

Abstract
The immature neonatal intestinal immune system hyperreacts to newly colonizing unfamiliar bacteria. The hypothesis that human milk oligosaccharides from colostrum (cHMOS) can directly modulate the signaling pathways of the immature mucosa was tested. Modulation of cytokine immune signaling by HMOS was measured ex vivo in intact immature (fetal) human intestinal mucosa. From the genes whose transcription was modulated by colostrum HMOS (cHMOS), Ingenuity Pathway Analysis identified networks controlling immune cell communication, intestinal mucosal immune system differentiation, and homeostasis. cHMOS attenuate pathogen-associated molecular pattern (PAMP)-stimulated acute phase inflammatory cytokine protein levels (IL-8, IL-6, MCP-1/2, IL-1β), while elevating cytokines involved in tissue repair and homeostasis. 3’-, 4-, and 6’-galactosyllactoses of cHMOS account for specific immunomodulation of PIC-induced IL-8 levels. cHMOS attenuate mucosal responses to surface inflammatory stimuli during early development, while enhancing signals that support maturation of the intestinal mucosal immune system.
INTRODUCTION
The intestinal mucosa defends against both pathogen infection (exogenous novel epitopes) and, conversely, autoimmunity (endogenous epitopes), in part by discriminating “self” from “non-self” antigens1, 2. The antigenic composition of the intestinal contents shifts rapidly after birth with the introduction of a plethora of novel antigens as dietary components, and as a consequence of early colonization by strains and species of microbiota novel to the newborn gut2. This coincides with a period during which the immature neonatal intestinal mucosal immune system is hyperinflammatory3. Subsequent immune maturation is required both for active responses to foreign antigens, and for proper control and regulation of such responses. Impaired immunological development due to preterm delivery, injury at the mucosal surface, pathogenic infection, and food allergy may disturb the ontogeny of homeostatic control of inflammatory processes, contributing to conditions such as necrotizing enterocolitis (NEC)4, 5, pediatric inflammatory bowel diseases (IBD) 6, Th2 disorders, and atopic dermatitis6.
Fortunately, human milk quenches inflammatory processes7, 8 and minimizes the incidence of many immune-based disorders9. In early postnatal developing gut, milk enhances the signals that facilitate appropriate immune responses and antigenic memory9. Moreover, human milk contains immune modulatory components and protects against development of IBD10, 11, 12. Human milk oligosaccharides (HMOS) are complex glycans containing a lactose moiety at the reducing end. At 5-15 mg/mL, HMOS are collectively the third most abundant solid component of human milk13, 14. Traditionally, up to 200 known individual HMOS are subdivided into neutral and acidic oligosaccharides13, 14. Some possess anti-inflammatory functions, including reduced leukocyte adhesion15, and some bind specifically to dendritic cells through the lectin receptor DC-SIGN (Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Non-integrin)16. The human milk oligosaccharide disialyllacto-N-tetraose reduces NEC-like inflammation in neonatal rats17. Some HMOS in colostrum, cHMOS, are present at higher concentration and have a different pattern than HMOS of mature milk (mHMOS)18,19. Some oligosaccharides observed in colostrum decline to undetectable levels as lactation proceeds20. The biologic activities of these colostrum-specific oligosaccharides had not been defined. Immature human intestine ex vivo was deemed the most relevant model for investigating effects of colostrum oligosaccharides on maturation- and inflammation-related signaling of immature intestinal mucosa of neonates.
The hypothesis tested was that HMOS from colostrum directly modulate mucosal signaling in immature human intestine. This was investigated in human fetal intestine explants. Changes induced by cHMOS were determined in basal and PAMP (or IL-1β)-stimulated intestinal mucosa. The predominant TLRs expressed in intestinal mucosa are TLR3 and TLR5 21, whose PAMPs are double-stranded RNA (polyinosinic:polycytodylic acid, PIC) and flagellin (FLA), respectively. Stimulation of the IL-1β receptor mediates a variety of signaling pathways associated with innate immunity and host responses to microbes. Signaling was measured through RT-PCR array analysis of the cytokinerelated transcriptome, and shifts in functional patterns of expression were confirmation by antibody arrays of the corresponding proteome, and ELISA of IL-8 levels, a prototypic inflammatory cytokine of human intestinal mucosa. Immunomodulation by individual HMOS candidates was determined in cultured human intestinal epithelial cells.
RESULTS
Modulation of inflammatory cytokines and receptors in basal epithelium by cHMOS
The initial condition of cultured fetal intestinal tissues was evaluated by IL-8 concentrations in the media. As shown in Supplemental Fig. 1, IL-8 concentrations were below 100±10 pg/mL in successfully cultured healthy tissue samples under quiescent conditions. This concentration represented basal expression of IL-8. When these intestinal samples were challenged with PAMPs (FLA, PIC) or the proinflammatory mediator IL-1β, secretion of IL-8 increased (Supplemental Fig. 1). Only intestinal samples with low basal IL-8 expression retain the ability to respond to exogenous proinflammatory stimuli, and these quiescent intestinal tissues were used in these experiments.
cHMOS, but not mHMOS (P = 0.036), directly modulated immunological signaling in the immature intestine, as measured on an 84-gene RT2 PCR array by probes against chemokine and chemokine receptor genes. The screening criteria described in the Methods section identified 45 genes whose transcription was up-regulated by cHMOS and 11 genes whose transcription was down-regulated by cHMOS (Table 1). The ten genes whose transcription was most up-regulated and down-regulated by the presence of cHMOS are listed in Fig. 1. Those genes whose transcription was highly up-regulated include several chemokines, such as CCL21, CCL11, CCL24, CXCL2, CXCL10 and CCL13. IL-10, usually considered an anti-inflammatory cytokine, also increased. Another significantly up-regulated gene transcription was CARD18 (Caspase recruitment domain-containing protein, also called Caspase-1 inhibitor Iceberg), which inhibits generation of IL-1β by interacting with caspase-1 and prevents it from forming a complex with RIP222. Among the genes whose transcription was down-regulated by cHMOS , BCL6 and IL-5 were the most highly suppressed, followed by chemokine receptors CCR1, CXCR2 and CCR2, and chemokines CCL1, CCL19, CCL15, and CCL17.
TABLE 1.
Summary of HMOS-regulated inflammatory genes in human fetal intestine
| Chemokines and receptors | Cytokines and receptors | Other genes involved in inflammatory responses | |||
|---|---|---|---|---|---|
|
| |||||
| Symbol | Fold Change | Symbol | Fold Change | Symbol | Fold Change |
| CCL2 (MCP-1) | 8.39 | IL1A | 13.4 | CARD18 | 35.46 |
| CCL3(MIP-1α) | 4.24 | IL1B | 4.71 | CRP | 17.52 |
| CCL11(Eotaxin) | 72.74 | IL8 | 12.03 | MIF | 13.19 |
| CCL13 | 16.62 | IL10 | 29.47 | LTB | 10.23 |
| CCL16 | 5.20 | IL13 | 13.47 | C4A | 6.11 |
| CCL20 (MIP-3α) | 5.28 | IL17C | 9.97 | IFNA2 | 4.77 |
| CCL21(MIP-2) | 208.61 | IL36G | 4.75 | AIMP1 | 3.96 |
| CCL23 | 3.81 | IL37 | 14.40 | CEBPB | 3.71 |
| CCL24(Eotaxin-2) | 41.97 | IL1R1 | 5.09 | BCL6 | 0.02 |
| CCL25 | 6.56 | IL1RN | 3.98 | ||
| CXCL2 | 33.31 | IL5RA | 4.93 | ||
| CXCL3 | 3.8 | IL10RA | 12.1 | ||
| CXCL6 | 8.04 | IL10RB | 15.61 | ||
| CXCL9 | 5.07 | IL13RA1 | 6.73 | ||
| CXCL10(IP-10) | 21.58 | IL36RN | 4.17 | ||
| CXCL11 | 13.13 | IL5 | 0.22 | ||
| CXCL13(MCP-4) | 13.5 | IL9R | 0.34 | ||
| CXCL14 | 7.4 | ||||
| CCR4 | 3.56 | ||||
| CCR6 | 16.46 | ||||
| CCR8 | 4.22 | ||||
| CX3CR1 | 6.52 | ||||
| CCL1(I-309) | 0.38 | ||||
| CCL4(MIP-1β) | 0.78 | ||||
| CCL15(MIP-1δ) | 0.57 | ||||
| CCL17 | 0.66 | ||||
| CCL19 | 0.43 | ||||
| CCR1 | 0.37 | ||||
| CCR2 | 0.77 | ||||
| CXCR2 | 0.58 | ||||
Data represent the fold changes (>3.5 or <0.8; P<0.05) of messenger RNA expression of cytokine, inflammatory regulator, cytokine receptor, chemokine and chemokine receptor genes in human fetal intestine in response to 2 d of HMOS exposure. Significance of differences [P] was measured by two-way RM ANOVA (Graph Pad Software, Inc. San Diego, CA). Abbreviations: IFN, interferon; IL, interleukin; CCL, chemokine (C-C motif) ligand; CXCL, chemokine (C-X-C motif); CCR, chemokine (C-C motif) receptor; CX3CR, chemokine (C-X3-C motif) receptor; CRP, C-reactive protein; MIF, macrophage migration inhibitory factor; LTB, lymphotoxin beta; C4A, Complement component 4A; AIMP, aminoacyl tRNA synthetase complex-interacting multifunctional protein; CEBPB, CCAAT/enhancer binding protein; BCL6, B-cell CLL/lymphoma 6.
Figure 1. Genes whose transcription are most modulated by cHMOS.
