

Abstract
Context:
Mutations of succinate dehydrogenase A/B/C/D genes (SDHx) increase susceptibility to development of pheochromocytomas and paragangliomas (PPGLs), with particularly high rates of malignancy associated with SDHB mutations.
Objective:
We assessed whether altered succinate dehydrogenase product-precursor relationships, manifested by differences in tumor ratios of succinate to fumarate or other metabolites, might aid in identifying and stratifying patients with SDHx mutations.
Design, Setting, and Patients:
PPGL tumor specimens from 233 patients, including 45 with SDHx mutations, were provided from eight tertiary referral centers for mass spectrometric analyses of Krebs cycle metabolites.
Main Outcome Measure:
Diagnostic performance of the succinate:fumarate ratio for identification of pathogenic SDHx mutations.
Results:
SDH-deficient PPGLs were characterized by 25-fold higher succinate and 80% lower fumarate, cis-aconitate, and isocitrate tissue levels than PPGLs without SDHx mutations. Receiver-operating characteristic curves for use of ratios of succinate to fumarate or to cis-aconitate and isocitrate to identify SDHx mutations indicated areas under curves of 0.94 to 0.96; an optimal cut-off of 97.7 for the succinate:fumarate ratio provided a diagnostic sensitivity of 93% at a specificity of 97% to identify SDHX-mutated PPGLs. Succinate:fumarate ratios were higher in both SDHB-mutated and metastatic tumors than in those due to SDHD/C mutations or without metastases.
Conclusions:
Mass spectrometric-based measurements of ratios of succinate:fumarate and other metabolites in PPGLs offer a useful method to identify patients for testing of SDHx mutations, with additional utility to quantitatively assess functionality of mutations and metabolic factors responsible for malignant risk.
Pheochromocytomas and paragangliomas (PPGLs) are adrenal and extra-adrenal tumors of neural crest origin. At least one-third of cases are explained by germline mutations in at least ten tumor-susceptibility genes (1): neurofibromatosis type 1 (NF1); rearranged during transfection (RET) protooncogene, transmembrane protein 127 (TMEM127); myc-associated factor X (MAX); von Hippel-Lindau tumor suppressor (VHL) or one of the genes for succinate dehydrogenase subunits (SDHA, B, C, D, AF2).
PPGLs due to mutations in SDHB are predominantly extra-adrenal with low catecholamine contents and associated with high risk for metastatic disease (2, 3). Screening for SDHx-related mutations is therefore important for identifying patients and family members at risk for developing malignancy, additional PPGLs as well as other types of tumors that also result from SDHx mutations. The latter are now established to include gastrointestinal stromal tumors and renal cell carcinomas (4, 5). The rationale is that earlier detection of tumors through screening of identified patients should result in improved therapeutic outcome and reduced risk of malignancy.
Identification of patients with PPGLs resulting from SDHx mutations can be facilitated by considerations of patterns of biochemical parameters and immunohistochemical staining for SDHB protein in resected tumor material (6, 7). With the advent of next-generation sequencing, such triaging for targeted genetic testing may become obsolete (8). Nevertheless, next-generation sequencing does not allow detection of SDHx gene deletions particularly important to consider in patients with PPGLs (9, 10). Also as outlined elsewhere (11), broad nonselective screening by next-generation sequencing has other potential limitations. A particularly major challenge concerns identification of gene variants that are pathogenic among a larger proportion of variants of uncertain significance. Gene prediction tools are used in this context, but the ideal method is assignment of functionality using quantitative readouts.
As demonstrated for myeloid leukemia and gliomas due to mutations of isocitrate dehydrogenase, measurements of metabolites of the enzyme can provide especially useful biomarkers of disease and quantitative tools for assessment of functionality and disease stratification (12–14). Assessment of Krebs cycle metabolism is, however, not only of interest in leukemia and gliomas, but also in many other neoplasms including PPGLs (15, 16). In particular, measurements of tumor tissue levels of succinate and fumarate, the respective substrate and metabolite of succinate dehydrogenase, have now been shown in a proof-of-principle pilot study to offer a potentially useful tool for identification of patients with SDHx mutations (17). Utility of the succinate to glutamate ratio for the same purpose has also been demonstrated in another small patient cohort (18).
The present study extends the above preliminary findings to a much larger cohort of 233 patients with PPGLs in whom tumor tissue levels of succinate, fumarate and other Krebs cycle metabolites were measured using a novel, rapid, and simple liquid chromatographic tandem mass spectrometric method. The primary objective of the study was to assess utility of the succinate:fumarate ratio for identification of patients with SDHx mutations. For this we used a training set of tumor samples from 49 patients to establish diagnostic cut-off values, followed by a blinded validation series of 184 samples to establish diagnostic efficacy. For the secondary objective, both sets of data were employed to establish utility for disease stratification according to the specific gene inactivated as well as tumor location and presence of metastatic disease.
