

Summary
Planarians can regenerate any missing body part in a process requiring dividing cells called neoblasts. Historically, neoblasts have largely been considered a homogeneous stem cell population. Most studies, however, analyzed neoblasts at the population rather than the single-cell level, leaving the degree of heterogeneity in this population unresolved. We combined RNA sequencing of neoblasts from wounded planarians with expression screening and identified 33 transcription factors transcribed in specific differentiated cells and in small fractions of neoblasts during regeneration. Many neoblast subsets expressing distinct tissue-associated transcription factors were present, suggesting candidate specification into many lineages. Consistent with this possibility, klf, pax3/7, and FoxA were required for the differentiation of cintillo-expressing sensory neurons, dopamine-β-hydroxylase-expressing neurons, and the pharynx, respectively. Together, these results suggest that specification of cell fate for most-to-all regenerative lineages occurs within neoblasts, with regenerative cells of blastemas being generated from a highly heterogeneous collection of lineage-specified neoblasts.
Highlights
-
•
Forty-one transcription factors are expressed in subsets of planarian neoblasts
-
•
Specific combinations of transcription factors mark different neoblast subsets
-
•
Specific cell-type regeneration failures follow transcription factor RNAi
-
•
The neoblast population contains many specified progenitors after wounding
Planarian neoblasts are dividing cells required for regeneration. Reddien and colleagues find evidence for extensive specialization of neoblasts into distinct subsets expressing different transcription factors. They propose that specification of regenerative cell fate occurs in neoblasts, with neoblasts comprising pluripotent stem cells and a heterogeneous collection of lineage progenitors.
Introduction
Planarians are flatworms capable of regenerating any missing tissue after injury. Regeneration in the planarian Schmidtea mediterranea requires a population of small mesenchymal cells called neoblasts, which are the only dividing cells of the adult animal. Irradiation eliminates neoblasts, blocking regeneration and tissue turnover (Reddien et al., 2005). Following injury, neoblasts rapidly divide throughout the animal, with mitotic numbers peaking at 6 hr after wounding. If the wound requires the replacement of missing tissue, a second peak of neoblast proliferation occurs at 48 hr (Wenemoser and Reddien, 2010). At this time, neoblasts accumulate at the wound site and their progeny form an unpigmented bud of regenerated tissue called the blastema.
Recently, two neoblast models for planarian regeneration have been proposed: the naive and specialized models (Reddien, 2013). The naive model posits that all neoblasts are stem cells with the same potential and are therefore a largely homogeneous population with fate specification occurring only in nondividing neoblast progeny. By contrast, the specialized model predicts that neoblasts involved in producing missing cells have largely restricted fates and are therefore a heterogeneous population containing many different lineage-committed dividing cells.
Neoblasts have frequently been considered as a uniform population of pluripotent stem cells. Indeed, some neoblasts, termed cNeoblasts, are pluripotent stem cells that can rescue tissue homeostasis and regeneration in lethally irradiated animals by single-cell transplantation (Wagner et al., 2011). The abundance of cNeoblasts in the neoblast population, however, is unknown. smedwi-1 encodes a PIWI-family protein that is expressed in all dividing adult planarian cells (Reddien et al., 2005) and is a canonical neoblast marker. All smedwi-1+ cells rapidly disappear within 1 day following irradiation (Eisenhoffer et al., 2008). Some smedwi-1+ cells have been found to express tissue-specific transcription factors required for specification of a few distinct tissues, such as the eye (Lapan and Reddien, 2011, 2012), the nephridia (Scimone et al., 2011), the anterior pole (Scimone et al., 2014), and some neurons (Cowles et al., 2013; Currie and Pearson, 2013; Wenemoser et al., 2012). Expression of these transcription factors is induced in a small number of smedwi-1+ cells following wounding, with only rare neoblasts expressing these transcription factors in intact animals (Cowles et al., 2013; Lapan and Reddien, 2012). These data provide support for the specialized neoblast model for at least several lineages.
Determining whether the specification of most, or all, cell lineages occurs within neoblasts is essential for understanding the cellular basis for planarian regeneration. Specifically, at what cellular step in regeneration is the identity of new cells specified? On the basis of prior results demonstrating the specialization of smedwi-1+ cells for case study tissues, such as the eye, we sought to test the breadth of the specialized neoblast model. Because it is possible that the smedwi-1+cell population contains both dividing cells (neoblasts) and immediate nondividing neoblast progeny cells, we purified S and G2/M phase neoblasts (X1 neoblasts) using fluorescence-activated cell sorting (FACS) and used RNA sequencing (RNA-seq) to identify transcription factors upregulated in X1 neoblasts following wounding. We combined this approach with broad gene expression screening to identify transcription factors expressed in many different tissues and in neoblasts of wounded planarians. Several conserved transcription factors expressed in neoblasts following wounding were required for regeneration of specific cell types, as predicted by the specialized neoblast model. Our results identify a large collection of transcription factors expressed in small subsets of neoblasts at wounds. Together with previous data, these results indicate that the neoblasts that produce regenerative lineages are a highly heterogeneous population consisting of pluripotent stem cells and lineage-restricted progenitors.
Results
X1 Neoblasts from Wounded Planarians Express Tissue-Associated Transcription Factors
Planarians are flatworms with complex internal anatomy (Figure 1A). They possess a branched intestine that connects to a centrally located pharynx (the only opening of the animal); a nervous system with two cephalic ganglia, two ventral nerve cords, sensory neurons including those of the eyes, and many peripheral neurons; an excretory system (a network of ciliated ducts); and body wall muscle fibers. Because a small planarian tissue fragment can regenerate an entire animal, regeneration requires mechanisms for the specification of the many distinct cell types of adult planarians.
Figure 1.
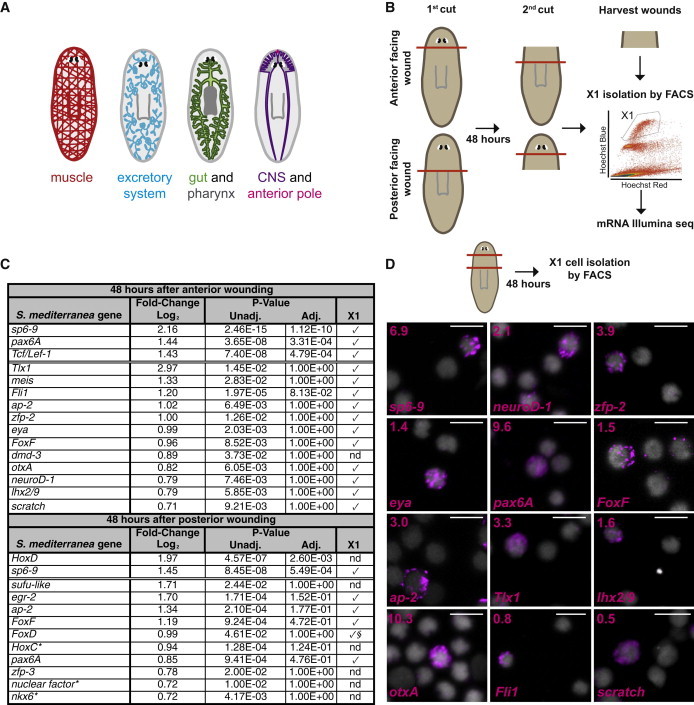
mRNA Sequencing Analysis of Sorted X1 Neoblasts from Regenerating Planarians
(A) Cartoon shows the planarian gastrovascular system with pharynx, the nervous system with photoreceptors, the excretory system, muscle, and the anterior pole.
(B) Schematic of the experimental approach. Prepharyngeal tissues were harvested 48 hr after amputation. Cell suspensions were labeled with Hoechst 33342 and X1 cells isolated by FACS. RNA was then purified from sorted X1 cells and mRNA Illumina sequencing performed.