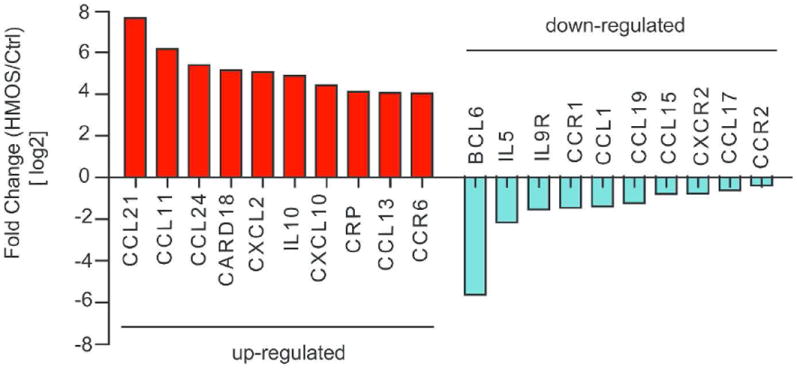
Red bars, ten genes whose transcription is most enhanced by cHMOS; blue bars, ten genes whose transcription is most attenuated by cHMOS.
Analysis of cHMOS-modulated gene expression in basal intestinal mucosa
cHMOS-induced changes in gene transcription were clustered computationally by Ingenuity Pathway Analysis (IPA) according to known canonical pathways and biofunctional networks. This program analyzes functional relationships among cHMOS-modulated genes within known signaling pathways. The discreet canonical pathways most strongly modified by cHMOS (Fig. 2) include those for T helper (Th) cell differentiation, immune cell communication, regulation of innate immunity, communication between innate and adaptive immune cells, and dendritic cell maturation.
Figure 2. Canonical pathways whose genes are most strongly modulated by cHMOS.
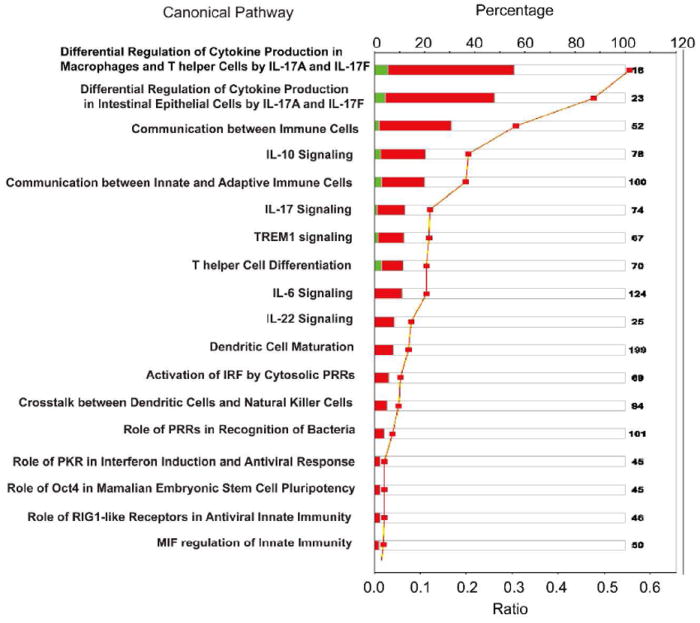
Canonical pathways whose transcription is influenced by cHMOS-treatment were identified by IPA analysis and rank ordered by the magnitude of their ratios. The names of canonical pathways are presented on the y-axis. Red bars, genes whose transcription was enhanced by cHMOS treatment; green bars, genes whose transcription was attenuated by cHMOS. Orange squares represented ratios calculated from changes in gene expression from Table 1.
IPA also identified four higher order functional networks composed of specific biological pathways of cytokine and chemokine genes whose transcription was strongly modified by cHMOS (Fig. 3). A pathway involved in immune cell trafficking is depicted in Fig. 3A, in which ERK1/2 (extracellular signal-regulated kinase) is the central signaling molecule. This network includes several genes whose transription is strongly influenced by cHMOS, including upregulation of CCL21, CCL11, and CRP, and attenuation of CCL19 expression. A network modulating development and function of specific cells of the hematological system, as well as antigen presentation, is shown in Fig. 3B. In this network, the genes whose transcrption is stimulated by cHMOS include CCL24, CXCL12, CCR6, and those whose transcription is attenuated by cHMOS include CCR1, CXCR2, CCL1 and CCL17. For these two networks, cHMOS treatment is expected to have specific effects on various constituent pathways.
Figure 3. Four functional networks whose gene expression was most modulated by cHMOS.
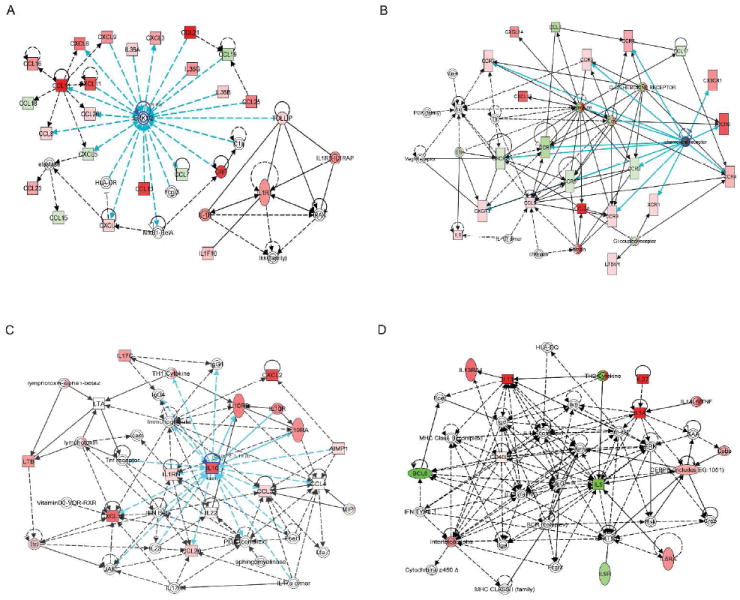
(A) Network 1 signaling pathways for “cell/cell interaction, cellular movement, and immune cell trafficking”. (B) Expression products of the genes of Network 2 comprise signaling pathways for “antigen presentation, hematological system development and function”. Solid lines, direct interaction; dashed lines, indirect interaction. Genes whose expression is enhanced by cHMOS are red; those whose expression was attenuated are shown in green. (C) Network 3 genes for cytokines promoting Th1 activity and differentiation that were stimulated by cHMOS. (D) Network 4 genes for cytokines promoting Th2 activity and differentiation were suppressed by cHMOS. Solid lines, direct interaction; dashed lines, indirect interaction. Red indicates gene transcription promoted by cHMOS; green indicates gene transcription suppressed by cHMOS.
A network that regulates inflammatory response is illustrated in Fig. 3C, in which cHMOS stimulate expression of CXCL2, CXCL10, and IL-10. Calculated global cytokine change scores in this network deduce a potential increase in Th1 (T helper cell 1) cytokines. The genes upregulated by cHMOS, which lead to a predominance of cytokines supporting a Th1 response, are labeled red.
In the network of genes that control signaling pathways for development and function of specific cell types during hematological system development (Fig. 3D), cHMOS attenuated transcription of BCL6, IL-5, and IL9R. The deduced cytokine expression changes suggest a global change of cytokines that would attenuate Th2 (T helper cell 2) cytokine functions. Genes whose expression is attenuated by cHMOS, and whose decrease would suppress cytokines supporting Th2 function, are labeled green.
cHMOS-modulated cytokine expression in basal immature intestinal mucosa
The above analyses of the functional significance of cHMOS modulation of signaling systems in quiescent basal mucosa were deduced from differences in gene transcription. Changes in expression of inflammatory cytokine protein levels induced by cHMOS in immature intestine were measured to scrutinize the conclusions that had been based on RNA levels. The cytokines were measured using antibody membrane arrays that detect 40 different cytokines simultaneously. In Supplemental Fig. 2, the position map of cytokines and the corresponding intensity of binding are shown. The change in expression of signaling molecules induced by cHMOS in basal unstimulated intestinal mucosa is summarized in Fig. 4. cHMOS suppressed translation of IL-17, IL-13, IL-4, IL-2, IL-11, G-CSF, TNF-β, TGF-β1, GM-CSF, and eotaxin. Among these are typical Th2 polarizing cytokines IL-13, IL-4, IL-1β, and IL-6. Likewise, the expression of Th17 polarizing cytokines IL-17, TNF-β, TGF-β1, and IL-4 was reduced by cHMOS. In contrast, translation of Th1 polarizing cytokines MCP-1 (CCL-2) and IL12-p70 was stimulated by cHMOS. The overall functional pattern of cHMOS modulation of signaling molecule translation is consistent with the overall functional pattern of the changes in transcription observed in the RT-PCR arrays. The concordance between the increase in MCP-1 protein and upregulation of its mRNA (CCL2) transcription is striking. However, decreased cytokine protein levels, such as IL-10, did not vary according to changes in their mRNA transcription following cHMOS treatment; this suggests that these cytokine protein levels may be regulated by factors other than gene transcription, such as the presence of miRNA, rates of degradation, or posttranslational modifications that are common in regulation of the rapidly changing levels of signal transduction molecules23 . That notwithstanding, the imputed global functional consequences of the protein and mRNA changes in response to cHMOS are congruent. Both suggest that cHMOS modulate differentiation of T helper cells toward a balanced Th1 expansion in unstimulated basal immature intestine.
Figure 4. Modulation of cytokine translation by cHMOS in quiescent unstimulated mucosa.