Materials and Methods
Patients and tumor procurement
PPGLs were collected from 233 patients (Table 1) undergoing surgical resection of primary tumors. Tumor procurement was approved under Intramural Review Board protocols at each participating center. All tumor specimens were provided as frozen fragments (10–50 mg). Fifteen cases were also available as formalin-fixed paraffin-embedded (FFPE) specimens to assess utility of such samples for metabolite analyses. As a first step, 50 frozen specimens from 49 patients with known mutational status were analyzed as a training set. The second step involved provision of 184 tumor samples as a validation set. These latter specimens were provided blinded to their mutational status. Krebs cycle metabolites were analyzed and an interpretation concerning the likelihood of an SDHx mutation was provided back to the centers. The primary study endpoint was the evaluation of diagnostic sensitivity and specificity using the succinate:fumarate ratio to define SDHx mutational status.
Table 1.
Patient Demographics and Clinical Characteristics
| Germline Mutation | No. of Patients | Sex (M/F) | Age Range (y; mean) | Tumor Location (A/E/HN) | Metastatic Disease |
|---|---|---|---|---|---|
| Training set | |||||
| SDHB | 8 | 4/4 | 15–46; 33 | 1/7/0 | 4 |
| SDHD | 3 | 3/0 | 32–46; 39 | 1/1/1 | 0 |
| VHL | 3 | 1/2 | 10–32; 24 | 2/1/0 | 0 |
| RET | 11 | 6/5 | 31–67; 44 | 11/0/0 | 0 |
| NF1 | 8 | 7/1 | 17–59; 42 | 8/0/0 | 0 |
| None | 16 | 7/9 | 18–76; 46 | 12/3a/0 (1U) | 1 |
| Validation set | |||||
| SDHB | 16 | 9/7 | 14–57; 34 | 2/11/3 | 4 |
| SDHD | 16 | 6/10 | 12–76; 37 | 1/1/14 | 1 |
| SDHC | 2 | 1/1 | 16–60; 38 | 0/1/1 | 0 |
| VHL | 8 | 6/2 | 11–43; 22 | 7/1/0 | 0 |
| RET | 19 | 7/12 | 18–65; 37 | 19/0/0 | 0 |
| NF1 | 6 | 2/4 | 38–76; 51 | 6/0/0 | 0 |
| TMEM127 | 3 | 1/2 | 21–54; 36 | 3/0/0 | 0 |
| Max | 1 | 0/1 | 46 | 1/0/0 | 1 |
| None | 113 | 62/51 | 13–78; 50 | 86/21/6 | 9 |
Abbreviations: U, unknown; A, adrenal; E, extra-adrenal (thoracic and abdominal); HN, head and neck.
For one patient, two simultaneously occurring extra-adrenal tumors were analysed.
Measurements of tissue metabolites
Processed tissues from primary tumors were analyzed by ultrahigh pressure liquid chromatography with tandem mass spectrometry (UPHPLC-MS/MS). Fresh frozen tumor tissue (5–10 mg) was homogenized in 500 μL LC/MS grade methanol containing the internal standard mixture (Supplemental Table 1) by vortexing with a metal bead for 2 min. To achieve separation of insoluble debris, homogenates were centrifuged at 2000xg for 5 min at 4°C. Supernatants were dried using a speed vac concentrator (Thermo Scientific) and stored at −80°C. On the day of analysis, residues were resuspended in mobile phase and cleared with a 0.2 μm centrifugal filter. FFPE tissue was sectioned, and metabolites were extracted from two slices of 50 μm thickness using the method of Kelly et al (19).
UHPLC-MS/MS was performed on the same instrumentation described for our routine diagnostic measurements of plasma and urinary normetanephrine, metanephrine, and methoxytyramine (20, 21). This included an Acquity UHPLC system (Waters), equipped with a binary pump manager, a sample manager and a column manager, coupled to an API QTRAP 5500 triple quadrupole mass spectrometer (AB Sciex). A Waters Acquity UPLC® HSS T3 column (1.8 μm, 2.1 × 100 mm) was used for chromatographic separation. Mobile phases consisted of 0.2% formic acid in water (A) and 0.2% formic acid in acetonitrile (B) set at a flow rate of 0.459 mL/min. For each injection an initial gradient of 5% mobile phase B for 0.37 min was increased to 30% at 4.87 min and 100% at 5.37 min; after column washing with 100% B for 0.5 min, the column was re-equilibrated with 5% mobile phase B ready for the next injection. Targeted analyses were performed in multiple reaction monitoring scan mode with use of negative electrospray ionization, as described elsewhere (22). Multiple reaction monitoring transitions for quantification and qualification are listed in Supplemental Table 2.
Interassay variation was established by measuring tissue aliquots of two different tumors at different days with one tumor showing moderate levels of succinate (QC1) and the other one having a high succinate concentration (QC2). Interassay coefficients of variation were determined at 6% in QC1 and 4.2% in QC2 for succinate, and 6.6% and 23.8% for fumarate, respectively. Interassay variations for all metabolites are recorded in Supplemental Table 1.
