

Abstract
The generation of chimeric embryos is a widespread and powerful approach to study cell fates, tissue interactions, and species-specific contributions to the histological and morphological development of vertebrate embryos. In particular, the use of chimeric embryos has established the importance of neural crest in directing the species-specific morphology of the craniofacial complex. The method described herein utilizes two avian species, duck and quail, with remarkably different craniofacial morphology. This method greatly facilitates the investigation of molecular and cellular regulation of species-specific pattern in the craniofacial complex. Experiments in quail and duck chimeric embryos have already revealed neural crest-mediated tissue interactions and cell-autonomous behaviors that regulate species-specific pattern in the craniofacial skeleton, musculature, and integument. The great diversity of neural crest derivatives suggests significant potential for future applications of the quail-duck chimeric system to understanding vertebrate development, disease, and evolution.
Keywords: Developmental Biology, Issue 87, neural crest, quail-duck chimeras, craniofacial development, epithelial-mesenchymal interactions, tissue transplants, evolutionary developmental biology
Introduction
The facial skeleton develops from the growth and fusion of multiple facial processes that are composed of neural crest and mesodermal mesenchyme surrounded by ectodermal and endodermal epithelial layers1-11. Morphogenetic events within each process are governed by distinct signaling interactions between the mesenchyme and surrounding epithelia12-16. Alterations to these signaling interactions and/or their downstream effectors contribute to disease phenotypes and may also be relevant to evolution of the craniofacial skeleton17,18. Therefore, elucidating the timing and nature of tissue interactions has great potential to increase our understanding of the developmental and evolutionary biology of the facial skeleton.
The use of chimeric embryos to investigate tissue interactions has a long history in developmental biology. This approach was pioneered by Hans Spemann and his lab who discovered embryonic “organizers” by transplanting tissues between embryos of different amphibian species. Spemann was a master of micro-surgical techniques whose hand-skills were complemented by his development of specialized tools, notably the Spemann pipette. Viktor Hamburger was a graduate student in the laboratory of Hans Spemann in Freiburg during the 1920s, which is when the original transplant experiments that led to Spemann’s Nobel Prize were performed. When Hamburger moved to Washington University in St. Louis in 1935, he detailed the process of making a Spemann micropipette in his Manual of Experimental Embryology19. Drew Noden was a graduate student in Hamburger’s lab at Washington University until 1972. After moving to the University of Massachusetts, Amherst and then to Cornell University, Noden continued fabricating and using Spemann micropipettes for his surgical transplants involving quail-chick chimeras. While a graduate student, one of the authors (Rich Schneider) trained with Drew Noden at Cornell from 1995 to 1998. The following protocol for making a Spemann micropipette is based on descriptions written by Hamburger and Noden, and includes subsequent modifications made by Schneider.
The use of quail-chick chimeras for the study of craniofacial development and especially for understanding the contributions of neural crest cells was pioneered by Noden and by Le Douarin in the early 1970s, reviewed in Le Douarin et al20. This approach has been broadly adopted in many studies and by numerous other investigators1,4,5,21-38. The equivalent rates of growth and morphology of quail and chick make transplants within them ideal for the study of cell fate and lineage tracing. However, because of the similarities between quail and chick, morphological changes induced by donor cells are difficult to decipher. In contrast, other avian chimeric systems have included domestic duck as a way to study mechanisms that make embryos anatomically distinct39-50. More specifically, the quail-duck chimeric system offers multiple benefits for discerning the effects of the donor on the host, and vice versa. First, quail and duck embryos are distinct in body size and shape, which provides a direct way to explore donor- or host-specific mechanisms of pattern formation by assaying for differential domains of gene expression (Figures 1 A and B). Second, quail and duck embryos have considerably different rates of maturation, with quail hatching in 17 days and duck hatching in 28 days. Transplanted neural crest maintains its intrinsic maturation rate within the host environment, and thus, identification of temporal changes in gene expression, tissue interactions, histogenesis, and morphogenesis is possible51-57. Finally, the anti-quail nuclear antibody (Q¢PN) allows donor and host cellular contributions to be permanently distinguished from one another by recognizing a protein that is ubiquitously expressed in quail cells but absent from duck cells.
