

Abstract
Global production of rice (Oryza sativa) grain is limited by water availability and the low ‘leaf-level’ photosynthetic capacity of many cultivars. Oryza sativa is extremely susceptible to water-deficits; therefore, predicted increases in the frequency and duration of drought events, combined with future rises in global temperatures and food demand, necessitate the development of more productive and drought tolerant cultivars. We investigated the underlying physiological, isotopic and morphological responses to water-deficit in seven common varieties of O. sativa, subjected to prolonged drought of varying intensities, for phenotyping purposes in open field conditions. Significant variation was observed in leaf-level photosynthesis rates (A) under both water treatments. Yield and A were influenced by the conductance of the mesophyll layer to CO2 (g m) and not by stomatal conductance (g s). Mesophyll conductance declined during drought to differing extents among the cultivars; those varieties that maintained g m during water-deficit sustained A and yield to a greater extent. However, the variety with the highest g m and yield under well-watered conditions (IR55419-04) was distinct from the most effective cultivar under drought (Vandana). Mesophyll conductance most effectively characterises the photosynthetic capacity and yield of O. sativa cultivars under both well-watered and water-deficit conditions; however, the desired attributes of high g m during optimal growth conditions and the capacity for g m to remain constant during water-deficit may be mutually exclusive. Nonetheless, future genetic and physiological studies aimed at enhancing O. sativa yield and drought stress tolerance should investigate the biochemistry and morphology of the interface between the sub-stomatal pore and mesophyll layer.
Introduction
Global production of rice (Oryza sativa) is limited by the availability of freshwater [1] and the tolerance of rice cultivars to drought stress [2]. Nonetheless, O. sativa is cultivated in a diverse range of climates and habitats, with lowland varieties generally considered to be more productive, but less tolerant of drought, than their upland counterparts [3]. Selective breeding of O. sativa varieties has increased the grain production of individual plants by 70% since 1965 [4]. However, these increases in productivity have been restricted by the comparatively low ‘leaf-level’ photosynthetic capacity of O. sativa relative to other C3 crops [5], [6]. Phenotypic screening of the photosynthetic and gas-exchange characteristics of O. sativa varieties under water deficit conditions may permit the identification of attributes that confer both drought tolerance and high productivity [7], [8].
Yield in varieties of O. sativa are closely related to photosynthesis under both well-watered and drought conditions [7], [9]. The uptake of CO2 for photosynthesis involves two major resistance steps at the stomata and at the surface of the mesophyll layer [10], [11]. Stomatal (g s) and mesophyll conductance (g m) to CO2 often decrease in unison in response to environmental stresses [7], [12]–[14]. Upland and lowland O. sativa cultivars exposed to a soil moisture gradient ranging from high water-availability to severe water-deficit exhibited identical reductions in both g s and g m that correlated with the rate of photosynthesis (A) and yield. Most notably, while there was no observable difference in A, g s or g m response to drought between the upland and the lowland varieties, those cultivars that exhibited the highest values of total conductance to CO2 (g t: where g t = [g s * g m]/[g s + g m]) supported higher A and yield under all levels of water availability [7]. Those O. sativa varieties with higher capacities for CO2 uptake are therefore the most likely to offer the greatest potential improvement of leaf-level photosynthesis towards achieving enhanced yields [9], [15], [16]. However, an increase in the capacity for leaf gas exchange may be accompanied by reduced drought tolerance [17], [18].
These differences in the photosynthetic and leaf gas-exchange capacities of O. sativa varieties have been ascribed to genetic variation [9]. The quantitative trait loci responsible for leaf gas-exchange and photosynthetic parameters in O. sativa tend to be found in adjacent regions of the plant genome; indicative of the exertion of selective pressures favouring coordination and high heritability of photosynthetic behaviour [19], [20]. Therefore, effective screening and analysis of the photosynthetic and gas exchange characteristics of O. sativa varieties under stress conditions has the potential to identify functional traits associated with optimisation of water use efficiency (WUE) and the maintenance of yield and A under drought [21]. Rates of photosynthesis are not only determined by diffusive CO2-uptake, but also by the activity of ribulose-1,5-bisphosphate (RubisCO) [22]. However, exposure of thirteen hydroponically grown O. sativa varieties to physiological drought did not suggest that the capacity of RubisCO carboxylation (Vcmax) or the maximum rate of electron transport required for ribulose-1,5-bisphosphate regeneration (J max) were associated with A or grain yield. Rather, A and yield were largely determined by g s and g m setting the limits for diffusional CO2 uptake [9], consistent with field based studies [7]. Furthermore, the intrinsic water-use efficiency (A/g s) of the O. sativa varieties was closely related to the ratio of g m to g s [9], and this attribute showed the greatest variability between cultivars, and thus potential in terms of possible improvements in yield and growth through maximising WUE [5], [9], [21]. In theory, those varieties with high g m to g s ratios should be most suited to maintaining photosynthesis and yield under drought stress conditions, while high values of both g m and g s should be favourable for varieties grown under constant high water availability [21].
The observation that A and yield of O. sativa closely correlate to g m [7], [9], [16] has led to a focus on the interface between the leaf mesophyll cells and internal sub-stomatal air space in the improvement of rice productivity [23]. Adachi et al. [16] crossed two varieties of O. sativa to produce a new cultivar with higher values of g m, which were associated with modifications of the mesophyll structure that increased the surface area available for CO2 uptake; thus demonstrating the potential for enhanced productivity through increased g m. In addition to morphological adaptations [15], differences in g m between O. sativa varieties are also likely affected by biochemical factors [23] such as the abundance of cooporin proteins responsible for active transport of CO2 [24], [25] (Kaldenhoff, 2012). Furthermore, the abundance and activity of these proteins has been observed to decline under water-stress [26]. Exposure to drought is associated with declines in values of A, g s and g m [11], [27]. Sustained impairment of A following the alleviation of drought stress is commonly related with continued biochemical constraints to the uptake and assimilation of CO2 [28]. However, experimental evidence suggests that O. sativa does not display prolonged reductions in A following the cessation of drought, and that the observed constraints to A under water-stress are largely physical impediments to CO2 diffusion [7], [9], [29], [30]. This suggests that artificial selection of those O. sativa varieties with high g m would enhance A while not negatively affecting drought tolerance or recovery.
Rice production is sensitive to water deficit at different developmental stages of plant growth. For example, early drought stress can affect leaf expansion [3], and later drought stress during flowering or grain-filling stages can disrupt grain formation and development [31], [32]. The photosynthetic and yield responses of seven O. sativa varieties (five upland and two lowland varieties chosen to represent genetic variability) were analysed in response to sustained drought. As different O. sativa varieties have different phenologies (for example Van forms flower inflorescences relatively earlier than Moro), through the use of a drip irrigation system it was possible to apply the stress at the same developmental stage for each variety. This has the advantage over a previous study into the drought tolerance of O. sativa varieties conducted at the IRRI in that all varieties experienced drought at the same phenological stage, therefore eliminating a crucial source of error whereby early flowering varieties outperformed late developing cultivars [cf. 7]. Mesophyll conductance to CO2 is of key interest to the productivity and drought tolerance of O. sativa [7], [16], however it is not possible to directly measure g m, and due to the uncertainties associated with its calculation [33] (particularly in the use of the variable J method under stress conditions) it is preferable to simultaneously use two independent methodologies in the estimation of g m [34]. We therefore employed two independent methodologies in the estimation of g m: firstly, based upon the synchronous measurement of leaf gas exchange and chlorophyll fluorescence parameters (variable J), and secondly, the analysis of carbon isotope discrimination (Δ13C) in recently formed metabolites to assess their effectiveness in an analysis of O. sativa drought tolerance in the field. Further details of the variable J and ‘Δ13C of recently synthesised sugars’ methods of g m analysis are presented in the Material and Methods section. This study aimed to: 1) assess the effect of g s and g m on A and yield in well-watered and drought conditions; 2) analyse differences in g m calculation between the ‘variable J’ [35] and ‘Δ13C of recently synthesised sugars’ [15] methods, and; 3) identify O. sativa varieties that exhibit drought tolerance and/or high productivity, and may be suitable for further genomic/biochemical development to further enhance grain production.