(C) The table lists all the transcription factors that showed upregulated expression in X1 cells from wounded animals with Padj ≤ 0.05 (above double line) and Punadj ≤ 0.05 with a log2 fold change ≥ 0.7 (below double line). Check marks indicate expression was validated in sorted X1 cells from wounded planarians; nd, not determined. §From published work (Scimone et al., 2014); ∗best BLASTx hit to human transcripts.
(D) Schematic: X1 cells were isolated from prepharyngeal regions by FACS 48 hr after amputations, and cell FISH was performed using RNA probes from genes found in (C). DAPI labeled DNA (gray). Percentages of X1 cells expressing each transcription factor are shown in the upper left corners. Total number of X1 cells counted: sp6-9+ 14/202; neuroD-1+ 22/1034; zfp-2+ 7/180; eya+ 9/635; pax6A+ 107/1113; FoxF+ 3/205; ap-2+ 34/1137; Tlx1+ 5/152; lhx2/9+ 20/1222; otxA+ 16/155; Fli1+ 11/1463; scratch+ 4/766. Images shown are maximal-intensity projections. Scale bars, 10 μm.
Transcription factors can orchestrate the differential expression of regulatory and structural genes needed for cell differentiation in development. In planarians, several transcription factors are expressed in neoblasts for the specification of neoblast fate. In order to establish whether expression of transcription factors in neoblasts following wounding is a hallmark of the specification of most regenerative cell lineages, we first utilized FACS to purify neoblasts that were in the S or G2/M phases of the cell cycle (the X1 fraction: DNA content of more than 2C) (Hayashi et al., 2006) of the prepharyngeal region 48 hr following either head (posterior-facing wound) or trunk (anterior-facing wound) amputations. We subjected these neoblasts to RNA-seq analysis (Figure 1B). This experimental design allowed identification of genes activated during regeneration and controlled for natural variability in gene expression across the anterior-posterior axis by using the same (prepharyngeal) neoblast population. Expression in X1 neoblasts would confirm these transcription factors are active in cells that undergo one or more cell divisions. A few transcription factors were significantly (Padj [adjusted p value] ≤ 0.05) upregulated in X1 neoblasts following wounding (Figure 1C). We also examined (and tested below) transcription factors that were upregulated with a fold-change of log2 ≥ 0.7 and Punadj (unadjusted p value) ≤ 0.05 (Figure 1C; Table S1 available online). By these criteria, neoblasts from wounded planarians expressed the previously described genes sp6-9, which is expressed in optic cup cells and peripheral neurons, and required for eye regeneration (Lapan and Reddien, 2011); meis and otxA, which are expressed in and required for the regeneration of photoreceptor neurons (Lapan and Reddien, 2012); eya, which is expressed in and required for regeneration of eyes and protonephridia (Lapan and Reddien, 2011, 2012; Scimone et al., 2011); and ap-2, which is expressed in multiple neuron types and required for TrpA+ neuron regeneration (Wenemoser et al., 2012) (Figure 1C). In addition, we identified several transcription factor homologs that were expressed in X1 neoblasts 48 hr following wounding (Figures 1C, S1, and S2A; Table S1). We examined the expression of some of these transcription factor homologs (7 out of 13 genes were tested). In wild-type and irradiated uninjured animals, most (six out of seven genes; zfp-2 did not show expression above background) were primarily expressed in differentiated cell types rather than in neoblasts (Figure S1C). To assess the expression in neoblasts, we isolated X1 cells of prepharyngeal regions at 48 hr following both anterior and posterior amputation and performed cell fluorescence in situ hybridizations (FISH). All of the tested transcription factors identified from this RNA-seq analysis (unknown and previously reported, 15 out of 23 genes were tested) were expressed in small subsets of X1 cells from wounded animals (Figures 1D, 2A, 3, and S1D).
Figure 2.
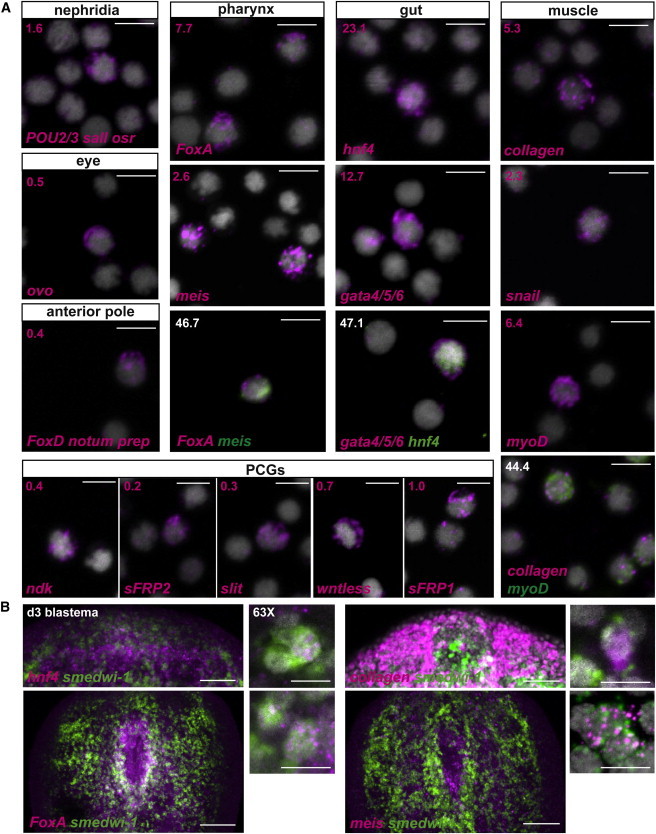
A Candidate Gene Approach Identified Tissue-Associated Transcription Factors Expressed in X1 Neoblasts from Regenerating Planarians
(A) FISH using previously known and new tissue-associated transcription factors with sorted X1 cells from prepharyngeal regions of amputated planarians. Percentages of X1 cells expressing the transcription factors are shown in the upper left corner. Total number of X1 cells counted: for protonephridia, a mixture of RNA probes POU2/3, odd-skipped related (osr), and sal-like (sall) n = 8/504; for the eye, ovo n = 1/194; for the anterior pole, a mixture of the RNA probes FoxD, prep, and notum n = 1/250; for pharynx, FoxA n = 54/704 and meis n = 20/777, with FoxA and meis coexpression observed in n = 7/15 of meis+ X1 cells; for the gut, hnf4 n = 337/1459 and gata4/5/6 n = 665/5249, with coexpression observed in n = 56/119 of hnf4+ X1 cells; for the muscle, collagen n = 72/1357 and the transcription factors myoD n = 47/732 and snail n = 45/1945, with myoD and collagen coexpression observed in n = 20/45 myoD+ X1 cells. Positional control genes were also expressed in wounded X1 cells: nou-darake (ndk, n = 23/6432), secreted related-frizzled 2 (sFRP-2, n = 9/4402), slit (n = 8/2763), wntless (n = 24/3367), and secreted related-frizzled 1 (sFRP-1; n = 7/732). Because some genes were expressed in multiple cell types, for some tissues, progenitor numbers will be an overestimate. DAPI labeled DNA (gray). Scale bars, 10 μm.
(B) Coexpression of the gut transcription factor hnf4, the muscle gene collagen, and the pharynx transcription factors FoxA and meis (magenta) with the neoblast marker smedwi-1 (green) in day 3 regenerating anterior blastemas or tail fragments (for the pharynx genes). Higher magnification on the right shows cells coexpressing both genes (scale bars, 10 μm). DAPI labeled DNA (gray). All images shown are maximal-intensity projections. Anterior, up. Scale bars, 100 μm.
Figure 3.
