

Abstract
AIM: To determine the underlying mechanisms of action and influence of Xiaotan Sanjie (XTSJ) decoction on gastric cancer stem-like cells (GCSCs).
METHODS: The gastric cancer cell line MKN-45 line was selected and sorted by FACS using the cancer stem cell marker CD44; the stemness of these cells was checked in our previous study. In an in vitro study, the expression of Notch-1, Hes1, Vascular endothelial growth factor (VEGF), and Ki-67 in both CD44-positive gastric cancer stem-like cells (GCSCs) and CD44-negative cells was measured by Western blot. The effect of XTSJ serum on cell viability and on the above markers was measured by MTT assay and Western blot, respectively. In an in vivo study, the ability to induce angiogenesis and maintenance of GCSCs in CD44-positive-MKN-45- and CD44-negative-engrafted mice were detected by immunohistochemical staining using markers for CD34 and CD44, respectively. The role of XTSJ decoction in regulating the expression of Notch-1, Hes1, VEGF and Ki-67 was measured by Western blot and real-time polymerase chain reaction.
RESULTS: CD44+ GCSCs showed more cell proliferation and VEGF secretion than CD44-negative cells in vitro, which were accompanied by the high expression of Notch-1 and Hes1 and positively associated with tumor growth (GCSCs vs CD44-negative cells, 2.72 ± 0.25 vs 1.46 ± 0.16, P < 0.05) and microvessel density (MVD) (GCSCs vs CD44-negative cells, 8.15 ± 0.42 vs 3.83 ± 0.49, P < 0.001) in vivo. XTSJ decoction inhibited the viability of both cell types in a dose-dependent manner in vitro. Specifically, a significant difference in the medium- (82.87% ± 6.53%) and high-dose XTSJ groups (77.43% ± 7.34%) was detected at 24 h in the CD44+ GCSCs group compared with the saline group (95.42% ± 5.76%) and the low-dose XTSJ group (90.74% ± 6.57%) (P < 0.05). However, the efficacy of XTSJ decoction was reduced in the CD44- groups; significant differences were only detected in the high-dose XTSJ group at 48 h (78.57% ± 6.94%) and 72 h (72.12% ± 7.68%) when compared with the other CD44- groups (P < 0.05). Notably, these differences were highly consistent with the Notch-1, Hes1, VEGF and Ki-67 expression in these cells. Similarly, in vivo, XTSJ decoction inhibited tumor growth in a dose-dependent manner. A significant difference was observed in the medium- (1.76 ± 0.15) and high-dose XTSJ (1.33 ± 0.081) groups compared with the GCSCs control group (2.72 ± 0.25) and the low-dose XTSJ group (2.51 ± 0.25) (P < 0.05). We also detected a remarkable decrease of MVD in the medium- (7.10 ± 0.60) and high-dose XTSJ (5.99 ± 0.47) groups compared with the GCSC control group (8.15 ± 0.42) and the low-dose XTSJ group (8.14 ± 0.46) (P < 0.05). Additionally, CD44 expression was decreased in these groups [medium- (4.43 ± 0.45) and high-dose XTSJ groups (3.56 ± 0.31) vs the GCSC control (5.96 ± 0.46) and low dose XTSJ groups (5.91 ± 0.38)] (P < 0.05). The significant differences in Notch-1, Hes1, VEGF and Ki-67 expression highly mirrored the results of XTSJ decoction in inhibiting tumor growth, MVD and CD44 expression.
CONCLUSION: Notch-1 may play an important role in regulating the proliferation of GCSCs; XTSJ decoction could attenuate tumor angiogenesis, at least partially, by inhibiting Notch-1.
Keywords: Gastric cancer stem-like cells, Xiaotan Sanjie decoction, Tumor angiogenesis, Notch-1, Vascular endothelial growth factor
Core tip: Cancer stem-like cells (CSCs) are involved in tumor angiogenesis and have a higher capacity for secreting vascular endothelial growth factor (VEGF) than non-CSCs. Notch-1 was important in the control of the proliferation of many CSCs; targeting Notch-1 could result in a reduced CSC pool size and decreased levels of VEGF. Our study indicated that CD44+ GCSCs showed superior proliferation and higher VEGF secretion than non-CSCs; these traits were accompanied by high expression of Notch-1. Xiaotan Sanjie decoction could inhibit the proliferation of GCSCs and down-regulate microvessel density in a dose-dependent manner, which could be at least partially explained by its role in manipulating Notch-1.
INTRODUCTION
Gastric cancer is “an enigmatic disease”[1]. Although treatments have greatly improved over the past 20 years, it remains a great therapeutic challenge for oncologists, and the 5-year survival rate is less than 20%[2].
The induction of angiogenesis, a hallmark of cancer, plays an essential role in tumor growth and metastasis[3]. Studies have shown that the vascular endothelial growth factor (VEGF) system is essential and specific to these processes[4]. In mammals, the VEGF system is comprised of five secreted ligands (VEGF-A, VEGF-B, VEGF-C, VEGF-D and PlGF) and three primary receptors (VEGF-R1, VEGF-R2 and VEGF-R3). VEGF-A (commonly referred to as VEGF) and VEGFR-2 constitute the key tissue signals for increased blood vessel growth and/or increased vascular function in various cancers[5]. To date, many inhibitors that block these molecular targets have been used in clinical practice. However, more and more studies indicate that these agents have a limited effect on the overall survival of patients[6]; surprisingly, these drugs may even promote the progression of cancer[7]. Based on this background, some investigators advocate further exploration of the underlying mechanisms that regulate tumor angiogenesis[5].
Cancer stem-like cells (CSCs), defined as “cells within a tumor that possess the capacity for self-renewal and that can cause the heterogeneous lineages of cancer cells that constitute the tumor”[8], have been identified in various cancers[9]. Studies of these cells provide novel insights regarding tumor initiation, progression, recurrence, drug resistance, and angiogenesis[10]. Recent studies indicate that CSCs are involved in tumor angiogenesis via complex mechanisms such as increasing VEGF secretion[11], and it is believed that the existence of CSCs in the tumor bulk is a strong stimulus for VEGF[12]. Gastric cancer stem-like cells (GCSCs) have also been identified in gastric cancer cell lines and primary tumors with the stem cell marker CD44[13,14]. Although data concerning the role of GCSCs in tumor angiogenesis remain rare, previous studies of ovarian cancer indicated that purified CD44-positive cells (also known as ovarian CSCs) possess a similar endothelial potential[15].
Notch signaling plays a fundamental role in controlling the fate of CSCs[16,17] and is believed to be a new target for future cancer treatments[18]. Numerous studies demonstrated that the receptors (Notch 1-4) and/or ligands (Dll-1, Dll-3, Dll-4 and Jag 1-2) of Notch play an important role in different cancers. In particular, Notch-1 has been shown to be associated with cell proliferation in many cancers. Recent studies indicated that Notch-1 is also involved in the control of cell proliferation in CSCs, and targeting Notch-1 could result in the decrease of the number of CSCs[19-21] and levels of VEGF[22]. In the gastrointestinal tract, Notch-1 is also involved in controlling the fate of intestinal stem cells by increasing the expression of a downstream gene, Hes1[23]. Previous studies indicated that the activation of Notch-1 could promote disease progression[22] and that the expression of Notch-1 is significantly higher in cancer cells than in normal tissues[23]. Taking these results into consideration, it is plausible that targeting Notch-1 could have an important role in tumor angiogenesis.
Increasingly, previous data suggest that traditional Chinese medicine can be effective in cancer management by targeting CSCs[24,25]. Xiaotan Sanjie (XTSJ) decoction, a traditional Chinese herbal decoction composed of 11 herbs (Table 1), has been shown to be effective in gastric cancer treatment in China. Previous observations indicated that it is effective in advanced gastric cancer patients[26]. Animal studies demonstrated that it could suppress tumor growth and inhibit angiogenesis via VEGF/KDR signaling[27,28], as well as inhibit the expression of CD44v6 in the gastric cancer cell line MKN-45[29]. In this study, we aimed to investigate the role of XTSJ decoction in GCSCs and the underlying mechanism.
Table 1.