Materials and Methods
Plant material and growth conditions
The experiment was conducted at the International Rice Research Institute Experimental Station at Los Banos, Philippines (14° 11′N 121° 15′E, 21 m above sea level) during the dry season (January to May). Rainfall was minimal, however, temperature and leaf to air vapour pressure difference (VPD) increased during the experimental period. The soil at the experimental site is a neutral Andaqueptic Haplaquol (pH = 6.5). Seven O. sativa cultivars were studied: Apo, IR55419-04 (IR55), IR64, IR71525-19-1-1 (IR71), Moroberekan (Moro), PSBRc80 (PS80) and Vandana (Van). Moroberekan failed to set or produce grains and seemed to suffer from disease and heat sensitivity, particularly under drought conditions and it was not possible to record physiological measurements for this variety under water deficit stress conditions. Seeds were sown in dry soil at a rate of 80 kg ha−1 in rows spaced ∼25 cm apart. Fertilisation with nitrogen was conducted on two occasions at application rates of 90 and 120 kg ha−1, alongside fertilisation of 30 kg ha−1 of phosphate and potassium. Weeds were controlled by the application of standard agricultural herbicides and manual picking. Pesticides were used to control insect pests; in particular stem borers.
Experimental plots for each O. sativa variety measured 3 m×2 m, with adjacent irrigated and drought-stressed treatments. The plots were arranged in a randomized block design. A sprinkler system was initially used to provide equal irrigation levels to all plants until flower (panicle) development, when the water deficit treatment was instigated. During the drought stress period, the well-watered treatments were irrigated by use of a drip irrigation system, with soil water potential maintained above -0.04 MPa (monitored using tensiometers placed at soil depths of 15 and 30 cm) – the level at which the more sensitive rice cultivars begin to exhibit a reduction in transpiration [36].
Leaf gas exchange measurements
Leaf gas-exchange and fluorescence of plants grown under both well-watered treatment or stress conditions were simultaneously measured with a LI-6400-40 leaf chamber fluorometer (Li-Cor, Inc., Nebraska, USA) on flag leaves enclosed in a temperature, light and humidity controlled cuvette. The measurements were made in situ between 11.00 and 15.00, in saturating PPFD (1400 µmol m−2s−1), with relative humidity ranging between 45–55%, and a leaf temperature of 30°C. Instantaneous measurements of steady-state A, g s, internal [CO2] (C i) and the quantum yield of PSII in the light (ΔF/Fm') were made on six to 14 plants per treatment. Measurements of dark respiration (R d) were also made at ambient CO2 concentration in the dark on the same leaves. Instantaneous transpiration efficiency was calculated as the ratio of A to g s.
Carbon isotope discrimination
Three of the same flag leaves used for in situ gas exchange measurements were collected in the evening, oven dried at 70°C until their weight remained constant and were then stored until soluble sugar extraction, following the protocol of Richter et al. [37]. Leaves were finely ground in liquid nitrogen and shaken for 60 min in water at room temperature. After centrifugation (15 min at 5000 g), the supernatant was sequentially mixed with cationic (Dowex-50) and anionic (Dowex-1) exchange resins. The residual solution of purified soluble sugars was freeze-dried and carbon isotope composition (δ13C) determined using a mass-spectrometer. Additional isotope ratio mass spectrometry analyses were performed on the pellet samples that remained after the extraction of soluble sugars. Samples of approximately 1 mg were used for δ13C analysis. Stable isotope ratios were determined using a continuous-flow triple-collector isotope ratio mass spectrometer (ISOPRIME, GV, Manchester, UK). Solid samples were quantitatively combusted in an elemental analyser (Model NA 1500, Carlo Erba, Milan, Italy) and CO2 was transferred in helium flow to the mass spectrometer. Calculations of carbon isotope discrimination (Δ13C) of solid samples were undertaken following the protocol of Farquhar et al. [38], assuming the carbon isotopic composition of CO2 in air (δair) to be −8.0‰.
Estimation of mesophyll conductance
Mesophyll conductance to CO2 diffusion is the inverse of the total resistance encountered in the transport of CO2 across the leaf mesophyll. Estimation of g m is subject to a number of assumptions and sources of error, and it is preferable to employ two independent approaches in its calculation [34], [39]. In this study g m was calculated using the ‘variable J’ and ‘Δ13C of recently synthesised sugars’ methods. The calculation of g m using the variable J method is based on simultaneous measurements of gas-exchange and chlorophyll fluorescence parameters as described by Harley et al. [35] and Loreto et al. [40] (equations 1 and 2):
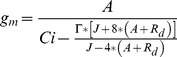 |
(1) |
where the electron transport rate (J F) is calculated from fluorescence [41]:
| (2) |
.
Where F m is the fluorescence maximum and the partitioning factor (β) between photosystems I and II was considered to be 0.5 and leaf absorbance (α) was measured directly [42]. The variable J method is sensitive to the estimation of the CO2 compensation point to photorespiration (Γ*) and R d. For the results of this study to be directly comparable with those of Centritto et al. [7], the value of Γ* used in the gas exchange algorithm was calculated using the Rubisco specific factor for an annual herb [43], consistent with previously published O. sativa Γ* values [44]. As measurements were conducted under field conditions it was not feasible to produce robust measurements of respiration in the light (R l); respiration in the dark was therefore used in the calculation of gm, as R l and R d have been observed to be broadly consistent when recycling of CO2 is taken into consideration [45]. Measurements of R d were performed at ambient [CO2] concentration in the dark on the same leaves by switching off the light in the leaf cuvette; when CO2 release from the leaf had become stable for approximately five to ten minutes this was recorded and considered to represent R d. To calibrate the values of J calculated by fluorescence (J F) and gas-exchange (J A), measurements were also conducted under non-photrespiratory conditions using 1% [O2] [35], [40]. Electron transport rate and R d values are presented in Figures S1 and S2. Total conductance to CO2 (g t) was calculated as: g t = g s * g m/(g s + g m). The concentration of CO2 at the chloroplast site (C c) was then calculated from mesophyll conductance values [46].
The method described by Lauteri et al. [47] for ‘on-line’ determinations was adapted to calculate g m by using the Δ13C of recently synthesized sugars [48]. Discrepancies between Δ13C of leaf soluble carbohydrates (Δobs) and Δ13C expected on the basis of gas-exchange measurements (Δexp), allow the estimation of g m utilising the approach of Evans et al. [49] that was subsequently modified by Lloyd et al. [50]:
| (3) |
Where: b is the discrimination associated with carboxylation reactions, taken to be 27.5‰; b s is the fractionation occurring when CO2 enters solutions (1.1‰ at 25°C); a 1 is the fractionation during diffusion in water (0.7‰); f is the fractionation associated with photorespiration, and; p a is the partial pressure of CO2 in air. Two g m calculations were performed and compared, since two values were taken into account for the fractionation factor f: 7‰ [50] and 0‰ [48], [51]. The closest relationship between g m estimates (R2 = 0.911) using the variable J and Δ13C of recently synthesized sugars methods in all varieties was found when using a f value of 0‰.
Harvesting
Plants were harvested at physiological maturity by sampling 1 m of the 2 central rows of each plot. Data collected included biomass, plant height, anthesis date, tiller number, straw production, grain yield, and harvest index (HI).
Statistical analysis
Analyses of variance were conducted using SAS to generate least squares means for each entry. Leaf gas exchange data were tested using a simple factorial ANOVA (three-way maximum interactions) and, where appropriate, the treatment means of leaf properties and gas exchange parameters were compared using a Tukey post-hoc test.
Results
Drought stress induced reductions in yield, A and rates of gas-exchange across all the O. sativa varieties analysed in this study. The duration of time required for 50% of plants to develop flowers (flower days) did not correlate with yield in well-watered plants, but those varieties with a shorter flowering day period exhibited enhanced yield under drought stress (Fig. 1a). Varieties with shorter flowering days allocated a higher proportion of their total biomass to yield (harvest index: HI) under control and drought treatments (Fig. 1b). Positive allometric correlations between plant height and both yield (Fig. 1c) and HI (Fig. 1d) were observed, both under watered or drought conditions.