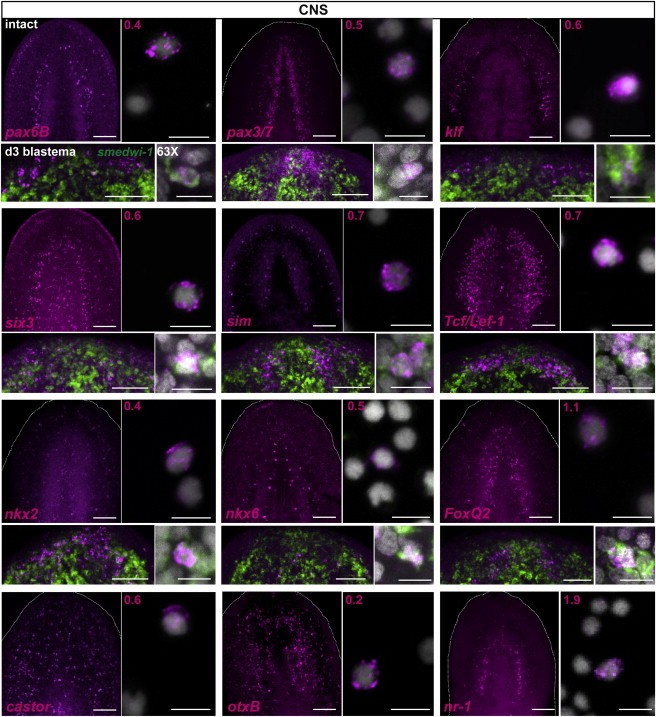
A Broad Panel of CNS-Associated Transcription Factors Was Expressed in X1 Neoblasts
For each gene tested (except last row), the left upper panel shows the expression of the transcription factor (magenta) in the intact head (scale bars, 100 μm), the right upper panel shows expression in sorted X1 cells from prepharyngeal regions of amputated animals 48 hr after wounding (scale bars, 10 μm), the left bottom panel shows the expression of the transcription factor and smedwi-1 (green) in day 3 regenerating anterior blastemas (scale bars, 100 μm), and the bottom right panel shows a higher magnification of a cell coexpressing the transcription factor (magenta) and smedwi-1 (green) in the regenerating blastema (scale bars, 10 μm). DAPI labeled DNA (gray). Percentages of X1 cells expressing the transcription factors are shown in the upper left corner. Numbers of X1 cells counted: pax6B n = 10/2302, pax3/7 n = 12/2299, klf n = 15/2746, six3 n = 11/1826, single minded (sim) n = 1/150, Tcf/Lef-1 n = 3/432, nkx2 n = 9/2435, nkx6 n = 2/395, FoxQ2 n = 2/180, castor n = 11/1954, otxB n = 4/2111, and nr-1 n = 31/1637. Images shown are maximal-intensity projections. Anterior, up.
Known Tissue-Associated Transcription Factors Are Expressed in X1 Neoblasts following Wounding
We reasoned that known tissue-associated transcription factors might be expressed in neoblasts from wounded planarians, but expression of such genes might not be significantly detectable by RNA-seq differential gene expression analysis as a consequence of expression occurring in only rare neoblasts. We therefore analyzed the X1 neoblast expression of transcription factors expressed in distinct tissues. First, we determined the expression frequency of transcription factors previously shown to be associated with distinct tissues and expressed in smedwi-1+ cells using FISH in sorted X1 neoblasts. As expected, a small fraction of X1 cells expressed the nephridia transcription factors POU2/3, sall, and osr (1.6%) (Scimone et al., 2011); the eye-specific transcription factor ovo (0.5%) (Lapan and Reddien, 2012); and the anterior pole transcription factors FoxD and prep and the marker notum (0.4%) (Scimone et al., 2014) (Figure 2A).
Second, we examined the expression of transcription factors known or predicted to be active in the pharynx, gut, and muscle (Figures 2A and S1E). FoxA is a forkhead transcription factor expressed in the endoderm lineage across metazoans. It is expressed in cells that intercalate, polarize, and form tight junctions in the digestive tracts of the mouse, the sea urchin, and the nematode C. elegans (de-Leon, 2011). A planarian FoxA homolog is expressed in the pharynx of the related species Dugesia japonica (Koinuma et al., 2000). We found that during regeneration, FoxA was expressed in the pharynx primordium in the planarian S. mediterranea (Smed-FoxA; Figures S2A, S2B, 2B and 6B). FoxA expression occurred in smedwi-1+ cells and in isolated X1 neoblasts following wounding (7.7% of X1 cells) (Figure 2). The homeobox gene meis was previously shown to be expressed in eyes and required for the regeneration of photoreceptor neurons (Lapan and Reddien, 2012) and was also expressed at the pharynx primordium during regeneration (Figures 2B and S2B). meis was expressed in isolated X1 neoblasts (2.6%) following wounding, and some meis+ X1 neoblasts also coexpressed FoxA (46.7% of meis+ cells, which is higher than expected if expression of these genes in neoblasts was independent; p = 0.0001, Fisher’s exact test), suggesting that both genes might be involved in pharynx specification (Figure 2A). Expression of meis was also observed in smedwi-1+ cells during pharynx regeneration in tail fragments 3 days following wounding (Figure 2B). FoxA and meis were expressed in the pharynx of intact animals (Figure S2C). Finally, a homolog of twist was expressed in the pharynx in intact animals as well as during regeneration (Cowles et al., 2013; Figures S2B and S2C), and it was expressed in smedwi-1+ cells during regeneration (Figure S2B). Moreover, twist was expressed in isolated X1 neoblasts following wounding (2.1%; Figure S2B).
Figure 6.
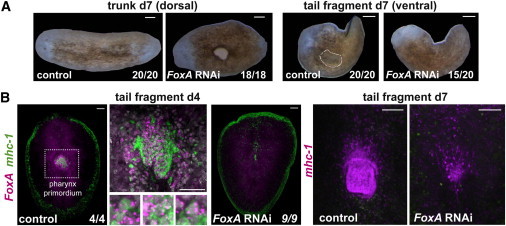
FoxA RNAi Disrupts Pharynx Regeneration
(A) FoxA(RNAi) animals show dorsal lesions near the old pharynx (in trunk pieces, n = 18/18) and do not regenerate a new pharynx (in tail fragments, n = 15/20) 7 days following amputation. Light images show no pharynx opening on the ventral side in regenerating tail fragments of FoxA(RNAi) animals.
(B) FISH using the RNA probes mhc-1 and FoxA shows a defective regenerating pharynx in FoxA(RNAi) tail fragments at 4 and 7 days following amputation. Coexpression of the muscle marker mhc-1 (green) and the pharynx transcription factor FoxA (magenta) is observed in cells of the pharynx primordium in control RNAi animals at 4 days following tail amputation. Right panels: mhc-1 expression (magenta) in control and FoxA RNAi day 7 regenerating tail fragments. DAPI labeled DNA (gray). Images are maximal intensity projections and representative of results seen in more than nine animals per panel. Anterior, up. Scale bars for all panels, 100 μm.
Endodermal transcription factors that control differentiation of the gut and its derivatives during development in multiple species include the hepatocyte nuclear factor (HNF) and GATA families (Zaret, 1999). An HNF homolog, hnf4, and a GATA4/5/6 homolog, gata4/5/6, are both expressed in the gut of S. mediterranea (Wagner et al., 2011). These genes were expressed together in the same cells during gut regeneration (day 3 anterior blastemas; Figure S2D) and were expressed in isolated X1 cells from wounded planarians (hnf4: 23.1% of X1 and gata4/5/6: 12.6%; Figure 2A). Moreover, hnf4 and gata4/5/6 were coexpressed together in the same X1 cells (47.1% of hnf4+ cells; p = 0.0038, Fisher’s exact test; Figure 2A). In addition, hnf4 was expressed in smedwi-1+ cells during gut regeneration (Figure 2B).
Muscle cells develop in most embryos from the mesoderm tissue layer and frequently involve expression of myoD, a well-known myogenic basic-helix-loop-helix transcription factor (Weintraub et al., 1991). Muscle cells are marked by collagen expression in S. mediterranea (Witchley et al., 2013). collagen and myoD (Cowles et al., 2013) were expressed in isolated X1 neoblasts following wounding (myoD: 6.4% and collagen: 5.3%), and 44.4% of myoD+ cells coexpressed collagen (greater than expected by chance: p < 0.0001, Fisher’s exact test; Figure 2A). myoD was also coexpressed with collagen in intact animals (95.1% of myoD+ cells; Figure S2E). In addition, collagen was coexpressed with smedwi-1 in cells of day 3 blastemas (Figure 2B). The Snail gene family encodes zinc finger proteins that can act as transcriptional regulators and control entry into myogenic differentiation (Soleimani et al., 2012). A planarian snail homolog (Figure S1A) was expressed together with collagen in intact animals (83.8% of snail+ cells; Figure S2E). snail was also expressed in X1 neoblasts (2.3%; Figure 2A) and was coexpressed with myoD in X1 cells from wounded animals (35.7%; Figure S2E) and expressed in smedwi-1+cells during regeneration (Figure S2E), suggesting a possible role for this gene in muscle specification.