Protocol
1. Prepare Tungsten Needles
Cut a tungsten rod in half using wire cutters so that the rod does not bend.
Thread the rod through the tapered end of a 5 3/4 in borosilicate glass Pasteur pipette.
Leaving approximately 3/4 in of the rod sticking out of the glass, adhere the rod to the pipette tip with a small bead of “hot melt adhesive” (HMA), which is the glue stick used for a glue gun. The HMA should be quickly melted in an alcohol flame, taking care not to burn the glue and generate black smoke.
Using a pair of forceps, bend the tip of the tungsten rod (about 1/4 in from the top end) until a 45° angle is reached.
Holding the Pasteur pipette, place about 1/8 in of the tip of the tungsten rod into the flame of a propane torch. Hold the tip steady and exactly perpendicular to the flame.
Pull the needle out of the flame the moment a tiny orange speck of tungsten flies off of the needle.
2. Prepare Spemann Pipettes
Using a Bunsen burner, heat the narrow end of a Pasteur pipette until the glass shortly beyond the taper begins to melt. Remove from flame and pull the tip until the pipette becomes sealed. Be sure that there remains a portion of the tapered part that has an O.D. of approximately 0.7-1.0 mm. Break the sealed end off about 4 cm from the taper.
Attach around 2 ft (60 cm) of rubber tubing to the wide (open) end of the pipette. Blow on the other end of the rubber tubing to ensure that no air can pass through the tapered part of the Pasteur pipette.
Using a propane fuel cylinder with an attached pencil flame torch, heat an oval area on one side of the pipette just below the start of the taper. Blow air very gently and constantly through the rubber tubing with a slight amount of pressure (approximately the amount of pressure needed to say the beginning of the letter “P” at conversation level). When the glass becomes soft on one side and begins to buckle outwards, quickly remove the pipette from the flame and blow hard and steady through the rubber tubing. This will cause the heated part of the glass to form a sausage-shaped bubble that may pop, which is fine. If the bubble is too small, try melting and blowing the glass again. The final open window in the glass should be around 0.25 in (6.35 mm) wide by 0.5 in (12.7 mm) long.
Pointing the tapered end upwards, scrape the bubble into a glass waste container while gently blowing through the tubing to prevent glass fragments from entering the pipette.
Using the propane flame, burn off the remaining edges of the bubble and fire-polish the sides of the open window. Take care not to melt the shaft of the pipette.
Using a diamond point pencil, score the tapered end of the pipette approximately 1.25 in (30 mm) from the window and break off the tip with forceps so that the pipette opening is cut cleanly (discard pipettes with broken or uneven tips).
Using forceps and the edge of a flame from an alcohol burner, bend the narrow portion of the pipette 0.375 in (9.5 mm) from the tip to an angle of approximately 60° so that the bent portion is in a vertical plane when the window opening is at the 1:00 position for right-handed usage and 11:00 for left-handed usage. This step is best conducted in a draft-free enclosure.
Fire-polish the tip using the alcohol flame under a dissecting microscope. Be sure not to close off the tip but rather keep the opening round and smooth without any constriction.
Cut a 1 in (25 mm) piece of rubber tubing, spray the piece of tubing with 70% ethanol, and slide the tubing over the shaft of the pipette until the window opening is completely covered. This functions as the “diaphragm” of the Spemann pipette.
Place a latex rubber 2 ml bulb over the open bottom of the pipette. The bulb maintains negative pressure in the Spemann pipette.
To practice using a Spemann pipette, transfer chromatography beads of around 100-200 µm in diameter from a marked spot in one Petri dish to a marked spot in another under a microscope.
To clean the Spemann pipette, remove the rubber bulb and place pipette, tip down, in a tall beaker lined with cotton or gauze on the bottom and filled with distilled water and glassware detergent.
3. Sterilize and Incubate Eggs
Wipe eggs with 70% ethanol and place the eggs on plastic egg trays.
Set eggs in a heated incubator at 37.5 °C and 85-87%.
