

Abstract
Aim
Here we investigated the role of complement activation in phagocytosis and the release of cytokines and chemokines in response to two clinical isolates: Borrelia afzelii K78, which is resistant to complement-mediated lysis, and Borrelia garinii LU59, which is complement-sensitive.
Methods
Borrelia spirochetes were incubated in hirudin plasma, or hirudin-anticoagulated whole blood. Complement activation was measured as the generation of C3a and sC5b-9. Binding of the complement components C3, factor H, C4, and C4BP to the bacterial surfaces was analyzed. The importance of complement activation on phagocytosis, and on the release of cytokines and chemokines, was investigated using inhibitors acting at different levels of the complement cascade.
Results
1) Borrelia garinii LU59 induced significantly higher complement activation than did Borrelia afzelii K78. 2) Borrelia afzelii K78 recruited higher amounts of factor H resulting in significantly lower C3 binding. 3) Both Borrelia strains were efficiently phagocytized by granulocytes and monocytes, with substantial inhibition by complement blockade at the levels of C3 and C5. 4) The release of the pro-inflammatory cytokines and chemokines IL-1β, IL-6, TNF, CCL20, and CXCL8, together with the anti-inflammatory IL-10, were increased the most (by>10-fold after exposure to Borrelia). 5) Both strains induced a similar release of cytokines and chemokines, which in contrast to the phagocytosis, was almost totally unaffected by complement blockade.
Conclusions
Our results show that complement activation plays an important role in the process of phagocytosis but not in the subsequent cytokine release in response to live Borrelia spirochetes.
Introduction
Lyme borreliosis is an infectious disease that is predominantly found in the northern hemisphere and is caused by the spirochetes of Borrelia burgdorferi sensu lato (s.l.) [1]. The infection can affect the skin, joints, heart, and/or nervous system [2], [3]. At least three genospecies in the B. burgdorferi s.l. group have been identified as human pathogens [4] and have been shown to be associated with different clinical manifestations: B. burgdorferi sensu stricto (s.s.) is mainly associated with arthritis, B. garinii with neuroborreliosis, and B. afzelii with skin manifestations [1], [5], [6].
To survive in different hosts and tissues, it is crucial for the Borrelia spirochetes to overcome the human host's immune response. The innate immune system is the first line of defence that the spirochetes encounter when entering the body. These reactions are the key determinants of the magnitude and quality of the early immune response, and they subsequently initiate and form the adaptive immune response [7]. The complement system is part of innate immunity and consists of a complex network of plasma and membrane-associated proteins that are activated in a cascade-like manner. It can be activated through three different pathways: the classical, lectin, and alternative, all of which converge in a central step, the activation of C3 to generate C3b, which serves as both a ligand for immune adhesion and a subunit of the alternative pathway convertase. Complement activation induced by microbes leads to important defense mechanisms such as phagocytosis of the target and the formation of the membrane attack complex (MAC).
Borrelia species differ in their ability to survive in the presence of complement, and they are classified as serum-resistant or serum-sensitive, based on a quantification of the amount of MAC formed on the bacterial membrane in vitro [8]. B. afzelii is classified as serum-resistant and B. burgdorferi s.s. as moderately resistant, whereas B. garinii is sensitive to complement-mediated killing [9]–[12]. Studies have shown that resistance to complement is correlated with an ability of the isolates to recruit fluid-phase immune regulators, factor H (FH) and factor H-like-protein-1 (FHL-1), to the bacterial surface. This process promotes the inactivation and degradation of the alternative pathway C3 convertase and C3b, which in turn hinder downstream activation, leading to decreased formation of MAC and a higher rate of spirochete survival [10], [13], [14]. In addition, it has recently been reported that B. burgdorferi s.l. is capable of binding C4-binding protein (C4BP), a regulator of the classical and lectin pathways [15].
The complement regulators of the FH family bind to complement regulator-acquiring surface proteins (CRASPs) that are expressed on the surface of B. burgdorferi s.l., thereby reducing the alternative pathway of complement activation [16], [17]. Expression of the five known CRASPs has been identified on the serum-resistant B. afzelii; in contrast, B. garinii, which is serum-sensitive, expresses only one CRASP with weak or no binding to FHL-1 and FH [17], [18], indicating that resistance to complement is correlated with the expression of CRASPs.
In addition to its role in phagocytosis and microbial killing, complement activation is a major factor in early inflammation and an important contributor to the release of cytokines and chemokines such as the pro-inflammatory IL1-β, IL-6, TNF, and CCL20 (MIP-3α). These mediators are important for the recruitment of other components of the innate immune response as well as signaling with the adaptive immune system [19]. It is therefore reasonable to hypothesize that the early innate immune response, including complement activation, phagocytic capacity, and cytokine/chemokine release, is of great importance for the response to B. burgdorferi s.l.
The overall aim of this study was to characterize complement activation and its role in early immune activation in response to Borrelia spirochetes. In vitro models based on clinical isolates of B. garinii and B. afzelii, sensitive and resistant to complement activation, respectively, were used to investigate the importance of complement activation in phagocytosis and in the generation of cytokines and chemokines.
Materials and Methods
Heparin treatment of equipment
All equipment that came in contact with blood and plasma (tubes and pipette tips) was furnished with a heparin surface according to the manufacturer's recommendations (Corline AB, Uppsala, Sweden).
Preparation of blood and plasma
Blood from healthy volunteers without (n = 8) or with (n = 1) specific anti-Borrelia antibodies was collected in 6-mL Vacutainer plastic tubes (BD Bioscience, Plymouth, UK) with the addition of the specific thrombin inhibitor hirudin (Refludan, Pharmion Ltd, Cambridge, UK), at a final concentration of 50 µg/mL blood. This study, using blood from healthy blood donors given their written consent, was performed with consent of the Ethical Committee of the University Hospital of Linköping, Sweden (#03-520). Plasma was collected by centrifugation at 3000 g for 20 min and stored at −80°C. For viability studies, aliquots of plasma were heat inactivated by incubation at 56°C for 30 min. For the phagocytosis experiments and cytokine release assays, blood was collected as described above and used within 30 min. Anti-Borrelia antibodies were measured in serum using the commercially available enzyme-linked immunosorbent assay (ELISA) kits Enzygnost Lyme link VlsE/IgG and Enzygnost Borreliosis IgM (DADE Behring, Marburg, Germany) on a BEP 2000 Advance System (Siemens Healthcare, Erlangen, Germany), according to the instructions from the manufacturer.
Bacterial strains and growth conditions
The Borrelia strains used in this study were isolated in Sven Bergström's laboratory, Umeå University, Sweden: B. garinii LU59 from human cerebrospinal fluid (CSF) and B. afzelii K78 from a human skin biopsy. Bacteria were grown in Barbour-Stoenner-Kelly (BSK) II medium [30] supplemented with 7% rabbit serum (Sigma Aldrich, St. Louis, MO, USA) at 37°C until the density reached 108–109 cells/mL.