Ingredients of Xiaotan Sanjie decoction and the corresponding percentage (accurate to 1 decimal place)
| Chinese name | Latin name | Percentage |
| Ban-xia | Pinelliae rhizoma | 12.7% |
| Nan-xing | Rhizoma arisaematis | 12.7% |
| Fu-ling | Poria cocos | 12.7% |
| Zhi-shi | Aurantii fructus immaturus | 8.5% |
| Chen-pi | Citri reticulatae viride pericarpium | 7.6% |
| Quan-xie | Scorpio | 5.1% |
| Wu-gong | Scolopendra | 7.6% |
| Ji-nei-jin | Galli gigerii endothelium corneum | 12.7% |
| Bei-mu | Fritillariae cirrhosae bulbus | 7.6% |
| Bai-jie-zi | Semen brassicae | 7.6% |
| Gan-cao | Glycyrrhiza uralensis Fisch | 5.1% |
MATERIALS AND METHODS
Animals
Twenty male Sprague-Dawley rats (weighing 200-250 g) and 56 male nude mice (weighing 20-25 g) were purchased from the Shanghai Institute of Material Medical (Chinese Academy of Science, Shanghai, China) and maintained under standard pathogen-free conditions. All of the rats and mice were handled according to the recommendations of the National Institutes of Health Guidelines for Care and Use of Laboratory Animals. The experimental protocol was approved by the Shanghai Medical Experimental Animal Care Commission.
Cell sorting
The human gastric cancer cell line MKN-45 was purchased from the Shanghai Cells Center, the Chinese Academy of Sciences (Shanghai, China). The GCSC marker CD44 was used in this study. FACS sorting was performed as previously described[13].
Preparation of serum samples containing the tested drugs
All of the herbal medicines were purchased from Shanghai LeiYunShang Co. Ltd., with a known production area. The water extract of the decoction was manufactured as previously described[27,30]. After mixing the herbal components according to the scheduled proportions as listed in Table 1, the herbals were suspended in distilled water for 30 min and then boiled out in a stainless steel pot for 40 min. Then, the drug juice was cooled down to room temperature and filtered using sterile gauze; quality control was performed by high-performance liquid chromatography, as shown in Figure 1. The method of obtaining serum samples containing the tested drugs was described previously[27]. Rats were randomly divided into five groups as follows (n = 5 in each group): control group, 0.9% saline group, and low-, medium- and high-dose XTSJ groups (crude water extracts, which correspond to 1.46, 2.92 and 5.84 g/mL, respectively). The water extract of the decoction and saline were delivered by oral gavage at a frequency of 4 mL per day for 2 wk. Blood samples were aseptically obtained from the abdominal aorta of the rats 2 h after the last administration, and the serum was acquired by centrifugation of the blood at 2000 rpm for 20 min. Following two filtrations through a 0.22-μm cellulose acetate membrane, the serum was bottled, heated in 56 °C water for 30 min and stored at -70 °C.
Figure 1.
High-performance liquid chromatography-DAD of the samples. The samples were randomly tested ten times, and the absorbance was read at 280 nm.
Cell viability and proliferation assay
Cell viability assay was conducted using a MTT kit (Beyotime, China). FACS-sorted CD44+ and CD44- cells were incubated in 96-well plates (5 × 104 cells/well). The following day, the cells were treated with 20 μL of the aforementioned groups of serum samples for 24, 48 or 72 h, with 6 replicates of each treatment. Twenty microliters of MTT reagent was added to each well (5 mg/mL) after incubation, and the cells were then incubated for another 3 h at 37 °C. Cell viability was determined using the following formula: cell viability = (A of the experimental sample/A of the control group) × 100%.
Xenografts and drug administration
Xenografts were created as previously described (Laboratory Animal Center, Second Military Medical University)[31]. The mice were randomized into the following groups: control CD44+ group, control CD44- group, and low-, medium- and high-dose XTSJ decoction groups (crude water extracts corresponding to 1.46, 2.92 and 5.84 g/mL, respectively), which included both CD44+ and CD44- cells (n = 6 in each group). For engraftment, both the CD44+ and CD44- cells were injected subcutaneously into the axilla of the mice using a handmade glass micropipette (1 × 105 cells per site). For drug administration, mice in the control groups received 0.4 mL of 0.9% sodium chloride via oral gavage; the XTSJ decoction groups synchronously received the same dose of the water extracts. Treatment was started 3 d after injection with a frequency of once every 3 d for 8 consecutive weeks. At the end of the 8th week, the mice were euthanized, and the tumors were carefully removed and measured.
Immunohistochemical staining for CD34 and CD44
The tumor tissues were processed by standard histological methods, and immunohistochemical (IHC) tests were performed as previously described[31]. For the measurement of MVD, quantitative vessel counts were performed by the method described by Vermeulen et al[32]. The final value of the IHC test was obtained by combining an estimate of the percentage of immunoreactive cells (quality score) with an estimate of staining intensity[33]. The percentage of immunoreactive cells was determined as follows: no staining = 0; 1%-10% staining = 1; 11%-50% staining = 2; 51%-80% staining = 3 and 81%-100% staining = 4. Staining intensity was rated on the following 0-3 scale: negative = 0, weak = 1, moderate = 2 and strong = 3.
Western blot analysis for Notch-1, Hes1, VEGF and Ki-67
The method of Western blot was previously described[31]. For the in vitro study, cell lysate proteins (40 μg) were separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and electrophoretically transferred to nitrocellulose membranes (Millipore, United States). For the in vivo study, the proteins were extracted from the tissues using RIPA lysis buffer and were separated via 10% SDS-polyacrylamide gel and transferred onto PVDF membranes (Millipore, United States). The primary antibodies used were rabbit anti-Notch-1 (dilution 1:500, Abcam, United States), anti-Hes1 (dilution 1:500, Santa, United States), anti-VEGF (1:1000, Abcam, United States), and anti-Ki-67 (1:500, Abcam, United States). Proteins were detected using the ECL detection reagent, and β-actin was used as a loading control. The results were analyzed using NIH Image J software.
Real-time quantitative polymerase chain reaction assay for Notch-1, Hes1, VEGF and Ki-67
The quantitative polymerase chain reaction (qPCR) method was previously described[31]. Fifty to one hundred milligrams of tissue was used to extract total RNA according to the protocol described for the BioEasy SYBR Green I Real-Time PCR Kit (Bo Ri Technology Co, Ltd, China). The primer sequences for specific gene amplification are shown in Table 2. Fold changes in gene expression were calculated using the 2-ΔΔCt method. The optical density (A) of the target genes were compared with that of GAPDH.
Table 2.
Base sequences of the primers used for real-time quantitative polymerase chain reaction
| Primer name | Sequence |
| Notch1 | |
| Forward | CACTGTGGGCGGGTCC |
| Reverse | GTTGTATTGGTTCGGCACCAT |
| Hes1 | |
| Forward | AGCCAACTGAAAACACCTGATT |
| Reverse | GGAGTTTATGATTAGCAGTGG |
| VEGF-A | |
| Forward | ATGACGAGGGCCTGGAGTGTG |
| Reverse | CCTATGTGCTGGCCTTGGTGAG |
| Ki-67 | |
| Forward | CTTTGGGTGCGACTTGACG |
| Reverse | GTCGACCCCGCTCCTTTT |
| GAPDH | |
| Forward | GGCATCCTGGGCTACACT |
| Reverse | CCACCACCCTGTTGCTGT |
GAPDH: Glyceraldehyde-3-phosphate dehydrogenase.
Statistical analysis
The data were processed using SPSS 18.0 and are presented as mean ± SE. Comparisons between different groups were evaluated by one-way ANOVA, followed by the Bonferroni test or Student’s t-test; ranked data were calculated by nonparametric tests. P < 0.05 was considered statistically significant.