Incubate eggs until they reach HH9.5, approximately 26 hr for quail and 48 hr for duck.
4. Window Eggs
Remove a small piece of outer shell from the top of the egg with forceps, taking care not to puncture the inner shell membrane.
Using a syringe with an 18 G by 1-inch needle, poke a hole at the narrowest tip of the egg and withdraw approximately 1-2 ml of albumen.
Place transparent tape over the hole and puncture mark in the shell.
Cut a “window” along the top surface of the egg (through the transparent tape) using curved scissors.
5. Visualize Embryos and Prepare for Surgery
Using a glass rod, gently brush a small amount of Neutral Red (0.02 g/ml in Hank’s BSS) over the embryo.
Cut the vitelline membrane and pull it over the embryo using a flame-sharpened tungsten needle.
Re-seal the egg by placing transparent tape over the window.
6. Separate the Donor Tissue from the Donor Embryo
Remove tape from the window on the egg of the donor embryo.
To cut the neural fold from the adjacent ectoderm of the donor embryo, make slits on both sides of the neural tube, using a flame-sharpened tungsten needle (manufactured as described above).
Then, cut across the neural tube at the desired anterior and posterior levels of the graft and separate the graft from the rest of the neural tube.
Remove the neural fold from the donor embryo using either a Spemann pipette or other type of micropipette.
Then, remove the tape from the window on the host egg and use the Spemann pipette to place the donor neural fold alongside the host embryo adjacent to the region of the neural tube that will receive the transplant.
7. Separate the Host Tissue and Transplant the Donor Tissue
Separate the neural fold from the neural tube of the host embryo, as done with the donor. Take care to remove a graft of equal size as the donor tissue.
Gently push this host tissue far away from the embryo.
Then, with a blunt tungsten needle or the rounded tip of a micropipette, gently move the donor neural fold into the host neural tube, maintaining the proper antero-posterior and dorso-ventral orientations. Make sure the graft is tucked in along the sides but be sure not to poke or damage to the underlying tissues. To help keep track of the original orientation note any asymmetry in the donor tissue or differential staining from the Neutral Red. We also photo-document the donor embryo with the accompanying excised tissue, as well as the chimera immediately following surgery.
Sterile saline should be added to the egg if the host embryo shows signs of desiccation.
Now, carefully re-seal and label the egg, and gently return it to a high humidity incubator (70-80%) where the chimeric embryo can develop to the desired stage for analysis.
8. Collection of Chimeras
Be sure to collect chimeric embryos in freshly made cold Serra’s fixative and immediately place them on a rocker at 4 °C for an O/N fixation. This will allow for more sensitive detection of quail cells with the anti-quail antibody.
For gene expression analyses via RT-qPCR, freeze embryos directly in liquid nitrogen prior to RNA extraction.
Representative Results
Prior to further analysis, the efficiency of the transplant needs to be assayed. For histological, morphological, or gene expression analyses on tissue samples, quail cells should be detected by immunohistochemistry using Q¢PN antibody as described36. For RNA analyses, species-specific contributions to tissues of interest can be calculated using a PCR-based strategy58. After the efficacy of the transplant has been validated, further morphological or molecular outcome measures can be evaluated in chimeras. Interactions between the donor neural crest cells and surrounding host-derived tissues that underlie proper histogenesis and morphogenesis of the craniofacial complex have previously been extensively studied. In particular, neural crest mesenchyme directs species-specific morphology of the face7,13,51,59, feather pattern52, muscle pattern56, and cartilage53,57 through the regulation of host gene expression. For example, neural crest mesenchyme dictates when bone forms in the mandible by temporally regulating Bmp4 expression55.
Recently, investigations have centered on tissue interactions occurring very early in craniofacial development. In this regard, experiments utilizing quail-duck chimeras have shown host embryos influence neural crest migration by determining morphological boundaries. That is, in chimeric quck embryos, donor quail neural crest cells migrate into the host duck mandibular arch in a duck-like pattern (Figure 1D). Despite this host contribution to the size of the neural crest population, the donor neural crest continues to generate a mandibular skeleton that is quail-like in size and shape (Figure 1E).