Susceptibility of B. garinii LU59 and B. afzelii K78 to complement
In initial experiments, the susceptibility of both Borrelia strains to complement was investigated. Aliquots of B. garinii LU59 and B. afzelii K78 were incubated for 60 min at 37°C in serially diluted heat inactivated or native hirudin plasma, collected from a blood donor without Borrelia-specific antibodies. After incubation, the number of motile and non-motile spirochetes was counted in a phase contrast microscope [20], [21].
Complement activation induced by Borrelia
Bacteria were washed with Dulbecco's PBS supplemented with 0.9 mM Ca2+ (PBSCa2+) by centrifugation at 4500 g for 5 min, then resuspended and diluted in PBSCa2+. Aliquots of 50 µL B. garinii LU59 and B. afzelii K78 spirochetes (109/mL plasma) were incubated in 450 µL hirudin plasma (n = 8) without Borrelia-specific antibodies for 60 min at 37°C in 2-mL tubes. The reactions were stopped by the addition of ethylenediaminetetraacetic acid (EDTA, 10 mM final concentration; Sigma Aldrich) and centrifuged at 4500 g for 5 min. All incubations were performed in duplicate. Hirudin plasma incubated without spirochetes was used as a control. Complement activation was monitored as the generation of activation product C3a and sC5b-9 complexes, measured in the plasma by ELISA as described previously [22], [23]. Data are presented as the activation compared to a plasma control incubated in parallel, but without the presence of spirochetes.
Quantification of C3/C4 fragments, FH, and C4BP by ELISA
B. garinii LU59 and B. afzelii K78 (109/mL plasma) were incubated in hirudin plasma (n = 3) or PBSCa2+ at 37°C for 30 min and then centrifuged at 4500 g for 5 min. The pellets with spirochetes were washed three times with 500 µL PBSCa2+ and then resuspended in PBSCa2+ and transferred to new tubes. Biotinylated antibodies against human C3c (1.09 µg/mL), C4c (0.56 µg/mL), (Dako, Glostrup, Denmark), FH (1.13 µg/mL) and C4BP (1.44 ug/mL) (Binding Site, Birmingham, UK) were added, and the samples were incubated for 45 min at room temperature (RT). The samples were then centrifuged at 4500 g for 5 min and washed with wash buffer (PBS with 0.05% Tween 20; Sigma Aldrich) three times, then incubated with streptavidin-HRP (1∶500; GE Healthcare, Uppsala, Sweden), washed four times, and stained with o-phenylenediamine (OPD; Sigma Aldrich). The absorbance was measured at 492 nm.
Visualization of C3 fragments and FH by immunostaining and Western blot
Spirochetes (109/mL plasma) were incubated for 30 min in hirudin plasma or PBSCa2+ at 37°C. Complement activation was stopped by the addition of EDTA to a final concentration of 10 mM. Samples were centrifuged at 4500 g for 5 min, and the pellet containing the spirochetes was washed four times with PBSCa2+ and analyzed for C3 fragments and FH by immunostaining and Western blot. For immunostaining, the spirochetes were incubated for 30 min with polyclonal anti-bodies against human C3c, which also detects intact C3, C3b, and iC3b (200 µg/mL; Dako), FH (250 µg/mL; Binding Site), and Borrelia (280 µg/mL; Acris, Herford, Germany). All antibodies were conjugated with Alexa Fluor Protein Labeling Kits (Life Technologies, Grand Island, NY, USA). Alexa Fluor 488 was used for anti-C3c and anti-FH and Alexa Fluor 555 for anti-Borrelia antibodies. Samples were washed, and fluorescence was examined with a Nikon Eclipse E600 Confocal Microscope and Nikon Confocal Microscope EZ-C1 2.20 software (Nikon, Chiyoda, Tokyo, Japan).
Analysis of adsorbed proteins was also performed by SDS-polyacrylamide gel electrophoresis under reducing conditions. The proteins were either silver stained (Bio-Rad, Hercules, CA, USA) or transferred to a polyvinylidene fluoride (PVDF) Immune-Blot membrane (Bio-Rad). Detection was performed using biotinylated antibodies against human FH (0.57 µg/ml; Binding Site) and C3c (1.09 µg/ml; Dako), followed by streptavidin-HRP (1∶500; GE Healthcare). Proteins were visualized with ECL Plus Western blot detection system (GE Healthcare). Human C3, C3b, iC3b and FH, purified as described earlier, served as controls [24]. The relative amount of Borrelia and total protein was visualized by SDS-PAGE followed by silver staining. A 20 kDa-band of bacterial orgin, most likely OspE [25] was used as loading control in the Western blot experiments [26]. Six separate blots were performed using plasma from different donors. Quantification of the bound proteins was performed using public domain software (Java-based ImageJ software) from the NIH. The β-chain was chosen for quantification of the amount of bound C3-fragments because it is not subject to proteolytic cleavage resulting in transformation to C3b and iC3b.
Labeling of spirochetes with FITC
Spirochetes were labeled by a modification of the procedure described by Hazenbos et al. [27]. B. garinii LU59 and B. afzelii K78 were washed with 2 mL of 50 mM sodium carbonate with 100 mM sodium chloride, pH 7.4 (Sigma Aldrich), then centrifuged at 4500 g for 5 min. The pellets were suspended in 1 mL fluorescein isothiocyanate (0.5 mg/mL; FITC; Sigma Aldrich) in 50 mM sodium carbonate and 100 mM sodium chloride, pH 7.4, and incubated at RT for 20 min on a shaker, washed three times with 4 mL PBSCa2+, centrifuged at 4500 g for 5 min, and suspended in 2 mL PBSCa2+. The spirochetes were visualized by fluorescence microscopy and their motility was unchanged by the labeling procedure [20], [21].
Phagocytosis of spirochetes
Samples of 50 µL FITC-labeled B. garinii LU59 or B. afzelii K78 spirochetes (108/mL blood) were incubated in 450 µL hirudin blood without (n = 3) or with (n = 1) Borrelia-specific antibodies for 30 min at 37°C in 2-mL tubes. EDTA, which binds the Ca2+ and Mg2+ that are necessary for a functional complement system and therefore completely inhibits complement activation, was used at a final concentration of 10 mM as a negative control. Two additional inhibitors were used, 50 µM compstatin (Ac-ICV(1-MeW)QDWGAHRCT-NH2), a synthetic peptide inhibiting the activation of C3 by the two C3 convertases [28]; and 5.5 µM C5a receptor antagonist (PMX53; C5aRa), which blocks the C5a-mediated up-regulation of granulocytes and monocytes. A scrambled peptide of C5aRa (5.5 µM) was used as a control. The peptides were a kind gift from Dr. John D. Lambris (the University of Pennsylvania, Philadelphia, PA, USA). Reactions were stopped by the addition of 10 mM EDTA (final concentration). Samples (100 µL) of hirudin blood from the incubations were transferred to new tubes, and 2 mL AKC lysis buffer (155 mM NH4Cl, 10 mM KHCO3, 0.1 M EDTA; Sigma Aldrich) was added to remove and lyse the red blood cells. The reaction was allowed to continue for 5–10 min until the supernatants were clear red. After centrifugation at 400 g for 5 min, the supernatants were discarded. The pellets (leukocytes) were washed with 3 mL PBS supplemented with 1% (w/v) bovine serum albumin (BSA; Sigma Aldrich), and then 1.5 mL PBS with 1% paraformaldehyde (Apoteket, Gothenburg, Sweden) and 0.075% trypan blue were added to quench extracellular fluorescence [29], [30]. Granulocytes and monocytes were analyzed according to size (forward scatter) and granularity (side scatter) using flow cytometry (Cell lab Quanta SC, Beckman Coulter, Miami, FL, USA); 5,000 granulocytes were collected, resulting in the collection of ∼500 monocytes. Data are presented as mean fluorescence intensity (MFI). Phagocytosis without complement inhibitors was set to 100%.