Genetic characterization
Genetic testing was performed to confirm or exclude the presence of germline mutations in SDHx genes. With the exception of patients in whom a germline or somatic mutation in another PPGL susceptibility gene (RET, NF1, TMEM127, MAX, VHL, HIF2A) had previously been identified, all patients were tested for SDHAF1, SDHAF2, SDHA, SDHB, SDHC or SDHD point mutations by automated sequencing, with detection of gross deletions by multiplex ligation-dependent probe amplification analysis. Diagnosis of NF1 was based on clinical presentation. Genetic testing for RET, TMEM127, MAX and VHL mutations was performed in 70% of the cases. PCR conditions and primers are available on request. The Alamut mutation interpretation software (http://www.interactive-biosoftware.com) was used to assess pathogenicity of previously unidentified variants.
Follow-up testing
Follow-up testing, utilizing additional tumor specimens and samples of germline DNA, was carried out in cases where tumor succinate:fumarate ratios indicated either false-negative results (ratios below the cut-off in patients with SDHx mutations) or false-positive results (ratios above the cut-off in patients without SDHx mutations), this according to a written study plan outlined to all investigators prior to their contributions of patient specimens for the validation series. Genetic testing in patients with false-positive results included evaluation of tumor tissues for somatic mutations of all SDHx genes as well as of SDHAF2 and SDHAF1. Loss of heterozygosity studies were also conducted mainly directed to false-negative results. For both false-positive and false-negative results, tissue samples were re-examined for metabolite levels, SDH enzyme activity, and protein content of SDHB. SDH activities were measured in tissue homogenates, and western blots were performed as described previously (23). Densitometry was undertaken using Chemidoc software.
Statistical analysis
Statistical analysis was performed using SigmaPlot 12.0. The differences between two groups were analyzed by a t-test, when data were distributed normally or alternatively by rank sum test. Comparisons between multiple groups were undertaken by one-way ANOVA on ranks. Logistic regression was used to establish receiver-operating characteristic (ROC) curves, which were used to identify the optimal cut-off values for discriminating PPGLs with SDHx-related mutations from others. Sensitivity was calculated by dividing the number of true positives by the sum of true positives and false negatives. Specificity was estimated by dividing the number of true negatives by the sum of true negatives and false positives. Values are provided as mean ± SEM.
Results
Succinate to fumarate ratios
Succinate:fumarate ratios for PPGLs of the training set were 454-fold higher (P < .001) in SDHx-mutated than other tumors, with values spanning a range of two orders of magnitude (Figure 1a). Using ROC curve analysis a cut-off of 97.7 for the succinate:fumarate ratio was determined to provide 100% sensitivity and 97.3% specificity (Supplemental Figure 1). In three paraganglial specimens, two of which were from separate extra-adrenal tumors from the same patient, elevated succinate:fumarate ratios were detected, but no germline SDHx mutation was noted (two false-positives in the training set).
Figure 1.

Box and whisker plots of succinate:fumarate-ratios for PPGLs analyzed in the training (A) and validation set (B). Boxes span the 25–75 percentile and whiskers span the 10–90 percentile. Cut-off, determined by ROC curve analysis of the training set (97.7), is marked as a dashed line. Black circles indicate samples with values below the cut-off for SDHx tumors or above the cut-off for non-SDHx PPGLs. Two samples were excluded from the analysis of the validation set due to unusually high fumarate and malate values.
Succinate:fumarate ratios for PPGLs of the validation set were also considerably higher (P < .001) in SDHx-mutated vs other PPGLs (Figure 1b), but the 94-fold difference was smaller than that for the training set. This smaller difference reflected five specimens with ratios above the cut-off, but no SDHx germline mutation (five false-positives in the validation set) and a further three specimens from patients with confirmed SDHx germline mutations, but with tumor succinate:fumarate ratios below the cut-off (three false-negatives). For two samples, no interpretation was provided back to contributing investigators due to findings of high tissue succinate levels with unusual out of range values for fumarate. These samples were thus not included in the final analysis.
Follow-up testing
Follow-up genetic testing confirmed the SDHx germline mutation status in all three patients with false-negative succinate:fumarate ratios (Supplemental Table 3). In all except one false-positive case, exhaustive testing of SDHx genes in tumor samples failed to reveal any additional genetic variant that could explain the high tumor succinate:fumarate ratios. That one case among the seven with false-positive succinate:fumarate ratios involved a patient in whom testing of the head and neck paraganglioma (HNP) revealed a somatic SDHB mutation (c.380T > G) confirmed at two independent centers. That mutation was predicted to be deleterious and the patient was transferred from a false- to a true-positive category (six false-positives in total).
All three patients with false-negative tumor succinate:fumarate ratios had HNPs (Supplemental Table 3). SDH enzyme activity and western blot analyses indicated results consistent with mutation-status in all three patients. Specifically, all SDHx-mutated tumors returning false-negative results for tumor succinate:fumarate ratios had reduced levels of SDH activity (33 ± 6%) compared to wild-type controls (93 ± 6%); tumor tissue also showed reduced SDHB protein to 36 ± 2% of controls by western blot.