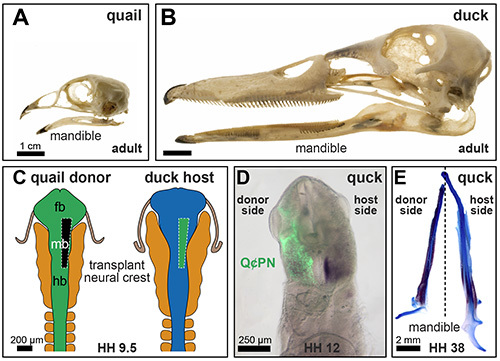 Figure 1. Quail-Duck Chimeric System. (A) Quail and (B) duck skulls display considerable differences in size and shape, and thus, are ideally suited for using a chimeric system to study craniofacial development (modified from Tokia et al.56). (C) Experimental design for generating unilateral chimeric quck embryos from stage-matched HH9.5 quail and duck embryos. The neural fold is removed from one side of quail embryos and transplanted into duck embryos after an equivalent piece of the neural fold has been removed. (D) Quail donor cells (green) can be followed in chimeras using an anti-quail antibody (Q¢PN) as shown in ventral view of HH12 chimeric embryo. (F) In quck mandibles at HH38, the quail donor-derived Meckel’s cartilage is shorter and straighter than that observed for the contralateral duck host-derived Meckel’s cartilage, which is larger and curved. Reprinted Tokita et al., Dev. Bio. 306, 377 (2007) with permission from Elsevier.
Figure 1. Quail-Duck Chimeric System. (A) Quail and (B) duck skulls display considerable differences in size and shape, and thus, are ideally suited for using a chimeric system to study craniofacial development (modified from Tokia et al.56). (C) Experimental design for generating unilateral chimeric quck embryos from stage-matched HH9.5 quail and duck embryos. The neural fold is removed from one side of quail embryos and transplanted into duck embryos after an equivalent piece of the neural fold has been removed. (D) Quail donor cells (green) can be followed in chimeras using an anti-quail antibody (Q¢PN) as shown in ventral view of HH12 chimeric embryo. (F) In quck mandibles at HH38, the quail donor-derived Meckel’s cartilage is shorter and straighter than that observed for the contralateral duck host-derived Meckel’s cartilage, which is larger and curved. Reprinted Tokita et al., Dev. Bio. 306, 377 (2007) with permission from Elsevier.
Discussion
The neural crest is a transient embryonic cell population that migrates extensively throughout the embryo and differentiates into diverse cell types, including chondrocytes and osteoblasts, that contribute to the craniofacial skeleton. Transplanting neural crest in the quail-duck chimeric system has contributed greatly to our understanding of the tissue interactions and signaling pathways that regulate development of the craniofacial skeleton. However, given the vast potential of neural crest to also generate smooth muscle cells, adipocytes, melanocytes, schwann cells, and neurons, the quail-duck chimera system has tremendous potential for future applications, particularly in conjunction with the rapid advancement of stem cell biology and regenerative medicine. Since quail and duck are both commercially bred species, a ready supply of relatively inexpensive fertilized eggs is available from a variety of farms. Thus, this technique should be accessible to researchers operating within a wide range of budgets and facility space.
Although this technique is very powerful, there remain several limitations. Like other surgical techniques, the quality and viability of quail-duck chimeras rely on the surgical skills of the researcher, and therefore, there will be more inter- and intra-individual variation between experiments as compared to other models, such as those utilizing mouse genetics. Moreover, there is also variation in the rates of development and stages of individual embryos that contributes to the reproducibility and success of each transplant. Avian embryos are also very susceptible to dehydration and therefore critical steps during surgery include keeping the light levels low, the time under the microscope to a minimum, the eggs sealed with tape as much as possible, and high humidity in the post-operative incubator to avoid desiccation.