Cytokine and chemokine release
Hirudin blood without anti-Borrelia antibodies (n = 3, same individuals as for the phagocytosis experiments described above) was used. Washed spirochetes (108/mL) were incubated in 450 µL of hirudin blood at 37°C for 4 h, with or without the addition of complement inhibitors at a final concentration of 50 µM for compstatin and 100 µg/ml for the monoclonal antibody eculizumab (Soliris, Alexion, Pharmaceuticals, Cheschire, CT, USA) which binds to C5 and prevents its activation. Hirudin blood incubated without spirochetes was used as a control. All incubations were performed in duplicate. Further complement activation was stopped by the addition of 10 mM EDTA (final concentration) and centrifuged at 3000 g for 20 min, and the resulting plasma samples were frozen immediately at −80°C. In addition, plasma samples for determination of base-line level of each cytokine or chemokine for each donor were collected directly, without incubation. Complement activation was monitored as the generation of the activation products C3a and sC5b-9 complexes (data not shown). Cytokine and chemokine concentrations were measured by Luminex xMAP technology (Milliplex Human Cytokine/Chemokine Kit, Merck Millipore, Billerica, MA, USA) according to the instructions provided by the manufacturer. The analyzed cytokines and chemokines were: IL-1β, IL-6, IL-10, IL-12(p70), IL-17A, IL-23, TNF, GM-CSF, CXCL10 (IP-10), CXCL1 (GRO-α), CCL22 (MDC), CXCL8 (IL-8), CCL20 (MIP3-α), and CCL2 (MCP-1). Values under the detection limit were given half the value of the lowest point of the standard curve. The inter-assay coefficients of variation (CV) were 3.5–16.4%, and the intra-assay CVs were 3.9–10.5%, according to the manufacturer.
Statistics
All data were normalized against plasma without bacteria that was treated in the same way as the samples; data are presented as means ± SEM. Statistical analysis of data was performed using Prism version 6.0 for Macintosh software (GraphPad, San Diego, CA, USA). Differences between means were evaluated using paired t-tests and one-way ANOVA followed by Dunnetts post hoc test, and p-values<0.05 were considered to be significant.
Results
Susceptibility of B. garinii LU59 and B. afzelii K78 to complement
B. garinii LU59 was highly sensitive to complement with 22 and 40% motile spirochetes remaining after incubation in hirudin plasma diluted 1∶3 respectively 1∶9. No motile spirochetes were found after incubation with undiluted plasma, while the presence of neither heat-inactivated nor native plasma in higher dilutions affected the viability. Complement activation by B. garinii LU59 was most likely induced by the alternative pathway since it requires high concentration (>10%) of all participating components [31]. Incubation with hirudin plasma under these conditions did not affect the motility of B. afzelii K78.
Complement activation induced by spirochetes
Complement activation by spirochetes in hirudin plasma was monitored by the generation of C3a and sC5b-9 complexes. When plasma without Borrelia-specific antibodies was used, higher levels of both activation products were found in plasma incubated with B. garinii LU59 when compared to plasma incubated with B. afzelii K78 (p<0.01) (Figure 1A). An identical experimental setup using plasma with anti-Borrelia IgG antibodies (n = 1) showed a higher activation of complement than when the antibodies were not present. In this setting, no significant difference was found between the two bacterial strains (data not shown).
Figure 1. Complement activation and adherence of complement proteins to Borrelia spirochetes incubated in hirudin plasma.

A) The ability of the Borrelia strains B. afzelii K78 and B.gariini LU59, resistant and sensitive to complement activation, respectively, to activate complement in hirudin plasma (n = 8) was assessed by analyzing the generation of C3a and sC5b-9 using ELISA. Data are normalized against plasma incubated without bacteria and are presented as means ± SEM. B) Borrelia were incubated with hirudin plasma (n = 6), and bound proteins were analyzed using an in-house-developed particle ELISA. Data are shown as absorbance values of bacteria incubated in plasma, normalized against bacteria incubated in PBSCa2+, and are presented as means ± SEM. C) Quantification of bound C3-fragments (using the β-chain that is not proteolytically cleaved) and FH. Results are presented normalized to either K78 or LU59 as mean of six Western blots. D) Silver stained SDS–PAGE of B. afzelii K78 and B.gariini LU59 non-opsonized (PBS) or opsonised (Pl). In addition, C3 and its fragments (E) and FH (F) bound to bacteria incubated with plasma (Pl) were visualized using Western blot with bacteria incubated in PBSCa2+ as controls, and purified C3, C3b and iC3b or FH as references. Insert: loading control, consisting of a 20kDa Borrelia protein that was detected on the membranes after detection with antibodies.
Quantification of C3/C4 fragments, FH, and C4BP bound to spirochetes
In order to attempt to explain the differences in complement activation between the two strains of bacteria, we analyzed the regulatory proteins FH and C4BP, as well as fragments of C3 and C4 bound to the spirochetes. An ELISA-like assay was developed in which spirochetes were incubated in plasma to allow the binding of complement proteins. B. garinii LU59 was found to bind significantly more C3-fragments compared to B. afzelii K78 (p p<0.05) while the opposite was true for binding of the regulatory protein FH (p<0.01) (Figure 1B). Similar differences, i.e., that the total amount of C3 bound to LU59 was more than double compared to K78 (p<0.0001), while the opposite binding was true for FH (p<0.01), were found by densitometric scanning of western blots (Figure 1C). In addition, both C4 (fragments) and the inhibitor C4BP were detected, albeit at low levels and with no significant difference between the two strains (data not shown).
In addition, immunostaining was used to ascertain that the detected C3 and FH indeed adhered to the spirochetes (data not shown). Silver staining of SDS-gels showed equal amount of bacteria in each sample and similar amounts of protein for both strains after incubation in plasma (Figure 1D). Western blots were performed to investigate the structure of the bound C3 (i.e. C3, C3b or iC3b), and whether FHL-1 and/or FH related proteins (FHR), bound in addition to FH (as well as for quantification). One representative example of six blots is shown (Figure 1E and F). These experiments were performed with an anti-C3c antibody that also detects degraded fragments of C3, and with an anti-FH antibody with an affinity for the FHL-1 and FHR proteins. Western blots showed that the C3 bound to both strains of spirochetes was quantitatively degraded to its inactive form iC3b, which is unable to form the alternative convertase but still functions as an opsonizing fragment. In the lanes containing opsonized Borrelia, the transition to iC3b is visualized by the total degradation of the α-chain and the appearance of the 40 kDa band (Figure 1E).