Figure 1. The performance of seven rice varieties under well-watered (open symbols) and drought conditions (closed symbols): a) relationship between yield and flowering days (period of time for 50% of plants to develop flowers) under full-water (linear regression: R2 = 0.0150; F 1,4 = 0.0609; P = 0.817) and drought (linear regression: R2 = 0.749; F 1,5 = 14.905; P = 0.0119); b) allometric relationship between yield and plant height (linear regression: R2 = 0.637; F 1,10 = 17.536; P = 0.00187); c) relationship between harvest index (HI:dry weight of grain relative to dry total plant biomass) and flowering days under full-water (linear regression: R2 = 0.637; F 1,5 = 8.760; P = 0.0315) and drought (linear regression: R2 = 0.752; F 1,5 = 15.144; P = 0.0115), and; d) relationship between HI and plant height (linear regression: R2 = 0.430; F 1,10 = 7.548; P = 0.0206).

Error bars indicate one standard error either side of the mean. Numbers next to data points indicate Oryza sativa variety: 1 = Apo; 2 = IR55; 3 = IR64; 4 = IR71; 5 = Moro; 6 = PSBRc80; 7 = Van.
The carbon isotopic composition of the leaves of the O. sativa varieties became increasingly enriched in the heavier 13C isotope following water-stress, with the exception of IR64. Changes in the carbon isotopic composition of soluble sugars induced by the drought treatment were less consistent, with three varieties exhibiting increases and four decreases in the ratio of stable carbon isotopes (Table 1). Despite the lack of genotype x water treatment interaction, the varieties yielded differences in the ‘long-term’ isotopic signal of the bulk leaf material; some genotypes showed wide variation (up to −1.5‰ in pellets of Van) while others were more negligible (e.g. 0.2‰ in pellets of IR64). Drought caused also different variation in Δ13C of soluble sugars: up to −3.5‰ in PS80 but 0.2‰ in IR71. Under both well-watered and rainfed conditions, soluble sugars showed generally lower values of carbon isotopic discrimination when compared with the average Δ13C values of the flag leaf pellets. Significant differences were observed in the carbon isotopic discrimination values of of leaf pellets between varieties, however this was not replicated in the analysis of soluble sugars (Table 1).
Table 1. Carbon isotope discrimination (‰) of the flag leaf pellet (bulk) and soluble sugars of seven rice cultivars grown under drought and well-watered conditions in ‰.
| Genotype | Habitat | Bulk Leaf Δ13C (‰) | Sugars Δ13C (‰) | ||||
| Well-watered | Drought | Mean | Well-watered | Drought | Mean | ||
| Apo | U | 21.6±0.9 | 20.6±0.3 | 21.0 ab | 18.3±1.3 | 18.7±0.6 | 18.5 a |
| IR55 | U | 21.4±0.5 | 20.8±0.0 | 21.1 a | 18.2±0.5 | 16.2±0.9 | 17.2 a |
| IR64 | L | 20.9±0.2 | 21.1±0.5 | 21.0 ab | 18.5±0.6 | 16.6±0.8 | 17.5 a |
| IR71 | U | 21.1±0.2 | 20.7±0.5 | 20.9 ab | 17.3±1.3 | 17.5±0.7 | 17.4 a |
| Moro | U | 19.9±0.4 | 19.7±0.2 | 19.8 b | 18.3±0.6 | 19.5±0.2 | 18.9 a |
| PS80 | L | 21.4±0.4 | 21.2±0.4 | 21.2 a | 19.3±0.3 | 15.8±0.2 | 17.2 a |
| Van | U | 21.5±0.3 | 20.0±0.5 | 20.7 b | 18.1±1.5 | 15.8±0.4 | 17.0 a |
| Mean | 21.1 a | 20.6 b | 18.2 a | 17.2 b | |||
Values indicate the mean of six plants; ± indicates the standard error of mean; different letters indicate significant differences (P≤0.05) among means according to an ANOVA.
A strong positive correlation was observed between the two estimates of g m in the O. sativa varieties grown under control and drought conditions (R2 = 0.930; P<0.01) (Fig. 2), excluding IR64 and PS80. Indeed, the lowland varieties IR64 and PS80 grown under well-watered conditions exhibited high values of g m derived from the application of the isotopic methodology. It was therefore decided to utilise g m measurements resulting from the variable J method in the subsequent analyses [an extended discussion of the contrasting merits of the approaches is given in 7]. Rates of A, g s and g m all showed consistent declines under water-stress conditions (Fig. 3). IR55 showed the highest yield and g m under well-watered conditions. Variety IR71 exhibited the highest general values of A, g s and g m under both well-watered and drought conditions, while Apo and PS80 showed the lowest values of these photosynthesis and gas-exchange parameters under control and water-stress. The A/g s ratio of the O. sativa varieties either increased (IR71; Van; IR55) or remained constant under drought stress in all varieties with the exception of PS80, where a 42.6% reduction was observed (Fig. 3d). The ratio of [CO2] within the substomatal pore (C i) to the external atmospheric [CO2] (C a) was largely unaffected by water-stress (Fig. 4a). In contrast, the ratio of the [CO2] within the chloroplast envelope (C c) to C a was diminished in all O. sativa varieties during drought treatment (Fig. 4b), indicative of transport limitations of CO2 in the mesophyll for photosynthesis. Oryza sativa variety IR71 showed the smallest proportional reduction of 16.8% in the C c/C a ratio, while Apo exhibited the largest reduction of 33.8%.
Figure 2. Comparison between the estimates of mesophyll conductance of CO2 (g m) obtained by applying two independent methods: the variable J method and the ‘δ13C of recently synthesised sugars’ method (linear regression: R2 = 0.932; F 1,9 = 122.750; P = 1.515×10−6).
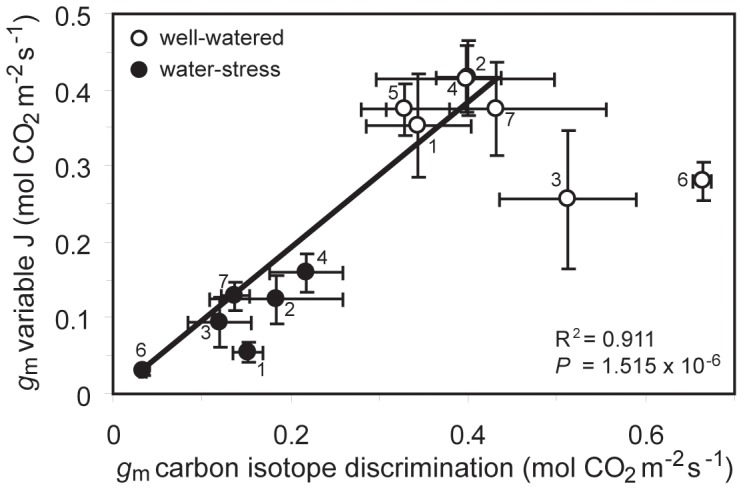
Each data point represents the average value of three observations based upon the Δ13C of recently synthesised sugars and six to fourteen gas-exchange measurements utilising the variable J method. Error bars as in Figure 1. The regression line excludes the two data points on the right of the graph (IR64 and PS80) with anomalously high g m derived from the δ13C of recently synthesised sugars. Numbers next to data points indicate Oryza sativa variety as in Figure 1.
Figure 3. Measurements of (a) photosynthesis rate (A), (b) stomatal conductance (g s), (c) mesophyll conductance (g m), and (d) intrinsic transpiration efficiency (A/g s) in control and water-stressed leaves of the seven Oryza sativa genotypes.