In terms of viability of the chimeras, usually between 50-75% survive, although these percentages can decrease the older the collection stage. In a typical 4-6 hr session of surgery, an experienced surgeon can generate 10-15 chimeras. The success of the transplants also depends greatly on the quality of the tools. Good tools lead to more consistent, reproducible results. Using a propane torch to make tungsten needles allows extremely sharp needles to be made. The type of torch used makes a big difference because it controls the size of the flame. Electrolytic sharpening can also be used, but this approach does not even come close to producing needles as sharp. Use tungsten rods instead of spooled wire so that the needles can be made straight.
The Spemann micropipette, while time-consuming and difficult to make, is an ideal instrument for tissue transfer. The pipette can be customized with different-sized openings, and can be used repeatedly. A critical factor for using a Spemann micropipette is to have some fluid in the pipette before touching the tip to the surface of the embryo. Some of the fluid will always flow out when contact is made with the meniscus over the embryo. Pressing on the diaphragm allows fluid and graft tissue to be ejected very precisely, whereas slightly letting up on the diaphragm gently sucks the donor graft tissue into the pipette. Maintaining a bit of positive pressure on the diaphragm keeps the donor graft tissue at the tip of pipette during transfer, and a little additional pressure on the diaphragm allows the donor graft tissue to be deliberately placed in the host.
For protection of the Spemann pipette during storage and sterilization, remove the bulb from the wide end and carefully insert the tapered tip into the bulb. Place the Spemann pipette in a glass test tube, cover the top with aluminum foil, and autoclave prior to surgery. After multiple pipettes are ready to be sterilized again for surgery, invert the pipettes, heat them nearly to a boil in the same solution of distilled water and glassware detergent, and then rinse repeatedly with distilled water. Autoclave pipettes in their individual tubes. The rubber tubing that forms the diaphragm and the rubber bulb should be replaced after several sterilizations or when they become darkened and stiff.
Many of the components of this protocol involve dangerous equipment. For example, the procedure for making a Spemann Micropipette involves three types of flames as well as heating, pulling, blowing, bending, cutting, and polishing glass. Therefore, wearing proper personal protective equipment (PPE) that increases safety such as goggles and a lab coat is critical. Additionally, because many people suffer from, or have the potential to develop, egg allergies, always use gloves when handling eggs. With these precautions in mind, the quail-duck chimeric system is a safe, efficient, and relatively accessible method that has many future applications.
Disclosures
The authors have nothing to disclose.
Acknowledgments
This work was funded by a National Institute of Dental and Craniofacial Research (NIDCR) F32 grant (DE021929) to J.L.F. and a NIDCR R01 grant DE016402 to R.A.S.
References
- Le Lièvre CS, Le Douarin NM. Mesenchymal derivatives of the neural crest: analysis of chimaeric quail and chick embryos. J Embryol Exp Morphol. 1975;34:125–154. [PubMed] [Google Scholar]
- Le Lièvre CS. Participation of neural crest-derived cells in the genesis of the skull in birds. J Embryol Exp Morphol. 1978;47:17–37. [PubMed] [Google Scholar]
- Noden DM. An analysis of the migratory behavior of avian cephalic neural crest cells. Dev Biol. 1975;42:106–130. doi: 10.1016/0012-1606(75)90318-8. [DOI] [PubMed] [Google Scholar]