Phagocytosis of spirochetes
Phagocytosis of B. afzelii K78 by granulocytes was more efficient than that of B. garinii LU59 within a 5- to 30-min time period, (Figure 2A) with significant difference (p<0.05) at 30 min). To elucidate the importance of complement activation for efficient phagocytosis, we used inhibitors acting on different levels of the complement cascade (Figure 2B). The C5aRa showed an approximately 80% inhibition for both strains, with no synergy between the two inhibitors. The decrease in phagocytosis of each Borrelia strain by the two inhibitors, alone or in combination, were highly significant (p<0.0001). Similar results were obtained regarding phagocytosis by monocytes (data not shown). In the presence of Borrelia-specific antibodies, phagocytosis of B. garinii LU59 was inhibited by 70% in the presence of either inhibitor. B. afzelii K78 was 45% inhibited by compstatin and 60% by C5aRa (data not shown).
Figure 2. Importance of complement activation for efficient phagocytosis.
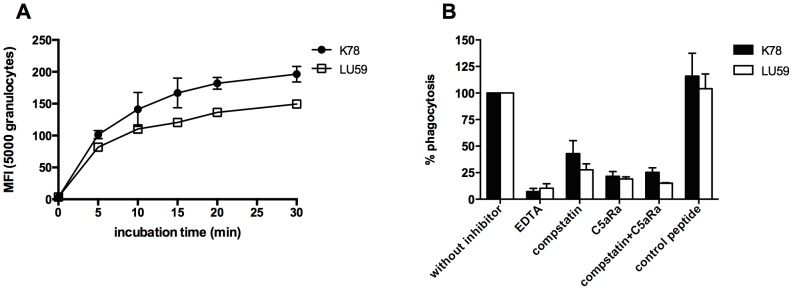
FITC-labeled B. afzelii K78 and B.garinii LU59 were incubated in hirudin blood to study phagocytosis by measuring the mean fluorescence intensity (MFI) for granulocytes. A) Phagocytosis of each bacterial strain was monitored for 30 min. Results are presented as mean ± SEM of two observations. B) The importance of complement activation in phagocytosis was analyzed by incubating Borrelia spirochetes in hirudin blood (n = 3) with the addition of complement inhibitors acting at different stages in the activation cascade. Compstatin selectively inhibits the proteolytic activation of C3 and a C5a receptor antagonist (C5aRa) that blocks the C5a-mediated up-regulation of granulocytes and monocytes. A scrambled peptide of C5aRa was used as a control. The mean fluorescence intensity (MFI) for each bacterial strain incubated in hirudin blood without complement inhibitors was set to 100%. EDTA: ethylene diaminetetraacetic acid. C5aRa: C5a receptor antagonist.
Cytokine and chemokine release
As a measurement of the early inflammatory response to Borrelia infection, we analyzed the levels of secreted cytokines and chemokines after 4 h of incubation of hirudin blood with or without Borrelia spirochetes. After exposure to Borrelia spirochetes, we saw increases in the secreted levels of IL-1β (10,000-fold), IL-6 (400-fold), TNF (140-fold), CCL20 (40-fold), IL-10 (35-fold), CXCL8 (15-fold), GM-CSF (10-fold), CXCL10 (8-fold), IL-23 (7-fold), IL-12p70 (4-fold), CXCL1 (4-fold), and IL-17A (1.5-fold), although the levels did not differ significantly between B. afzelii K78 and B. garinii LU59 (Figure 3A). CCL22 and CXCL1 did not reach levels above the detection limit of the assay. The pro-inflammatory cytokines IL-1β, IL-6, TNF, CCL20, and CXCL8, together with the anti-inflammatory IL-10, increased the most, i.e., more than 10-fold in comparison with the control. Therefore, these cytokines and chemokines were used to further elucidate the importance of complement activation for their release (Figure 3B and C). Inhibition of complement at the levels of C3a and C5a generation showed no significant effect on cytokine or chemokine release, the only exception being CXCL8, which showed a significant 60% decrease for LU59 in the presence of compstatin (Figure 3C). The background levels of all cytokines (i.e. in plasma which was collected immediately, without incubation) were all below the detection limit in each blood donor.
Figure 3. Cytokine and chemokine levels in plasma as a response to incubation with Borrelia spirochetes.

Spirochetes were incubated in whole blood (n = 3) in duplicate for 4 h, and the cytokine/chemokine production was then measured using a Luminex-based assay. Results are normalized against the values for plasma without the addition of bacteria. (A) The x-fold increase in the panel of cytokines and chemokines analyzed in this report showed no difference between the complement-resistant B. afzelii K78 and -sensitive B. garinii LU59. (B and C) The importance of complement activation was further investigated using two complement inhibitors: compstatin, which selectively inhibits the proteolytic activation of C3, and eculizumab, an anti-C5 antibody blocking the cleavage of C5 and therefore further complement activation and C5a mediated signaling. Data are presented as means ± SEM.
Discussion
In the present study, we have investigated functional aspects of the early innate immune response to B. afzelii K78 and B. garinii LU59 by examining complement activation, phagocytosis, and cytokine and chemokine release.
One main finding regarding complement activation was the significant difference in found between the two strains of Borrelia. In accordance with previous findings [32], we found that the complement-sensitive strain B. garinii LU59 induced a higher level of complement activation, as measured by the generation of C3a and sC5b-9 in plasma and detected by ELISA. Complement activation induced by the presence of Borrelia-specific antibodies resulted in a significant increase in generated sC5b-9, in agreement with other studies of the deposition of C5b-9 on the bacterial surface [8], [18], [33], [34]. Moreover, although they were measured in only one case (because we identified only one donor with anti-Borrelia-specific antibodies, despite having screened a large number of blood donors) the presence of Borrelia-specific antibodies compensated for the differences in complement activation between the sensitive and the resistant strain, thus indicating the benefit of pre-existing antibodies for efficient clearance.
It has previously been shown that all three pathways of complement activation are involved in complement-mediated killing of Borrelia spirochetes, although the alternative pathway is believed to be the most important [12]. Recruitment of complement regulators to the bacterial surface is an established mechanism used by pathogens to avoid complement attack [35]–[37]. Here we investigated the two main regulators of the alternative and classical pathway, FH and C4BP and both of which act as co-factors for factor I (FI)-mediated degradation of C3b and C4b, respectively [38]–[40]. Regulatory proteins bound to the membrane of B. garinii LU59 and B. afzelii K78 were quantified by an in-house-developed particle ELISA, as well as by western blotting (FH) confirming that B. afzelii K78 had recruited more FH (at least twice as much) than had the complement-sensitive B. garinii LU59, an observation also made by others [10], [13], [41]. Furthermore, we detected significantly higher amounts of deposited C3 on LU59 by both techniques, more than twice the amount found on K78, which is in agreement with the higher amount of C3a (and subsequently sC5b-9), generated by LU59. Interestingly, western blot analysis showed that the deposited C3 had been quantitatively degraded to iC3b on both strains despite the fourfold difference in C3/FH ratio, suggesting that FH may not be the limiting factor under the experimental conditions used here.