A different category of genes, the positional control genes (PCGs), described or predicted to have roles in planarian patterning (Reddien, 2011), are highly expressed in planarian muscle cells (Witchley et al., 2013), and are required for the patterning of regenerated tissue. We observed that in addition to this muscle expression, several PCGs were expressed at low frequency in the X1 compartment during regeneration (Figure 2A); these PCGs included slit, a midline-expressed repulsive axon guidance cue; nou-darake (ndk), which is required for proper restriction of the brain to the planarian head; sFRP1 and sFRP2, which are expressed as anterior gradients; and the wound-induced and posteriorly expressed gene wntless (Reddien, 2011). Although these PCGs are not transcription factors, these results further demonstrate the gene expression heterogeneity that exists in neoblasts during regeneration.
Numerous Transcription Factors Expressed in Cells of the Nervous System Are Expressed in Neoblasts following Wounding
Despite the ability of planarians to completely regenerate the nervous system following head amputation, few genes required for the regeneration of specific neuronal lineages have been described (Cowles et al., 2013; Currie and Pearson, 2013; März et al., 2013; Wenemoser et al., 2012). We reasoned that many transcription factors might exist that specify the diverse, numerous cell types in the brain, and the specialized neoblast model predicts many of these would be expressed in neoblasts during regeneration. The nervous system was therefore a good target for testing the specialized neoblast hypothesis.
The neoblast RNA-seq data described above identified upregulation of several transcription factors associated with the nervous system in the neoblast population following wounding (Figure 1C), such as sp6-9, eya, ap-2, pax6A, and otxA (Figure S1C). In addition, expression of other nervous system-associated genes such as Tcf/Lef-1, neuroD-1 (Cowles et al., 2013), and lhx2/9 was observed in neoblasts following wounding (Figure 1C). These genes were indeed expressed in the nervous system of S. mediterranea (Figures S1C, S3A, and 3), as well as in X1 neoblasts from wounded planarians and in smedwi-1+ cells near day 3 regenerating anterior blastemas (Figures 1D, 3, and S3B).
We next searched for conserved transcription factors with roles in nervous system development in other organisms. We found 40 putative transcription factors homologs that were expressed in the planarian brain, coexpressing or closely associated with the neuronal marker cholinergic acetyltransferase (ChAT) gene. From those 40 genes, 18 genes were detectably expressed following wounding either in smedwi-1+ cells in regenerating blastemas and/or in isolated X1 neoblasts of wounded animals (Figures 3, S3B, and S3C). Collectively, from the RNA-seq and candidate approaches, a total of 26 neuron-associated transcription factors displayed expression in neoblasts of regenerating planarians.
Paired-box homeodomain-encoding genes are transcription factors with essential roles in organogenesis, including brain development and patterning (Robson et al., 2006). Planarians have two genes encoding Pax6 homologs, pax6A and pax6B (Figure S4A; Pineda et al., 2002). pax6A was abundantly expressed throughout the brain (Figures S1C and S3A), was significantly upregulated in X1 cells (Figure 1C), and was expressed in isolated X1 cells from wounded planarians (9.6%; Figure 1D). pax6A was also expressed in smedwi-1+ cells during regeneration (Wenemoser et al., 2012; Figure S3B). pax6B expression was most abundant in the lateral cephalic ganglia of the planarian brain in intact animals (Figures 3 and S3A), was present in rare X1 cells from wounded animals (0.4%), and was coexpressed with smedwi-1 during head regeneration (Figure 3). In addition, a planarian gene encoding a homolog of Pax3/7-family transcription factors (Figure S4A) was expressed in the ventral midline of the brain and within the ventral nerve cords (Figures 3 and S3A). pax3/7 was expressed in isolated X1 cells (0.5%) from wounded planarians and coexpressed with smedwi-1+ cells during anterior regeneration (Figure 3).
Drosophila cephalic gap genes have important roles in brain development, as do their orthologs in vertebrates (Kammermeier and Reichert, 2001). Planarians have two homologs of the Drosophila orthodenticle gene, otxA and otxB. Both genes were abundantly expressed in the planarian brain (Figures 3 and S3A; Umesono et al., 1999; Lapan and Reddien, 2011). otxA upregulation in X1 neoblasts was indicated by RNA-seq (Figure 1C), was detected in isolated X1 cells from wounded animals (10.3%; Figure 1D), and was coexpressed with smedwi-1 during anterior regeneration (Figure S3B); otxB was also expressed in X1 cells from wounded animals (0.2%; Figure 3).
The nkx2.2 gene functions in establishing progenitor domains of the ventral spinal cord and hindbrain of vertebrates during embryogenesis (Briscoe et al., 1999). A planarian homolog, nkx2 (Figure S1A), was expressed in the medial planarian brain; cells expressing nkx2 mostly localized to the space between the midline neurons and the lateral region of the cephalic ganglia (Figures 3 and S3A). nkx6.1 is another homeobox gene with important roles in patterning of the ventral vertebrate CNS (Sander et al., 2000). A planarian homolog, nkx6 (Figure S1A), was expressed in scattered cells throughout the medial brain region (Figures 3 and S3A). nkx2 and nkx6 were expressed in smedwi-1+ cells during regeneration and in isolated X1 cells from wounded animals (nkx2, 0.4% and nkx6, 0.5%; Figure 3).
castor encodes a zinc-finger transcription factor expressed in a subset of Drosophila embryonic neuroglioblasts and controls neural differentiation (Mellerick et al., 1992). A planarian castor homolog (Figure S1B) was expressed ventrally in many scattered cells, with some of these cells coexpressing the neuronal marker ChAT (Figure S3A). Following wounding, castor was expressed in 0.6% of X1 neoblasts (Figure 3).
Nuclear receptors are ligand-activated transcription factors that have several roles during development. We found a nuclear receptor (nr-1) gene in S. mediterranea to be expressed in neurons in an inner arc of the brain (Figures 3 and S3A). nr-1 was also expressed in 1.9% of isolated X1 neoblasts from wounded animals (Figure 3).
Several genes found in a transcriptome study of the planarian eye (Lapan and Reddien, 2012) were also expressed in different domains of the brain. For example, the forkhead-family gene FoxQ2, as well as klf, are expressed in the eye and required for photoreceptor neuron regeneration (Lapan and Reddien, 2012). In addition, FoxQ2 was expressed in the ventral midline of the brain and in ventral nerve cords, and klf was expressed in peripheral sensory neurons and in the outer brain branches (Figures 3 and S3A). Both of these genes were also expressed in isolated X1 cells from wounded planarians (1.1% and 0.6%, respectively) and were expressed in smedwi-1+ cells during anterior regeneration (Figure 3).
The HLH-Pas domain transcription factor single minded (sim) is expressed in the planarian brain and is required for the regeneration of several neuron types (Cowles et al., 2013). In agreement with this report, we found expression of sim in isolated X1 neoblasts from wounded planarians (0.7%) and in smedwi-1+ cells in day 3 regenerating anterior blastemas (Figure 3).