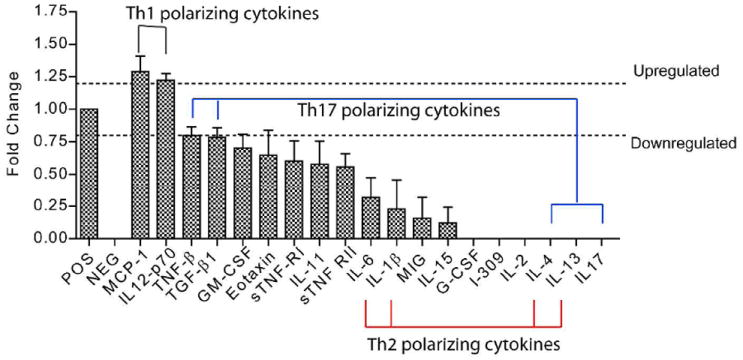
Cytokine translation was assessed by antibody membrane array. Cytokine expression in quiescent mucosa was significantly modulated by cHMOS, with changes of >1.2 fold considered up-regulated, and changes of <0.8 considered down-regulated (comparison with basal levels, P<0.05).
cHMOS-modulated cytokine expression after proinflammatory stimulation of immature intestinal mucosa
To query whether cHMOS influence innate immune responses in inflamed immature intestine, the tissues were treated with three distinct PAMP-related inflammatory stimuli, FLA, PIC, and IL-1β at concentrations previously found to be effective inducers of inflammatory signaling in this system24. Fig. 5 illustrates that cHMOS significantly modulated a wide spectrum of cytokine protein expression induced by PAMP or IL-1β stimulation. Several cytokines were up-regulated by cHMOS. In tissues challenged with FLA, cHMOS upregulated MIP-1δ (CCL15) and ICAM-1 expression (Fig. 5A). Each of these promotes the uptake of bacterial pathogens by recruiting macrophages and neutrophils, and lead to bacteria clearance25, 26. In intestine challenged with PIC, cHMOS upregulated TIMP-2 and PDGF-BB (Fig. 5C), both of which enhance the regulation of epithelial cell functions and promote mucosal repair27. In intestine stimulated by IL-1β, cHMOS upregulated expression of MIP-1δ (CCL15), MIP-1β (CCL4), and TIMP-2 (Fig. 5B). TIMP-2 inhibits matrix metalloproteinase, and is critical in maintaining tissue homeostasis28.
Figure 5. Modulation of cytokine translation by cHMOS in PAMP-stimulated mucosa.
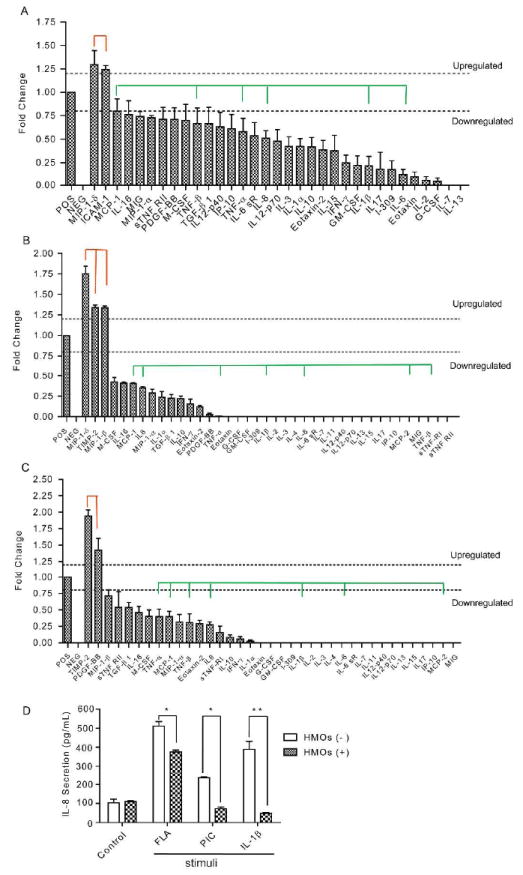
(A) cHMOS modulated the FLA-induced changes in cytokine profile of immature intestinal mucosa. (B) cHMOS modulated the IL-1β-induced changes in cytokine profile of immature intestinal mucosa. (C) cHMOS modulated the PIC-induced changes in cytokine profile of immature intestinal mucosa. Green lines indicate acute inflammatory cytokines suppressed by cHMOS. Red lines indicate gene translation promoted by cHMOS. Data were duplicates from three independent experiments (n=6, P<0.05). (D) cHMOS treatment attenuated the stimulation of IL-8 cytokine secretion induced by these three major PAMP related proinflammatory molecules. (*P<0.05; ** P<0.01).
Although modulation of cytokine expression by cHMOS varied in tissues stimulated by different PAMP-related stimuli, those intracellular cytokines mediating acute phase inflammation, including IL-8, IL-6, IL-1β, TNF (-α or -β), and MCP (-1 or -2), were suppressed by cHMOS in all cases (Fig. 5A-C). This ability of cHMOS to attenuate PAMP-induced inflammation was corroborated by measuring IL-8 secretion by ELISA in the tissue supernatant. IL-8 is a chemokine produced by macrophages and epithelial cells of the intestinal mucosa. As a prototypic marker of inflammation, IL-8 recruits neutrophils to sites of inflammation in response to infection, trauma, ischemia, and cancer29. The basal IL-8 concentration is not significantly changed by cHMOS. PAMP-related stimuli increase IL-8 secretion in immature intestine. These increased IL-8 secretions were abrogated by the presence of cHMOS (Fig. 5D). In FLA (100 ng/mL) stimulated tissue, cHMOS treatment significantly reduced IL-8 expression by approximately 25% (P<0.05). In PIC (10 ng/mL) and IL-1β (10 ng/mL) stimulated tissues, cHMOS suppressed IL-8 expression dramatically, from concentrations of 235 pg/mL (P<0.05) and 384 pg/mL (P<0.001), respectively, to basal levels of expression (~100 pg/mL). Attenuation of secreted IL-8 by ELISA was consistent with the results of the cytokine membrane array, demonstrating that the cHMOS fraction is anti-inflammatory, protecting immature infant intestinal mucosa from inflammation triggered by various PAMP or PAMP-related stimuli.
LC-MS profile of active HMOS
The quantities and concentrations of individual oligosaccharides vary between individuals and over the course of lactation14, 19. HMOS of different mothers can vary in activity19. In preliminary experiments, different milk preparations varied greatly in their ability to quench inflammation (manifested as elevated IL-8 levels). Therefore the differences between early milk and mature milk oligosaccharides were studied systematically as part of the experimental paradigm. Of the eight HMOS preparations in this study, only the two that had been prepared from colostrum (cHMOS) demonstrated the robust and consistent immune modulatory activities in fetal intestinal tissue shown above (P = 0.036). The HMOS preparations, six from mature milks, two from colostrum, were each prepared from milks pooled from a discrete population of 20-50 mothers. The individual oligosaccharides of each were resolved and quantified by LC-MS (high-performance liquid chromatography with tandem mass spectrometry). All eight preparations contained a typical pattern of HMOS of pooled milk, including a preponderance of 2’-fucosyllactose (2’-FL), lacto-N-fucopentaose 1 (LNF1, or LNFP1), lacto-N-tetraose (LNT) and lactodifucotetraose (LDFT) (Fig. 6A). However, the two cHMOS preparations contained higher concentrations of several larger oligosaccharides whose structures terminate with complex fucosylated and/or sialylated moieties, and two distinct peaks of galactosyloligosaccharides previously reported only in colostrum, 6’-galactosyllactose and 3’-galactosyllactose30 (Fig. 6A). The variation in amounts of individual oligosaccharides among the eight preparations is shown in Fig. 6B.
Figure 6. Characterization of active HMOS profile by LC-MS.
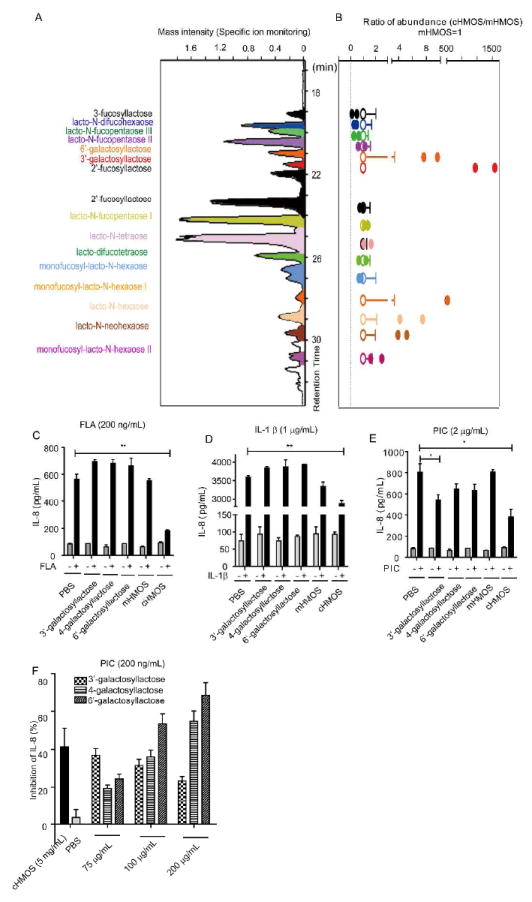
(A) Representative profile of HMOS from colostrum by LC-MS. Major peaks were identified by comparing retention times and mass spectra with authentic isolated standards. (B) Variation in relative amounts of the major oligosaccharides isolated from mature human milk (open circles) and colostrum (filled circles). The two colostrum preparations were highly active immunomodulators, as described above, while the mature milk HMOS did not display consistent robust activity. (C) Inhibition by galactosyllactoses relative to HMOS from colostrum and mature milk on IL-8 secretion in FLA (200 ng/mL) stimulated immature intestinal epithelial cells. (D) Inhibition by galactosyllactoses relative to HMOS from colostrum and mature milk on IL-8 secretion in IL-1β (1 μg/mL) stimulated immature intestinal epithelial cells. (E) Inhibition by galactosyllactoses relative to HMOS from colostrum and mature milk on IL-8 secretion in PIC (2 μg/mL) stimulated immature intestinal epithelial cells. (F) Dose-related inhibition by galactosyllactoses (75-200 μg/mL) relative to cHMOS on IL-8 secretion in PIC (200 μg/mL) stimulated immature intestinal epithelial cells.