RESULTS
GCSCs show high expression of Notch-1, Hes1 and VEGF
In our previous study, CD44+ GCSCs showed increased cell proliferation, spheroid colony formation, and tumorigenicity compared with CD44- cells[31]. Ki-67 is a useful marker for indicating cell proliferation in normal or cancer stem cells[23,34], and VEGF is closely related to tumor angiogenesis[4,35]. In this study, the Western blot at 48 h after FACS sorting indicated that GCSCs exhibited superior cell proliferation and VEGF secretion compared with CD44- cells, and this result was accompanied by high expression of Notch-1 and Hes1 (Figure 2). Because Notch-1 plays an important role in cell proliferation in CSCs, we speculate that the enhanced cell proliferation of GCSCs was related to the expression of Notch-1.
Figure 2.
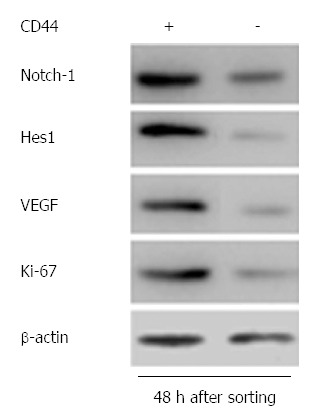
Results of Western blot 48 h after FACS sorting, indicating significant differences in Notch-1, Hes1, VEGF and Ki-67 expression between CD44+ and CD44- cells.
XTSJ decoction inhibits cell viability in a
dose-dependent manner in vitro, most likely by
down-regulating Notch-1
As shown in Figure 3A and B, cell viability analysis found no significant difference between the saline group (95.42% ± 5.76%) and the low-dose group (90.74% ± 6.57%) after incubation (P > 0.05). However, we detected a significant difference in the medium- (82.87% ± 6.53%) and high-dose groups (77.43% ± 7.34%) at 24 h in the CD44+ groups (P < 0.05), indicating that the XTSJ decoction could inhibit cell viability in a dose-dependent manner in GCSCs. Interestingly, the efficacy of XTSJ decoction was reduced in the CD44- groups; we only detected significant differences in the high-dose group at 48 h (78.57% ± 6.94%) and 72 h (72.12% ± 7.68%) when compared with the other CD44- groups (P < 0.05). Using Western blot, we further confirmed the expression of Notch-1, Hes1, VEGF, and Ki-67 at 48 h after different incubations (Figure 3C and D). The Western blot results indicated that the efficacy of XTSJ decoction in the inhibition of cell proliferation and VEGF secretion was accompanied by a down-regulation of Notch-1 and Hes1 (Figure 3E and F) (P < 0.05). These results indicated that XTSJ decoction plays an important role in inhibiting cell proliferation and VEGF secretion in GCSCs, while Notch-1 is involved in the regulation of these processes. However, these conclusions may not be exclusive because some pro-inflammatory cytokines contained in the serum could also affect cell proliferation.
Figure 3.
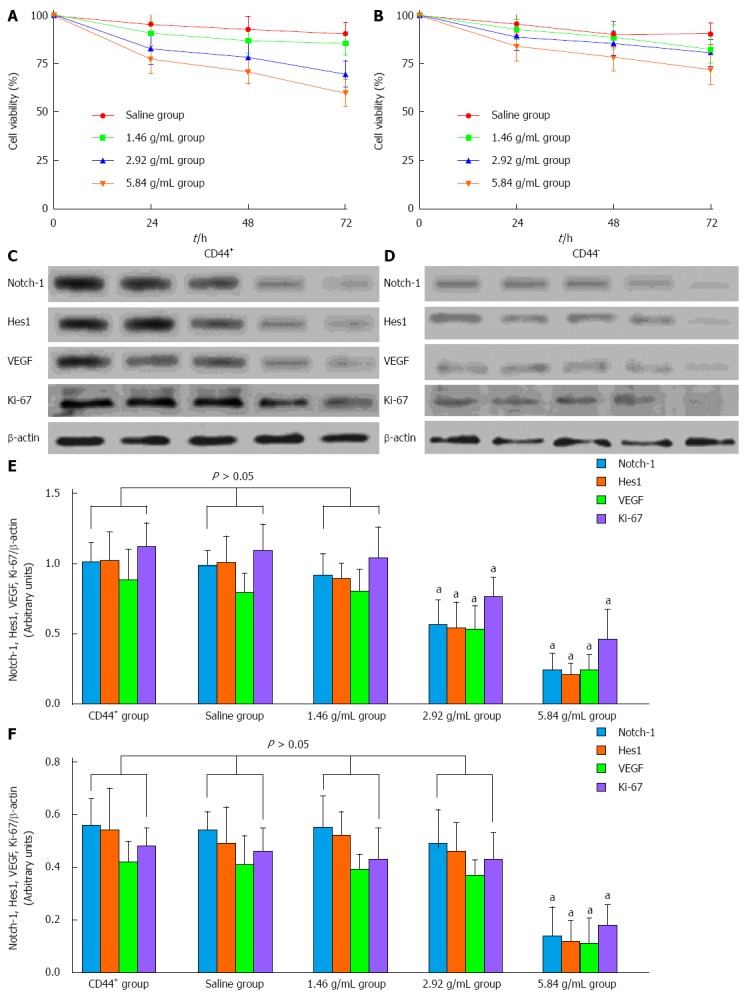
Xiaotan Sanjie decoction inhibits cell viability in a dose-dependent manner. A, B: The effect of Xiaotan Sanjie (XTSJ) decoction on the viability of CD44+ and CD44- cells. The cells were treated with XTSJ decoction for 24, 48 or 72 h. Cell viability was determined by MTT assay. Dots: Mean of six independent experiments; bars: SD; C, D: Results of the Western blot for the expression of Notch-1, Hes1, VEGF, and Ki-67 in the cells incubated with XTSJ decoction for 48 h; E, F: The statistical analysis of the expression of Notch-1, Hes1, VEGF and Ki-67. The densitometry analysis results of the displayed bands were normalized to β-actin; the data are expressed as arbitrary units (one-way ANOVA, followed by the Bonferroni test). aP < 0.05 vs CD44+ group.
GCSCs are superior in microvessel formation, and XTSJ decoction inhibits tumor angiogenesis in a dose-dependent manner
As shown in Figure 4A, tumor weight was significantly different between the control CD44+ group (2.72 ± 0.25) and the control CD44- group (1.46 ± 0.16) at the end of the experiment (P < 0.05). Notably, XTSJ decoction inhibited tumor growth in a dose-dependent manner in the CD44+ groups; significant differences were observed in the medium- (1.76 ± 0.15) and high-dose XTSJ (1.33 ± 0.081) groups compared with the GCSC control and low-dose XTSJ groups (2.51 ± 0.25) (P < 0.05). We also detected a remarkable decrease in MVD in the medium- (7.10 ± 0.60) and high-dose XTSJ (5.99 ± 0.47) groups compared with the GCSC control (8.15 ± 0.42) and low-dose XTSJ groups (8.14 ± 0.46) (P < 0.05). Additionally, CD44 expression in these groups was decreased [medium- (4.43 ± 0.45) and high-dose XTSJ groups (3.56 ± 0.31) vs the GCSC control (5.96 ± 0.46) and low dose XTSJ groups (5.91 ± 0.38)] (P < 0.05). These data indicate that XTSJ decoction could inhibit tumor angiogenesis in a dose-dependent manner. Moreover, we detected no significant differences among the control CD44- group and the low- and medium-dose XTSJ decoction CD44- groups (P > 0.05), although there was a significant difference between the control CD44- group and the high-dose XTSJ decoction group (P < 0.05) (Figure 4B and C). These results indicated that targeting GCSCs plays an important role in inhibiting tumor angiogenesis.
Figure 4.