In addition, low amounts of C4BP were detected on both strains, with a non-significantly higher level on B. garinii LU59. Binding of C4BP to B. garinii in the literature is contradictory, studies in which no binding of C4BP was detected when a similar experimental setup has been reported [13], [32] as well as studies in line with data presented here where binding of C4BP is observed [15].
Microbes are generally cleared from the body by phagocytosis. The Borrelia spirochetes used in this study were phagocytosed by both granulocytes and monocytes, a finding that has previously been demonstrated using other B. burgdorferi s.l [42]–[44]. For these studies we used FITC-labeling, which is a standard technique to study opsonization and phagocytosis of pathogens [45], [46]. Intriguingly, the phagocytosis assay showed that B. afzelii K78 was more extensively phagocytosed than was B. garinii LU59 at all time points, a result that has not been reported previously. It is most unlikely that this difference in phagocytosis is a consequence of differences in viability since live and dead bacteria are phagocytosed by granulocytes at similar rate [47].
When we examined the importance of complement activation in phagocytosis by using several complement inhibitors interfering with the activation of complement at different stages in the activation cascade, we found that phagocytosis was obliterated in the presence of the complement inhibitor EDTA, a chelator that binds Ca2+ and Mg2+. These ions are necessary for a functional complement system but also for a number of other biochemical reactions so the addition of EDTA could possibly have affected other mechanisms involved in phagocytosis, and not uniquely the complement system. Therefore, we also tested the effects of complement inhibitors acting at the levels of C3 and C5, respectively. Both inhibitors produced a 50–80% reduction in phagocytosis, without any obvious synergistic effects. Although it can not be totally excluded that the FITC-labeling procedure per se may affect activation and binding of complement proteins to the pathogen surface, the marked effect on phagocytosis seen using compstatin and C5aRa makes this option less likely. The effect of complement inhibition on phagocytosis was less prominent in the presence of Borrelia-specific antibodies. Consequently, complement activation is most important but not completely essential for phagocytosis. This observation is supported by other in vitro studies of human neutrophils as well as by in vivo studies using C3 knockout mice [42], [48].
Regarding the analysis of early cytokine and chemokine release in human blood in response to live spirochetes, we found a rapid and pronounced pro-inflammatory response, with high levels of major inflammatory mediators such as IL-1β, IL-6, TNF, and CXCL8, an observation that is in line with previous reports [19], [49], [50]. Interestingly, the T helper (Th) 17-recruiting chemokine CCL20 was also highly induced, as well as the Th17-inducing cytokine IL-23. In addition, pro-inflammatory IL-6 aids in Th17 induction, and CXCL8 is in part associated with Th17. These findings are in line with recent observations of Th17-like immunity in neuroborreliosis [51]–[53] and Lyme arthritis [54]. Furthermore, Th1-inducing IL-12p70, as well as Th1-recruiting CXCL10, were among the highly induced cytokines, whereas the Th2-recruiting CCL22 was not induced at all. Thus, the pattern we observed could reflect the ensuing adaptive response that is characterized by Th1 predominance over Th2 reactivity [51], [53], [55]–[57]. Notably, we found that already after 4 h of incubation with Borrelia spirochetes, there were significantly elevated levels of IL-10. IL-10 is considered a key negative regulator of pro-inflammatory cytokine release and function [58]. It has been shown that B. burgdorferi s.l. induces the secretion of IL-10 in mononuclear cells, and the clearance of B. burgdorferi s.l. in IL-10-deficient mice is 10-fold higher than that in wild-type mice [59]. These studies, together with our findings suggest, that B. burgdorferi s.l. itself may induce IL-10 to inhibit the host defense.
Activation of pattern recognition receptors (PRR), such as toll-like receptors (TLR), by conserved lipoproteins on the spirochetal membrane enhances phagocytosis and antigen presentation, as well as triggering the production and release of inflammatory cytokines [60], [61]. TLR activation induced by Borrelia spirochetes has been shown to induce the activation of the transcription factor NFκβ, which is involved in the production of pro-inflammatory cytokines [62], [63]. In the present study, we showed a 10- to 10,000-fold increase in the release of pro-inflammatory cytokines when compared to background levels. IL-1β increased the most (i.e., 10 times more than IL-6). IL-1β has previously been connected with the activation of PRR as a response to Borrelia spirochetes [64]. It has recently been suggested that autophagy is responsible for an increased production of IL-1β and IL-6 in peripheral mononuclear blood cells as a response to B. burgdorferi s.l. [65]. Furthermore, differences in cytokine response has been described following phagocytosis depending on the viability of E.coli: both dead and live bacteria are reported to induce, e.g. IL-6 to similar degree, while generation of IL-1β is much reduced after phagocytosis of dead bacteria. This difference has been attributed to the intracellular release of bacterial mRNA from live, ingested bacteria, which leads to inflammasome activation which is required for processing of pro-IL-1β to mature IL-1β [66].
Our results showed that inhibition of complement activation decreased phagocytosis without affecting the release of any of the detected cytokines and chemokines, except for CXCL8, indicating that phagocytosis is not crucial for the pro-inflammatory response. This finding is in contrast to other observations that have shown a significant decrease in pro-inflammatory cytokines when phagocytosis or phagosomal signalling is blocked [43], [67]. However, these studies were made using systems of purified monocytes, without the presence of active serum, so the impact of complement activation could not be evaluated.
Based on our findings, we propose that the complement-sensitive strain B. garinii LU59, which is less prone to inhibit the alternative pathway by recruiting FH, would seek an immunologically privileged site such as the central nervous system (CNS). This behavior would constitute an optimal strategy for escaping the effects of complement in early infection and could at least in part explain the neurotropism observed in this genospecies [6]. However, it cannot be the full explanation, since B. garinii OspA serotype 4, which has been repeatedly isolated from human CSF [68], is able to bind FHL-1 quite efficiently [69]. During established CNS borreliosis, we have previously demonstrated evidence of complement activation via the classical pathway in CSF [70]. However, B. garinii has previously been shown to be able to bind C4BP [15], a finding that is consistent with the in vitro experiments performed here. Therefore, C4BP could constitute an important virulence factor for spirochetal survival in the presence of antibodies.