An ortholog of the sine oculis/Six family transcription factor six3 (Pineda and Saló, 2002; Wenemoser et al., 2012) was expressed in the lateral cephalic ganglia of the planarian brain and in peripheral neurons (Figures 3 and S3A) and in X1 neoblasts following wounding (0.6%) and was coexpressed with smedwi-1 in day 3 regenerating blastemas (Figure 3). Moreover, genes encoding two SOX proteins (soxB-2 and soxB; Figure S1A; Lapan and Reddien, 2012), as well as the transcription factor-encoding genes glass, lhx3/4, prox-2, and elf-1 (Figures S1A and S1B), were coexpressed with smedwi-1+ cells during anterior regeneration (Figure S3B), and some of these were expressed in isolated X1 cells from wounded planarians (Figure S3C).
In conclusion, the large number of transcription factors expressed in distinct brain regions and in neoblasts from wounded planarians supports a model of neuronal fate specification occurring for most or all lineages within the neoblast compartment during regeneration.
Tissue-Associated Transcription Factors Are Expressed in Nonoverlapping Subsets of Neoblasts from Wounded Planarians
We hypothesized that if the neoblast expression of tissue-associated transcription factors is a result of the specification of distinct lineages, as opposed to reflecting stochastic gene expression or other roles for these genes, only specific combinations of genes would be expressed together in the same neoblast cells. Indeed, for the eye and protonephridia, combinations of transcription factors expressed in differentiated cells are coexpressed in a small number of neoblasts (Lapan and Reddien, 2011, 2012; Scimone et al., 2011). We found multiple other instances of transcription factors (and/or specific markers) expressed together in the same differentiated tissue in intact animals that were coexpressed in the same isolated X1 neoblasts following wounding (Figure 2). Specifically, hnf4 and gata4/5/6 (gut-associated), collagen, myoD, and snail (muscle-associated), and FoxA and meis (pharynx-associated) were expressed together in neoblasts (Figure 2).
To test for the possibility that coexpression of transcription factors could be coincidental, we performed double FISH with isolated X1 cells from wounded planarians using combinations of RNA probes for genes expressed in distinct differentiated cell types. In all such tested cases, we failed to detect any neoblast that displayed coexpression of transcription factors associated with different tissues (taking these results together, these data suggest substantial restriction against coexpression; one-tailed paired t test, p = 0.0064; Figure 4). Specifically, pax6A (neurons) was never detectably coexpressed in the same X1 cell with FoxA (pharynx) or with POU2/3, sall, and osr (nephridia) or with the anterior pole genes FoxD, notum, and prep (Figure 4). Similarly, FoxA was never detectably coexpressed in the same X1 cell together with the nephridia genes, the muscle gene collagen, or the eye gene ovo (Figure 4). Moreover, the gut genes hnf4 and gata4/5/6 were never detectably coexpressed with the neuronal and eye gene sp6-9, the neuronal genes neuroD-1 and pax6A, or the nephridia genes in X1 cells (Figure 4). Together, these results indicate that multiple distinct neoblast populations express specific combinations of transcription factors during regeneration that are associated with differentiated tissues.
Figure 4.
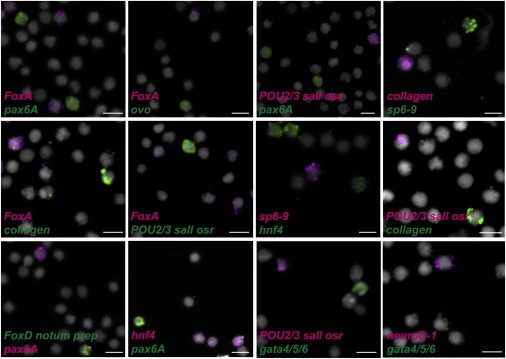
Nonoverlapping Expression of Different Tissue-Associated Genes in X1 Neoblasts
Cell FISH from sorted X1 cells from prepharyngeal regions 48 hr following amputations using several combinations of genes expressed in different tissues. Pharynx: FoxA; CNS: pax6A, neuroD-1, and sp6-9; eye: ovo and sp6-9; protonephridia: POU2/3, sall, osr; muscle: collagen; anterior pole: FoxD, notum, prep, and gut: hnf4 and gata4/5/6. No overlapping expression within the same X1 cells was detected (paired t test one-tailed, p = 0.0064) when compared to expected frequencies. Total number of X1 cells counted: FoxA/pax6A n = 693, FoxA/ovo n = 198, POU2/3/sall/osr/pax6A n = 489, FoxD/prep/notum/pax6A n = 255, collagen/sp6-9 n = 198, collagen/FoxA n = 443, FoxA/POU2/3/sall/osr n = 295, hnf4/sp6-9 n = 448, collagen/POU2/3/sall/osr n = 427, hnf4/pax6A n = 216, POU2/3/sall/osr/gata4/5/6 n = 2422, neuroD-1/gata4/5/6 n = 201. DAPI labeled DNA (gray). Images shown are maximal-intensity projections. Scale bars, 10 μm.
Tissue-Associated Transcription Factors Are Required for Regeneration of Distinct Cell Types
The large collection of transcription factors identified here provides a resource for detailed investigation of different candidate lineage-specification events in planarian regeneration. The specialized neoblast model predicts that most of the transcription factors described above will be essential for the specification of regenerative lineages. Identifying all of such candidate lineages is beyond the scope of this work; however, we sought to test this prediction by the analysis of case studies. To assess the role of some of the identified neoblast-expressed transcription factors in the regeneration of specific cell types, we performed RNAi and analyzed regeneration outcomes. We restricted our analysis to three transcription factors expressed in cell types for which we had other markers as the result of extensive in situ screening: the neuronal expressed genes klf and pax3/7 and the pharynx gene FoxA.
klf was shown to be required for the proper formation of photoreceptor neurons during eye regeneration (Lapan and Reddien, 2012). However, klf had a second expression domain in the outer branches of the planarian head, suggesting a possible role in the differentiation of sensory neurons (Figure 5A). cintillo encodes a protein similar to the degenerin/epithelial superfamily of sodium channels and is expressed in the anterior dorsal margin of the planarian head (Oviedo et al., 2003). We found that neurons expressing klf in the outer head rim also expressed cintillo both in intact and regenerating animal heads (Figures 5A and S4B). pax3/7 was expressed in the ventral midline of the planarian brain (Figure 5A). We identified the planarian homolog of the enzyme dopamine β-hydroxylase (DBH), which is required for the conversion of dopamine into the neurotransmitter norepinephrine. DBH was also expressed in scattered cells on the ventral midline of the planarian brain, and some of these cells coexpressed pax3/7 in intact and regenerating planarians (Figures 5A and S4B).
Figure 5.
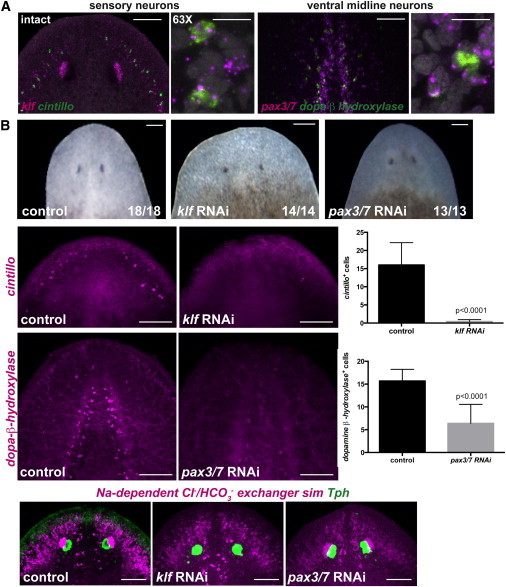
Regeneration Defects of Neural Cell Types following Tissue-Associated Transcription Factor RNAi
(A) Two regions of the brain are shown: the left panel depicts sensory neurons coexpressing the transcription factor klf and the marker cintillo, and the right panel shows ventral midline neurons coexpressing the transcription factor pax3/7 and the enzyme dopamine-β-hydroxylase (scale bars, 100 μm). Higher-magnification panels show coexpression of the genes analyzed in intact head regions (scale bars, 10 μm). DAPI labeled DNA (gray).