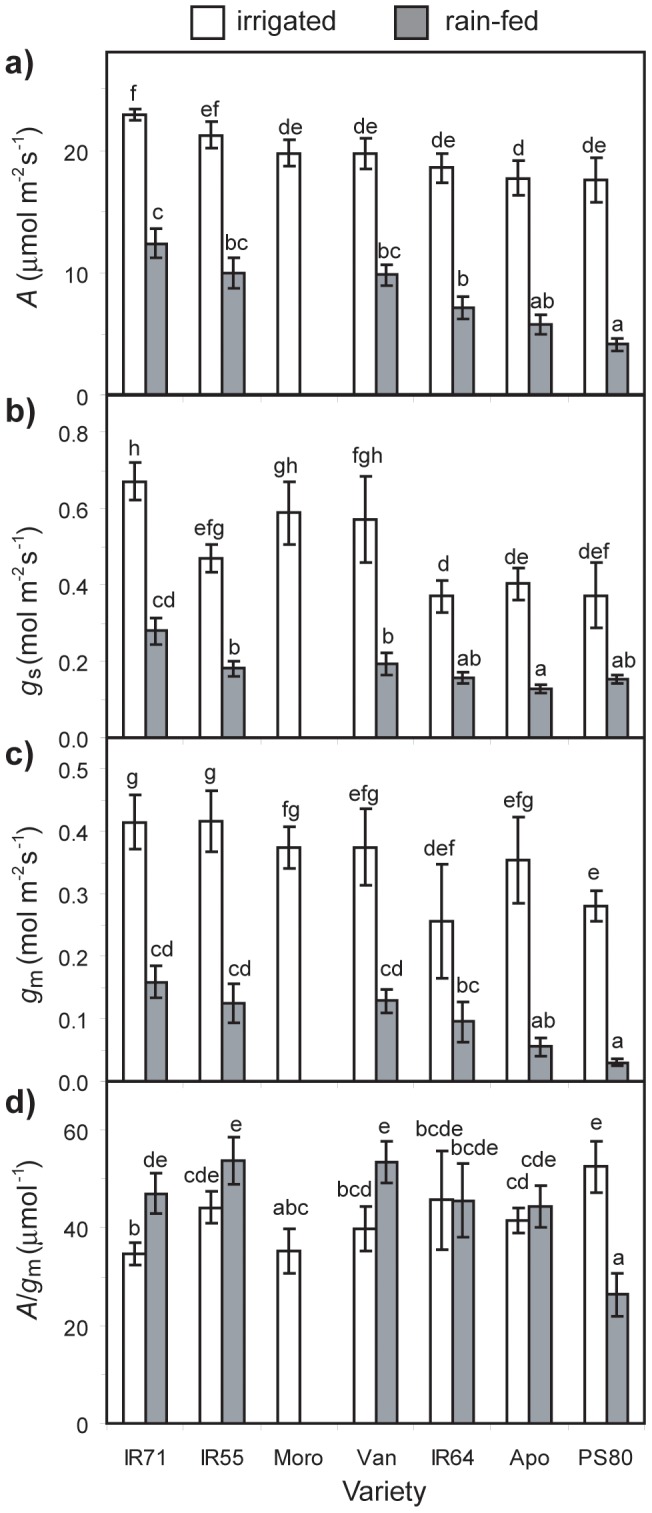
The measurements were made on the flag leaf in saturating PPFD (1400 µmol m−2s−1), with relative humidity ranging between 45–55%, and a leaf temperature of 30°C. Data are means of 4 to 7 plants per treatment. Error bars as in Figure 1. Different letters denote significant differences among means derived using a factorial ANOVA and Tukey post-hoc test.
Figure 4. Measurements of (a) the intercellular [CO2] (C i) to the ambient [CO2] (C a) ratio (C i/C a), and (b) the chloroplastic [CO2] (C c) to the ambient [CO2] ratio (C c/C a) in control and water-stressed leaves of the seven Oryza sativa genotypes.
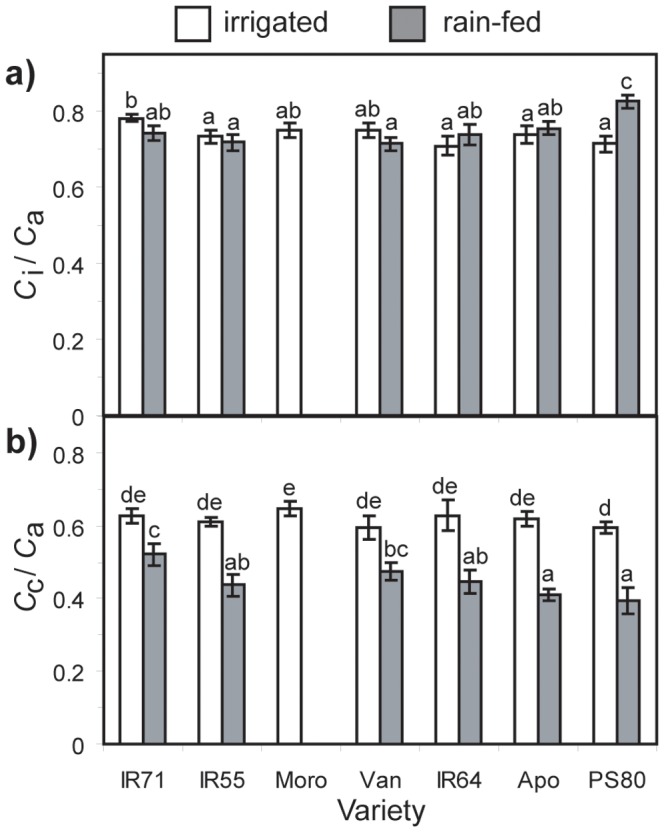
The measurements were made on the flag leaf in saturating PPFD (1400 µmol m−2s−1), with relative humidity ranging between 45–55%, and a leaf temperature of 30°C. Data are means of 4 to 7 plants per treatment. Error bars as in Figure 1. Different letters denote significant differences among means derived using a factorial ANOVA and Tukey post-hoc test.
Yield and A exhibited significant positive correlations with g s (Fig. 5a and 5b). However, the positive correlations between yield and A with g m (Fig. 5c and 5d) were stronger than those observed with g s or total conductance (g tot); suggesting that g m is the major determinant of yield and A under both well-watered and water-stressed conditions. The yield of the O. sativa varieties was strongly related to A under both control and drought treatments (Fig. 5g). In order to gauge the response of these diffusive limitations to CO2 transport and their effect on yield and A, the proportional change (Δ) in response to drought was calculated. The proportional reduction in yield (Δyield) was not related to Δg s, Δg m or Δg tot (Fig. 6a; 6b; 6c). However, ΔA was significantly correlated to Δg m and Δg tot. IR71 exhibited the highest levels of A and g m (Fig. 3a and 3c), but also the lowest proportional reduction in g m and A (Fig. 6d). The highest yields observed under control conditions were found in IR55 (3039.6±359.1 kg ha−1) and IR71 (2407.4±284.4 kg ha−1); under water-stress the yield of IR55 fell to 552.7±80.3 kg ha−1 and IR71 to 640.3±93.0 kg ha−1. The lowest Δyield reduction under drought conditions was observed in variety Van (Fig. 6), which also produced the highest yield under water-stress (935.3±206.0 kg ha−1). IR71 exhibited the second lowest Δyield and produced second largest yield during water-stress of the varieties analysed (Fig. 6g). The A/g s ratio was not significantly related to either yield or HI under both well-watered or drought conditions (Fig. 7a and 7c). However, the g m:g s ratio was significantly correlated with both yield and HI (Fig. 7b and 7d), suggesting that those O. sativa varieties with greater g m/g s ratio will exhibit enhanced productivity under both well-watered and water-stressed conditions.
Figure 5. Interaction of diffusive conductance parameters to CO2 uptake with yield and photosynthesis (A) under well-watered (open symbols) and drought conditions (closed symbols): a) relationship between yield and stomatal conductance (g s) (linear regression: R2 = 0.696; F 1,10 = 22.900; P = 0.000740); b) relationship between A and g s (linear regression: R2 = 0.873; F 1,11 = 75.721; P = 2.911×10–6); c) relationship between yield and mesophyll conductance (g m) (linear regression: R2 = 0.850; F 1,10 = 56.611; P = 2.911×10–5); d) relationship between A and g m (linear regression: R2 = 0.952; F 1,11 = 217.071; P = 1.376×10–8); e) relationship between yield and total conductance (g tot) (linear regression: R2 = 0.806; F 1,10 = 41.418; P = 7.483×10–5); f) relationship between A and g tot (linear regression: R2 = 0.952; F 1,11 = 216.173; P = 1.407×10–8), and; g) relationship between yield and A (linear regression: R2 = 0.795; F 1,10 = 38.665; P = 9.909×10–5).

