PubMed] [Google Scholar]
- 15.Pawela CP, Biswal BB, Hudetz AG, et al. A protocol for use of medetomidine anesthesia in rats for extended studies using task- induced BOLD contrast and resting-state functional connectivity. Neuroimage. 2009;46(4):1137–1147. doi: 10.1016/j.neuroimage.2009.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Oxford Center for Functional Magnetic Resonance of the Brain; [Accessed April 25, 2012]. Linear Image Registration Tool [Computer program] Available: http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/ [Google Scholar]
- 17.Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res. 1996;29(3):162–173. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- 18.Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. 5th. New York: Academic Press; 2004. [Google Scholar]
- 19.Galtrey CM, Fawcett JW. Characterization of tests of functional recovery after median and ulnar nerve injury and repair in the rat forelimb. J Periph Nerv Syst. 2007;12(1):11–27. doi: 10.1111/j.1529-8027.2007.00113.x. [DOI] [PubMed] [Google Scholar]
- 20.Tos P, Ronchi G, Nicolino S, et al. Employment of the mouse median nerve model for the experimental assessment of peripheral nerve regeneration. J Neurosci Methods. 2008;169(1):119–127. doi: 10.1016/j.jneumeth.2007.11.030. [DOI] [PubMed] [Google Scholar]
- 21.Navarro X, Vivo M, Valero-Cabre A. Neural plasticity after peripheral nerve injury and regeneration. Prog Neurobiol. 2007;82(4):163–201. doi: 10.1016/j.pneurobio.2007.06.005. [DOI] [PubMed] [Google Scholar]
- 22.Lundborg G. Brain plasticity and hand surgery: an overview. J Hand Surg Br. 2000;25(3):242–252. doi: 10.1054/jhsb.1999.0339. [DOI] [PubMed] [Google Scholar]
- 23.Chen CJ, Liu HL, Wei FC, Chu NS. fMRI of the human sensorimotor cortex after toe-to-finger transplantation. Am J Neuroradiol. 2006;27(8):1617–1621. [PMC free article] [PubMed] [Google Scholar]
- 24.Pawela CP, Biswal BB, Hudetz AG, et al. Interhemispheric neuroplasticity following limb deafferentation detected by resting-state functional connectivity magnetic resonance imaging (fcMRI) and functional magnetic resonance imaging (fMRI) Neuroimage. 2010;49(3):2467–2478. doi: 10.1016/j.neuroimage.2009.09.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cao XC, Ling LJ. Anatomic basis and technical aspects of a new brachial plexus avulsion injury model in the rat. Plast Reconstr Surg. 2003;111(7):2488–2490. doi: 10.1097/01.PRS.0000063136.86358.12. [DOI] [PubMed] [Google Scholar]
- 26.Gu YD, Ma MK. Nerve transfer for treatment of root avulsion of the brachial plexus: experimental studies in a rat model. J Reconstr Microsurg. 1991;7(1):15–22. doi: 10.1055/s-2007-1006759. [DOI] [PubMed] [Google Scholar]
- 27.Pelled G, Chuang KH, Dodd SJ, Koretsky AP. Functional MRI detection of bilateral cortical reorganization in the rodent brain following peripheral nerve deafferentation. Neuroimage. 2007;37(1):262–273. doi: 10.1016/j.neuroimage.2007.03.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Werhahn KJ, Mortensen J, Van Boven RW, Cohen LG. Bihemispheric plasticity after acute hand deafferentation. Suppl Clin Neurophysiol. 2003;(56):232–241. doi: 10.1016/s1567-424x(09)70227-2. [DOI] [PubMed] [Google Scholar]
- 29.Gu YD. Contralateral C7 root transfer over the last 20 years in China. Chin Med J. 2007;120(13):1123–1126. [PubMed] [Google Scholar]
- 30.Liu J, Pho RW, Kour AK, Zhang AH, Ong BK. Neurologic deficit and recovery in the donor limb following cross-C7 transfer in brachial-plexus injury. J Reconstr Microsurg. 1997;13(4):237–242. doi: 10.1055/s-2007-1000230. [DOI] [PubMed] [Google Scholar]