(B) Upper row: normal head regeneration (day seven) in control, klf, or pax3/7 RNAi animals. Animals were fed four times followed by head amputation. Second row: cintillo+ cells are not detected in klf(RNAi) animals; graph on the right shows total number of cintillo+ cells (mean ± SD) in regenerating heads (n > 14 animals per RNAi condition; Student’s t test, p < 0.0001). Third row: dopamine-β-hydroxylase-expressing cells are strongly reduced in regenerating pax3/7(RNAi) animals; graph on the right shows total number of dopamine-β-hydroxylase cells (mean ± SD) in regenerating heads (n > 13 animals per RNAi condition; Student-t test, p < 0.0001). Fourth row: dorsal view of control labelings using the RNA probes sim, the outer branches and photoreceptor neuron gene Na+-dependent Cl−/HCO3−, and the trypthophan hydroxylase (Tph) gene, show normal expression of these genes in the regenerating heads of the different RNAi animals. Images are maximal-intensity projections and representative of results seen in more than five animals (except otherwise labeled) per panel. Anterior, up. Scale bars, 100 μm.
klf, pax-3/7, and control RNAi animals regenerated anterior blastemas with largely normal brain formation and patterning (Figures 5B and S4C). Specifically, normal expression of Smed-tryptophan hydroxylase (tph) (Nishimura et al., 2007; Lapan and Reddien, 2012), sim, and a gene expressed in eyes and cephalic branches with homology to a sodium-dependent chloride/bicarbonate exchanger (Na+-dependent Cl−/HCO3−) was observed (Figures 5B and S4C). klf and pax3/7 transcripts were greatly reduced in klf and pax3/7 RNAi animals, respectively (Figure S5D). Despite displaying normal regeneration of many neurons, klf(RNAi) animals had a near-complete absence of cintillo-expressing cells (n = 14/14; Figure 5B) and also lacked photoreceptor neurons (Na+-dependent Cl−/HCO3−-expressing cells) (Lapan and Reddien, 2012), suggesting a specific role for klf in the regeneration of this subset of peripheral sensory neurons. Similarly, pax3/7(RNAi) animals showed significantly reduced numbers of DBH-expressing cells (n = 12/13; Figure 5B), consistent with a role for this gene in the specification of this subset of neurons. In both cases, whether neuron classes are absent or simply not expressing the appropriate genes is unknown.
FoxA was expressed in the pharynx primordium and in neoblasts during pharynx regeneration (Figures 2 and S2B). We inhibited FoxA with RNAi and amputated animals into head, trunk, and tail fragments. Seven days later, all regenerating trunk pieces (which started regeneration with pharynges) had lesions at the pharynx (n = 18/18 animals; Figure 6A). Most tail fragments failed to regenerate a pharynx (n = 15/20; Figure 6A). Because the pharynx is a muscular organ, the mhc-1 muscle marker was used to determine the presence and structure of the pharynx. We found that FoxA (RNAi) animals showed limited pharynx regeneration (n = 9/9), indicating a requirement for FoxA in pharynx specification (Figure 6B). FoxA mRNA levels in FoxA(RNAi) animals were significantly reduced compared to control RNAi animals (Figure S5A). Regenerating tail fragments of FoxA(RNAi) animals 4 days after amputation completely lacked expression of both FoxA and mhc-1 (n = 9/9; Figure 6B). By contrast, control RNAi animals showed FoxA expression together with mhc-1 in the pharynx primordium (Figure 6B). FoxA expression was not exclusively restricted to the pharynx in intact animals (Figure S2C). Similarly, the gut transcription factor hnf4 was expressed in few scattered cells in and around the pharynx (Figure S5B) in intact animals. Cells around the pharynx coexpressed FoxA and hnf4 in intact animals (Figure S5B). Moreover, some isolated X1 cells from wounded animals coexpressed FoxA and hnf4 (27.1% of FoxA+ cells; Figure S5C). Anterior blastemas in FoxA(RNAi) animals showed defects in the regeneration of gut branches (Figure S5C), reflecting either a direct role in anterior gut formation or a secondary effect of failed pharynx regeneration. Animals from cnidarians to humans have FoxA transcription factors that are commonly associated with development of digestive tracts (Mango, 2009). The phenotype observed in FoxA(RNAi) in planarians is reminiscent of the C. elegans pha-4 mutants that lack a pharynx and Drosophila forkhead mutants that lack foregut and hindgut, consistent with an evolutionary ancient role for FoxA transcription factors in foregut/pharynx specification and differentiation (Mango, 2009).
Discussion
Planarian neoblasts have long attracted interest as adult dividing cells that are required for regeneration and cell turnover. A largely unaddressed question has been whether neoblasts are a homogeneous population or are constituted of multiple different cell types, such as stem cells and lineage-committed progenitors. Two models for regeneration have been considered (Reddien, 2013): the naive neoblast model unites all neoblasts as pluripotent stem cells, with fate specification occurring in nondividing neoblast progeny. The specialized neoblast model posits that fate of regenerating cells is specified in neoblasts themselves.
A variety of recent reports have demonstrated instances of smedwi-1+ cell specialization. Transcription factors required for the regeneration of the eye (Lapan and Reddien, 2011, 2012), nephridia (Scimone et al., 2011), the anterior pole (Scimone et al., 2014), and several neuron classes (Cowles et al., 2013; Currie and Pearson, 2013; Wenemoser et al., 2012) are expressed in smedwi-1+ cells during regeneration. These data provide support for the specialized neoblast hypothesis. If this hypothesis is correct, it predicts that numerous additional transcription factors - perhaps for the specification of every missing cell type - would be expressed in distinct subsets of neoblasts during regeneration.
We tested this prediction here by seeking transcription factors expressed in small numbers of X1 neoblasts and/or smedwi-1+ cells following wounding. We coupled an RNA-seq approach with expression screening approaches using transcription factors expressed in specific differentiated planarian tissues. In total, 41 transcription factors (8 known from prior reports and 33 reported here) were expressed in X1 neoblasts and/or smedwi-1+ cells (Figure S1E). A total of 36 out of these 41 transcription factors were detectably expressed in sorted X1 neoblasts from wounded planarians, indicating expression in cells that will complete at least one round of division. These identified transcription factors displayed expression in specific differentiated cells and in small numbers of neoblasts at wound sites, consistent with the specialized neoblast model. Many specific lineages could be investigated to understand the formation of specific cell types during regeneration, and the transcription factors described here provide a resource for such inquiry. In vivo lineage-tracing methods are presently limited in planarians. Therefore, in principle, a transcription factor induced in neoblasts at wounds might have some role in neoblast physiology other than lineage specification. However, two lines of evidence suggest that many or all of these transcription factors will be involved in specifying progenitors for the regeneration of specific differentiated cell types. First, RNAi of a number of identified transcription factors, reported here and in published work, ablate the regeneration of their specific tissues. Here, we demonstrated a requirement for FoxA for the pharynx; pax3/7 for DBH+ ventral midline neurons, and klf for cintillo+ sensory neurons. In prior work, sp6-9, eya, and ovo were required for the eye (Lapan and Reddien, 2011, 2012); six1/2-2, POU2/3, and sall were required for the nephridia (Scimone et al., 2011); FoxD was required for the anterior pole (Scimone et al., 2014); and ap-2, lhx1/5-1, pitx, coe, hesl, and sim were required for different subsets of neurons (Cowles et al., 2013; Currie and Pearson, 2013; Wenemoser et al., 2012). Second, we found that transcription factors that were expressed in different differentiated tissues were expressed in distinct subsets of neoblasts. By contrast, in multiple cases, transcription factors expressed together in the same differentiated tissues were expressed together in the same neoblasts. Some described transcription factors might prove to have roles in the specification of more than one lineage. For instance, klf is required for the specification of both photoreceptor neurons as well as for the sensory cintillo+ neurons (this study and Lapan and Reddien, 2012). Similarly, eya has been shown to be required for the specification of eye progenitors (Lapan and Reddien, 2011) as well as nephridia cells (Scimone et al., 2011).