The higher concentration of galactosyllactoses in cHMOS relative to mHMOS, when considered together with immunomodulation by only cHMOS, suggested galactosyllactoses as candidate molecules for immunomodulation. The 3’- and 6’-galactosyllactoses that were detected in the active cHMOS fractions, as well as the 4-galactosyllactose reported to be in human colostrum, were tested at a concentration of 200 μg/mL for immunomodulation and compared to cHMOS at 5 mg/ml. These concentrations are within the physiologic range, when considering the highly variable expression of HMOS in colostrum. The activities were tested in H4 cells, a line of immature intestinal epithelial cells that provide an expedient and relevant model for screening these activities, as 80% of the cells in the 18-22 week old intestinal mucosa are intestinal epithelial cells31. Exposure of the epithelial cells to proinflammatory PAMPs (FLA, PIC) or IL-1β at levels previously found to be effective in cell culture (unpublished data) induced pronounced increases in IL-8 levels. None of the three galactosyllactoses attenuated the IL-8 inflammatory responses to FLA or IL-1β (Fig. 6C &D). In contrast, the IL-8 response to 2 μg/mL PIC was significantly attenuated specifically by 3’-galactosyllactose, but not 4- or 6’-galactosyllactose relative to the PBS control group (Fig. 6E). However, if the cells stimulated by 2 μg/mL PIC and treated with mHMOS (which exhibited no inhibition) are combined with the PBS treated positive control cells also stimulated by 2 μg/mL PIC, then relative to this expanded control group, 3’-, 4- and 6’-galactosyllactose inhibition was significant at P = 0.001, 0.03, and 0.03, respectively. When the stimulation was reduced to 200 ng/mL PIC (Fig. 6F), the induction of IL-8 was inhibited by each of the three galactosyloligosaccharides at 75, 100, and 200 μg/mL. Thus, galactosyllactose replicates the ability of the native cHMOS fraction to attenuate the IL-8 inflammatory response to PIC at physiologically meaningful concentrations in vitro, but the remaining activities of the cHMOS fraction cannot be accounted for by the galactosyllactoses tested, indicating that additional oligosaccharide components of cHMOS remain candidates for mediating the residual immunomodulation exhibited by cHMOS.
DISCUSSION
The intestine is the largest immune organ in the body, and is proximate to a diverse pool of immune effector cells. In comparison to intestinal epithelial cells of the mature mucosa, neonatal intestinal epithelial cells overexpress innate inflammatory genes such as NFκB, MyD88, TLR2, TLR4, and TRAF, with scant expression of negative feedback regulator genes32. Accordingly, immature intestinal mucosa is prone to exaggerated responses to proinflammatory stimuli, increasing the risk of inflammatory diseases of the intestine in the neonate32.
Immature infant intestinal mucosa also contain fewer total T cells than mature mucosa33, and those present are skewed toward a Th2 bias. During pregnancy, Th2 responses prevent adverse immunological reactions between the mother and her fetus34 (Fig. 7A). This residual Th2 bias leaves neonatal intestinal mucosa more vulnerable to bacterial infection and food allergy35, and the intestinal immune system must mature to meet immune challenges of adult life.
Figure 7. Hypothetical model of colostrum HMOS as immune modulators in human fetal intestine.
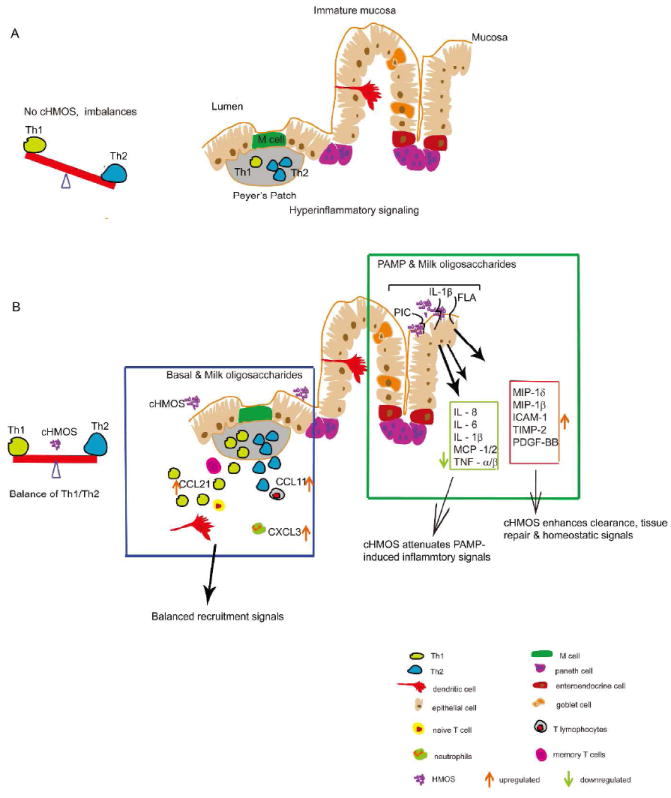
(A) The neonatal intestinal immune system is immature at birth with an imbalance between Th1-Th2 and fewer T cells. (B) Multiple effects of cHMOS on immune modulation in immature intestine.
The hypersensitivity to inflammatory stimuli and the skew towards Th2 are major contributors toward the extraordinary reactiveness of the neonatal, and especially premature, intestinal mucosa to external stimuli. Breastfeeding and human milk per se are known to counter adverse outcomes associated with inappropriately high responses to mucosal stimulation. For example, feeding human milk to preterm infants lowers incidence and severity of infectious and autoimmune diseases12, 36. Many human milk components are biologically active, including anti-inflammatory cytokines, chemokines, hormones, and growth factors37, 38. The natural mixture of human milk oligosaccharides alters cytokine production of T-cells from cord blood39. Plant-derived oligosaccharides designed to mimic those of human milk increase the Th1 response of Th2-skewed populations39. Thus, human milk components, especially HMOS, are putative external agents that induce tolerance, protecting infants from allergy development, while supporting maturation of the infant’s immune system7, 40, 41. The studies herein directly tested the ability of cHMOS to modulate signaling in human immature intestinal mucosa ex vivo and identified signaling pathways that provide potential mechanisms for such activity.
Development of lymphoid tissues, including differentiation of T lymphocytes, is a complex process and is regulated by diverse cytokine signals42. In the ex vivo human immature intestinal mucosa, cHMOS modulated expression of a wide range of cytokines at both transcription (mRNA) and translation (protein) levels (Fig. 1, Table 1, Fig. 4). To interpret these changes in the context of discrete immunological signaling pathways, these changes were clustered into known canonical pathways (recognized pathways from the literature) and biofunctional networks (imputed de novo from the data) by the Ingenuity Pathway Analysis (IPA) software system. This analysis of the data, discussed below, strongly suggests that in basal, unstimulated immature intestinal mucosa (a model for early life, especially for the ELBW neonate), cHMOS modulate immune cell trafficking, maturation, and balance.
cHMOS modulated intrinsic expression of inflammatory markers associated with cell trafficking. In basal, unstimulated mucosa, cHMOS enhanced expression of cytokines associated with recruitment of immune cells, including CCL21, CCL11, CCL13, and CXCL3 (Table 1). CCL21 recruits CCR7-bearing naïve T cells, natural killer cells, memory T cells, and dendritic cells43. CCL11 recruits lymphocytes bearing CCR344. CCL13 (MCP-4) participates in recruitment of monocytes, eosinophils, T lymphocytes, and basophils45. CXCL3 (MIP-2β, macrophage inflammation protein 2β) attracts neutrophils46. The common downstream molecule of these genes is ERK1/2 (extracellular signal-regulated kinase, mitogen-activated protein kinase), whose extracellular signal-regulated kinase activity is central to signal pathways mediating immune cell trafficking47 (Fig. 3C).
cHMOS also modulate signaling pathways related to maturation of lymphoid tissue (Fig. 2). Genes whose expression was significantly changed by cHMOS included signaling molecules for key canonical pathways of immune maturation, including T helper cell differentiation, dendritic cell maturation, and immune cell intercommunication, especially communication between innate and adaptive immune cells. As in the trafficking pathway above, again ERK1/2 is a signaling molecule identified by IPA analysis (Fig. 3A) as one of the most strongly affected by cHMOS. However, in this pathway its mitogen-activated protein kinase activity plays a central role in promoting cellular proliferation48. This suggests that cHMOS stimulates ERK1/2 pathway signaling cytokines that promote intestinal immune system development (Fig. 7B) through rapid lymphoid tissue expansion49.
In this basal, unstimulated mucosa tissue, cHMOS also influenced cytokine and chemokine networks that regulate Th1/Th2 lymphocyte balance (Fig. 7B). Th1 cells drive the type-1 pathway (“cellular immunity”) to fight viruses and other intracellular pathogens and to expunge cancerous cells50. Th2 cells drive the type-2 pathway (“humoral immunity”) to upregulate antibody production to fight extracellular organisms51. However, Th1 secrete cytokines that inhibit Th2 proliferation while Th2 secrete cytokines that inhibit Th1 cell proliferation51, a reciprocal inhibition that is essential for immune homeostasis. Among the cHMOS-up-regulated genes, CCL21 recruits CCR7-bearing Th1 cells52, while IL-36 signaling enhances Th1 proliferation and Th1 polarization of naive CD4+ T cells53. Although cHMOS upregulate CCL11, which induces recruitment of CCR3-bearing Th2 cells44, it attenuates BCL-6 and IL-5, regulators of differentiation of germinal center B cells54, 55, suppressing differentiation of B cells. Downstream, cHMOS treatment stimulates Th1 cytokine production while suppressing Th2 cytokine production (Fig. 3C-D). These cHMOS-related changes in functional patterns of mRNA in quiescent mucosal tissues were confirmed by cytokine (protein) membrane array. Archetypal Th2 and Th17 polarizing cytokines are suppressed by cHMOS while Th1 polarizing cytokines are up-regulated (Fig. 4). The improved balance between Th1 and Th2 fostered by cHMOS may enhance defense against pathogen infection and attenuate susceptibility to food allergy and autoimmune diseases in neonatal intestinal mucosa.