- Noden DM. The control of avian cephalic neural crest cytodifferentiation. I. Skeletal and connective tissues. Dev Biol. 1978;67:296–312. doi: 10.1016/0012-1606(78)90201-4. [DOI] [PubMed] [Google Scholar]
- Couly GF, Coltey PM, Le Douarin NM. The triple origin of skull in higher vertebrates: a study in quail-chick chimeras. Development. 1993;117:409–429. doi: 10.1242/dev.117.2.409. [DOI] [PubMed] [Google Scholar]
- Noden DM, Schneider RA. Neural crest cells and the community of plan for craniofacial development: historical debates and current perspectives. Adv Exp Med Biol. 2006;589:1–23. doi: 10.1007/978-0-387-46954-6_1. [DOI] [PubMed] [Google Scholar]
- Schneider RA. Developmental mechanisms facilitating the evolution of bills and quills. J Anat. 2005;207:563–573. doi: 10.1111/j.1469-7580.2005.00471.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperber GH. Craniofacial embryology. 4th edn. Wright; 1989. [Google Scholar]
- Chai Y, Maxson RE. Recent advances in craniofacial morphogenesis. Dev Dyn. 2006;235:2353–2375. doi: 10.1002/dvdy.20833. [DOI] [PubMed] [Google Scholar]
- Gitton Y, et al. Evolving maps in craniofacial development. Semin Cell Dev Biol. 2010;21:301–308. doi: 10.1016/j.semcdb.2010.01.008. [DOI] [PubMed] [Google Scholar]
- Szabo-Rogers HL, Smithers LE, Yakob W, Liu KJ. New directions in craniofacial morphogenesis. Dev Biol. 2010;341:84–94. doi: 10.1016/j.ydbio.2009.11.021. [DOI] [PubMed] [Google Scholar]
- Francis-West PH, Tatla T, Brickell PM. Expression patterns of the bone morphogenetic protein genes Bmp-4 and Bmp-2 in the developing chick face suggest a role in outgrowth of the primordia. Dev Dyn. 1994;201:168–178. doi: 10.1002/aja.1002010207. [DOI] [PubMed] [Google Scholar]
- Schneider RA. How to tweak a beak: molecular techniques for studying the evolution of size and shape in Darwin's finches and other birds. Bioessays. 2007;29:1–6. doi: 10.1002/bies.20517. [DOI] [PubMed] [Google Scholar]
- Richman JM, Lee SH. About face: signals and genes controlling jaw patterning and identity in vertebrates. Bioessays. 2003;25:554–568. doi: 10.1002/bies.10288. [DOI] [PubMed] [Google Scholar]
- Graham A, Okabe M, Quinlan R. The role of the endoderm in the development and evolution of the pharyngeal arches. J Anat. 2005;207:479–487. doi: 10.1111/j.1469-7580.2005.00472.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neubüser A, Peters H, Balling R, Martin GR. Antagonistic interactions between FGF and BMP signaling pathways: a mechanism for positioning the sites of tooth formation. Cell. 1997;90:247–255. doi: 10.1016/s0092-8674(00)80333-5. [DOI] [PubMed] [Google Scholar]
- Wilkie AO, Morriss-Kay GM. Genetics of craniofacial development and malformation. Nature reviews. Genetics. 2001;2:458–468. doi: 10.1038/35076601. [DOI] [PubMed] [Google Scholar]
- Fish JL, et al. modularity, and the evolvability of the vertebrate jaw. Evol Dev. 2011;13:549–564. doi: 10.1111/j.1525-142X.2011.00511.x. [DOI] [PubMed] [Google Scholar]
- Hamburger V. A Manual of Experimental Embryology. Chicago: 1942. [Google Scholar]
- Le Douarin NM, Creuzet S, Couly G, Dupin E. Neural crest cell plasticity and its limits. Development. 2004;131:4637–4650. doi: 10.1242/dev.01350. [DOI] [PubMed] [Google Scholar]
- Baker CV, Bronner-Fraser M, Le Douarin NM, Teillet MA. Early- and late-migrating cranial neural crest cell populations have equivalent developmental potential in vivo. Development. 1997;124:3077–3087. doi: 10.1242/dev.124.16.3077. [DOI] [PubMed] [Google Scholar]
- Baker CV, Stark MR, Marcelle C, Bronner-Fraser M. Competence, specification and induction of Pax-3 in the trigeminal placode. Development. 1999;126:147–156. doi: 10.1242/dev.126.1.147. [DOI] [PubMed] [Google Scholar]