Error bars as in Figure 1. Numbers next to data points indicate Oryza sativa variety as in Figure 1.
Figure 6. Changes in yield and photosynthesis in relation to modification of diffusive resistances to CO2 uptake following water-stress.

Those varieties that experienced smaller reductions in parameters were more tolerant of drought. a) relationship between Δyield and Δg s (linear regression: R2 = 0.337; F 1,4 = 2.032; P = 0.227); b) relationship between Δyield and Δg m (linear regression: R2 = 0.134; F 1,4 = 0.618; P = 0.476); c) relationship between Δyield and Δg tot (linear regression: R2 = 0.0818; F 1,4 = 0.356; P = 0.583); d) relationship between ΔA and Δg s (linear regression: R2 = 0.0003; F 1,4 = 0.00106; P = 0.976); e) relationship between ΔA and Δg m (linear regression: R2 = 0.742; F 1,4 = 11.527; P = 0.0274); f) relationship between ΔA and Δg tot (linear regression: R2 = 0.715; F 1,4 = 10.042; P = 0.0339), and; g) relationship between Δyield and ΔA (linear regression: R2 = 0.427; F 1,4 = 2.979; P = 0.159). Error bars as in Figure 1. Numbers next to data points indicate Oryza sativa variety as in Figure 1.
Figure 7. Interaction of yield and A with transpiration efficiency (A/g s) and the ratio of g m to g s in well-watered (open symbols) and drought conditions (closed symbols): a) relationship between yield and A/g s under full (linear regression: R2 = 0.0595; F 1,4 = 0.253; P = 0.641) and water-stressed (linear regression: R2 = 0.434; F 1,4 = 3.072; P = 0.155) conditions; b) relationship between harvest index (HI) and A/g s under full (linear regression: R2 = 0.205; F 1,4 = 1.032; P = 0.367) and water-stressed (linear regression: R2 = 0.185; F 1,4 = 0.909; P = 0.394) conditions; c) relationship between yield and g m:g s (linear regression: R2 = 0.456; F 1,10 = 8.379; P = 0.0160), and; d) relationship between HI and g m:g s (linear regression: R2 = 0.359; F 1,10 = 5.610; P = 0.0394).

Error bars as in Figure 1. Numbers next to data points indicate Oryza sativa variety as in Figure 1.
Discussion
The seven O. sativa varieties exhibited a diverse range of morphological, isotopic, photosynthetic and leaf gas-exchange responses to drought stress. A number of factors determined the yield of the O. sativa cultivars analysed in this study. Despite water deficit being instigated at the same developmental stage for all varieties, phenology had an effect on yield under drought as rapidly developing plants maintained yield to a greater extent than slower developers (Fig. 1a) [52]. Despite the type and phenolgical timing of the drought stress being different to that utilised in the investigation of Centritto et al. [7], the effect on A and yield observed in the two studies are broadly consistent. Those plants that were able to accumulate biomass rapidly produced higher grain yields than those that accumulated biomass less efficiently under both well-watered and drought conditions. This may be due to allometric relationships between biomass and yield [53], or the positive influence of increased photosynthetic area on ‘whole-plant’ carbon uptake [54]. Leaf-level photosynthetic rates were also closely related to yield in all varieties under both well-watered and drought conditions (Fig. 5g). Photosynthesis rates were in turn determined by diffusive limitations to CO2-uptake (Fig. 5b; 5d; 5f). The most significant of these resistances to CO2-uptake in O. sativa occurs at the interface between the internal sub-stomatal air-space and the mesophyll layer [15], [55]. Consistent with other studies, g m in O. sativa correlated most closely with A and yield (Fig. 5d) [7], [9], and represents a critical ‘rate-limiting’ step in the determination of grain productivity and assessment of the photosynthetic response to drought stress.
The ‘bulk’ carbon isotope discrimination values of the O. sativa varieties decreased following drought treatment; indicative of water-deficits over the life-span of the flag-leaf reducing discrimination against the heavier 13C isotope [56], [57]. However, this pattern of reduced carbon isotope discrimination is not apparent in the carbon isotopic composition of recently synthesised sugars (Table 1). These findings may reflect ‘shorter-term’ variations in WUE in the plants experiencing drought stress, a modification of the photosynthetic pathway affecting sugar metabolism [28] or the breakdown of previously stored carbohydrates with a different isotopic signature into more simple sugars over the course of the stress treatment [58], [59]. Differences in the diffusional uptake of CO2 likely affected carbon isotopic composition [60]; however, this was not apparent from our g s, g m (variable J) and g tot datasets (Fig. 5), suggesting that other factors affected carbon isotopic discrimination and possibly the effectiveness of the Δ13C of recently synthesised sugars method of estimating g m in varieties IR64 and PS80 under control conditions (Fig. 2). It is noteworthy that under water-deficit conditions, g m estimates based on the variable J method closely correlated with those derived from the Δ13C of recently synthesised sugars (Fig. 2). These findings do not support the suggestion that reductions in C i may affect estimation of the total photosynthetic electron transport rate associated with enhanced photorespiration, thus rendering the variable J method unreliable in the investigation of g m in plant responses to environmental stress [34].
Selective breeding has improved the productivity of O. sativa varieties through improved growth form and the increased proportional allocation of biomass as grain [4]; yet these increases in yield have been constrained by the relatively low leaf-level photosynthetic rate of O. sativa in comparison to other C3 crops [5], [61]. The results of this study indicate that under well-watered and water-deficit conditions, A is an important determinant of yield in O. sativa and is largely related to the rate of transport of CO2 across the mesophyll (Fig. 5d). Further improvements in O. sativa grain yield may likely be achieved through the enhancement of g m to CO2 inducing enhanced A [6], [9], [16], [62]. Those O. sativa varieties that showed the greatest photosynthetic performance and yield under control conditions were not necessarily the most effective varieties under drought conditions. Varieties that maintained transport of CO2 during water-stress sustained A and yield to a greater extent than varieties that exhibited more pronounced diffusive limitations to photosynthetic CO2-uptake (Fig. 5). Drought stress induced a relatively consistent mean reduction in g s in the O. sativa varieties (Fig. 3), yet the C i:C a ratio was unaffected (Fig. 4), indicating that g s was not limiting to A under water-deficit, or that the gas-exchange data were possibly biased by the occurrence of non-uniform stomatal closure over the leaf lamina and consequent overestimation of C i [27], [63], [64]. However, the C i/C a ratio in the O. sativa varieties was reduced by 16.8 to 33.8% after drought, consistent with the impairment of A through mesophyll diffusive limitations [7], [11], [12], [65], [66]. Mesophyll conductance to CO2 declined during the drought treatment, constraining A and growth (Fig. 5; 6). However, significant variability in the extent of reductions in g m were observed in the O. sativa cultivars ranging from −61.7% (IR71) to −89.3% (PS80). Furthermore, those varieties that showed the smallest reductions in g m also exhibited greater A; demonstrating that g m largely determines A in O. sativa under differing degrees of water-deficit [7], [9]. These differences in the ability to sustain g m values under drought stress are likely related to the maintenance of biochemical function in the mesophyll such as the activity of cooporin and carbonic anhydrase [eg. 25,67].
The conductance of the mesophyll layer to the transport of CO2 determined A and yield in the seven O. sativa varieties studied. Under conditions of water-stress it has been suggested that a low g s (to reduce transpirative water-loss) and a high g m (to maintain the uptake of CO2) are beneficial [11], [68]. However, in the case of O. sativa, yield and HI were only weakly related to the ratio of g m:g s (R2<0.460) (Fig. 7). The reduction in g s under water-deficit is consistent with active stomatal control in the optimisation of WUE [69], [70]; nevertheless, the lack of a strong correlation to the g m:g s ratio may be indicative of the lack of a stomatal limitation to A under drought stress in the O. sativa varieties apparent in the C i:C a ratios (Fig. 4a). The photosynthetic and yield responses of O. sativa under well-watered and drought conditions were more dependent upon g m than g s (Fig. 5b and 5d). This is consistent with earlier studies reporting that modification and improvement of g m is more effective in the enhancement of O. sativa productivity [16] and likely drought tolerance [7], [9]. The strong dependence of A and yield on g m is also evident in the lack of a relationship between A/g s and yield or HI (Fig. 7a and 7c). This may suggest that stomatal control is either less effective [70], [71], or plays a reduced role, in respect to mesophyll limitations to CO2 in a drought intolerant crop such as O. sativa in comparison to crops adapted to more water-limited environments [72].