Taken together, our findings suggest that the early innate immune response, including complement activation, phagocytosis, and the release of cytokines and chemokines, may influence the course of Borrelia infection. We have shown: 1) that the complement-resistant strain B. afzelii K78 recruits more FH and hence induces a lower activation of complement than does the complement-sensitive B. garinii LU59, a finding that in part could explain the neurotropism of the sensitive strain; 2) that both granulocytes and monocytes participate in the phagocytosis of Borrelia spirochetes and that complement activation is extremely important, but not crucial, for spirochetal phagocytosis; 3) that the early cytokine/chemokine response to Borrelia spirochetes was dominated by pro-inflammatory mediators and also by inducers and recruiters of Th1- and Th17-like immunity; 4) that the assessed cytokine/chemokine response was not affected by the complement system in this in vitro model. Since only minor differences were found in response to the two strains, a valid conclusion from this work could be that there is enough redundancy in complement sufficient subjects to eradicate Borrelia infections.
Acknowledgments
The authors would like to thank MSc Petra Cassel, Division of Clinical Immunology, Department of Clinical and Experimental Medicine, Linköping University, for help with the Luminex assay; Ms Bodil Bagert for help with the initial motility determinations; Professor John Lambris, University of Pennsylvania, Philadelphia, PA, USA, for providing compstatin and the C5aRa; Professor Bo-Eric Malmvall, Futurum Academy for Health and Care, Ryhov County Hospital, Jönköping, for giving advice on the manuscript; and Associate Professor Maria Jenmalm, Division of Clinical Immunology, Department of Clinical and Experimental Medicine, Linköping University, for valuable discussions. We also thank Dr. Deborah McClellan for excellent editorial assistance. This work was supported by grants from Futurum Academy for Health and Care, Jönköping County Council, Sweden, the Family Olinder-Nielsen's Foundation, FORSS – Medical Research Council of Southeast Sweden, and the Linnaeus University.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are included within the paper.
Funding Statement
AJH was supported by Futurum Academy for Health and Care, Jönköping County Council, Sweden, and Family Olinder-Nielsen's Foundation. AJH, JE, and KNE were supported by FORSS – Medical Research Council of Southeast Sweden. KNE was also supported by Linnaeus University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Stanek G, Wormser GP, Gray J, Strle F (2012) Lyme borreliosis. Lancet 379: 461–473. [DOI] [PubMed] [Google Scholar]
- 2. Berglund J, Eitrem R, Ornstein K, Lindberg A, Ringer A, et al. (1995) An epidemiologic study of Lyme disease in southern Sweden. N Engl J Med 333: 1319–1327. [DOI] [PubMed] [Google Scholar]
- 3. Cimmino MA (1998) Relative frequency of Lyme borreliosis and of its clinical manifestations in Europe. European Community Concerted Action on Risk Assessment in Lyme Borreliosis. Infection 26: 298–300. [DOI] [PubMed] [Google Scholar]
- 4. Baranton G, Postic D, Saint Girons I, Boerlin P, Piffaretti JC, et al. (1992) Delineation of Borrelia burgdorferi sensu stricto, Borrelia garinii sp. nov., and group VS461 associated with Lyme borreliosis. Int J Syst Bacteriol 42: 378–383. [DOI] [PubMed] [Google Scholar]
- 5. Bacon RM, Kugeler KJ, Mead PS (2008) Surveillance for Lyme disease–United States, 1992-2006. MMWR Surveill Summ 57: 1–9. [PubMed] [Google Scholar]
- 6. van Dam AP, Kuiper H, Vos K, Widjojokusumo A, de Jongh BM, et al. (1993) Different genospecies of Borrelia burgdorferi are associated with distinct clinical manifestations of Lyme borreliosis. Clin Infect Dis 17: 708–717. [DOI] [PubMed] [Google Scholar]
- 7. Salazar JC, Pope CD, Sellati TJ, Feder HM Jr, Kiely TG, et al. (2003) Coevolution of markers of innate and adaptive immunity in skin and peripheral blood of patients with erythema migrans. J Immunol 171: 2660–2670. [DOI] [PubMed] [Google Scholar]
- 8. Breitner-Ruddock S, Wurzner R, Schulze J, Brade V (1997) Heterogeneity in the complement-dependent bacteriolysis within the species of Borrelia burgdorferi. Med Microbiol Immunol 185: 253–260. [DOI] [PubMed] [Google Scholar]
- 9. Kraiczy P, Hunfeld KP, Breitner-Ruddock S, Wurzner R, Acker G, et al. (2000) Comparison of two laboratory methods for the determination of serum resistance in Borrelia burgdorferi isolates. Immunobiology 201: 406–419. [DOI] [PubMed] [Google Scholar]
- 10. Kraiczy P, Skerka C, Kirschfink M, Zipfel PF, Brade V (2001) Mechanism of complement resistance of pathogenic Borrelia burgdorferi isolates. Int Immunopharmacol 1: 393–401. [DOI] [PubMed] [Google Scholar]
- 11. Kurtenbach K, De Michelis S, Etti S, Schafer SM, Sewell HS, et al. (2002) Host association of Borrelia burgdorferi sensu lato–the key role of host complement. Trends Microbiol 10: 74–79. [DOI] [PubMed] [Google Scholar]
- 12. van Dam AP, Oei A, Jaspars R, Fijen C, Wilske B, et al. (1997) Complement-mediated serum sensitivity among spirochetes that cause Lyme disease. Infect Immun 65: 1228–1236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Alitalo A, Meri T, Ramo L, Jokiranta TS, Heikkila T, et al. (2001) Complement evasion by Borrelia burgdorferi: serum-resistant strains promote C3b inactivation. Infect Immun 69: 3685–3691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Hellwage J, Meri T, Heikkila T, Alitalo A, Panelius J, et al. (2001) The complement regulator factor H binds to the surface protein OspE of Borrelia burgdorferi. J Biol Chem 276: 8427–8435. [DOI] [PubMed] [Google Scholar]
- 15. Pietikainen J, Meri T, Blom AM, Meri S (2010) Binding of the complement inhibitor C4b-binding protein to Lyme disease Borreliae. Mol Immunol 47: 1299–1305. [DOI] [PubMed] [Google Scholar]
- 16. Bykowski T, Woodman ME, Cooley AE, Brissette CA, Wallich R, et al. (2008) Borrelia burgdorferi complement regulator-acquiring surface proteins (BbCRASPs): Expression patterns during the mammal-tick infection cycle. Int J Med Microbiol 298 Suppl 1249–256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Jozsi M, Zipfel PF (2008) Factor H family proteins and human diseases. Trends Immunol 29: 380–387. [DOI] [PubMed] [Google Scholar]
- 18. Kraiczy P, Skerka C, Kirschfink M, Brade V, Zipfel PF (2001) Immune evasion of Borrelia burgdorferi by acquisition of human complement regulators FHL-1/reconectin and Factor H. Eur J Immunol. 31: 1674–1684. [DOI] [PubMed] [Google Scholar]