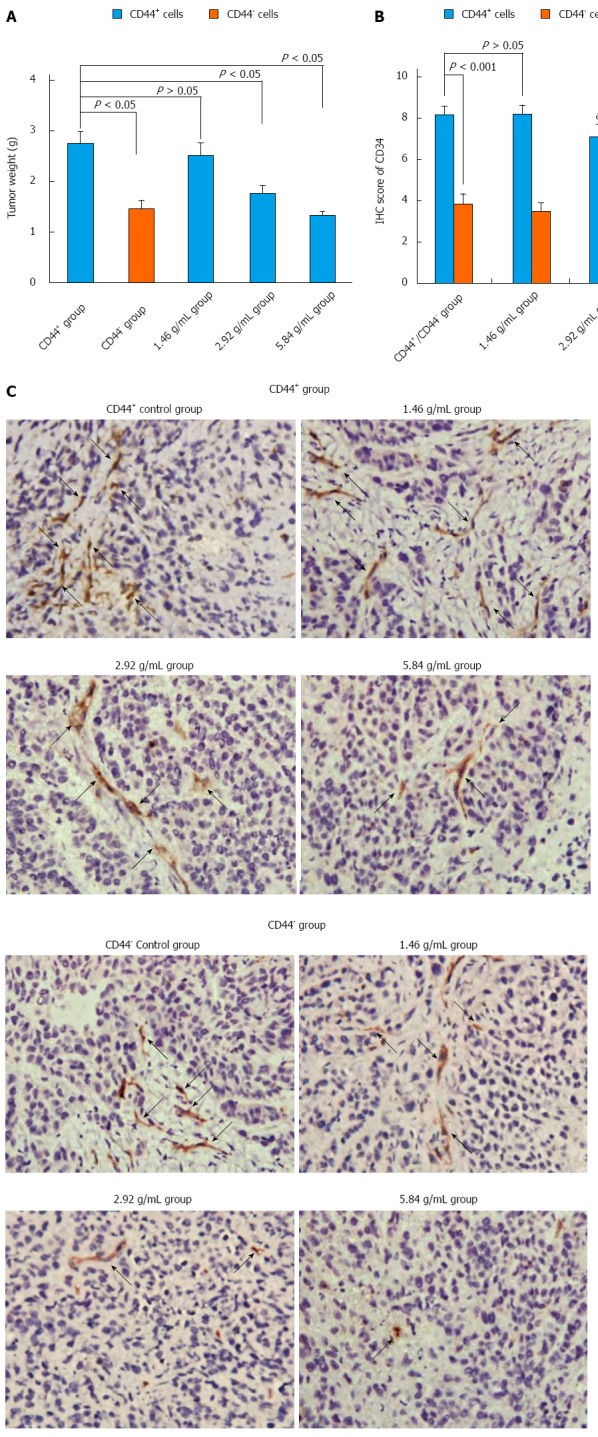
Xiaotan Sanjie decoction inhibits tumor weight and angiogenesis in a dose-dependent manner. A: Tumor weight after eight consecutive weeks of treatment (n = 6 in each group). Control CD44+ group had larger tumors than control CD44- group (Student’s t-test). Treatment with Xiaotan Sanjie (XTSJ) decoction inhibited tumor growth in a dose-dependent manner in gastric cancer stem-like cell transplanted mice (one-way ANOVA, followed by the Bonferroni test); B: Comparison of the microvessel density (MVD) in different groups. The control CD44+ mice showed higher MVD than the control CD44- mice (n = 6, P < 0.05), and XTSJ decoction reduced the MVD in a dose-dependent manner. Statistically significant differences in MVD could be detected in the medium- and high-dose CD44+ groups (Nonparametric tests); C: Immunohistochemical (IHC) results of the MVD. The MVD was defined as a discrete CD34-positive endothelial cell aggregate, with or without definable lumina. (original magnification × 400, positive areas are indicated by black arrows). aP < 0.05 vs CD44+ group; cP < 0.05 vs CD44- group.
In our study, although the control CD44+ group and the CD44+ XTSJ decoction groups were grafted with CD44+ cells, the IHC results indicated that not all of the cells were CD44+ at the end of the experiment, indicating that CD44+ cells were likely to give rise to CD44- cells. The variation of CD44 expression in the XTSJ decoction group was consistent with the results of the MVD (Figure 5A and B), which indicated that these cells are positively associated with tumor angiogenesis.
Figure 5.
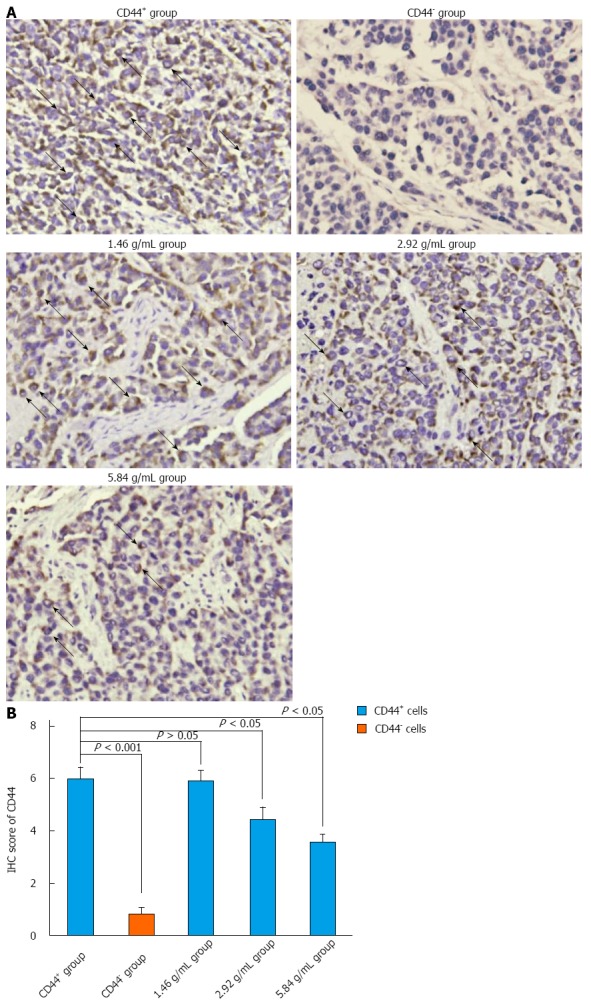
Xiaotan Sanjie decoction inhibits CD44 expression in a dose-dependent manner. A: Immunohistochemical (IHC) staining for CD44. CD44 was highly expressed in the cancer cell membrane (original magnification × 400, positive areas are indicated by black arrows); B: The comparison of CD44 in different groups. CD44 was highly expressed in the control CD44+ group compared with the rest of the groups and was rarely detected in the control CD44- group. The results also showed that not all of the cells in the CD44+ group were CD44 positive. Xiaotan Sanjie decoction inhibited the expression of CD44 in a dose-dependent manner. Statistically significant differences in the expression of CD44 were also detected in the medium- and high-dose groups in CD44+ cells (nonparametric tests).
Capacity of XTSJ decoction to inhibit tumor angiogenesis is related to the down-regulation of Notch-1 and Hes1
As shown in Figure 6A, we detected significant differences in the expression of Notch-1, Hes1, VEGF, and Ki-67 between the control CD44+ and CD44- groups by Western blot. These results indicated that the expression of Notch-1 and Hes1 is positively associated with cell proliferation and tumor angiogenesis. XTSJ decoction inhibited cell proliferation and VEGF secretion in a dose-dependent manner with the down-regulation of Notch-1 and Hes1 (Figure 6B). These results suggest that Notch-1 and Hes1 could be regarded as a potential mechanism for the efficacy of XTSJ decoction. These results were confirmed by qPCR (Figure 6C).
Figure 6.