The concept of neoblast specialization at wounds is important in understanding planarian regeneration and opens many avenues for future inquiry. For example, how is the neoblast response tailored to the identity of missing tissues? Do specialized neoblasts amplify their population, or do specialized neoblasts rapidly cease division and differentiate? There is an ongoing need for differentiated cells in planarians, and low levels of specialized neoblasts appear to exist in intact animals (Cowles et al., 2013; Lapan and Reddien, 2012). For example, the ovo+ eye progenitors are abundant near anterior-facing wounds at 48 hr following amputation, but a small number of them can also be observed near the head of intact animals (Lapan and Reddien, 2012). Similarly, we observed some X1 neoblasts expressing FoxA, klf, and pax3/7 from macerated prepharyngeal regions of uninjured animals, with an increased number present at 48 hr following wounding (Figure S6).
Regeneration involves instructions that specify the identity of missing tissue to be replaced. To understand the connection between regenerative instructions and the production of appropriate cell types, it is essential to address the cellular step at which specification of the fate of regenerative cells occurs. We propose that cNeoblasts (directly, or via their descendants) begin expressing numerous transcription factors of specific lineages in distinct neoblast cells (Figure 7). In this model, almost all of the lineages formed during development could be reconstituted during regeneration, with progenitors that generate and comprise planarian blastemas being a heterogeneous patchwork of lineage-specified cells.
Figure 7.
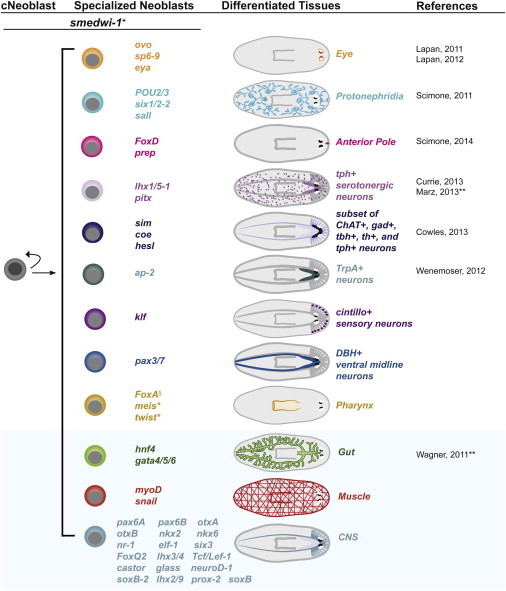
Proposed Model: Specialization of Neoblasts into Different Lineages following Wounding
cNeoblasts will give rise, directly or indirectly, to specialized neoblasts for most-to-all tissues. Both the cNeoblasts and the specialized neoblasts express the smedwi-1 marker. As specialized neoblasts further differentiate, they will lose expression of smedwi-1. A summary of all known transcription factors expressed in neoblasts and functionally associated with distinct lineages by RNAi analysis are shown in the upper part of the model (white background). The transcription factors expressed in neoblasts and in specific tissues, but that have not been functionally shown to be required for their specification, are shown in the lower part of the model (blue background). §During the production phase of this manuscript, the role of FoxA in pharynx regeneration was also described in (Adler et al., 2014). ∗Functional requirement for pharynx specification was not tested. ∗∗Coexpression with the SMEDWI-1 protein was shown. The transcription factors dlx, six1/2, soxB, and otxA, as well as hunchback and eya, have been shown to be required for the regeneration of the eye (Lapan and Reddien, 2011, 2012) and the nephridia (Scimone et al., 2011), but no coexpression with the neoblast gene smedwi-1 or expression in X1 neoblasts has been tested.
Experimental Procedures
Animals, Radiation Treatment, and RNAi
Asexual S. mediterranea (CIW4) animals starved 7–14 days prior experiments were used. Animals were exposed to 6,000 rads of radiation using a dual Gammacell-40 137 cesium source and fixed 3 days after irradiation. Double-stranded RNA-expressing bacteria cultures were mixed with 70% liver solution in a 1:300 ratio. RNAi animals have been fed four times (days 0, 4, 7, and 11) and amputated at day 12. Seven days following amputation, animals were fixed and in situ hybridizations performed. Details of histological methods can be found in Supplemental Experimental Procedures.
mRNA Purification and Illumina Sequencing
Total RNA was purified using TRIzol reagent, and mRNA sequenced using TruSeq RNA Sample Preparation Kit v2 (Illumina) on a HiSeq. Details of RNA-seq analysis can be found in Supplemental Experimental Procedures.
Statistical Analysis
Coexpression frequencies were tested using a two-tailed Fisher’s exact test. Nonoverlapping expression in neoblast subsets was tested using a one-tailed paired t test. Comparisons between RNAi treatments were analyzed using unpaired t tests. Details can be found in Supplemental Experimental Procedures.
Author Contributions
M.L.S., K.M.K., S.W.L., and P.W.R were responsible for the overall study design and analysis. M.L.S. and K.M.K. performed RNAi experiments; K.M.K. and M.L.S. performed X1 FISH; K.M.K. performed X1 RNA sequencing; M.L.S. and S.W.L. performed in vivo neoblast FISH; M.L.S. and S.W.L. performed the neural transcription factor screen; and K.M.K., M.L.S., and S.W.L. performed other transcription factor identification. M.L.S., K.M.K., S.W.L., and P.W.R. wrote the manuscript.
Acknowledgments
P.W.R. is an HHMI Investigator and an associate member of the Broad Institute of Harvard and MIT. We acknowledge support from the NIH (R01GM080639). K.M.K. is supported by National Science Foundation graduate research fellowship grant 1122374. Any opinion, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation. We thank M. Srivastava and J. Meisel for phylogenetic analysis advice, I. Wang for illustrations and cell FISH advice, J. van Wolfswinkel for cell FISH advice and transcriptome analysis, and members of the Reddien lab.
Footnotes
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
Accession Numbers
Illumina sequencing data were deposited to the Gene Expression Omnibus with the accession number GSE57226. Sequences of genes identified in this study were deposited to GenBank with the accession numbers KJ934799–KJ934818.
Supplemental Information
All contigs that follow the criteria for upregulation 48 hours after anterior and posterior amputation used in this study (fold-change expression of log2 ≥ 0.7 with a Padj ≤ 0.05 and Punadj ≤ 0.05, with a BLASTx E-value < 10−5). Known planarian genes were identified by BLASTn to GenBank sequences. Previously unknown planarian transcription factors were defined by best human BLASTx homology, followed by PFAM domain analysis (Figure S1B) and, in some cases, phylogenetics (Figure S1A). The transcriptome is known to be partial redundant, as is in the case of ap-2, which is assembled as two contigs.