The disparity in A values observed between the cultivars raises the possibility of crossing different varieties to enhance leaf-level A and yield in subsequent generations [6], [8], [16]. Selection of drought tolerance on the basis of the ability of a particular genotype to maintain g m function under drought stress would be viable [11], [73]. However, genetic sequencing and identification of quantitative trait loci permit the possibility of extracting genetic material from a O. sativa cultivar with a valuable characteristic (such as high g m and/or stable g m under water deficit) and in the creation of a new variety with a suite of desired attributes [17], [19], [74]–[76]. For example, from the results of this study it would be desirable to combine the high g m and yield of IR55 under well-watered conditions with the capacity to sustain g m under water-stress exhibited by IR71 and water-stressed yield of Van (Fig. 3). In this context it may be possible to phenotypically screen a large number of O. sativa genotypes to identify quantitative trait loci responsible for g m values and g m response to drought. Those varieties developed to be resistant to drought may be suited to more marginal land-types or in scenarios of insufficient water availability. Nonetheless, given the sporadic and unpredictable nature of sustained and persistent drought events it is unlikely that large numbers of farmers would select a drought tolerant variety such as Van over a more productive cultivar such as IR55, and thus limit their harvest to accommodate the risk of interruption to water supplies [73], [77]. An ideal O. sativa variety would therefore possess both values of g m capable of sustaining high A and the ability to maintain g m in the event of water-deficits [8], [78]. However, the results of this study suggest that those attributes may be mutually exclusive in the species analysed. Furthermore, no single parameter analysed in this study exclusively accounted for yield under either well-watered or drought conditions; therefore attempts to improve the productivity of O. sativa cultivars may also involve selection for a suite of characteristics that improve productivity under specific conditions of water availability. Nonetheless, future efforts to improve O. sativa productivity should focus on biochemical [eg. 25] and physical modification [eg. 16] of the interface between the mesophyll layer and sub-stomatal air-space to enhance CO2 uptake and A.
Conclusions
Physiological analysis of O. sativa has great potential in identifying those photosynthetic and leaf gas-exchange characteristics that confer attributes such as high yield and tolerance to drought. The analysis of the seven O. sativa varieties indicates that under both well-watered and drought stressed conditions yield and A are determined by diffusional constraints to CO2 uptake and not by biochemical limitations. The varieties showed significant variation in leaf-level photosynthetic rates, symptomatic of the photosynthetic constraints to O. sativa production that have previously been recorded. Selection of those O. sativa genotypes that exhibit higher rates of leaf-level A would likely be beneficial for grain yields. Photosynthesis was most closely related to g m, and the tolerance of the O. sativa varieties to drought was largely determined by the ability to maintain g m values under water-deficits. However, the highest g m and yield under well-watered conditions (IR55) and the most pronounced ability to sustain g m under water-deficit (IR71) were observed in different varieties. This may suggest that the desired attributes for enhanced yield during high water availability and the ability to maintain g m, and thus sustain grain production under water-deficits, may be mutually exclusive. Nonetheless, the results of studies such as this could make it possible through selective breeding, or the identification of a particular genetic expression associated with a desired attribute, to increase leaf-level photosynthetic rates and drought tolerance in the same variety of O. sativa. These findings indicate that the interface between the internal sub-stomatal air-space and the mesophyll layer offers the greatest potential in terms of possible modification to enhance the performance of O. sativa under optimal and stressed growth conditions in the future.
Supporting Information
Dark respiration ( R d) values of seven Oryza sativa varieties under irrigated (white) and rain-fed (grey) conditions used in the calculation of g m using the variable J method.
(PDF)
Electron transport rate (ETR) values of seven Oryza sativa varieties under irrigated (white) and rain-fed (grey) conditions used in the calculation of gm using the variable J method.
(PDF)
Acknowledgments
The following are thanked for scientific discussion and technical assistance: L. Spaccino for IRMS analyses (CNR-IBAF) and R. Torres G. Dimayuga and the drought physiology team at IRRI for technical support (IRRI, Philippines). The comments of two anonymous referees significantly improved this manuscript.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.
Funding Statement
This work was funded by: Ministero dell'Istruzione, dell'Università e della Ricerca of Italy (PRIN) grant: “Going to the root of plant productivity: how the rhizosphere interact with the aboveground armament for indirect and direct defense against abiotic and biotic stressors (PRO-ROOT)” URL: http://hubmiur.pubblica.istruzione.it/web/universita/home and Marie Curie Intra European Fellowship (2010-275626) - HAWORTH - Evoutionary adaptation to atmospheric carbon dioxide, URL: http://ec.europa.eu/research/mariecurieactions/about-mca/actions/ief/index_en.htm. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Pandey S, Bhandari H, Ding S, Prapertchob P, Sharan R, et al. (2007) Coping with drought in rice farming in Asia: insights from a cross-country comparative study. Agricultural Economics 37: 213–224. [Google Scholar]
- 2. Serraj R, McNally KL, Slamet-Loedin I, Kohli A, Haefele SM, et al. (2011) Drought resistance improvement in rice: an integrated genetic and resource management strategy. Plant Production Science 14: 1–14. [Google Scholar]
- 3. Parent B, Suard B, Serraj R, Tardieu F (2010) Rice leaf growth and water potential are resilient to evaporative demand and soil water deficit once the effects of root system are neutralized. Plant, Cell & Environment 33: 1256–1267. [DOI] [PubMed] [Google Scholar]
- 4. Peng S, Cassman KG, Virmani SS, Sheehy J, Khush GS (1999) Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential. Crop Science 39: 1552–1559. [Google Scholar]
- 5. Hubbart S, Peng S, Horton P, Chen Y, Murchie EH (2007) Trends in leaf photosynthesis in historical rice varieties developed in the Philippines since 1966. Journal of Experimental Botany 58: 3429–3438. [DOI] [PubMed] [Google Scholar]
- 6. Gu J, Yin X, Stomph TJ, Struik PC (2014) Can exploiting natural genetic variation in leaf photosynthesis contribute to increasing rice productivity? A simulation analysis. Plant, cell & environment 37: 22–34. [DOI] [PubMed] [Google Scholar]
- 7. Centritto M, Lauteri M, Monteverdi MC, Serraj R (2009) Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. Journal of Experimental Botany 60: 2325–2339. [DOI] [PubMed] [Google Scholar]
- 8. Long SP (2014) We need winners in the race to increase photosynthesis in rice, whether from conventional breeding, biotechnology or both. Plant, Cell & Environment 37: 19–21. [DOI] [PubMed] [Google Scholar]
- 9. Gu J, Yin X, Stomph T-J, Wang H, Struik PC (2012) Physiological basis of genetic variation in leaf photosynthesis among rice (Oryza sativa L.) introgression lines under drought and well-watered conditions. Journal of Experimental Botany 63: 5137–5153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Niinemets Ü, Diaz-Espejo A, Flexas J, Galmes J, Warren CR (2009) Role of mesophyll diffusion conductance in constraining potential photosynthetic productivity in the field. Journal of Experimental Botany 60: 2249–2270. [DOI] [PubMed] [Google Scholar]