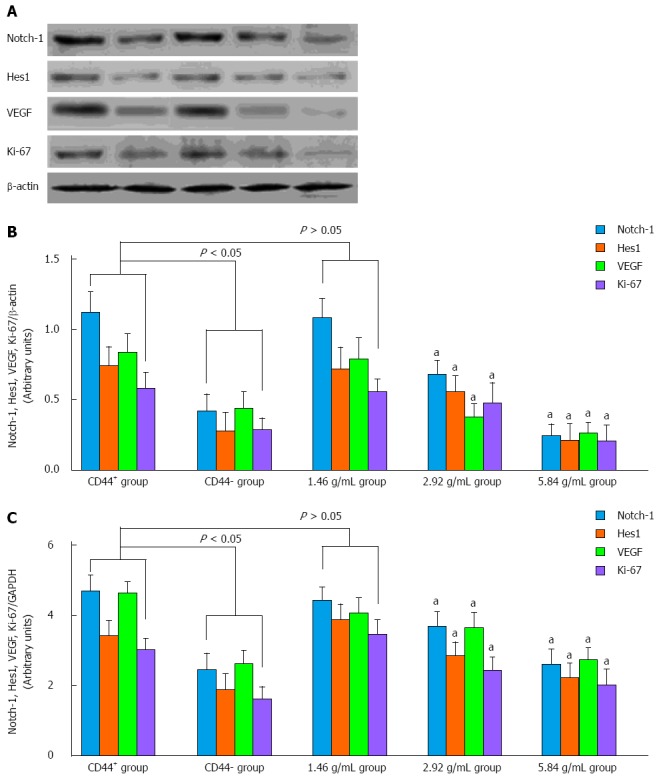
Western blot and real-time quantitative polymerase chain reaction analysis of Notch-1, Hes1, VEGF and Ki-67 expression in vivo. A, B: Western blot revealed that Notch-1 and Hes1 were highly expressed in the control CD44+ group compared with the control CD44- group. Xiaotan Sanjie decoction inhibited the expression of Notch-1 and Hes1 in a dose-dependent manner, and significant differences could be detected in the medium- and high-dose groups (P < 0.05); C: The variation in Notch-1, Hes1, VEGF and Ki-67 expression in the experimental groups was confirmed by quantitative polymerase chain reaction (one-way ANOVA, followed by the Bonferroni test). aP < 0.05 vs CD44+ group.
DISCUSSION
In this study, CD44 positive GCSCs were shown to have higher cell proliferation (in vitro and in vivo) and superior induction of angiogenesis (in vivo) than their counterparts. We also detected remarkably high expression of Notch-1 and its target gene Hes1 in these cells. CD44 is a transcriptional target of Notch-1[36], apparently downstream of Hes1/Hey1[37]. These results, together with previous studies of CSCs and tumor angiogenesis[11,38-40], may indicate that GCSCs could play an important role in angiogenesis, which is at least partially attributable to Notch-1-regulated cell proliferation. In addition, XTSJ decoction was shown to be effective in inhibiting cell viability (in vitro), tumor growth and angiogenesis (in vivo) in a dose-dependent manner in GCSCs; these processes were also accompanied by the down-regulation of Notch-1 and Hes1. Thus, manipulating Notch-1-regulated proliferation of GCSCs could be regarded as a potential gastric cancer treatment in the future.
VEGF has an established role in tumor angiogenesis. Recent studies of CSCs revealed that these cells could secrete markedly elevated levels of VEGF compared with non-CSCs[11,38-40], and targeting these cells may contribute to the inhibition of angiogenesis in the tumor mass. Notch-1 is one of the most important receptors of Notch signaling. However, previous studies indicated that it is a poor prognostic factor for various cancers[41-43], including gastric cancer[44]. Notably, the high expression of Notch-1 is related to enhanced cell proliferation in many cancers[45], and its down-regulation contributes to cell growth inhibition[46,47]. Interestingly, the inhibition of Notch-1 results in the reduction of CSCs[19-21] and a decrease in VEGF[22]. The gastric cancer cell line MKN-45 was previously shown to have a high concentration of GCSCs[13], which further increased the possibility that these cells would act as a major source of VEGF in the tumor. Interestingly, compared to other gastric cell lines, such as MKN-74, MKN-28, and KATO-III, MKN-45 was previously shown to have superior CD44, VEGF and p21 expression[48,49]. XTSJ decoction was shown to be effective in restraining tumor angiogenesis, which was accompanied by a down-regulation of Notch-1. Although related data are still scarce, these results may be regarded as evidence of a possible approach to anti-angiogenesis therapy in the future.
Increasing numbers of studies have indicated that natural derivatives could target CSCs via Notch-1[24,25,50,51]. Interestingly, traditional Chinese medicine has also been shown to be effective for targeting Notch-1 in CSCs[24,25,51]; however, most of these studies were of herbal extracts or single agents, and data for herbal remedies are still rare. The efficacy of XTSJ decoction in gastric cancer treatment has been established by our group. Previous studies indicated that it could inhibit proliferation and induce apoptosis of MKN-45 cells by manipulating genes such as p53 and p21[27,52]. The latter is also down-stream of Notch. In addition, an active ingredient analysis of the herbals that comprise this decoction could further elucidate the reasons for its efficacy. For example, Chen et al[53] demonstrated that the water-soluble polysaccharide isolated from Nan-xing (Rhizoma arisaematis) caused tumor growth inhibition in human breast cancer MCF-7 cells. Chen et al[54] indicated that the extracts isolated from Fu-ling (Poria cocos) possessed anti-proliferative effects on human leukemic U937 and HL-60 cells. Zhang et al[55] demonstrated that the extract of Fu-ling could also induce cell cycle arrest and apoptosis in MCF-7 cells by the premature expression of cell cyclin D1 and cyclin E, as well as Bcl-2. Gapter et al[56] also observed its role in inducing the apoptosis of prostate cancer cells by manipulating p21 and Bcl-2. Kang et al[57] indicated that the extract from Chen-pi (Citri reticulatae viride pericarpium) induced apoptosis in human Snu-C4 colon cancer cells by Bcl-2. Kwon et al[58] reported that the water exact from Quan-xie (Scorpio) induced apoptosis in human hepatoma HepG2 cells. Zhao et al[59] showed that the polysaccharide-protein complex from Wu-gong (Scolopendra) inhibited tumor growth. Chung et al[60] and Kitadai[61] showed that the extracts of Gan-cao (Glycyrrhiza uralensis Fisch) inhibited cell proliferation in MCF-7 human breast cancer cells. Notably, some of the mechanisms described above for XTSJ decoction or its ingredients could participate in complex crosstalk with Notch-1. For example, studies have demonstrated that there is crosstalk between Notch and p53 in cancer growth control[62]. Ling et al[63] observed that Notch-1 signaling could promote the generation of mammary tumor-initiating cells by a cyclin D1-dependent approach. Ye et al[64] indicated that Notch-1 signaling was also involved in Bcl-2 regulated cell growth and apoptosis in cancer. Wang et al[65] and Gao et al[66] also demonstrated that silencing Notch-1 induced apoptosis and increased chemosensitivity in cancer cells related to Bcl-2 and Bax. Based on these results, we speculate that these signaling pathways contribute to the effect of XTSJ decoction in GCSCs. It is also plausible that the effects of XTSJ decoction on Notch-1 would affect other signaling pathways. However, additional studies are needed to verify this surmise.
In conclusion, our study indicated that CD44-positive GCSCs exhibited higher tumor angiogenesis compared with non-CSCs and that Notch-1-regulated cell proliferation plays an important role in these processes. XTSJ decoction inhibits tumor angiogenesis in a dose-dependent manner; the underlying mechanisms may lie in the manipulation of Notch-1-regulated proliferation of GCSCs.
ACKNOWLEDGMENTS
We thank Prof. Jian-Wen Liu at Pharmacy College of East China University of Science and Technology for technical support and constructive advice.
COMMENTS
Background
Emerging evidence indicates that cancer stem-like cells (CSCs) are likely to be involved in tumor angiogenesis and to have a superior capacity to secrete vascular endothelial growth factor (VEGF) than non-CSCs. Notch-1 plays a fundamental role in the control of the proliferation of many CSCs, and targeting Notch-1 could result in a reduced CSC pool size and decreased levels of VEGF. An increasing number of studies have indicated that Traditional Chinese medicine (TCM) could regulate CSCs; however, the relationship of gastric cancer stem-like cells (GCSCs) and angiogenesis as well as the role of TCM in these processes are rarely addressed.
Research frontiers
CSCs have been identified in various cancers in recent years. Studies of these cells have provided novel insights into tumor angiogenesis and have uncovered that these cells are involved in tumor angiogenesis via complex mechanisms such as increasing VEGF secretion. GCSCs have also been identified in gastric cancer cell lines and primary tumors with the stem cell marker CD44; however, data concerning the role of GCSCs in tumor angiogenesis remain rare.