References
- Adler C.E., Seidel C.W., McKinney S.A., Sánchez Alvarado A. Selective amputation of the pharynx identifies a FoxA-dependent regeneration program in planaria. eLife. 2014;3:e02238. doi: 10.7554/eLife.02238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Briscoe J., Sussel L., Serup P., Hartigan-O’Connor D., Jessell T.M., Rubenstein J.L., Ericson J. Homeobox gene Nkx2.2 and specification of neuronal identity by graded Sonic hedgehog signalling. Nature. 1999;398:622–627. doi: 10.1038/19315. [DOI] [PubMed] [Google Scholar]
- Cowles M.W., Brown D.D., Nisperos S.V., Stanley B.N., Pearson B.J., Zayas R.M. Genome-wide analysis of the bHLH gene family in planarians identifies factors required for adult neurogenesis and neuronal regeneration. Development. 2013;140:4691–4702. doi: 10.1242/dev.098616. [DOI] [PubMed] [Google Scholar]
- Currie K.W., Pearson B.J. Transcription factors lhx1/5-1 and pitx are required for the maintenance and regeneration of serotonergic neurons in planarians. Development. 2013;140:3577–3588. doi: 10.1242/dev.098590. [DOI] [PubMed] [Google Scholar]
- de-Leon S.B. The conserved role and divergent regulation of foxa, a pan-eumetazoan developmental regulatory gene. Dev. Biol. 2011;357:21–26. doi: 10.1016/j.ydbio.2010.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenhoffer G.T., Kang H., Sánchez Alvarado A. Molecular analysis of stem cells and their descendants during cell turnover and regeneration in the planarian Schmidtea mediterranea. Cell Stem Cell. 2008;3:327–339. doi: 10.1016/j.stem.2008.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi T., Asami M., Higuchi S., Shibata N., Agata K. Isolation of planarian X-ray-sensitive stem cells by fluorescence-activated cell sorting. Dev. Growth Differ. 2006;48:371–380. doi: 10.1111/j.1440-169X.2006.00876.x. [DOI] [PubMed] [Google Scholar]
- Kammermeier L., Reichert H. Common developmental genetic mechanisms for patterning invertebrate and vertebrate brains. Brain Res. Bull. 2001;55:675–682. doi: 10.1016/s0361-9230(01)00559-7. [DOI] [PubMed] [Google Scholar]
- Koinuma S., Umesono Y., Watanabe K., Agata K. Planaria FoxA (HNF3) homologue is specifically expressed in the pharynx-forming cells. Gene. 2000;259:171–176. doi: 10.1016/s0378-1119(00)00426-1. [DOI] [PubMed] [Google Scholar]
- Lapan S.W., Reddien P.W. dlx and sp6-9 Control optic cup regeneration in a prototypic eye. PLoS Genet. 2011;7:e1002226. doi: 10.1371/journal.pgen.1002226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapan S.W., Reddien P.W. Transcriptome analysis of the planarian eye identifies ovo as a specific regulator of eye regeneration. Cell Reports. 2012;2:294–307. doi: 10.1016/j.celrep.2012.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mango S.E. The molecular basis of organ formation: insights from the C. elegans foregut. Annu. Rev. Cell Dev. Biol. 2009;25:597–628. doi: 10.1146/annurev.cellbio.24.110707.175411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- März M., Seebeck F., Bartscherer K. A Pitx transcription factor controls the establishment and maintenance of the serotonergic lineage in planarians. Development. 2013;140:4499–4509. doi: 10.1242/dev.100081. [DOI] [PubMed] [Google Scholar]
- Mellerick D.M., Kassis J.A., Zhang S.D., Odenwald W.F. castor encodes a novel zinc finger protein required for the development of a subset of CNS neurons in Drosophila. Neuron. 1992;9:789–803. doi: 10.1016/0896-6273(92)90234-5. [DOI] [PubMed] [Google Scholar]
- Nishimura K., Kitamura Y., Inoue T., Umesono Y., Yoshimoto K., Takeuchi K., Taniguchi T., Agata K. Identification and distribution of tryptophan hydroxylase (TPH)-positive neurons in the planarian Dugesia japonica. Neurosci. Res. 2007;59:101–106. doi: 10.1016/j.neures.2007.05.014. [DOI] [PubMed] [Google Scholar]
- Oviedo N.J., Newmark P.A., Sánchez Alvarado A. Allometric scaling and proportion regulation in the freshwater planarian Schmidtea mediterranea. Developmental dynamics: an official publication of the American Association of Anatomists. 2003;226:326–333. doi: 10.1002/dvdy.10228. [DOI] [PubMed] [Google Scholar]
- Pineda D., Saló E. Planarian Gtsix3, a member of the Six/so gene family, is expressed in brain branches but not in eye cells. Gene Expr. Patterns. 2002;2:169–173. doi: 10.1016/s0925-4773(02)00341-6. [DOI] [PubMed] [Google Scholar]
- Pineda D., Rossi L., Batistoni R., Salvetti A., Marsal M., Gremigni V., Falleni A., Gonzalez-Linares J., Deri P., Saló E. The genetic network of prototypic planarian eye regeneration is Pax6 independent. Development. 2002;129:1423–1434. doi: 10.1242/dev.129.6.1423. [DOI] [PubMed] [Google Scholar]
- Reddien P.W. Constitutive gene expression and the specification of tissue identity in adult planarian biology. Trends Genet. 2011;27:277–285. doi: 10.1016/j.tig.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddien P.W. Specialized progenitors and regeneration. Development. 2013;140:951–957. doi: 10.1242/dev.080499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddien P.W., Oviedo N.J., Jennings J.R., Jenkin J.C., Sánchez Alvarado A. SMEDWI-2 is a PIWI-like protein that regulates planarian stem cells. Science. 2005;310:1327–1330. doi: 10.1126/science.1116110. [DOI] [PubMed] [Google Scholar]
- Robson E.J., He S.J., Eccles M.R. A PANorama of PAX genes in cancer and development. Nat. Rev. Cancer. 2006;6:52–62. doi: 10.1038/nrc1778. [DOI] [PubMed] [Google Scholar]
- Sander M., Paydar S., Ericson J., Briscoe J., Berber E., German M., Jessell T.M., Rubenstein J.L. Ventral neural patterning by Nkx homeobox genes: Nkx6.1 controls somatic motor neuron and ventral interneuron fates. Genes Dev. 2000;14:2134–2139. doi: 10.1101/gad.820400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scimone M.L., Srivastava M., Bell G.W., Reddien P.W. A regulatory program for excretory system regeneration in planarians. Development. 2011;138:4387–4398. doi: 10.1242/dev.068098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scimone M.L., Lapan S.W., Reddien P.W. A forkhead transcription factor is wound-induced at the planarian midline and required for anterior pole regeneration. PLoS Genet. 2014;10:e1003999. doi: 10.1371/journal.pgen.1003999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soleimani V.D., Yin H., Jahani-Asl A., Ming H., Kockx C.E., van Ijcken W.F., Grosveld F., Rudnicki M.A. Snail regulates MyoD binding-site occupancy to direct enhancer switching and differentiation-specific transcription in myogenesis. Mol. Cell. 2012;47:457–468. doi: 10.1016/j.molcel.2012.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umesono Y., Watanabe K., Agata K. Distinct structural domains in the planarian brain defined by the expression of evolutionarily conserved homeobox genes. Dev. Genes Evol. 1999;209:31–39. doi: 10.1007/s004270050224. [DOI] [PubMed] [Google Scholar]
- Wagner D.E., Wang I.E., Reddien P.W. Clonogenic neoblasts are pluripotent adult stem cells that underlie planarian regeneration. Science. 2011;332:811–816. doi: 10.1126/science.1203983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weintraub H., Davis R., Tapscott S., Thayer M., Krause M., Benezra R., Blackwell T.K., Turner D., Rupp R., Hollenberg S. The myoD gene family: nodal point during specification of the muscle cell lineage. Science. 1991;251:761–766. doi: 10.1126/science.1846704. [DOI] [PubMed] [Google Scholar]
- Wenemoser D., Reddien P.W. Planarian regeneration involves distinct stem cell responses to wounds and tissue absence. Dev. Biol. 2010;344:979–991. doi: 10.1016/j.ydbio.2010.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wenemoser D., Lapan S.W., Wilkinson A.W., Bell G.W., Reddien P.W. A molecular wound response program associated with regeneration initiation in planarians. Genes Dev. 2012;26:988–1002. doi: 10.1101/gad.187377.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witchley J.N., Mayer M., Wagner D.E., Owen J.H., Reddien P.W. Muscle cells provide instructions for planarian regeneration. Cell Reports. 2013;4:633–641. doi: 10.1016/j.celrep.2013.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaret K. Developmental competence of the gut endoderm: genetic potentiation by GATA and HNF3/fork head proteins. Dev. Biol. 1999;209:1–10. doi: 10.1006/dbio.1999.9228. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
All contigs that follow the criteria for upregulation 48 hours after anterior and posterior amputation used in this study (fold-change expression of log2 ≥ 0.7 with a Padj ≤ 0.05 and Punadj ≤ 0.05, with a BLASTx E-value < 10−5). Known planarian genes were identified by BLASTn to GenBank sequences. Previously unknown planarian transcription factors were defined by best human BLASTx homology, followed by PFAM domain analysis (Figure S1B) and, in some cases, phylogenetics (Figure S1A). The transcriptome is known to be partial redundant, as is in the case of ap-2, which is assembled as two contigs.