- 11. Flexas J, Ribas-Carbó M, Diaz-Espejo A, Galmēs J, Medrano H (2008) Mesophyll conductance to CO2: current knowledge and future prospects. Plant, Cell & Environment 31: 602–621. [DOI] [PubMed] [Google Scholar]
- 12. Centritto M, Loreto F, Chartzoulakis K (2003) The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings. Plant, Cell & Environment 26: 585–594. [Google Scholar]
- 13. Pallozzi E, Fortunati A, Marino G, Loreto F, Agati G, et al. (2013) BVOC emission from Populus × canadensis saplings in response to acute UV-A radiation. Physiologia Plantarum 148: 51–61. [DOI] [PubMed] [Google Scholar]
- 14. Hu L, Wang Z, Huang B (2010) Diffusion limitations and metabolic factors associated with inhibition and recovery of photosynthesis from drought stress in a C3 perennial grass species. Physiologia Plantarum 139: 93–106. [DOI] [PubMed] [Google Scholar]
- 15. Scafaro AP, Von Caemmerer S, Evans JR, Atwell BJ (2011) Temperature response of mesophyll conductance in cultivated and wild Oryza species with contrasting mesophyll cell wall thickness. Plant Cell and Environment 34: 1999–2008. [DOI] [PubMed] [Google Scholar]
- 16. Adachi S, Nakae T, Uchida M, Soda K, Takai T, et al. (2013) The mesophyll anatomy enhancing CO2 diffusion is a key trait for improving rice photosynthesis. Journal of Experimental Botany 64: 1061–1072. [DOI] [PubMed] [Google Scholar]
- 17. Manickavelu A, Nadarajan N, Ganesh SK, Gnanamalar RP, Chandra Babu R (2006) Drought tolerance in rice: morphological and molecular genetic consideration. Plant Growth Regulation 50: 121–138. [Google Scholar]
- 18. Huang X-Y, Chao D-Y, Gao J-P, Zhu M-Z, Shi M, et al. (2009) A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes & Development 23: 1805–1817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Hu SP, Zhou Y, Zhang L, Zhu XD, Li L, et al. (2009) Correlation and quantitative trait loci analyses of total chlorophyll content and photosynthetic rate of rice (Oryza sativa) under water stress and well-watered conditions. Journal of Integrative Plant Biology 51: 879–888. [DOI] [PubMed] [Google Scholar]
- 20. Gu J, Yin X, Struik PC, Stomph TJ, Wang H (2012) Using chromosome introgression lines to map quantitative trait loci for photosynthesis parameters in rice (Oryza sativa L.) leaves under drought and well-watered field conditions. Journal of Experimental Botany 63: 455–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Flexas J, Niinemets Ü, Gallé A, Barbour M, Centritto M, et al. (2013) Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynthesis Research 117: 45–59. [DOI] [PubMed] [Google Scholar]
- 22. Farquhar GD, Caemmerer S, Berry JA (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149: 78–90. [DOI] [PubMed] [Google Scholar]
- 23. Giuliani R, Koteyeva N, Voznesenskaya E, Evans MA, Cousins AB, et al. (2013) Coordination of leaf photosynthesis, transpiration, and structural traits in rice and wild relatives (Genus Oryza). Plant Physiology 162: 1632–1651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kaldenhoff R (2012) Mechanisms underlying CO2 diffusion in leaves. Current Opinion in Plant Biology 15: 276–281. [DOI] [PubMed] [Google Scholar]
- 25. Hanba YT, Shibasaka M, Hayashi Y, Hayakawa T, Kasamo K, et al. (2004) Overexpression of the barley aquaporin HvPIP2;1 increases internal CO2 conductance and CO2 assimilation in the leaves of transgenic rice plants. Plant and Cell Physiology 45: 521–529. [DOI] [PubMed] [Google Scholar]
- 26. Tyerman SD, Niemietz CM, Bramley H (2002) Plant aquaporins: multifunctional water and solute channels with expanding roles. Plant, Cell & Environment 25: 173–194. [DOI] [PubMed] [Google Scholar]
- 27. Loreto F, Centritto M (2008) Leaf carbon assimilation in a water-limited world. Plant Biosystems 142: 154–161. [Google Scholar]
- 28. Chaves M, Flexas J, Pinheiro C (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of Botany 103: 551–560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Cornic G (2000) Drought stress inhibits photosynthesis by decreasing stomatal aperture – not by affecting ATP synthesis. Trends in Plant Science 5: 187–188. [Google Scholar]
- 30. Pinheiro C, Chaves MM (2011) Photosynthesis and drought: can we make metabolic connections from available data? Journal of Experimental Botany 62: 869–882. [DOI] [PubMed] [Google Scholar]
- 31. Kamoshita A, Rodriguez R, Yamauchi A, Wade LJ (2004) Genotypic variation in response of rainfed lowland rice to prolonged drought and rewatering. Plant Production Science 7: 406–420. [Google Scholar]
- 32. Boonjung H, Fukai S (1996) Effects of soil water deficit at different growth stages on rice growth and yield under upland conditions. 2. Phenology, biomass production and yield. Field Crops Research 48: 47–55. [Google Scholar]
- 33. Pons TL, Flexas J, von Caemmerer S, Evans JR, Genty B, et al. (2009) Estimating mesophyll conductance to CO2: methodology, potential errors, and recommendations. Journal of Experimental Botany 60: 2217–2234. [DOI] [PubMed] [Google Scholar]
- 34. Gilbert ME, Pou A, Zwieniecki MA, Holbrook NM (2012) On measuring the response of mesophyll conductance to carbon dioxide with the variable J method. Journal of Experimental Botany 63: 413–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Harley PC, Loreto F, Dimarco G, Sharkey TD (1992) Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2 . Plant Physiology 98: 1429–1436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Bois JF, Couchat PH, Lasceve G (1985) Relationships between transpiration and photosynthesis during a water stress. Acta Horticulturae 171: 297–314. [Google Scholar]
- 37. Richter A, Wanek W, Werner RA, Ghashghaie J, Jaeggi M, et al. (2009) Preparation of starch and soluble sugars of plant material for the analysis of carbon isotope composition: a comparison of methods. Rapid Communications in Mass Spectrometry 23: 2476–2488. [DOI] [PubMed] [Google Scholar]
- 38. Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40: 503–537. [Google Scholar]
- 39. Rodeghiero M, Niinemets Ü, Cescatti A (2007) Major diffusion leaks of clamp-on leaf cuvettes still unaccounted: how erroneous are the estimates of Farquhar et al. model parameters? Plant, cell & environment 30: 1006–1022. [DOI] [PubMed] [Google Scholar]
- 40. Loreto F, Harley PC, Dimarco G, Sharkey TD (1992) Estimation of mesophyll conductance to CO2 flux by three different methods. Plant Physiology 98: 1437–1443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Genty B, Briantais J-M, Baker NR (1989) The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta (BBA) - General Subjects 990: 87–92. [Google Scholar]
- 42. Laisk A, Loreto F (1996) Determining photosynthetic parameters from leaf CO2 exchange and chlorophyll fluorescence (ribulose-1, 5-bisphosphate carboxylase/oxygenase specificity factor, dark respiration in the light, excitation distribution between photosystems, alternative electron transport rate, and mesophyll diffusion resistance. Plant Physiology 110: 903–912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Galmes J, Flexas J, Keys AJ, Cifre J, Mitchell RAC, et al. (2005) Rubisco specificity factor tends to be larger in plant species from drier habitats and in species with persistent leaves. Plant Cell and Environment 28: 571–579. [Google Scholar]
- 44. Furbank RT, von Caemmerer S, Sheehy J, Edwards G (2009) C4 rice: a challenge for plant phenomics. Functional Plant Biology 36: 845–856. [DOI] [PubMed] [Google Scholar]
- 45. Pinelli P, Loreto F (2003) 12CO2 emission from different metabolic pathways measured in illuminated and darkened C-3 and C-4 leaves at low, atmospheric and elevated CO2 concentration. Journal of Experimental Botany 54: 1761–1769. [DOI] [PubMed] [Google Scholar]
- 46. Loreto F, Dimarco G, Tricoli D, Sharkey TD (1994) Measurements of mesophyll conductance, photosynthetic electron transport and alternative electron sinks of field grown wheat leaves. Photosynthesis Research 41: 397–403. [DOI] [PubMed] [Google Scholar]