Among the patients with false-positive elevations of succinate:fumarate ratios, only one presented with an adrenal tumor, the others with paragangliomas (Supplemental Table 3). SDH western blot and activity measurements supported the findings of high tumor tissue succinate:fumarate ratios in four of the six patients with false-positive results despite lack of evidence for SDHx mutations.
Profiling of Krebs cycle metabolites
Most Krebs cycle metabolites showed significant differences between tumors with and without SDHx mutations (Figure 2). Succinate was 25-fold higher in PPGLs with SDHx mutations, whereas fumarate, citrate, cis-aconitate and isocitrate were, respectively, 80%, 60%, 82%, and 80% lower. No significant differences between groups were observed for α-ketoglutarate, malate, pyruvate, and lactate.
Figure 2.

Box and whisker plots comparing metabolite levels between 45 PPGLs with SDHx mutations and 189 other PPGLs. Boxes span the 25–75 percentile and whiskers span the 10–90 percentile, with outliers as points. Y-axes indicate tissue levels of metabolites in ng/mg tissue. Significance was established by the Mann-Whitney Rank sum test.
Analysis of subcomponents of the data set showed that PPGLs due to VHL mutations (germline or somatic), similar to those due to SDHx mutations, also contain less (P < .05) fumarate, citrate, cis-aconitate, and isocitrate compared to tumors harboring RET, NF1 or TMEM127 mutations (Supplemental Figure 2, b-e). Succinate, however, was only elevated in tumors with SDH inactivation (Supplemental Figure 2a). Pyruvate was slightly higher in SDHx and VHL tumors compared to PPGLs with NF1, RET or TMEM127 mutations, but significance was only reached for VHL tumors (Supplemental Figure 2f).
SDHx mutational status, malignancy, and tumor location
Tumor succinate levels were similar among tumors due to SDHB and SDHC/SDHD mutations; however, fumarate levels were lower and succinate:fumarate ratios higher (P ≤ .01) in SDHB compared to SDHC/D tumors (Figure 3a). Among tumors due to SDHx mutations, fumarate was lower (P = .007) and succinate:fumarate ratios higher (P = .022) in those associated with metastatic disease than in those without metastatic disease (Figure 3b). Tumors at head and neck locations had higher (P < .001) fumarate levels and lower (P < .001) succinate:fumarate ratios compared to those at adrenal, abdominal or thoracic locations (Figure 3c).
Figure 3.
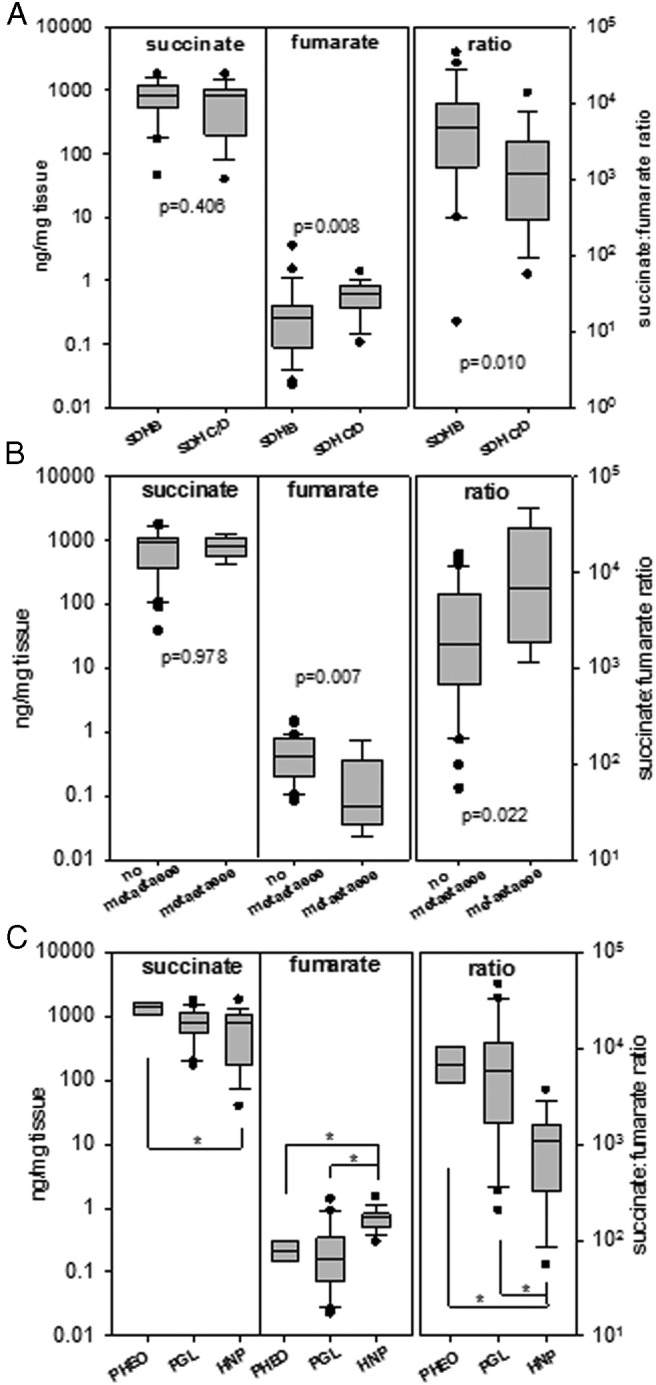
Box and whisker plots comparing succinate and fumarate levels and succinate:fumarate ratios between subgroups of SDHx-related PPGLs (45 tumors in total). (A) 24 PPGLs with SDHB vs 21 with SDHC/D mutations; (B) 9 PPGLs with vs 36 without metastases; (C) adrenal (PHEO, 5) vs abdominal or thoracic tumor location (PGL, 21) vs head and neck paragangliomas (HNP, 19). Boxes span the 25–75 percentile and whiskers span the 10–90 percentile, with outliers as points. Significance was established by Mann-Whitney Rank sum test or ANOVA on Ranks/Dunn's test (asterisks, P < .05).