- 19. McKisic MD, Barthold SW (2000) T-cell-independent responses to Borrelia burgdorferi are critical for protective immunity and resolution of lyme disease. Infect Immun 68: 5190–5197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Meri T, Cutler SJ, Blom AM, Meri S, Jokiranta TS (2006) Relapsing fever spirochetes Borrelia recurrentis and B. duttonii acquire complement regulators C4b-binding protein and factor H. Infect Immun. 74: 4157–4163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Shamaei-Tousi A, Burns MJ, Benach JL, Furie MB, Gergel EI, et al. (2000) The relapsing fever spirochaete, Borrelia crocidurae, activates human endothelial cells and promotes the transendothelial migration of neutrophils. Cell Microbiol 2: 591–599. [DOI] [PubMed] [Google Scholar]
- 22. Nilsson Ekdahl K, Nilsson B, Pekna M, Nilsson UR (1992) Generation of iC3 at the interface between blood and gas. Scand J Immunol 35: 85–91. [DOI] [PubMed] [Google Scholar]
- 23. Mollnes TE, Lea T, Froland SS, Harboe M (1985) Quantification of the terminal complement complex in human plasma by an enzyme-linked immunosorbent assay based on monoclonal antibodies against a neoantigen of the complex. Scand J Immunol 22: 197–202. [DOI] [PubMed] [Google Scholar]
- 24. Hammer CH, Wirtz GH, Renfer L, Gresham HD, Tack BF (1981) Large scale isolation of functionally active components of the human complement system. J Biol Chem 256: 3995–4006. [PubMed] [Google Scholar]
- 25. Welinder C, Ekblad L (2011) Coomassie staining as loading control in Western blot analysis. J Proteome Res 10: 1416–1419. [DOI] [PubMed] [Google Scholar]
- 26. Alitalo A, Meri T, Chen T, Lankinen H, Cheng ZZ, et al. (2004) Lysine-dependent multipoint binding of the Borrelia burgdorferi virulence factor outer surface protein E to the C terminus of factor H. J Immunol. 172: 6195–6201. [DOI] [PubMed] [Google Scholar]
- 27. Hazenbos WL, van den Berg BM, van't Wout JW, Mooi FR, van Furth R (1994) Virulence factors determine attachment and ingestion of nonopsonized and opsonized Bordetella pertussis by human monocytes. Infect Immun 62: 4818–4824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Katragadda M, Magotti P, Sfyroera G, Lambris JD (2006) Hydrophobic effect and hydrogen bonds account for the improved activity of a complement inhibitor, compstatin. J Med Chem 49: 4616–4622. [DOI] [PubMed] [Google Scholar]
- 29. Nuutila J, Lilius EM (2005) Flow cytometric quantitative determination of ingestion by phagocytes needs the distinguishing of overlapping populations of binding and ingesting cells. Cytometry A 65: 93–102. [DOI] [PubMed] [Google Scholar]
- 30. Klippel N, Cui S, Groebe L, Bilitewski U (2010) Deletion of the Candida albicans histidine kinase gene CHK1 improves recognition by phagocytes through an increased exposure of cell wall beta-1,3-glucans. Microbiology 156: 3432–3444. [DOI] [PubMed] [Google Scholar]
- 31. Nilsson UR, Storm KE, Elwing H, Nilsson B (1993) Conformational epitopes of C3 reflecting its mode of binding to an artificial polymer surface. Mol Immunol 30: 211–219. [DOI] [PubMed] [Google Scholar]
- 32. Schwab J, Hammerschmidt C, Richter D, Skerka C, Matuschka FR, et al. (2013) Borrelia valaisiana resist complement-mediated killing independently of the recruitment of immune regulators and inactivation of complement components. PLoS One 8: e53659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Brade V, Kleber I, Acker G (1992) Differences of two Borrelia burgdorferi strains in complement activation and serum resistance. Immunobiology 185: 453–465. [DOI] [PubMed] [Google Scholar]
- 34. Kochi SK, Johnson RC, Dalmasso AP (1991) Complement-mediated killing of the Lyme disease spirochete Borrelia burgdorferi. Role of antibody in formation of an effective membrane attack complex. J Immunol 146: 3964–3970. [PubMed] [Google Scholar]
- 35. Horstmann RD, Sievertsen HJ, Knobloch J, Fischetti VA (1988) Antiphagocytic activity of streptococcal M protein: selective binding of complement control protein factor H. Proc Natl Acad Sci U S A. 85: 1657–1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Janulczyk R, Iannelli F, Sjoholm AG, Pozzi G, Bjorck L (2000) Hic, a novel surface protein of Streptococcus pneumoniae that interferes with complement function. J Biol Chem 275: 37257–37263. [DOI] [PubMed] [Google Scholar]
- 37. Ram S, McQuillen DP, Gulati S, Elkins C, Pangburn MK, et al. (1998) Binding of complement factor H to loop 5 of porin protein 1A: a molecular mechanism of serum resistance of nonsialylated Neisseria gonorrhoeae. J Exp Med 188: 671–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Kuhn S, Skerka C, Zipfel PF (1995) Mapping of the complement regulatory domains in the human factor H-like protein 1 and in factor H1. J Immunol 155: 5663–5670. [PubMed] [Google Scholar]
- 39. Whaley K, Ruddy S (1976) Modulation of C3b hemolytic activity by a plasma protein distinct from C3b inactivator. Science 193: 1011–1013. [DOI] [PubMed] [Google Scholar]
- 40. Zipfel PF, Skerka C (2009) Complement regulators and inhibitory proteins. Nat Rev Immunol 9: 729–740. [DOI] [PubMed] [Google Scholar]
- 41. Kraiczy P, Hellwage J, Skerka C, Becker H, Kirschfink M, et al. (2004) Complement resistance of Borrelia burgdorferi correlates with the expression of BbCRASP-1, a novel linear plasmid-encoded surface protein that interacts with human factor H and FHL-1 and is unrelated to Erp proteins. J Biol Chem 279: 2421–2429. [DOI] [PubMed] [Google Scholar]
- 42. Suhonen J, Hartiala K, Tuominen-Gustafsson H, Viljanen MK (2000) Borrelia burgdorferi—induced oxidative burst, calcium mobilization, and phagocytosis of human neutrophils are complement dependent. J Infect Dis 181: 195–202. [DOI] [PubMed] [Google Scholar]