Profuse expression of innate inflammatory genes and sparse expression of negative feedback regulator genes32 underlies the bias toward production of pro-inflammatory cytokines, including IL-6 and IL-8, by basal unstimulated immature mucosa33. IL-6 and IL-8 are acute-phase mediators of the innate immune response34, 56 in recruiting neutrophils. Neutrophils are the most abundant leukocyte in humans and kill microorganisms in an immediate, non-specific manner57. IL-6 combined with IL-23 promotes Th17 responses that enhance epithelial expression of antimicrobial peptides58 and Th17 cells direct neutrophil recruitment into sites of bacterial and fungal infection59. However, overwhelming expression of acute phase cytokines produces excessive neutrophil recruitment, resulting in tissue destruction and inflammatory tissue damage57. Moreover, elevated Th17 responses are associated with the pathogenesis of several autoimmune and inflammatory diseases, including Crohn’s disease60. cHMOS curbed excessive expression of Th17-polarizing cytokines including TNF-β, TGF-β1, IL-4, and IL-17 (Fig. 4). cHMOS also attenuated expression of acute phase inflammatory cytokines, including IL-6 and IL-1β. IL-6 controls Treg (regulatory T cell)/Th17 balance61 and negatively regulates FoxP3+ Treg differentiation. FoxP3+ Tregs attenuate experimental necrotizing enterocolitis62 and its numbers are lower in NEC patients than in normal control groups63. Deceased IL-6 promotes FoxP3+ Treg differentiation; Therefore, the suppression of IL-6 levels by cHMOS may contribute to the lower incidence of NEC in infants fed human milk11, 17. Indeed, the ability of cHMOS to accelerate maturation of immature intestinal mucosa while modulating acute phase signaling and promoting homeostasis may contribute to the ability of human milk feeding to decrease risk of many clinical conditions in neonates.
This modulation of basal expression of signaling agents by cHMOS addresses the ability of cHMOS to promote homeostasis of intrinsic cytokine levels in the absence of external influences. However, a critical feature of the immature gut is the excessive perturbation of its innate immune system by bacterial ligands. Pathogen associated molecular patterns (PAMPs) are powerful ligands to the sentinel molecules of the innate immune system, especially the toll-like receptors (TLRs). cHMOS modulates the response of the immature mucosa to PAMPs common in newly colonizing organisms, represented by flagellin, and PIC, and by a common mediator of inflammation, IL-1β. Flagellin, a principal component of bacterial flagella, is a virulence factor recognized by the innate immune system via Toll-like receptor (TLR) 564. PIC, a synthetic double stranded-RNA molecule, mimics RNA virus infection65 via stimulation of TLR3. IL-1β initiates and amplifies a wide variety of effects associated with innate immunity and host responses to microbial invasion and tissue injury66 mediated through stimulation of the IL-1β receptor. cHMOS modulated the increased expression of a wide spectrum of cytokines in immature intestinal tissue that typically follows exposure to PAMPs and IL-1β (Fig. 5). Two general trends were apparent: The first was that acute phase inflammatory cytokines, including IL-6, IL-8, TNF-α, TNF-β and MCP-1, were suppressed. The second was that cytokines involved in tissue repair and tissue homeostasis such as MIP-1-δ, MIP-1-β, TIMP-2 and PDGF were enhanced. These results strongly suggest potential anti-inflammatory and mucosal protective functions of cHMOS in PAMP or IL-1β stimulated immature intestine. This is consistent with, and may account for, the decreased incidence of infant inflammatory diseases among breast-fed infants, including NEC, asthma, and IBD36, 67. The direct immunomodulation described herein could be synergistic with the prebiotic effect whereby HMOS regulates the microbiome68, 69, and the microbiome promotes recovery from chemically induced (dextran sulfate sodium, DSS) injury of the intestinal mucosa70, 71.
The recognized biological functions of natural mixtures of HMOS include inhibition of adhesion of pathogens to the cell surface receptors of their target cells, an essential first step in pathogenesis. In addition, HMOS stimulate colonization of the infant gut with symbiotic mutualist bacteria of the microbiota (a prebiotic effect), which has several potential health benefits72. Both of these effects would indirectly influence inflammatory signaling in the immature intestinal mucosa. For example, dietary prebiotic oligosaccharides of plant origin are structurally dissimilar to HMOS, but their prebiotic effect is reported to enhance Th1-dependent vaccination responses in mice through modulation of the microbiome73. This report supports the concept that HMOS also directly modulate the immune system of intestinal mucosa15, 16, and provides potential pathways for their mechanism of immunomodulation.
HMOS expression is especially heterogenous, with maternal glycosyltransferase polymorphisms, notably of the fucosyltransferases, being a major factor driving variation among individuals and populations74. Moreover, the expression of HMOS is distinct in both quality and quantity between colostrum, a mother’s first milk (here, prior to day eight of lactation), and the mature milk that is produced from the second week of lactation onward. Variation in individual patterns of HMOS expression underlies differences in biological activities among different preparations of HMOS. Of the eight HMOS preparations tested herein, six were from mature milks pooled from multiple mothers (pooling decreases differences due to individual genetic variation), and two were prepared from pooled colostrums. The activities described above were robust and reproducible only in the colostrum preparations. cHMOS and mHMOS compositions were compared by LC-MS. Consistent with previous reports, cHMOS preparations had higher relative amounts of complex structures, such as lacto-N-neohexaose and monofucosyl-lacto-N-hexaose I. cHMOS also contained 6’-galactosyllactose and 3’-galactosyllactose, oligosaccharides not typically reported in human milk, but that had been reported in an analysis of colostrum20. In the H4 immature human intestinal epithelial cells, these three galactosyloligosaccharides do not have the ability exhibited by cHMOS preparations to attenuate inflammation induced by FLA and IL-1β (Fig 6C-E). In contrast, 200 μg/mL 3’-galactosyllactose, 6’-galactosyllactose and 4-galactosyllactose each inhibited the inflammatory activity in 2 μg/mL PIC stimulated H4 cells Fig 6. In the presence of 200 ng/mL PIC in H4 cells, the induction of IL-8 protein levels was inhibited in a dose-dependent manner by from 75-200 μg/mL 6’-galactosyllactose and 4-galactosyllactose. 3’-galactosyllactose inhibited induction of IL-8 across this same range. These are within the range of concentrations measured in the cHMOS preparations: 3’-galactosyllactose was 47 and 79 μg/mL, 4-galactosyllactose was 0 and 80 μg/mL, and 6’-galactosyllactose was 125 and 157 μg/mL. Thus, the galactosyllactoses may account for one of the pathways attenuated by cHMOS.
Further research can explore additional individual pathways modulated by the remaining individual oligosaccharides of cHMOS. The individual roles of each cell type of the intestinal mucosa could be defined, as well as the signaling interrelationships among the different intestinal mucosal cell types. In such studies, the numbers of observations in each treatment group could be expanded to allow more fine-grained analysis of the interactions among signals than was possible in this study.
To summarize, in quiescent non-inflamed immature human intestine, cHMOS enhance expression of genes involved in immune cell trafficking and proliferation through an ERK1/2 pathway. Simultaneously, cHMOS heighten signaling for recruitment of immune cells to the mucosal surface and intestinal immune development: cHMOS stimulates expression of Th1 polarization cytokines and suppresses Th2/Th17 polarizing cytokines, thereby moving Th1/Th2 balance toward that of mature tissues (Fig 7B). In inflamed mucosa, cHMOS suppress PAMP-induced release of acute phase inflammatory factors. Concurrently, cHMOS stimulate expression of tissue repair and tissue homeostasis cytokines. Milk oligosaccharides from colostrum are more active than oligosaccharides from mature milk, and their activities are complementary and synergistic. The immature intestinal mucosa is highly inflammatory, and therefore, the greatest need for modulation of signaling coincides with the period of colostrum production. Human milk oligosaccharides may constitute a major innate immune system that protects the infant immature intestinal mucosa, directs maturation of the immune system, and colonizes the intestine. That cHMOS per se are immune regulators could explain the clinical association between human milk consumption and reduced risk of early inflammatory bowel diseases. These data support use of human milk as a standard of care in the neonatal intensive care unit; moreover, individual cHMOS may become novel prophylactic and therapeutic agents.
METHODS
Ethics
Human milk collected within 7 days after delivery (colostrum) was obtained from the Central Massachusetts Breastmilk Bank. Human small intestinal samples were a gift of Dr. Nanda Nanthakumar (Mucosal Immunology Lab, Massachusetts General Hospital, Harvard Medical School) approved by the Committee for the Protection of Human Subjects from Research Risks (Brigham and Women’s Hospital), and the Human Investigation Committee (Massachusetts General Hospital). The research was approved by Partners Human Research Committee.
Purification of HMOS
HMOS were prepared75 from colostrum (cHMOS) or mature milk (mHMOS) by pooling individual samples into 1 L batches representing 20-50 donors. During production of early milk, volumes are limited. Even by defining any early milk (0-7 days) as ‘Colostrum’, only two batches of colostrum could be prepared from all of the milks of convenience available to us. In contrast, mature milk was not limiting, and 6 batches were prepared to increase the statistical power of the comparison. After centrifugation at 3000 × g for 40 min at 4°C, decanting eliminated the cream. The clear aqueous phase was made 67% in ethanol at 4°C and stirred overnight to precipitate protein. After centrifugation at 3000 × g for 40 min at 4°C, and decanting through a filter, the crude oligosaccharide fraction was applied to a 2 L charcoal celite column, the column was washed with 5% ethanol, and the HMOS were eluted with 65% ethanol and dried by rotary evaporation followed by lyophilization. Potential endotoxin contamination was removed by passing the HMOS preparation through a polymyxin B column until the content of endotoxin activity was below 0.01 units (Limulus Amebocyte Lysate, LAL, GenScript, Piscataway, NJ). HMOS from pooled mature milk (mHMOS) was prepared identically. HMOS preparations were lyophilized and stored at -20°C until use. The two cHMOS and six mHMOS preparations were compared for content of individual oligosaccharides by LC-MS, and their activities compared in epithelial cells by IL-8 ELISA.