Succinate:fumarate ratio determined in FFPE tissue
In a subset of samples, direct comparison of the succinate:fumarate ratios between fresh-frozen and FFPE tissue showed similar values for non-SDHx tumors (Supplemental Figure 3). Five of six SDHx-related tumors had succinate:fumarate ratios above the cut-off for fresh-frozen tissue.
Identification of SDHx mutations based on Krebs cycle metabolite levels
For all tumors in the validation set considered together, diagnostic sensitivity and specificity, respectively, reached 90.9% and 97.3%; however, when tumors were separated according to location, sensitivity increased to 100% for adrenal, abdominal, and thoracic PPGLs (Table 2). For head and neck tumors, sensitivity was 83.3% and specificity 80.0%.
Table 2.
Diagnostic Sensitivity and Specificity for Succinate:Fumarate Ratios to Identify SDHx Mutations
| All PPGLs | PPGLs Excl. HNP | HNP | |
|---|---|---|---|
| Validation set | |||
| Sensitivity (%) | 90.9 (30/33) | 100 (15/15) | 83.3 (15/18) |
| Specificity (%) | 97.3 (145/149) | 97.9 (141/144) | 80.0 (4/5) |
| Training and validation set | |||
| Sensitivity (%) | 93.2 (41/44) | 100 (25/25) | 84.2 (16/19) |
| Specificity (%) | 96.8 (181/187) | 97.3 (177/182) | 80.0 (4/5) |
Analysis excluded two SDHx tumors with low succinate:fumarate ratios, which are the result of extremely high fumarate values. These samples also showed high, for SDHx mutations typical succinate levels.
Abbreviation: HNP, head and neck paraganglioma.
For the succinate:fumarate ratio, an area under the ROC curve of 0.94 was calculated (Figure 4a) with an overall sensitivity for the combined training and validation set of 93.2% and specificity of 96.8%. Succinate to citrate, succinate to cis-aconitate and succinate to isocitrate ratios also provided high diagnostic performance with respective areas under ROC curves of 0.94, 0.95 and 0.96 (Figure 4b).
Figure 4.
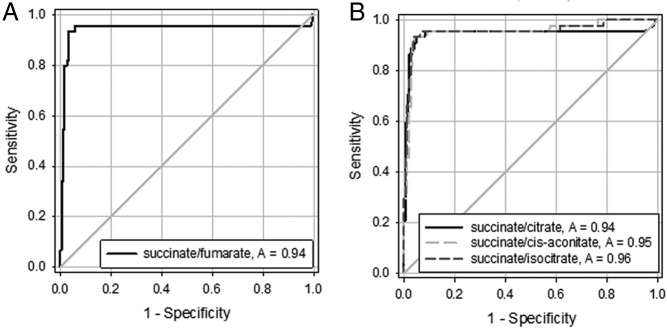
ROC curves for identification of SDHx mutations according to succinate to fumarate ratios in panel A, and succinate to citrate, succinate to cis-aconitate, and succinate to isocitrate in panel B. ROC curves are derived from both training and validation set.
Discussion
The present study is the first comprehensive analysis of a large tumor set from multiple centers establishing the validity of measuring succinate:fumarate ratios for both identifying PPGLs due to SDHx mutations and quantitatively confirming functionality of identified mutations. Differences in Krebs cycle metabolite profiles according to driver mutation, metastasis and tumor location indicate further potential as a quantitative method for disease stratification.