Innovations and breakthroughs
An increasing number of studies have indicated that natural derivatives could target CSCs via Notch-1, which would result in a reduced CSC pool size and decreased levels of VEGF. Interestingly, traditional Chinese medicine has also been shown to be effective in regulating CSCs by targeting Notch-1, but the majority of these studies are of herbal extracts or single agents, and data for herbal decoctions are still rare. In the present study, the authors observed that GCSCs could play an important role in tumor angiogenesis by over-expressing Notch-1. Furthermore, the authors found that the Xiaotan Sanjie decoction could inhibit tumor angiogenesis at least partially by manipulating the Notch-1-regulated proliferation of GCSCs.
Applications
The present study provided additional evidence for the role of CSCs in tumor angiogenesis. In addition, it further indicated that some TCM decoctions could inhibit tumor angiogenesis by manipulating the Notch-1-regulated proliferation of CSCs. All of these results could contribute to the elucidation of the underlying mechanisms of tumor angiogenesis and provide potential therapeutic targets for TCM in cancer in the future.
Terminology
“Cancer stem cells” refers to a subpopulation of cancer cells that possess the capacity for self-renewal and that can cause the heterogeneous lineages of cancer cells that constitute a tumor. Notch signaling is an evolutionarily conserved signaling pathway that consists of four vertebrate Notch receptors (Notch 1-4) and five ligands (Dll-1, Dll-3, Dll-4 and Jag 1-2), and its typical target genes include Hes, Myc, and p21. XTSJ decoction is a traditional Chinese herbal decoction composed of 11 herbs.
Peer review
Tumor angiogenesis plays an essential role in tumor growth and metastasis; however, the majority of currently available anti-angiogenesis-based therapies failed to extensively prolong the overall survival of patients. Therefore, it is perhaps better that the outcome of such treatments should be established by first understanding the underlying mechanisms of tumor angiogenesis. The current study provides additional insights into the role of CSCs in tumor angiogenesis, specifically, in gastric cancer, which could contribute to the understanding of the complex mechanisms of tumor angiogenesis. In addition, it further indicates that some TCMs could attenuate tumor angiogenesis by inhibiting the Notch-1 regulated proliferation of CSCs, which could not only contribute to the elucidation of the potential therapeutic mechanisms of TCMs but also shed light on possible targets for the treatment of CSCs.
Footnotes
Supported by Project of Experimental Animal Research (the Science and Technology Commission of Shanghai), No. 13140901803
P- Reviewer: Abdel-Salam OME S- Editor: Qi Y L- Editor: Wang TQ E- Editor: Ma S
References
- 1.Shah MA, Ajani JA. Gastric cancer--an enigmatic and heterogeneous disease. JAMA. 2010;303:1753–1754. doi: 10.1001/jama.2010.553. [DOI] [PubMed] [Google Scholar]
- 2.Paoletti X, Oba K, Burzykowski T, Michiels S, Ohashi Y, Pignon JP, Rougier P, Sakamoto J, Sargent D, Sasako M, et al. Benefit of adjuvant chemotherapy for resectable gastric cancer: a meta-analysis. JAMA. 2010;303:1729–1737. doi: 10.1001/jama.2010.534. [DOI] [PubMed] [Google Scholar]
- 3.Kakeji Y, Maehara Y, Sumiyoshi Y, Oda S, Emi Y. Angiogenesis as a target for gastric cancer. Surgery. 2002;131:S48–S54. doi: 10.1067/msy.2002.119304. [DOI] [PubMed] [Google Scholar]
- 4.Fondevila C, Metges JP, Fuster J, Grau JJ, Palacín A, Castells A, Volant A, Pera M. p53 and VEGF expression are independent predictors of tumour recurrence and survival following curative resection of gastric cancer. Br J Cancer. 2004;90:206–215. doi: 10.1038/sj.bjc.6601455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Thurston G, Kitajewski J. VEGF and Delta-Notch: interacting signalling pathways in tumour angiogenesis. Br J Cancer. 2008;99:1204–1209. doi: 10.1038/sj.bjc.6604484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li JL, Sainson RC, Shi W, Leek R, Harrington LS, Preusser M, Biswas S, Turley H, Heikamp E, Hainfellner JA, et al. Delta-like 4 Notch ligand regulates tumor angiogenesis, improves tumor vascular function, and promotes tumor growth in vivo. Cancer Res. 2007;67:11244–11253. doi: 10.1158/0008-5472.CAN-07-0969. [DOI] [PubMed] [Google Scholar]
- 7.Ellis LM, Reardon DA. Is there really a yin and yang to VEGF-targeted therapies? Lancet Oncol. 2010;11:809–811. doi: 10.1016/S1470-2045(10)70161-3. [DOI] [PubMed] [Google Scholar]
- 8.Clarke MF, Dick JE, Dirks PB, Eaves CJ, Jamieson CH, Jones DL, Visvader J, Weissman IL, Wahl GM. Cancer stem cells--perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res. 2006;66:9339–9344. doi: 10.1158/0008-5472.CAN-06-3126. [DOI] [PubMed] [Google Scholar]
- 9.Takebe N, Harris PJ, Warren RQ, Ivy SP. Targeting cancer stem cells by inhibiting Wnt, Notch, and Hedgehog pathways. Nat Rev Clin Oncol. 2011;8:97–106. doi: 10.1038/nrclinonc.2010.196. [DOI] [PubMed] [Google Scholar]
- 10.Zhao Y, Bao Q, Renner A, Camaj P, Eichhorn M, Ischenko I, Angele M, Kleespies A, Jauch KW, Bruns C. Cancer stem cells and angiogenesis. Int J Dev Biol. 2011;55:477–482. doi: 10.1387/ijdb.103225yz. [DOI] [PubMed] [Google Scholar]
- 11.Akimoto K, Kimura K, Nagano M, Takano S, To’a Salazar G, Yamashita T, Ohneda O. Umbilical cord blood-derived mesenchymal stem cells inhibit, but adipose tissue-derived mesenchymal stem cells promote, glioblastoma multiforme proliferation. Stem Cells Dev. 2013;22:1370–1386. doi: 10.1089/scd.2012.0486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jordan CT, Guzman ML, Noble M. Cancer stem cells. N Engl J Med. 2006;355:1253–1261. doi: 10.1056/NEJMra061808. [DOI] [PubMed] [Google Scholar]
- 13.Takaishi S, Okumura T, Tu S, Wang SS, Shibata W, Vigneshwaran R, Gordon SA, Shimada Y, Wang TC. Identification of gastric cancer stem cells using the cell surface marker CD44. Stem Cells. 2009;27:1006–1020. doi: 10.1002/stem.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Xue Z, Yan H, Li J, Liang S, Cai X, Chen X, Wu Q, Gao L, Wu K, Nie Y, et al. Identification of cancer stem cells in vincristine preconditioned SGC7901 gastric cancer cell line. J Cell Biochem. 2012;113:302–312. doi: 10.1002/jcb.23356. [DOI] [PubMed] [Google Scholar]