- Borue X, Noden DM. Normal and aberrant craniofacial myogenesis by grafted trunk somitic and segmental plate mesoderm. Development. 2004;131:3967–3980. doi: 10.1242/dev.01276. [DOI] [PubMed] [Google Scholar]
- Cobos I, Shimamura K, Rubenstein JL, Martinez S, Puelles L. Fate map of the avian anterior forebrain at the four-somite stage, based on the analysis of quail-chick chimeras. Dev Biol. 2001;239:46–67. doi: 10.1006/dbio.2001.0423. [DOI] [PubMed] [Google Scholar]
- Couly G, Le Douarin NM. Head morphogenesis in embryonic avian chimeras: evidence for a segmental pattern in the ectoderm corresponding to the neuromeres. Development. 1990;108:543–558. doi: 10.1242/dev.108.4.543. [DOI] [PubMed] [Google Scholar]
- Couly GF, Coltey PM, Le Douarin NM. The developmental fate of the cephalic mesoderm in quail-chick chimeras. Development. 1992;114:1–15. doi: 10.1242/dev.114.1.1. [DOI] [PubMed] [Google Scholar]
- Köntges G, Lumsden A. Rhombencephalic neural crest segmentation is preserved throughout craniofacial ontogeny. Development. 1996;122:3229–3242. doi: 10.1242/dev.122.10.3229. [DOI] [PubMed] [Google Scholar]
- Noden DM, Francis-West P. The differentiation and morphogenesis of craniofacial muscles. Dev Dyn. 2006;235:1194–1218. doi: 10.1002/dvdy.20697. [DOI] [PubMed] [Google Scholar]
- Le Douarin NM. A biological cell labelling technique and its use in experimental embryology. Dev Biol. 1973;30:217–222. doi: 10.1016/0012-1606(73)90061-4. [DOI] [PubMed] [Google Scholar]
- Lwigale PY. Embryonic origin of avian corneal sensory nerves. Dev Biol. 2001;239:323–337. doi: 10.1006/dbio.2001.0450. [DOI] [PubMed] [Google Scholar]
- Marcucio RS, Cordero DR, Hu D, Helms JA. Molecular interactions coordinating the development of the forebrain and face. 2005. [DOI] [PubMed]
- Noden DM. The Role of the Neural Crest in Patterning of Avian Cranial Skeletal, Connective, and Muscle Tissues. Dev Biol. 1983;96:144–165. doi: 10.1016/0012-1606(83)90318-4. [DOI] [PubMed] [Google Scholar]
- Noden DM. Patterning of Avian Craniofacial Muscles. Dev Biol. 1986;116:347–356. doi: 10.1016/0012-1606(86)90138-7. [DOI] [PubMed] [Google Scholar]
- Olivera-Martinez I, Coltey M, Dhouailly D, Pourquie O. Mediolateral somitic origin of ribs and dermis determined by quail-chick chimeras. Development. 2000;127:4611–4617. doi: 10.1242/dev.127.21.4611. [DOI] [PubMed] [Google Scholar]
- Schienda J, et al. Somitic origin of limb muscle satellite and side population cells. Proc Natl Acad Sci U S A. 2006;103:945–950. doi: 10.1073/pnas.0510164103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider RA. Neural crest can form cartilages normally derived from mesoderm during development of the avian head skeleton. Dev Biol. 1999;208:441–455. doi: 10.1006/dbio.1999.9213. [DOI] [PubMed] [Google Scholar]
- Schneider RA, Hu D, Rubenstein JL, Maden M, Helms JA. Local retinoid signaling coordinates forebrain and facial morphogenesis by maintaining FGF8 and SHH. Development. 2001;128:2755–2767. doi: 10.1242/dev.128.14.2755. [DOI] [PubMed] [Google Scholar]
- Selleck MAJ, Bronner-Fraser M. Origins of the avian neural crest: The role of neural plate-epidermal interactions. Development. 1995;121:525–538. doi: 10.1242/dev.121.2.525. [DOI] [PubMed] [Google Scholar]
- Waddington CH. Developmental mechanics of chick and duck embryos. Nature. 1930;125:924–925. [Google Scholar]
- Waddington CH. Experiments on the Development of Chick and Duck Embryos, Cultivated in vitro. Philosophical transactions of the Royal Society of London. 1932;221:179–230. [Google Scholar]
- Hampe A. Role of mesoderm and ectoderm of leg bud in exchanges between duck and chicken. C R Hebd Seances Acad Sci. 1957;244:3179–3181. [PubMed] [Google Scholar]
- Zwilling E. Interaction between ectoderm and mesoderm in duck-chicken limb bud chimaeras. J Exp Zool. 1959;142:521–532. doi: 10.1002/jez.1401420124. [DOI] [PubMed] [Google Scholar]