- 43. Moore MW, Cruz AR, LaVake CJ, Marzo AL, Eggers CH, et al. (2007) Phagocytosis of Borrelia burgdorferi and Treponema pallidum potentiates innate immune activation and induces gamma interferon production. Infect Immun 75: 2046–2062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Cruz AR, Moore MW, La Vake CJ, Eggers CH, Salazar JC, et al. (2008) Phagocytosis of Borrelia burgdorferi, the Lyme disease spirochete, potentiates innate immune activation and induces apoptosis in human monocytes. Infect Immun 76: 56–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Fijen CA, Bredius RG, Kuijper EJ, Out TA, De Haas M, et al. (2000) The role of Fcgamma receptor polymorphisms and C3 in the immune defence against Neisseria meningitidis in complement-deficient individuals. Clin Exp Immunol 120: 338–345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Ko YP, Kuipers A, Freitag CM, Jongerius I, Medina E, et al. (2013) Phagocytosis escape by a Staphylococcus aureus protein that connects complement and coagulation proteins at the bacterial surface. PLoS Pathog 9: e1003816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Peterson PK, Verhoef J, Sabath LD, Quie PG (1976) Extracellular and bacterial factors influencing staphylococcal phagocytosis and killing by human polymorphonuclear leukocytes. Infect Immun 14: 496–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. van Burgel ND, Balmus NC, Fikrig E, van Dam AP (2011) Infectivity of Borrelia burgdorferi sensu lato is unaltered in C3-deficient mice. Ticks Tick Borne Dis 2: 20–26. [DOI] [PubMed] [Google Scholar]
- 49. Suhonen J, Komi J, Soukka J, Lassila O, Viljanen MK (2003) Interaction between Borrelia burgdorferi and immature human dendritic cells. Scand J Immunol 58: 67–75. [DOI] [PubMed] [Google Scholar]
- 50. Dennis VA, Dixit S, O'Brien SM, Alvarez X, Pahar B, et al. (2009) Live Borrelia burgdorferi spirochetes elicit inflammatory mediators from human monocytes via the Toll-like receptor signaling pathway. Infect Immun 77: 1238–1245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Henningsson AJ, Tjernberg I, Malmvall BE, Forsberg P, Ernerudh J (2011) Indications of Th1 and Th17 responses in cerebrospinal fluid from patients with Lyme neuroborreliosis: a large retrospective study. J Neuroinflammation 8: 36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Nordberg M, Forsberg P, Johansson A, Nyman D, Jansson C, et al. (2011) Cytotoxic mechanisms may play a role in the local immune response in the central nervous system in neuroborreliosis. J Neuroimmunol 232: 186–193. [DOI] [PubMed] [Google Scholar]
- 53. Liba Z, Kayserova J, Komarek V (2013) Th1 and Th17 but no Th2-related cytokine spectrum in the cerebrospinal fluid of children with Borrelia-related facial nerve palsy. Fluids Barriers CNS 10: 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Codolo G, Amedei A, Steere AC, Papinutto E, Cappon A, et al. (2008) Borrelia burgdorferi NapA-driven Th17 cell inflammation in lyme arthritis. Arthritis Rheum 58: 3609–3617. [DOI] [PubMed] [Google Scholar]
- 55. Ekerfelt C, Ernerudh J, Bunikis J, Vrethem M, Aagesen J, et al. (1997) Compartmentalization of antigen specific cytokine responses to the central nervous system in CNS borreliosis: secretion of IFN-gamma predominates over IL-4 secretion in response to outer surface proteins of Lyme disease Borrelia spirochetes. J Neuroimmunol 79: 155–162. [DOI] [PubMed] [Google Scholar]
- 56. Widhe M, Jarefors S, Ekerfelt C, Vrethem M, Bergstrom S, et al. (2004) Borrelia-specific interferon-gamma and interleukin-4 secretion in cerebrospinal fluid and blood during Lyme borreliosis in humans: association with clinical outcome. J Infect Dis 189: 1881–1891. [DOI] [PubMed] [Google Scholar]
- 57. Sjowall J, Carlsson A, Vaarala O, Bergstrom S, Ernerudh J, et al. (2005) Innate immune responses in Lyme borreliosis: enhanced tumour necrosis factor-alpha and interleukin-12 in asymptomatic individuals in response to live spirochetes. Clin Exp Immunol 141: 89–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Giambartolomei GH, Dennis VA, Philipp MT (1998) Borrelia burgdorferi stimulates the production of interleukin-10 in peripheral blood mononuclear cells from uninfected humans and rhesus monkeys. Infect Immun 66: 2691–2697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Lazarus JJ, Meadows MJ, Lintner RE, Wooten RM (2006) IL-10 deficiency promotes increased Borrelia burgdorferi clearance predominantly through enhanced innate immune responses. J Immunol 177: 7076–7085. [DOI] [PubMed] [Google Scholar]
- 60. Steere AC, Angelis SM (2006) Therapy for Lyme arthritis: strategies for the treatment of antibiotic-refractory arthritis. Arthritis Rheum 54: 3079–3086. [DOI] [PubMed] [Google Scholar]
- 61. Blander JM, Medzhitov R (2006) Toll-dependent selection of microbial antigens for presentation by dendritic cells. Nature 440: 808–812. [DOI] [PubMed] [Google Scholar]
- 62. Hirschfeld M, Kirschning CJ, Schwandner R, Wesche H, Weis JH, et al. (1999) Cutting edge: inflammatory signaling by Borrelia burgdorferi lipoproteins is mediated by toll-like receptor 2. J Immunol 163: 2382–2386. [PubMed] [Google Scholar]
- 63. Alexopoulou L, Thomas V, Schnare M, Lobet Y, Anguita J, et al. (2002) Hyporesponsiveness to vaccination with Borrelia burgdorferi OspA in humans and in TLR1- and TLR2-deficient mice. Nat Med 8: 878–884. [DOI] [PubMed] [Google Scholar]
- 64. Petzke MM, Brooks A, Krupna MA, Mordue D, Schwartz I (2009) Recognition of Borrelia burgdorferi, the Lyme disease spirochete, by TLR7 and TLR9 induces a type I IFN response by human immune cells. J Immunol 183: 5279–5292. [DOI] [PubMed] [Google Scholar]
- 65. Buffen K, Oosting M, Mennens S, Anand PK, Plantinga TS, et al. (2013) Autophagy modulates Borrelia burgdorferi-induced production of interleukin-1beta (IL-1beta). J Biol Chem 288: 8658–8666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Underhill DM, Goodridge HS (2012) Information processing during phagocytosis. Nat Rev Immunol 12: 492–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Cervantes JL, Dunham-Ems SM, La Vake CJ, Petzke MM, Sahay B, et al. (2011) Phagosomal signaling by Borrelia burgdorferi in human monocytes involves Toll-like receptor (TLR) 2 and TLR8 cooperativity and TLR8-mediated induction of IFN-beta. Proc Natl Acad Sci U S A 108: 3683–3688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Wilske B, Busch U, Eiffert H, Fingerle V, Pfister HW, et al. (1996) Diversity of OspA and OspC among cerebrospinal fluid isolates of Borrelia burgdorferi sensu lato from patients with neuroborreliosis in Germany. Med Microbiol Immunol 184: 195–201. [DOI] [PubMed] [Google Scholar]
- 69. van Burgel ND, Kraiczy P, Schuijt TJ, Zipfel PF, van Dam AP (2010) Identification and functional characterisation of Complement Regulator Acquiring Surface Protein-1 of serum resistant Borrelia garinii OspA serotype 4. BMC Microbiol 10: 43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Henningsson AJ, Ernerudh J, Sandholm K, Carlsson SA, Granlund H, et al. (2007) Complement activation in Lyme neuroborreliosis—increased levels of C1q and C3a in cerebrospinal fluid indicate complement activation in the CNS. J Neuroimmunol 183: 200–207. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are included within the paper.