With an area under the ROC curve of 0.94 and overall diagnostic sensitivity of 93% and specificity of 97%, use of tumor tissue succinate:fumarate ratios to identify presence of SDHx mutations is in par with the diagnostic performance of many other tests used in the routine laboratory. Moreover with the exclusion of HNPs, the diagnostic sensitivity of succinate:fumarate ratios to identify tumors due to SDHx mutations increased to 100%. Thus, for thoracic, abdominal and adrenal tumors, a succinate:fumarate ratio below the cut-off reliably excludes an SDHx mutation. HNPs are biochemically different to other PPGLs, since these tumors mostly lack catecholamine production (24). Intensity of the succinate to fumarate signal is also lower in SDHx-mutated HNPs than in other SDHx-mutated PPGLs, which might reflect differences in the homogeneity of chromaffin tumor cell populations and overall tumor tissue loss of SDH.
With a specificity of 97% the method provides a high level of confidence that patients with positive results do in fact harbor an SDHx mutation. Of interest, among the patients with false-positive elevations of succinate:fumarate ratios most had other evidence pointing to loss of SDH function. Despite exhaustive follow-up mutation analysis no explanation was found except for one patient with a somatic SDHB mutation. Possibly chromosomal translocations, gene methylation, or mutations in promoter regions of SDHx genes may provide explanations. Further unexplored possibilities include mutations in genes influencing the functionality of the SDH enzyme complex or metabolic flux through the Krebs cycle. In a subgroup of HNPs from patients without SDHx germline mutations, signaling of the HIF-1alpha/miRNA-210 axis resulted in decreased iron-sulfur cluster scaffold protein, which is suspected to influence SDH protein stability (25). Furthermore, substantial increases in succinate accumulation have been demonstrated through processes involving increased import and metabolism of glutamine (26). Another study identified the mitochondrial chaperone TRAP1 as an inhibitor of SDH also leading to increases in succinate levels (27).
In addition to identification and quantitative functional characterization of SDHx mutations, profiling of Krebs cycle metabolites may also be useful for disease stratification, this following from the findings of Letouze et al that increased tumor levels of succinate lead to DNA hypermethylation as a critical tumorigenic mechanism (28). In that study stronger DNA hypermethylation and the more aggressive nature of PPGLs due to SDHB than SDHD mutations was hypothesized to be linked to differences in tumor succinate levels. Our findings of higher succinate to fumarate ratios in PPGLs due to mutations of SDHB than due to mutations of SDHC/D supports the suggestion by Letouze et al of stronger functional effects of the former than of the latter mutations. The higher tumor tissue succinate:fumarate ratios in PPGLs associated with metastases than in those without metastatic involvement further supports the possibility that these measurements might have prognostic value for assessing likelihood of malignancy. Further studies are required, however, to clarify any confounding influence of tumor location.
Increased succinate levels in SDHx mutated tumors have been previously described in small sets of PPGLs (28–30) and cell culture models (31, 32). Our findings of low fumarate in SDHx mutated compared to other tumors are also consistent with studies in SDH-deficient yeast strains (32). In addition to the above changes, we also show that citrate, cis-aconitate, and isocitrate were all decreased by SDHx mutations reflecting their lowered rates of production and indicating additional diagnostic utility to fumarate. Lower levels of fumarate, citrate, cis-aconitate, and isocitrate in SDHx- and VHL-related PPGLs compared to PPGLs due to RET, NF1, and TMEM127 mutations are consistent with another study demonstrating decreased oxidative phosphorylation in all PPGLs characterized by a pseudohypoxic signature (30, 33).
Importantly, the present data were generated on mass-spectrometric instrumentation, which is rapidly becoming the gold-standard technique for measurements of low molecular weight analytes in the routine diagnostic laboratory. More specifically, the measurements of Krebs cycle metabolites were performed using an instrument also used for measurements of plasma and urine metanephrines in the routine diagnosis of PPGLs (20, 21). Versatility of the instrumentation, achieved through high sample throughput and ease in changing from one analytical application to another, ensures utility for multiple applications. The simplicity of sample preparation also ensures low running costs of consumables. Furthermore, our finding that succinate, fumarate, and other metabolites are detectable in paraffin-fixed tissue indicates that the method is not restricted to frozen specimens, but may also be useful for retrospective analyses of tumor specimens stored by pathology laboratories.
In summary, mass spectrometry-based measurements of Krebs cycle metabolites in PPGLs provide a useful tool for identifying underlying SDHx mutations. The method also offers a window into assessing functionality of mutations; this not only has potential for stratifying risk for disease aggressiveness, but is also useful to assess functionality associated with gene variants of uncertain significance, thereby enabling damaging mutations to be distinguished from nonfunctional polymorphisms. Such a case has been described recently in a patient with PPGL, who carried a novel SDHD missense mutation subsequently identified as nonpathogenic (34). Furthermore, the method may be applicable to other neoplasms, such as GI stromal tumors and renal cell carcinomas that may occur due to mutations of SDHx or genes encoding other Krebs cycle enzymes (29, 35).
Acknowledgments
Spanish specimens were collected by hospitals through the Spanish National Tumor Bank Network (CNIO).