- 15.Alvero AB, Chen R, Fu HH, Montagna M, Schwartz PE, Rutherford T, Silasi DA, Steffensen KD, Waldstrom M, Visintin I, et al. Molecular phenotyping of human ovarian cancer stem cells unravels the mechanisms for repair and chemoresistance. Cell Cycle. 2009;8:158–166. doi: 10.4161/cc.8.1.7533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sikandar SS, Pate KT, Anderson S, Dizon D, Edwards RA, Waterman ML, Lipkin SM. NOTCH signaling is required for formation and self-renewal of tumor-initiating cells and for repression of secretory cell differentiation in colon cancer. Cancer Res. 2010;70:1469–1478. doi: 10.1158/0008-5472.CAN-09-2557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pannuti A, Foreman K, Rizzo P, Osipo C, Golde T, Osborne B, Miele L. Targeting Notch to target cancer stem cells. Clin Cancer Res. 2010;16:3141–3152. doi: 10.1158/1078-0432.CCR-09-2823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Garber K. Notch emerges as new cancer drug target. J Natl Cancer Inst. 2007;99:1284–1285. doi: 10.1093/jnci/djm148. [DOI] [PubMed] [Google Scholar]
- 19.Qiu M, Peng Q, Jiang I, Carroll C, Han G, Rymer I, Lippincott J, Zachwieja J, Gajiwala K, Kraynov E, et al. Specific inhibition of Notch1 signaling enhances the antitumor efficacy of chemotherapy in triple negative breast cancer through reduction of cancer stem cells. Cancer Lett. 2013;328:261–270. doi: 10.1016/j.canlet.2012.09.023. [DOI] [PubMed] [Google Scholar]
- 20.Sharma A, Paranjape AN, Rangarajan A, Dighe RR. A monoclonal antibody against human Notch1 ligand-binding domain depletes subpopulation of putative breast cancer stem-like cells. Mol Cancer Ther. 2012;11:77–86. doi: 10.1158/1535-7163.MCT-11-0508. [DOI] [PubMed] [Google Scholar]
- 21.Wang J, Wang C, Meng Q, Li S, Sun X, Bo Y, Yao W. siRNA targeting Notch-1 decreases glioma stem cell proliferation and tumor growth. Mol Biol Rep. 2012;39:2497–2503. doi: 10.1007/s11033-011-1001-1. [DOI] [PubMed] [Google Scholar]
- 22.Wang Z, Banerjee S, Li Y, Rahman KM, Zhang Y, Sarkar FH. Down-regulation of notch-1 inhibits invasion by inactivation of nuclear factor-kappaB, vascular endothelial growth factor, and matrix metalloproteinase-9 in pancreatic cancer cells. Cancer Res. 2006;66:2778–2784. doi: 10.1158/0008-5472.CAN-05-4281. [DOI] [PubMed] [Google Scholar]
- 23.Fre S, Huyghe M, Mourikis P, Robine S, Louvard D, Artavanis-Tsakonas S. Notch signals control the fate of immature progenitor cells in the intestine. Nature. 2005;435:964–968. doi: 10.1038/nature03589. [DOI] [PubMed] [Google Scholar]
- 24.Zhen Y, Zhao S, Li Q, Li Y, Kawamoto K. Arsenic trioxide-mediated Notch pathway inhibition depletes the cancer stem-like cell population in gliomas. Cancer Lett. 2010;292:64–72. doi: 10.1016/j.canlet.2009.11.005. [DOI] [PubMed] [Google Scholar]
- 25.Sun H, Zhang S. Arsenic trioxide regulates the apoptosis of glioma cell and glioma stem cell via down-regulation of stem cell marker Sox2. Biochem Biophys Res Commun. 2011;410:692–697. doi: 10.1016/j.bbrc.2011.06.060. [DOI] [PubMed] [Google Scholar]
- 26.Lv DL, Wei PK. Clinical observaton of Xiaotan SanJie decoction on 15 advanced gastric cancer patients. Zhonghua Xiaohua Waike Zazhi. 2007;15:401–402. [Google Scholar]
- 27.Gui MW, Wei PK, Lu Y, Guo W, Qin ZF, Sun DZ. [Effects of Xiaotan Sanjie Decoction-containing serum on proliferation and apoptosis of human gastric cancer cells MKN-45] Zhong Xi Yi Jie He Xuebao. 2010;8:250–255. doi: 10.3736/jcim20100309. [DOI] [PubMed] [Google Scholar]
- 28.Xu L, Su XM, Chen YL, Wei PK. Xiaotan Sanjie decoction contribute to the VEGF, KDRmRNA suppression in human gastric cancer in situ transplantation model. Shijie Huaren Xiaohua Zazhi. 2004;12:988–990. [Google Scholar]
- 29.Wang JP, Wei PK, Li YH, Xu L. Xiaotan Sanjie decoction suppress the CD44v6 expression on the MKN-45 grafted nude mice. Chengdu ZhongyiYao Daxue Xuebao. 2001;24:20–21. [Google Scholar]
- 30.Guo XD, Wei PK. Effect of Xiaotan Sanjie decoction on growth of MKN-45 transplanted nude mices and expression of proliferating cell nuclear antigen. Zhongxiyi Jiehe Xuebao. 2007;5:432–436. doi: 10.3736/jcim20070414. [DOI] [PubMed] [Google Scholar]
- 31.Yan B, Zhou Y, Feng S, Lv C, Xiu L, Zhang Y, Shi J, Li Y, Wei P, Qin Z. β -Elemene-Attenuated Tumor Angiogenesis by Targeting Notch-1 in Gastric Cancer Stem-Like Cells. Evid Based Complement Alternat Med. 2013;2013:268468. doi: 10.1155/2013/268468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Vermeulen PB, Gasparini G, Fox SB, Toi M, Martin L, McCulloch P, Pezzella F, Viale G, Weidner N, Harris AL, et al. Quantification of angiogenesis in solid human tumours: an international consensus on the methodology and criteria of evaluation. Eur J Cancer. 1996;32A:2474–2484. doi: 10.1016/s0959-8049(96)00379-6. [DOI] [PubMed] [Google Scholar]
- 33.Soslow RA, Dannenberg AJ, Rush D, Woerner BM, Khan KN, Masferrer J, Koki AT. COX-2 is expressed in human pulmonary, colonic, and mammary tumors. Cancer. 2000;89:2637–2645. doi: 10.1002/1097-0142(20001215)89:12<2637::aid-cncr17>3.0.co;2-b. [DOI] [PubMed] [Google Scholar]
- 34.Rodilla V, Villanueva A, Obrador-Hevia A, Robert-Moreno A, Fernández-Majada V, Grilli A, López-Bigas N, Bellora N, Albà MM, Torres F, et al. Jagged1 is the pathological link between Wnt and Notch pathways in colorectal cancer. Proc Natl Acad Sci U S A. 2009;106:6315–6320. doi: 10.1073/pnas.0813221106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Song ZJ, Gong P, Wu YE. Relationship between the expression of iNOS,VEGF,tumor angiogenesis and gastric cancer. World J Gastroenterol. 2002;8:591–595. doi: 10.3748/wjg.v8.i4.591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Deftos ML, Huang E, Ojala EW, Forbush KA, Bevan MJ. Notch1 signaling promotes the maturation of CD4 and CD8 SP thymocytes. Immunity. 2000;13:73–84. doi: 10.1016/s1074-7613(00)00009-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fischer A, Schumacher N, Maier M, Sendtner M, Gessler M. The Notch target genes Hey1 and Hey2 are required for embryonic vascular development. Genes Dev. 2004;18:901–911. doi: 10.1101/gad.291004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Beck B, Driessens G, Goossens S, Youssef KK, Kuchnio A, Caauwe A, Sotiropoulou PA, Loges S, Lapouge G, Candi A, et al. A vascular niche and a VEGF-Nrp1 loop regulate the initiation and stemness of skin tumours. Nature. 2011;478:399–403. doi: 10.1038/nature10525. [DOI] [PubMed] [Google Scholar]
- 39.Folkins C, Shaked Y, Man S, Tang T, Lee CR, Zhu Z, Hoffman RM, Kerbel RS. Glioma tumor stem-like cells promote tumor angiogenesis and vasculogenesis via vascular endothelial growth factor and stromal-derived factor 1. Cancer Res. 2009;69:7243–7251. doi: 10.1158/0008-5472.CAN-09-0167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sun H, Jia J, Wang X, Ma B, Di L, Song G, Ren J. CD44+/CD24- breast cancer cells isolated from MCF-7 cultures exhibit enhanced angiogenic properties. Clin Transl Oncol. 2013;15:46–54. doi: 10.1007/s12094-012-0891-2. [DOI] [PubMed] [Google Scholar]
- 41.Lin JT, Chen MK, Yeh KT, Chang CS, Chang TH, Lin CY, Wu YC, Su BW, Lee KD, Chang PJ. Association of high levels of Jagged-1 and Notch-1 expression with poor prognosis in head and neck cancer. Ann Surg Oncol. 2010;17:2976–2983. doi: 10.1245/s10434-010-1118-9. [DOI] [PubMed] [Google Scholar]