- Pautou MP. Determining role of the mesoderm in the specific differentiation of the leg in birds. Arch Anat Microsc Morphol Exp. 1968;57:311–328. [PubMed] [Google Scholar]
- Sohal GS, et al. Development of the trochlear nucleus in quail and comparative study of the trochlear nucleus, nerve, and innervation of the superior oblique muscle in quail, chick, and duck. Journal of Comparative Neurology. 1985;239:227–236. doi: 10.1002/cne.902390209. [DOI] [PubMed] [Google Scholar]
- Sohal GS. Effects of reciprocal forebrain transplantation on motility and hatching in chick and duck embryos. Brain Res. 1976;113:35–43. doi: 10.1016/0006-8993(76)90004-4. [DOI] [PubMed] [Google Scholar]
- Sohal GS, et al. Synapse formation on quail trochlear neurons transplanted in duck embryos before naturally occurring motor neuron death. International Journal of Developmental Neuroscience. 1990;8:9–16. doi: 10.1016/0736-5748(90)90019-x. [DOI] [PubMed] [Google Scholar]
- Yamashita T, Sohal GS. Development of smooth and skeletal muscle cells in the iris of the domestic duck, chick and quail. Cell and Tissue Research. 1986;244:121–131. doi: 10.1007/BF00218389. [DOI] [PubMed] [Google Scholar]
- Yamashita T, Sohal GS. Embryonic origin of skeletal muscle cells in the iris of the duck and quail. Cell and Tissue Research. 1987;249:31–37. doi: 10.1007/BF00215415. [DOI] [PubMed] [Google Scholar]
- Dhouailly D. Analysis of the factors in the specific differenciation of the neoptile feathers in the duck and chicken. J Embryol Exp Morphol. 1967;18:389–400. [PubMed] [Google Scholar]
- Dhouailly D. The determination of specific differentiation of neoptile and teleoptile feathers in the chick and the duck. J Embryol Exp Morphol. 1970;24:73–94. [PubMed] [Google Scholar]
- Schneider RA, Helms JA. The cellular and molecular origins of beak morphology. Science. 2003;299:565–568. doi: 10.1126/science.1077827. [DOI] [PubMed] [Google Scholar]
- Eames BF, Schneider RA. Quail-duck chimeras reveal spatiotemporal plasticity in molecular and histogenic programs of cranial feather development. Development. 2005;132:1499–1509. doi: 10.1242/dev.01719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eames BF, Schneider RA. The genesis of cartilage size and shape during development and evolution. Development. 2008;135:3947–3958. doi: 10.1242/dev.023309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lwigale PY, Schneider RA. Other chimeras: quail-duck and mouse-chick. Methods Cell Biol. 2008;87:59–74. doi: 10.1016/S0091-679X(08)00203-3. [DOI] [PubMed] [Google Scholar]
- Merrill AE, Eames BF, Weston SJ, Heath T, Schneider RA. Mesenchyme-dependent BMP signaling directs the timing of mandibular osteogenesis. Development. 2008;135:1223–1234. doi: 10.1242/dev.015933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tokita M, Schneider RA. Developmental origins of species-specific muscle pattern. Dev Biol. 2009;331:311–325. doi: 10.1016/j.ydbio.2009.05.548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solem RC, Eames BF, Tokita M, Schneider RA. Mesenchymal and mechanical mechanisms of secondary cartilage induction. Dev Biol. 2011;356:28–39. doi: 10.1016/j.ydbio.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ealba EL, Schneider RA. A simple PCR-based strategy for estimating species-specific contributions in chimeras and xenografts. Development. 2013;140:3062–3068. doi: 10.1242/dev.092676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jheon AH, Schneider RA. The cells that fill the bill: neural crest and the evolution of craniofacial development. J Dent Res. 2009;88:12–21. doi: 10.1177/0022034508327757. [DOI] [PMC free article] [PubMed] [Google Scholar]