Fetal intestinal organ culture
Cultures of fetal small intestine samples24 utilized fresh tissue samples from elective prostaglandin/saline-induced therapeutic abortion of 22 wk fetuses that had been transported to the laboratory in ice-cold fresh DMEM/F12 medium containing 40 mg/mL of penicillin and gentamicin. 22 weeks is approaching the age of the most immature premature infants, i.e., Extremely Low Birthweight (ELBW) neonates, and the intestine is already differentiated into colon, ileum, jejunum, and duodenum. Each experimental group was duplicate independent samples from three individuals. Tissues were rinsed in fresh ice-cold DMEM/F12 media and the mesentery and connective tissues were removed. The intestine was opened longitudinally with scissors and the mucosal side was gently washed with medium to remove meconium and other debris. The tissue was oriented under the dissecting microscope with the mucosal side up, cut into cubes of 0.5 cm on edge, transferred to 24 well cell culture plates, and incubated with 0.5 mL of DMEM/F12 media at 5% CO2 and 37°C.
Treatment of intestinal mucosa with PAMP-related stimuli and cHMOS
For each intestine, the inflammatory state of the cultured intestinal cubes was determined as IL-8 content of the supernatant. The tissue was deemed useful for these experiments only if the inflammatory state for most wells was at quiescent baseline levels (~100 pg/mL), indicating that the gut was not already inflamed due to prior damage. Half of the healthy samples were treated with 20 μL HMOS in PBS to achieve a final physiological concentration of 5 mg/mL in each well, and further incubated (basal, cHMOS) for 2 days; the other half received 20 μL of PBS alone (basal). After a 2 day incubation, some of the basal and basal-HMOS groups were further supplemented with 20 μL of an individual PAMP or IL-1β in PBS to achieve a final concentration per well of 10 ng/mL IL-1β, 100 ng/mL FLA, or 10 ng/mL PIC24. PBS alone was used for the unstimulated controls. Tissues were maintained in 5% CO2 at 37°C for an additional 18 h. Supernatants were collected for IL-8 ELISA analysis. Tissues were homogenized on ice in lysis buffer (Cell Signaling, Danvers, MA) supplemented with protease inhibitor cocktail (Cell Signaling, Danvers, MA); protein concentration was determined by BCA Protein Assay (Pierce, Rockford, Il).
Treatment of immature intestinal epithelial H4 cells with PAMP-related stimuli and cHMOS
Aliquots of the immature intestinal epithelial cell line H4 (103 cells) were cultured for two days in 96 well plates, whereupon they received 4 μL of milk oligosaccharides (final concentrations: HMOS, 5 mg/mL; 3’-GL, 4-GL and 6’-GL [Carbosynth, Compton, Berkshire, UK], 200 μg/mL), or PBS alone (basal). After an additional two days of incubation, designated wells received 20 μL of individual PAMPs or IL-1β in PBS to achieve a final concentration of 1 μg/mL of IL-1β, 200 ng/mL of FLA, or 2 μg/mL of PIC. These concentrations had been found to be effective in a variety of IEC lines in previous work (unpublished data). The unstimulated control received 20 μL of PBS without PAMP-related molecules. Cells were maintained in DMEM (Dulbecco’s Modified Eagle’s Medium) containing 10% FBS and 30 ng/mL EGF (epithelial growth factor) in 5% CO2 at 37°C for 18 h. Supernatants were collected for IL-8 ELISA analysis. Results are normalized to cell number.
IL-8 enzyme-linked immunosorbent assay
IL-8 was measured in supernatant after brief centrifugation according to the manufacturer’s instructions (R&D Systems, Minneapolis, MN). The color generated at 450 nm was measured on a Versa Max plate reader (Molecular Devices, Sunnyvale, CA). Values are normalized to the total protein concentrations of tissue lysates.
Tissue RNA extraction
RNA was extracted from homogenized human immature intestinal tissue using Trizol (Invitrogen, Grand Island, NY). Purification was by RNeasy minikit and Qiashredder columns from Qiagen (Valencia, CA) according to the manufacturer’s instructions.
RT2 profiler PCR array
Transcription of human inflammatory cytokines and receptor genes was measured in a human RT2 Profiler PCR Inflammatory Cytokines and Receptors Array (SABiosciences, PAHS-011ZA, Qiagen, Valencia, CA). Reverse transcription of 2 μg RNA from tissue was by RT2 First Strand Kit, and amplified samples were applied to PCR array plates. The RT2 Profiler arrays were probed with SYBR green/fluorescein master mix using the Profiler PCR array system. A 2-step real-time PCR reaction was initiated at 95°C (10 minutes) for 1 cycle and followed alternation of 95°C (15 seconds) and 60°C (1 minute) for 40 cycles (7500 Fast Real-Time PCR System, Applied Biosystems, Grand Island, NY). Data were analyzed using automated baseline and threshold cycle detection and interpreted via an excel-based PCR array data analytical tool from SABiosciences. Results were normalized to the expression of housekeeping genes. Up-regulation greater than 3.5 fold or down-regulation to below 0.8 demarked significant differences.
Antibody array of human signaling cytokines
Changes in cytokine and chemokine expression in intestinal tissue lysates were monitored by the RayBio Human Inflammation Antibody Array III (RayBiotech, Norcross, GA). Homogenized tissue lysates of 40 μg protein were diluted in blocking buffer and bound to array membranes overnight at 4°C. Membranes were processed according to the manufacturer’s instructions and relative quantities of inflammatory cytokines were visualized by X-ray film exposure. Spots were quantified using Image J analysis software (Eastman Kodak, Rochester, NY). Up-regulation of greater than 1.2-fold and down-regulation of lower than 0.8-fold were deemed significant.
Ingenuity pathways analysis
Data from the RT2 profiler PCR array were imported into Ingenuity Systems (www.ingenuity.com) to perform pathway analysis. Both up- and down-regulated identifiers were defined as value parameters. The generated networks were rank-ordered according to their significance score. Significance of biofunctional and canonical pathways are indicated by the Fisher Exact Test p-value. Canonical pathways are grouped into signaling pathway networks. Canonical pathways were rank-ordered according to the IPA-generated relevance ratios. This pathway analysis also generated networks of differentially regulated genes based on previously known associations between genes or proteins, but independent of established canonical pathways.
HMOS profile by LC-MS
Purified HMOS (1 mg) were dissolved in 1 mL of Milli-Q water and reduced with an excess of NaBH4 at 25°C for 16 hours. The reaction was stopped by dropwise addition of glacial HAc and samples were evaporated under nitrogen. After two washings with MeOH/AcOH 95/5 and an additional two with MeOH, samples were dissolved in 2 mL distilled water and desalted with 25 μL of DOWEX-50W resin (Sigma, St. Louis, MO), which had been pre-washed with 1 M HCl. Each fraction (3 μL) was analyzed by LC-MS (Agilent 1100 HPLC equipped with an Agilent ESI-TOF 6220 mass spectrometer, Agilent Technologies, Santa Clara, CA). LC separation was achieved on a graphitized carbon stationary phase (Hypercarb, 100 × 2.1 mm, Thermo Fisher Scientific, Waltham, MA). The mobile phase was a gradient of AcCN (B) in H2O (A) at a flow rate of 0.20 mL/min as follows: 0.0–2.0 min, 0% B; 2.0–6.0 min, 0-5% B; 6.0–9 min, 5% B; 9.0-20.0 min, 5–12% B; 20.0–22.0 min, 12% B; 22.0-26.0 min, 12-20% B; 26.0–32.0 min, 20-50% B; 32.0–35.0 min, 50-90% B; 35.0-40.0 min, 90% B; 40.0-41.0 min, 90-0% B. Each injection was followed by 10 minute equilibration at 0% B. Ion spray voltage was - 3500 V and the gas temperature was 350 °C. The fragmentor was set at 70 V. Data acquisition was performed in the negative mode starting at minute 6 of the elution, covering the m/z range 350-2300 at a 1.03 spectrum/sec scan rate.
Statistical analysis
All data represent duplicate independent values from each tissue from at least three different tissues (n=6) and are presented as the mean ± SEM. The significance of differences between mean values was determined using two-way repeated measures analysis of variance (RM ANOVA) which incorporates post-test Bonferroni correction for multiple comparisons in Prism software (Graph Pad Software, Inc. San Diego, CA). Comparison of differences between cHMOS and mHMOS activities was evaluated by Fisher’s exact ratios. Differences with p-values equal to or lower than 0.05 were considered significant.
Supplementary Material
Acknowledgments
Partially supported by NIH grants (R01HD059140, U01AI075563, and P01HD013021) and Abbott Nutrition. We thank Dr. Nanda Nanthakumar (Massachusetts General Hospital, Harvard Medical School) for providing human intestinal tissue and assisting with organ culture, and Dr. Lei Lu (Massachusetts General Hospital, Harvard Medical School) for providing H4 cells.