This work was funded by the European Union Seventh Framework Programme (FP7/2007-2013) under Grant Agreement No. 259735 (project ENS@T-Cancer) (S.R., M.P., N.Q., G.E., A.A.dC., M.R., E.R., M.M., J.U.R., H.J.T., F.B.), the Deutsche Forschungsgesellschaft (EI855/1-1) (S.R., M.P., N.Q., G.E., J.W.L.), the Fondo de Investigaciones Sanitarias (Project PI11/01359)(A.A.dC, M.R.), and the Eunice Kennedy Shriver National Institute of Child Health and Human Development (K.P.).
Disclosure Summary: The authors have nothing to disclose.
Footnotes
- FFPE
- formalin-fixed paraffin-embedded
- HNP
- head and neck paraganglioma
- MAX
- myc-associated factor X
- NF1
- neurofibromatosis type 1
- PPGLs
- pheochromocytomas and paragangliomas
- RET
- rearranged during transfection
- ROC
- receiver-operating characteristic
- TMEM127
- transmembrane protein 127
- VHL
- von Hippel-Lindau tumor suppressor.
References
- 1. Gimenez-Roqueplo AP, Dahia PL, Robledo M. An update on the genetics of paraganglioma, pheochromocytoma, and associated hereditary syndromes. Horm Metab Res. 2012;44:328–333 [DOI] [PubMed] [Google Scholar]
- 2. Gimenez-Roqueplo AP, Favier J, Rustin P, et al. Mutations in the SDHB gene are associated with extra-adrenal and/or malignant phaeochromocytomas. Cancer Res. 2003;63:5615–5621 [PubMed] [Google Scholar]
- 3. Eisenhofer G, Lenders JW, Siegert G, et al. Plasma methoxytyramine: a novel biomarker of metastatic pheochromocytoma and paraganglioma in relation to established risk factors of tumour size, location and SDHB mutation status. Eur J Cancer. 2012;48:1739–1749 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Ricketts C, Woodward ER, Killick P, et al. Germline SDHB mutations and familial renal cell carcinoma. J Natl Cancer Inst. 2008;100:1260–1262 [DOI] [PubMed] [Google Scholar]
- 5. Janeway KA, Kim SY, Lodish M, et al. Defects in succinate dehydrogenase in gastrointestinal stromal tumors lacking KIT and PDGFRA mutations. Proc Natl Acad Sci USA. 2011;108:314–318 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Eisenhofer G, Tischler AS, de Krijger RR. Diagnostic tests and biomarkers for pheochromocytoma and extra-adrenal paraganglioma: from routine laboratory methods to disease stratification. Endocr Pathol. 2012;23:4–14 [DOI] [PubMed] [Google Scholar]
- 7. Jafri M, Maher ER. The genetics of phaeochromocytoma: using clinical features to guide genetic testing. Eur J Endocrinol. 2012;166:151–158 [DOI] [PubMed] [Google Scholar]
- 8. Rattenberry E, Vialard L, Yeung A, et al. A comprehensive next generation sequencing-based genetic testing strategy to improve diagnosis of inherited pheochromocytoma and paraganglioma. J Clin Endocrinol Metab. 2013;98:E1248–1256 [DOI] [PubMed] [Google Scholar]
- 9. McWhinney SR, Pilarski RT, Forrester SR, et al. Large germline deletions of mitochondrial complex II subunits SDHB and SDHD in hereditary paraganglioma. J Clin Endocrinol Metab. 2004;89:5694–5699 [DOI] [PubMed] [Google Scholar]
- 10. Cascón A, Montero-Conde C, Ruiz-Llorente S, et al. Gross SDHB deletions in patients with paraganglioma detected by multiplex PCR: a possible hot spot? Genes Chromosomes Cancer. 2006;45:213–219 [DOI] [PubMed] [Google Scholar]
- 11. Jacob HJ. Next-generation sequencing for clinical diagnostics. N Engl J Med. 2013;369:1557–1558 [DOI] [PubMed] [Google Scholar]
- 12. Ward PS, Cross JR, Lu C, et al. Identification of additional IDH mutations associated with oncometabolite R(-)-2-hydroxyglutarate production. Oncogene. 2012;31:2491–2498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Choi C, Ganji SK, DeBerardinis RJ, et al. 2-hydroxyglutarate detection by magnetic resonance spectroscopy in IDH-mutated patients with gliomas. Nat Med. 2012;18:624–629 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. DiNardo CD, Propert KJ, Loren AW, et al. Serum 2-hydroxyglutarate levels predict isocitrate dehydrogenase mutations and clinical outcome in acute myeloid leukemia. Blood. 2013;121:4917–4924 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Pollard PJ, Ratcliffe PJ. Cancer. Puzzling patterns of predisposition. Science. 2009;324:192–194 [DOI] [PubMed] [Google Scholar]
- 16. Grossman AB. Where metabolism meets oncogenesis: another false lead? Lancet Oncol. 2010;11:309–310 [DOI] [PubMed] [Google Scholar]