- 47. Lauteri M, Scartazza A, Guido MC, Brugnoli E (1997) Genetic variation in photosynthetic capacity, carbon isotope discrimination and mesophyll conductance in provenances of Castanea sativa adapted to different environments. Functional Ecology 11: 675–683. [Google Scholar]
- 48. Scartazza A, Lauteri M, Guido MC, Brugnoli E (1998) Carbon isotope discrimination in leaf and stem sugars, water-use efficiency and mesophyll conductance during different developmental stages in rice subjected to drought. Australian Journal of Plant Physiology 25: 489–498. [Google Scholar]
- 49. Evans JR, Sharkey TD, Berry JA, Farquhar GD (1986) Carbon isotope discrimination measured concurrently with gas exchange to investigate CO2 diffusion in leaves of higher plants. Australian Journal of Plant Physiology 13: 281–292. [Google Scholar]
- 50. Lloyd J, Syvertsen JP, Kriedemann PE, Farquhar GD (1992) Low conductances for CO2 diffusion from stomata to the sites of carboxylation in leaves of woody species. Plant Cell and Environment 15: 873–899. [Google Scholar]
- 51. von Caemmerer S, Evans JR (1991) Determination of the average partial pressure of CO2 in chloroplasts from leaves of several C3 plants. Australian Journal of Plant Physiology 18: 287–305. [Google Scholar]
- 52. Pantuwan G, Fukai S, Cooper M, Rajatasereekul S, O'Toole JC (2002) Yield response of rice (Oryza sativa L.) genotypes to different types of drought under rainfed lowlands: Part 1. Grain yield and yield components. Field Crops Research 73: 153–168. [Google Scholar]
- 53. Siddique K, Kirby E, Perry M (1989) Ear: stem ratio in old and modern wheat varieties; relationship with improvement in number of grains per ear and yield. Field Crops Research 21: 59–78. [Google Scholar]
- 54. Koyama K, Kikuzawa K (2009) Is whole-plant photosynthetic rate proportional to leaf area? A test of scalings and a logistic equation by leaf demography census. The American Naturalist 173: 640–649. [DOI] [PubMed] [Google Scholar]
- 55. Tomás M, Flexas J, Copolovici L, Galmés J, Hallik L, et al. (2013) Importance of leaf anatomy in determining mesophyll diffusion conductance to CO2 across species: quantitative limitations and scaling up by models. Journal of experimental botany 64: 2269–2281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Zhao B, Kondo M, Maeda M, Ozaki Y, Zhang J (2004) Water-use efficiency and carbon isotope discrimination in two cultivars of upland rice during different developmental stages under three water regimes. Plant and Soil 261: 61–75. [Google Scholar]
- 57. Tipple BJ, Berke MA, Doman CE, Khachaturyan S, Ehleringer JR (2013) Leaf-wax n-alkanes record the plant–water environment at leaf flush. Proceedings of the National Academy of Sciences 110: 2659–2664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Quick W, Chaves M, Wendler R, David M, Rodrigues M, et al. (1992) The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions. Plant, Cell and Environment 15: 25–35. [Google Scholar]
- 59. Yang J, Zhang J, Wang Z, Zhu Q (2001) Activities of starch hydrolytic enzymes and sucrose-phosphate synthase in the stems of rice subjected to water stress during grain filling. Journal of Experimental Botany 52: 2169–2179. [DOI] [PubMed] [Google Scholar]
- 60. Scheidegger Y, Saurer M, Bahn M, Siegwolf R (2000) Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model. Oecologia 125: 350–357. [DOI] [PubMed] [Google Scholar]
- 61. Schmitt MR, Edwards GE (1981) Photosynthetic capacity and nitrogen use efficiency of maize, wheat, and rice - a comparison between C-3 and C-4 Photosynthesis. Journal of Experimental Botany 32: 459–466. [Google Scholar]
- 62. Kiran TV, Rao YV, Subrahmanyam D, Rani NS, Bhadana VP, et al. (2013) Variation in leaf photosynthetic characteristics in wild rice species. Photosynthetica 51: 350–358. [Google Scholar]
- 63. Terashima I (1992) Anatomy of non-uniform leaf photosynthesis. Photosynthesis research 31: 195–212. [DOI] [PubMed] [Google Scholar]
- 64. Mott KA, Buckley TN (2000) Patchy stomatal conductance: emergent collective behaviour of stomata. Trends in Plant Science 5: 258–262. [DOI] [PubMed] [Google Scholar]
- 65. Sun P, Wahbi S, Tsonev T, Haworth M, Liu S, et al. (2014) On the use of leaf spectral indices to assess water status and photosynthetic limitations in Olea europaea L. during water-stress and recovery. Plos One 9: e105165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Tsonev T, Wahbi S, Sun P, Sorrentino G, Centritto M (2014) Gas exchange, water relations and their relationships with photochemical reflectance index in Quercus ilex plants during water stress and recovery. International Journal of Agriculture and Biology 16: 335–341. [Google Scholar]
- 67.Warren CR. Stand aside stomata, another actor deserves centre stage: the forgotten role of the internal conductance to CO2 transfer; 2007 Jul 22–27; Glasgow, Scotland. Oxford University Press. pp.1475–1487. [DOI] [PubMed] [Google Scholar]
- 68.Centritto M, Tognetti R, Leitgeb E, Střelcová K, Cohen S (2011) Above ground processes - anticipating climate change influences. In: Bredemeier M, Cohen S, Godbold DL, Lode E, Pichler V et al., editors. Forest Management and the Water Cycle: An Ecosystem-Based Approach. London: Springer. pp.31–64. [Google Scholar]
- 69. Haworth M, Elliott-Kingston C, McElwain J (2013) Co-ordination of physiological and morphological responses of stomata to elevated [CO2] in vascular plants. Oecologia 171: 71–82. [DOI] [PubMed] [Google Scholar]
- 70. Haworth M, Elliott-Kingston C, McElwain JC (2011) Stomatal control as a driver of plant evolution. Journal of Experimental Botany 62: 2419–2423. [DOI] [PubMed] [Google Scholar]
- 71. Haworth M, Gallagher A, Elliott-Kingston C, Raschi A, Marandola D, et al. (2010) Stomatal index responses of Agrostis canina to carbon dioxide and sulphur dioxide: implications for palaeo-[CO2] using the stomatal proxy. New Phytologist 188: 845–855. [DOI] [PubMed] [Google Scholar]
- 72. Kubiskem ME, Abrams MD (1993) Stomatal and nonstomatal limitations of photosynthesis in 19 temperate tree species on contrasting sites during wet and dry years. Plant, Cell and Environment 16: 1123–1129. [Google Scholar]
- 73. Venuprasad R, Lafitte HR, Atlin GN (2007) Response to direct selection for grain yield under drought stress in rice. Crop Sci 47: 285–293. [Google Scholar]
- 74. Karaba A, Dixit S, Greco R, Aharoni A, Trijatmiko KR, et al. (2007) Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt tolerance gene. Proceedings of the National Academy of Sciences of the United States of America 104: 15270–15275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Lanceras JC, Pantuwan G, Jongdee B, Toojinda T (2004) Quantitative trait loci associated with drought tolerance at reproductive stage in rice. Plant Physiology 135: 384–399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Xu Y, This D, Pausch RC, Vonhof WM, Coburn JR, et al. (2009) Leaf-level water use efficiency determined by carbon isotope discrimination in rice seedlings: genetic variation associated with population structure and QTL mapping. Theoretical and Applied Genetics 118: 1065–1081. [DOI] [PubMed] [Google Scholar]
- 77.Wassmann R, Jagadish SVK, Heuer S, Ismail A, Redona E, et al. (2009) Climate change affecting rice production: the physiological and agronomic basis for possible adaptation strategies. In: Sparks DL, editor. Advances in Agronomy, Vol 101. pp.59–122.
- 78. Blum A (2005) Drought resistance, water-use efficiency, and yield potential–are they compatible, dissonant, or mutually exclusive? Australian Journal of Agricultural Research 56: 1159–1168. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Dark respiration ( R d) values of seven Oryza sativa varieties under irrigated (white) and rain-fed (grey) conditions used in the calculation of g m using the variable J method.
(PDF)
Electron transport rate (ETR) values of seven Oryza sativa varieties under irrigated (white) and rain-fed (grey) conditions used in the calculation of gm using the variable J method.
(PDF)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.