References
- 1.Mannick E, Udall JN., Jr Neonatal gastrointestinal mucosal immunity. Clin Perinatol. 1996;23(2):287–304. [PubMed] [Google Scholar]
- 2.Bailey M, Haverson K, Inman C, Harris C, Jones P, Corfield G, et al. The development of the mucosal immune system pre- and post-weaning: balancing regulatory and effector function. Proc Nutr Soc. 2005;64(4):451–457. doi: 10.1079/pns2005452. [DOI] [PubMed] [Google Scholar]
- 3.Rognum TO, Thrane S, Stoltenberg L, Vege A, Brandtzaeg P. Development of intestinal mucosal immunity in fetal life and the first postnatal months. Pediatr Res. 1992;32(2):145–149. doi: 10.1203/00006450-199208000-00003. [DOI] [PubMed] [Google Scholar]
- 4.Schnabl KL, Van Aerde JE, Thomson AB, Clandinin MT. Necrotizing enterocolitis: a multifactorial disease with no cure. World J Gastroenterol. 2008;14(14):2142–2161. doi: 10.3748/wjg.14.2142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lin PW, Stoll BJ. Necrotising enterocolitis. Lancet. 2006;368(9543):1271–1283. doi: 10.1016/S0140-6736(06)69525-1. [DOI] [PubMed] [Google Scholar]
- 6.Schirbel A, Fiocchi C. Targeting the innate immune system in pediatric inflammatory bowel disease. Expert Review of Gastroenterology & Hepatology. 2011;5(1):33–41. doi: 10.1586/egh.10.76. [DOI] [PubMed] [Google Scholar]
- 7.Buescher ES. Anti-inflammatory characteristics of human milk. Bioactive Components of human milk. 1997:207–222. [Google Scholar]
- 8.Ballard O, Morrow AL. Human milk composition: nutrients and bioactive factors. Pediatr Clin North Am. 2013;60(1):49–74. doi: 10.1016/j.pcl.2012.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kelly D, Coutts AG. Early nutrition and the development of immune function in the neonate. Proc Nutr Soc. 2000;59(2):177–185. doi: 10.1017/s0029665100000197. [DOI] [PubMed] [Google Scholar]
- 10.Schanler RJ, Shulman RJ, Lau C. Feeding strategies for premature infants: beneficial outcomes of feeding fortified human milk versus preterm formula. Pediatrics. 1999;103(6 Pt 1):1150–1157. doi: 10.1542/peds.103.6.1150. [DOI] [PubMed] [Google Scholar]
- 11.Lucas A, Cole TJ. Breast milk and neonatal necrotising enterocolitis. Lancet. 1990;336(8730):1519–1523. doi: 10.1016/0140-6736(90)93304-8. [DOI] [PubMed] [Google Scholar]
- 12.Klement E, Cohen RV, Boxman J, Joseph A, Reif S. Breastfeeding and risk of inflammatory bowel disease: a systematic review with meta-analysis. Am J Clin Nutr. 2004;80(5):1342–1352. doi: 10.1093/ajcn/80.5.1342. [DOI] [PubMed] [Google Scholar]
- 13.Newburg DS, Ruiz-Palacios GM, Morrow AL. Human milk glycans protect infants against enteric pathogens. Annu Rev Nutr. 2005;25:37–58. doi: 10.1146/annurev.nutr.25.050304.092553. [DOI] [PubMed] [Google Scholar]
- 14.Kunz C, Rudloff S, Baier W, Klein N, Strobel S. Oligosaccharides in human milk: structural, functional, and metabolic aspects. Annu Rev Nutr. 2000;20:699–722. doi: 10.1146/annurev.nutr.20.1.699. [DOI] [PubMed] [Google Scholar]
- 15.Kunz C, Rudloff S. Potential anti-inflammatory and anti-infectious effects of human milk oligosaccharides. Adv Exp Med Biol. 2008;606:455–465. doi: 10.1007/978-0-387-74087-4_18. [DOI] [PubMed] [Google Scholar]
- 16.Naarding MA, Ludwig IS, Groot F, Berkhout B, Geijtenbeek TBH, Pollakis G, et al. Lewis X component in human milk binds. The Journal of Clinical Investigation. 2005;115(11):3256–3264. doi: 10.1172/JCI25105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jantscher-Krenn E, Zherebtsov M, Nissan C, Goth K, Guner YS, Naidu N, et al. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut. 2012;61(10):1417–1425. doi: 10.1136/gutjnl-2011-301404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Coppa GV, Pierani P, Zampini L, Carloni I, Carlucci A, Gabrielli O. Oligosaccharides in human milk during different phases of lactation. Acta Paediatr Suppl. 1999;88(430):89–94. doi: 10.1111/j.1651-2227.1999.tb01307.x. [DOI] [PubMed] [Google Scholar]
- 19.Chaturvedi P, Warren CD, Altaye M, Morrow AL, Ruiz-Palacios G, Pickering LK, et al. Fucosylated human milk oligosaccharides vary between individuals and over the course of lactation. Glycobiology. 2001;11(5):365–372. doi: 10.1093/glycob/11.5.365. [DOI] [PubMed] [Google Scholar]
- 20.Sumiyoshi W, Urashima T, Nakamura T, Arai I, Nagasawa T, Saito T, et al. Galactosyllactoses in the Milk of Japanese Women: Changes in Concentration during the Course of Lactation. Journal of Applied Glycoscience. 2004;51(4):341–344. [Google Scholar]
- 21.Zarember KA, Godowski PJ. Tissue expression of human Toll-like receptors and differential regulation of Toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokines. J Immunol. 2002;168(2):554–561. doi: 10.4049/jimmunol.168.2.554. [DOI] [PubMed] [Google Scholar]
- 22.Humke EW, Shriver SK, Starovasnik MA, Fairbrother WJ, Dixit VM. ICEBERG: a novel inhibitor of interleukin-1beta generation. Cell. 2000;103(1):99–111. doi: 10.1016/s0092-8674(00)00108-2. [DOI] [PubMed] [Google Scholar]
- 23.Vogel C, Marcotte EM. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat Rev Genet. 2012;13(4):227–232. doi: 10.1038/nrg3185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nanthakumar NN, Fusunyan RD, Sanderson I, Walker WA. Inflammation in the developing human intestine: A possible pathophysiologic contribution to necrotizing enterocolitis. Proc Natl Acad Sci U S A. 2000;97(11):6043–6048. doi: 10.1073/pnas.97.11.6043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Youn BS, Zhang SM, Lee EK, Park DH, Broxmeyer HE, Murphy PM, et al. Molecular cloning of leukotactin-1: a novel human beta-chemokine, a chemoattractant for neutrophils, monocytes, and lymphocytes, and a potent agonist at CC chemokine receptors 1 and 3. J Immunol. 1997;159(11):5201–5205. [PubMed] [Google Scholar]
- 26.Yang L, Froio RM, Sciuto TE, Dvorak AM, Alon R, Luscinskas FW. ICAM-1 regulates neutrophil adhesion and transcellular migration of TNF-alpha-activated vascular endothelium under flow. Blood. 2005;106(2):584–592. doi: 10.1182/blood-2004-12-4942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Clements JM, Bawden LJ, Bloxidge RE, Catlin G, Cook AL, Craig S, et al. Two PDGF-B chain residues, arginine 27 and isoleucine 30, mediate receptor binding and activation. EMBO J. 1991;10(13):4113–4120. doi: 10.1002/j.1460-2075.1991.tb04988.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Morgunova E, Tuuttila A, Bergmann U, Tryggvason K. Structural insight into the complex formation of latent matrix metalloproteinase 2 with tissue inhibitor of metalloproteinase 2. Proc Natl Acad Sci U S A. 2002;99(11):7414–7419. doi: 10.1073/pnas.102185399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Baggiolini M, Walz A, Kunkel SL. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils. J Clin Invest. 1989;84(4):1045–1049. doi: 10.1172/JCI114265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sumiyoshi W, Urashima T, T N, Saito T, Tsumura N, Wang B, Brand-Miller J, Watanabe Y, Kimura K. Galactosyllactoses in the Milk of Japanese Women: Changes in Concentration during the Course of Lactation. J Appl Glycosci. 2004;51(4):341–344. [Google Scholar]
- 31.McCracken VJ, Lorenz RG. The gastrointestinal ecosystem: a precarious alliance among epithelium, immunity and microbiota. Cell Microbiol. 2001;3(1):1–11. doi: 10.1046/j.1462-5822.2001.00090.x. [DOI] [PubMed] [Google Scholar]
- 32.Nanthakumar N, Meng D, Goldstein AM, Zhu W, Lu L, Uauy R, et al. The Mechanism of Excessive Intestinal Inflammation in Necrotizing Enterocolitis: An Immature Innate Immune Response. PLoS ONE. 2011;6(3):e17776. doi: 10.1371/journal.pone.0017776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kuo S, El Guindy A, Panwala CM, Hagan PM, Camerini V. Differential appearance of T cell subsets in the large and small intestine of neonatal mice. Pediatr Res. 2001;49(4):543–551. doi: 10.1203/00006450-200104000-00017. [DOI] [PubMed] [Google Scholar]
- 34.Levy O. Innate immunity of the newborn: basic mechanisms and clinical correlates. Nat Rev Immunol. 2007;7(5):379–390. doi: 10.1038/nri2075. [DOI] [PubMed] [Google Scholar]
- 35.Abrahamsson TR, Sandberg Abelius M, Forsberg A, Bjorksten B, Jenmalm MC. A Th1/Th2-associated chemokine imbalance during infancy in children developing eczema, wheeze and sensitization. Clin Exp Allergy. 2011;41(12):1729–1739. doi: 10.1111/j.1365-2222.2011.03827.x. [DOI] [PubMed] [Google Scholar]
- 36.Gartner LM, Morton J, Lawrence RA, Naylor AJ, O’Hare D, Schanler RJ, et al. Breastfeeding and the use of human milk. Pediatrics. 2005;115(2):496–506. doi: 10.1542/peds.2004-2491. [DOI] [PubMed] [Google Scholar]
- 37.Newburg DS. Neonatal protection by an innate immune system of human milk consisting of oligosaccharides and glycans. J Anim Sci. 2009;87(13 Suppl):26–34. doi: 10.2527/jas.2008-1347. [DOI] [PubMed] [Google Scholar]
- 38.Goldman AS, Goldblum RM, Hanson LA. Anti-inflammatory systems in human milk. Adv Exp Med Biol. 1990;262:69–76. doi: 10.1007/978-1-4613-0553-8_6. [DOI] [PubMed] [Google Scholar]