- 17. Lendvai N, Pawlosky R, Bullova P, et al. Succinate-to-fumarate ratio as a new metabolic marker to detect the presence of SDHB/D-related paraganglioma: initial experimental and ex vivo findings. Endocrinology. 2014;155:27–32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Imperiale A, Moussallieh FM, Sebag F, et al. A new specific succinate-glutamate metabolomic hallmark in SDHx-related paragangliomas. PLoS One. 2013;8:e80539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kelly AD, Breitkopf SB, Yuan M, Goldsmith J, Spentzos D, Asara JM. Metabolomic profiling from formalin-fixed, paraffin-embedded tumor tissue using targeted LC/MS/MS: application in sarcoma. PLoS One. 2011;6:e25357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Peitzsch M, Pelzel D, Glöckner S, et al. Simultaneous liquid chromatography tandem mass spectrometric determination of urinary free metanephrines and catecholamines, with comparisons of free and deconjugated metabolites. Clin Chim Acta. 2013;418:50–58 [DOI] [PubMed] [Google Scholar]
- 21. Peitzsch M, Prejbisz A, Kroiß M, et al. Analysis of plasma 3-methoxytyramine, normetanephrine and metanephrine by ultraperformance liquid chromatography-tandem mass spectrometry: utility for diagnosis of dopamine-producing metastatic phaeochromocytoma. Ann Clin Biochem. 2013;50:147–155 [DOI] [PubMed] [Google Scholar]
- 22. Juratli TA, Peitzsch M, Geiger K, Schackert G, Eisenhofer G, Krex D. Accumulation of 2-hydroxyglutarate is not a biomarker for malignant progression in IDH-mutated low-grade gliomas. Neuro Oncol. 2013;15:682–690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Rapizzi E, Ercolino T, Canu L, et al. Mitochondrial function and content in pheochromocytoma/paraganglioma of succinate dehydrogenase mutation carriers. Endocr Relat Cancer. 2012;19:261–269 [DOI] [PubMed] [Google Scholar]
- 24. Capatina C, Ntali G, Karavitaki N, Grossman AB. The management of head-and-neck paragangliomas. Endocr Relat Cancer. 2013;20:R291–R305 [DOI] [PubMed] [Google Scholar]
- 25. Merlo A, de Quiros SB, Secades P, et al. Identification of a signaling axis HIF-1α/microRNA-210/ISCU independent of SDH mutation that defines a subgroup of head and neck paragangliomas. J Clin Endocrinol Metab. 2012;97:E2194–E2200 [DOI] [PubMed] [Google Scholar]
- 26. Tannahill GM, Curtis AM, Adamik J, et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature. 2013;496:238–242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Sciacovelli M, Guzzo G, Morello V, et al. The Mitochondrial Chaperone TRAP1 Promotes Neoplastic Growth by Inhibiting Succinate Dehydrogenase. Cell Metab. 2013;17:988–999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Letouzé E, Martinelli C, Loriot C, et al. SDH mutations establish a hypermethylator phenotype in paraganglioma. Cancer Cell. 2013;23:739–752 [DOI] [PubMed] [Google Scholar]
- 29. Pollard PJ, Brière JJ, Alam NA, et al. Accumulation of Krebs cycle intermediates and over-expression of HIF1alpha in tumours which result from germline FH and SDH mutations. Hum Mol Genet. 2005;14:2231–2239 [DOI] [PubMed] [Google Scholar]
- 30. Rao JU, Engelke UF, Rodenburg RJ, et al. Genotype-specific abnormalities in mitochondrial function associate with distinct profiles of energy metabolism and catecholamine content in pheochromocytoma and paraganglioma. Clin Cancer Res. 2013;19:3787–3795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Selak MA, Armour SM, MacKenzie ED, et al. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-alpha prolyl hydroxylase. Cancer Cell. 2005;7:77–85 [DOI] [PubMed] [Google Scholar]
- 32. Smith EH, Janknecht R, Maher LJ., 3rd Succinate inhibition of alpha-ketoglutarate-dependent enzymes in a yeast model of paraganglioma. Hum Mol Genet. 2007;16:3136–3148 [DOI] [PubMed] [Google Scholar]
- 33. Favier J, Briere JJ, Burnichon N, et al. The Warburg effect is genetically determined in inherited pheochromocytomas. PLoS One. 2009;4:e7094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Canu L, Rapizzi E, Zampetti B, et al. Pitfalls in genetic analysis of pheochromocytomas/paragangliomas-case report. J Clin Endocrinol Metab. 2014;99:2321–2326 [DOI] [PubMed] [Google Scholar]
- 35. O'Flaherty L, Adam J, Heather LC, et al. Dysregulation of hypoxia pathways in fumarate hydratase-deficient cells is independent of defective mitochondrial metabolism. Hum Mol Genet. 2010;19:3844–3851 [DOI] [PMC free article] [PubMed] [Google Scholar]