- 42.Reedijk M, Odorcic S, Chang L, Zhang H, Miller N, McCready DR, Lockwood G, Egan SE. High-level coexpression of JAG1 and NOTCH1 is observed in human breast cancer and is associated with poor overall survival. Cancer Res. 2005;65:8530–8537. doi: 10.1158/0008-5472.CAN-05-1069. [DOI] [PubMed] [Google Scholar]
- 43.Chu D, Li Y, Wang W, Zhao Q, Li J, Lu Y, Li M, Dong G, Zhang H, Xie H, et al. High level of Notch1 protein is associated with poor overall survival in colorectal cancer. Ann Surg Oncol. 2010;17:1337–1342. doi: 10.1245/s10434-009-0893-7. [DOI] [PubMed] [Google Scholar]
- 44.Yeh TS, Wu CW, Hsu KW, Liao WJ, Yang MC, Li AF, Wang AM, Kuo ML, Chi CW. The activated Notch1 signal pathway is associated with gastric cancer progression through cyclooxygenase-2. Cancer Res. 2009;69:5039–5048. doi: 10.1158/0008-5472.CAN-08-4021. [DOI] [PubMed] [Google Scholar]
- 45.Purow BW, Haque RM, Noel MW, Su Q, Burdick MJ, Lee J, Sundaresan T, Pastorino S, Park JK, Mikolaenko I, et al. Expression of Notch-1 and its ligands, Delta-like-1 and Jagged-1, is critical for glioma cell survival and proliferation. Cancer Res. 2005;65:2353–2363. doi: 10.1158/0008-5472.CAN-04-1890. [DOI] [PubMed] [Google Scholar]
- 46.Wang M, Wu L, Wang L, Xin X. Down-regulation of Notch1 by gamma-secretase inhibition contributes to cell growth inhibition and apoptosis in ovarian cancer cells A2780. Biochem Biophys Res Commun. 2010;393:144–149. doi: 10.1016/j.bbrc.2010.01.103. [DOI] [PubMed] [Google Scholar]
- 47.Xiao X, Ning L, Chen H. Notch1 mediates growth suppression of papillary and follicular thyroid cancer cells by histone deacetylase inhibitors. Mol Cancer Ther. 2009;8:350–356. doi: 10.1158/1535-7163.MCT-08-0585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ninomiya S, Inomata M, Tajima M, Ali AT, Ueda Y, Shiraishi N, Kitano S. Effect of bevacizumab, a humanized monoclonal antibody to vascular endothelial growth factor, on peritoneal metastasis of MNK-45P human gastric cancer in mice. J Surg Res. 2009;154:196–202. doi: 10.1016/j.jss.2008.08.017. [DOI] [PubMed] [Google Scholar]
- 49.Yokozaki H. Molecular characteristics of eight gastric cancer cell lines established in Japan. Pathol Int. 2000;50:767–777. doi: 10.1046/j.1440-1827.2000.01117.x. [DOI] [PubMed] [Google Scholar]
- 50.Lin L, Liu Y, Li H, Li PK, Fuchs J, Shibata H, Iwabuchi Y, Lin J. Targeting colon cancer stem cells using a new curcumin analogue, GO-Y030. Br J Cancer. 2011;105:212–220. doi: 10.1038/bjc.2011.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Li Y, Zhang J, Ma D, Zhang L, Si M, Yin H, Li J. Curcumin inhibits proliferation and invasion of osteosarcoma cells through inactivation of Notch-1 signaling. FEBS J. 2012;279:2247–2259. doi: 10.1111/j.1742-4658.2012.08607.x. [DOI] [PubMed] [Google Scholar]
- 52.Guo XD, Wei PK. [Effect of Xiaotan Sanjie Recipe on growth of transplanted tumor and expressions of proliferating cell nuclear antigen and epidermal growth factor receptor in tissue of gastric carcinoma of nude mice] Zhong Xi Yi Jie He Xue Bao. 2007;5:432–436. doi: 10.3736/jcim20070414. [DOI] [PubMed] [Google Scholar]
- 53.Chen G, Xu J, Miao X, Huan Y, Liu X, Ju Y, Han X. Characterization and antitumor activities of the water-soluble polysaccharide from Rhizoma Arisaematis. Carbohydr Polym. 2012;90:67–72. doi: 10.1016/j.carbpol.2012.04.063. [DOI] [PubMed] [Google Scholar]
- 54.Chen YY, Chang HM. Antiproliferative and differentiating effects of polysaccharide fraction from fu-ling (Poria cocos) on human leukemic U937 and HL-60 cells. Food Chem Toxicol. 2004;42:759–769. doi: 10.1016/j.fct.2004.01.018. [DOI] [PubMed] [Google Scholar]
- 55.Zhang M, Chiu LC, Cheung PC, Ooi VE. Growth-inhibitory effects of a beta-glucan from the mycelium of Poria cocos on human breast carcinoma MCF-7 cells: cell-cycle arrest and apoptosis induction. Oncol Rep. 2006;15:637–643. [PubMed] [Google Scholar]
- 56.Gapter L, Wang Z, Glinski J, Ng KY. Induction of apoptosis in prostate cancer cells by pachymic acid from Poria cocos. Biochem Biophys Res Commun. 2005;332:1153–1161. doi: 10.1016/j.bbrc.2005.05.044. [DOI] [PubMed] [Google Scholar]
- 57.Kang SA, Park HJ, Kim MJ, Lee SY, Han SW, Leem KH. Citri Reticulatae Viride Pericarpium extract induced apoptosis in SNU-C4, human colon cancer cells. J Ethnopharmacol. 2005;97:231–235. doi: 10.1016/j.jep.2004.11.012. [DOI] [PubMed] [Google Scholar]
- 58.Kwon KB, Kim EK, Lim JG, Jeong ES, Shin BC, Jeon YS, Kim KS, Seo EA, Ryu DG. Molecular mechanisms of apoptosis induced by Scorpio water extract in human hepatoma HepG2 cells. World J Gastroenterol. 2005;11:943–947. doi: 10.3748/wjg.v11.i7.943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhao H, Li Y, Wang Y, Zhang J, Ouyang X, Peng R, Yang J. Antitumor and immunostimulatory activity of a polysaccharide-protein complex from Scolopendra subspinipes mutilans L. Koch in tumor-bearing mice. Food Chem Toxicol. 2012;50:2648–2655. doi: 10.1016/j.fct.2012.05.018. [DOI] [PubMed] [Google Scholar]
- 60.Chung WT, Lee SH, Kim JD, Sung NS, Hwang B, Lee SY, Yu CY, Lee HY. Effect of the extracts from Glycyrrhiza uralensis Fisch on the growth characteristics of human cell lines: Anti-tumor and immune activation activities. Cytotechnology. 2001;37:55–64. doi: 10.1023/A:1016111713056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kitadai Y. Angiogenesis and lymphangiogenesis of gastric cancer. J Oncol. 2010;2010:468725. doi: 10.1155/2010/468725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Dotto GP. Crosstalk of Notch with p53 and p63 in cancer growth control. Nat Rev Cancer. 2009;9:587–595. doi: 10.1038/nrc2675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ling H, Jolicoeur P. Notch-1 signaling promotes the cyclinD1-dependent generation of mammary tumor-initiating cells that can revert to bi-potential progenitors from which they arise. Oncogene. 2013;32:3410–3419. doi: 10.1038/onc.2012.341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ye QF, Zhang YC, Peng XQ, Long Z, Ming YZ, He LY. Silencing Notch-1 induces apoptosis and increases the chemosensitivity of prostate cancer cells to docetaxel through Bcl-2 and Bax. Oncol Lett. 2012;3:879–884. doi: 10.3892/ol.2012.572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wang Z, Azmi AS, Ahmad A, Banerjee S, Wang S, Sarkar FH, Mohammad RM. TW-37, a small-molecule inhibitor of Bcl-2, inhibits cell growth and induces apoptosis in pancreatic cancer: involvement of Notch-1 signaling pathway. Cancer Res. 2009;69:2757–2765. doi: 10.1158/0008-5472.CAN-08-3060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Gao F, Yao M, Shi Y, Hao J, Ren Y, Liu Q, Wang X, Duan H. Notch pathway is involved in high glucose-induced apoptosis in podocytes via Bcl-2 and p53 pathways. J Cell Biochem. 2013;114:1029–1038. doi: 10.1002/jcb.24442. [DOI] [PubMed] [Google Scholar]