- 39.Eiwegger T, Stahl B, Schmitt J, Boehm G, Gerstmayr M, Pichler J, et al. Human milk--derived oligosaccharides and plant-derived oligosaccharides stimulate cytokine production of cord blood T-cells in vitro. Pediatr Res. 2004;56(4):536–540. doi: 10.1203/01.PDR.0000139411.35619.B4. [DOI] [PubMed] [Google Scholar]
- 40.Armogida SA, Yannaras NM, Melton AL, Srivastava MD. Identification and quantification of innate immune system mediators in human breast milk. Allergy Asthma Proc. 2004;25(5):297–304. [PubMed] [Google Scholar]
- 41.Kramer MS, Chalmers B, Hodnett ED, Sevkovskaya Z, Dzikovich I, Shapiro S, et al. Promotion of Breastfeeding Intervention Trial (PROBIT): a randomized trial in the Republic of Belarus. JAMA. 2001;285(4):413–420. doi: 10.1001/jama.285.4.413. [DOI] [PubMed] [Google Scholar]
- 42.Hofmann SR, Ettinger R, Zhou YJ, Gadina M, Lipsky P, Siegel R, et al. Cytokines and their role in lymphoid development, differentiation and homeostasis. Curr Opin Allergy Clin Immunol. 2002;2(6):495–506. doi: 10.1097/00130832-200212000-00004. [DOI] [PubMed] [Google Scholar]
- 43.Takeuchi H, Fujimoto A, Tanaka M, Yamano T, Hsueh E, Hoon DS. CCL21 chemokine regulates chemokine receptor CCR7 bearing malignant melanoma cells. Clin Cancer Res. 2004;10(7):2351–2358. doi: 10.1158/1078-0432.ccr-03-0195. [DOI] [PubMed] [Google Scholar]
- 44.Sallusto F, Mackay CR, Lanzavecchia A. Selective expression of the eotaxin receptor CCR3 by human T helper 2 cells. Science. 1997;277(5334):2005–2007. doi: 10.1126/science.277.5334.2005. [DOI] [PubMed] [Google Scholar]
- 45.Garcia-Zepeda EA, Combadiere C, Rothenberg ME, Sarafi MN, Lavigne F, Hamid Q, et al. Human monocyte chemoattractant protein (MCP)-4 is a novel CC chemokine with activities on monocytes, eosinophils, and basophils induced in allergic and nonallergic inflammation that signals through the CC chemokine receptors (CCR)-2 and -3. J Immunol. 1996;157(12):5613–5626. [PubMed] [Google Scholar]
- 46.Ahuja SK, Murphy PM. The CXC chemokines growth-regulated oncogene (GRO) alpha, GRObeta, GROgamma, neutrophil-activating peptide-2, and epithelial cell-derived neutrophil-activating peptide-78 are potent agonists for the type B, but not the type A, human interleukin-8 receptor. J Biol Chem. 1996;271(34):20545–20550. doi: 10.1074/jbc.271.34.20545. [DOI] [PubMed] [Google Scholar]
- 47.Shannon LA, Calloway PA, Welch TP, Vines CM. CCR7/CCL21 Migration on Fibronectin Is Mediated by Phospholipase Cγ1 and ERK1/2 in Primary T Lymphocytes. Journal of Biological Chemistry. 2010;285(50):38781–38787. doi: 10.1074/jbc.M110.152173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Meloche S, Pouyssegur J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene. 2007;26(22):3227–3239. doi: 10.1038/sj.onc.1210414. [DOI] [PubMed] [Google Scholar]
- 49.Maheshwari MZ A. ontogeny of the intestinal immune system. Immunology and Infection. 2006;2(10):18–26. [Google Scholar]
- 50.Neurath MF, Finotto S, Glimcher LH. The role of Th1/Th2 polarization in mucosal immunity. Nat Med. 2002;8(6):567–573. doi: 10.1038/nm0602-567. [DOI] [PubMed] [Google Scholar]
- 51.Kidd P. Th1/Th2 balance: the hypothesis, its limitations, and implications for health and disease. Altern Med Rev. 2003;8(3):223–246. [PubMed] [Google Scholar]
- 52.Randolph DA, Huang G, Carruthers CJ, Bromley LE, Chaplin DD. The role of CCR7 in TH1 and TH2 cell localization and delivery of B cell help in vivo. Science. 1999;286(5447):2159–2162. doi: 10.1126/science.286.5447.2159. [DOI] [PubMed] [Google Scholar]
- 53.Vigne S, Palmer G, Martin P, Lamacchia C, Strebel D, Rodriguez E, et al. IL-36 signaling amplifies Th1 responses by enhancing proliferation and Th1 polarization of naive CD4+ T cells. Blood. 2012;120(17):3478–3487. doi: 10.1182/blood-2012-06-439026. [DOI] [PubMed] [Google Scholar]
- 54.Shaffer AL, Yu X, He Y, Boldrick J, Chan EP, Staudt LM. BCL-6 represses genes that function in lymphocyte differentiation, inflammation, and cell cycle control. Immunity. 2000;13(2):199–212. doi: 10.1016/s1074-7613(00)00020-0. [DOI] [PubMed] [Google Scholar]
- 55.Takatsu K. Interleukin 5 and B cell differentiation. Cytokine Growth Factor Rev. 1998;9(1):25–35. doi: 10.1016/s1359-6101(97)00034-8. [DOI] [PubMed] [Google Scholar]
- 56.Clinton C. Development of the infant immune function and the effects of breast milk. Natural medicine Journal. 2010;2(8):3–6. [Google Scholar]
- 57.Kumar V, Sharma A. Neutrophils: Cinderella of innate immune system. International Immunopharmacology. 2010;10(11):1325–1334. doi: 10.1016/j.intimp.2010.08.012. [DOI] [PubMed] [Google Scholar]
- 58.Liang SC, Tan X-Y, Luxenberg DP, Karim R, Dunussi-Joannopoulos K, Collins M, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. The Journal of Experimental Medicine. 2006;203(10):2271–2279. doi: 10.1084/jem.20061308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu Rev Immunol. 2009;27:485–517. doi: 10.1146/annurev.immunol.021908.132710. [DOI] [PubMed] [Google Scholar]
- 60.Maddur MS, Miossec P, Kaveri SV, Bayry J. Th17 cells: biology, pathogenesis of autoimmune and inflammatory diseases, and therapeutic strategies. Am J Pathol. 2012;181(1):8–18. doi: 10.1016/j.ajpath.2012.03.044. [DOI] [PubMed] [Google Scholar]
- 61.Kimura A, Kishimoto T. IL-6: regulator of Treg/Th17 balance. Eur J Immunol. 2010;40(7):1830–1835. doi: 10.1002/eji.201040391. [DOI] [PubMed] [Google Scholar]
- 62.Dingle BM, Liu Y, Fatheree NY, Min J, Rhoads JM, Tran DQ. FoxP3(+) Regulatory T Cells Attenuate Experimental Necrotizing Enterocolitis. PLoS One. 2013;8(12):e82963. doi: 10.1371/journal.pone.0082963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Weitkamp J-H, Koyama T, Rock MT, Correa H, Goettel JA, Matta P, et al. Necrotising enterocolitis is characterised by disrupted immune regulation and diminished mucosal regulatory (FOXP3)/effector (CD4, CD8) T cell ratios. Gut. 2012 doi: 10.1136/gutjnl-2011-301551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hayashi F, Smith KD, Ozinsky A, Hawn TR, Yi EC, Goodlett DR, et al. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature. 2001;410(6832):1099–1103. doi: 10.1038/35074106. [DOI] [PubMed] [Google Scholar]
- 65.De Miranda J, Yaddanapudi K, Hornig M, Lipkin WI. Astrocytes recognize intracellular polyinosinic-polycytidylic acid via MDA-5. FASEB J. 2009;23(4):1064–1071. doi: 10.1096/fj.08-121434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Eder C. Mechanisms of interleukin-1β release. Immunobiology. 2009;214(7):543–553. doi: 10.1016/j.imbio.2008.11.007. [DOI] [PubMed] [Google Scholar]
- 67.Sullivan S, Schanler RJ, Kim JH, Patel AL, Trawöger R, Kiechl-Kohlendorfer U, et al. An Exclusively Human Milk-Based Diet Is Associated with a Lower Rate of Necrotizing Enterocolitis than a Diet of Human Milk and Bovine Milk-Based Products. The Journal of Pediatrics. 2010;156(4):562–567. e561. doi: 10.1016/j.jpeds.2009.10.040. [DOI] [PubMed] [Google Scholar]
- 68.Yu ZT, Chen C, Kling DE, Liu B, McCoy JM, Merighi M, et al. The principal fucosylated oligosaccharides of human milk exhibit prebiotic properties on cultured infant microbiota. Glycobiology. 2013;23(2):169–177. doi: 10.1093/glycob/cws138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Yu ZT, Chen C, Newburg DS. Utilization of major fucosylated and sialylated human milk oligosaccharides by isolated human gut microbes. Glycobiology. 2013;23(11):1281–1292. doi: 10.1093/glycob/cwt065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nanthakumar NN, Meng D, Newburg DS. Glucocorticoids and microbiota regulate ontogeny of intestinal fucosyltransferase 2 requisite for gut homeostasis. Glycobiology. 2013;23(10):1131–1141. doi: 10.1093/glycob/cwt050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Weiss GA, Hennet T. The role of milk sialyllactose in intestinal bacterial colonization. Adv Nutr. 2012;3(3):483S–488S. doi: 10.3945/an.111.001651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Coppa GV, Bruni S, Morelli L, Soldi S, Gabrielli O. The First Prebiotics in Humans: Human Milk Oligosaccharides. Journal of Clinical Gastroenterology. 2004;38:S80–S83. doi: 10.1097/01.mcg.0000128926.14285.25. [DOI] [PubMed] [Google Scholar]
- 73.Vos AP, Haarman M, van Ginkel JW, Knol J, Garssen J, Stahl B, et al. Dietary supplementation of neutral and acidic oligosaccharides enhances Th1-dependent vaccination responses in mice. Pediatr Allergy Immunol. 2007;18(4):304–312. doi: 10.1111/j.1399-3038.2007.00515.x. [DOI] [PubMed] [Google Scholar]
- 74.Newburg DS, Ruiz-Palacios GM, Altaye M, Chaturvedi P, Meinzen-Derr J, Guerrero MdL, et al. Innate protection conferred by fucosylated oligosaccharides of human milk against diarrhea in breastfed infants. Glycobiology. 2004;14(3):253–263. doi: 10.1093/glycob/cwh020. [DOI] [PubMed] [Google Scholar]
- 75.Crane JK, Azar SS, Stam A, Newburg DS. Oligosaccharides from human milk block binding and activity of the Escherichia coli heat-stable enterotoxin (STa) in T84 intestinal cells. J Nutr. 1994;124(12):2358–2364. doi: 10.1093/jn/124.12.358. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.