

Abstract
Socioeconomic and environmental influences are established factors promoting cancer disparity but the contribution of biological factors is not clear. We report a mechanistic link between carbohydrate derived metabolites and cancer which may provide a biological consequence of established factors of cancer disparity. Glycation is the non-enzymatic glycosylation of carbohydrates to macromolecules which produces reactive metabolites called advanced glycation end products (AGEs). A sedentary lifestyle and poor diet all promote disease and the AGE accumulation pool in our bodies and also increase cancer risk. We examined AGE metabolites in clinical specimens of African American and European American prostate cancer patients and found a higher AGE concentration in these specimens among African American patients when compared to European American patients. Elevated AGE levels corresponded with expression of the receptor for AGE (RAGE or AGER). We show that AGE mediated increases in cancer associated processes is dependent upon RAGE. Aberrant AGE accumulation may represent a metabolic susceptibility difference that contributes to cancer disparity.
Keywords: Glycation, Prostate Cancer, Cancer Disparity, Advanced Glycation End Product, Receptor for Advanced Glycation End Product, Immune response
Introduction
African American prostate cancer patients are more likely to develop aggressive prostate cancer and die of their disease than their European American counterparts (1-4). Poor diet and a lack of physical activity are potential risk factors for prostate cancer. Men who eat higher quantities of red meat and dairy products rather than fresh fruits and vegetables may have a higher chance of developing prostate cancer (5, 6). A healthy diet may also associate with improved clinical outcome and increased quality of life (6). While no association between physical activity and prostate cancer risk has yet been identified, research suggests that it may be associated with reduced risk of aggressive disease (7). Increased physical activity is also associated with increased survival in prostate cancer patients after diagnosis (8).
It is becoming increasingly apparent that molecular differences in tumor biology play a significant role in cancer disparity. In particular, established factors such as low income, poor diet and a sedentary lifestyle may confer molecular effects that alter cell signaling and gene expression patterns, including expression of non-coding RNAs, and effect chromosomal stability and therefore accelerate tumor development and progression (1-4). In other words our lifestyle choices may have profound effects on tumor development and progression which may contribute to health disparity outcomes such as the earlier development of cancer or the progression to more aggressive disease. Combined approaches to define the molecular consequences of the socioeconomic and environmental factors that drive health disparity may lead to the discovery of novel biomarkers indicative of disease risk and progression as well as avenues for treatment and prevention strategies.
Glycation represents a metabolic susceptibility difference that may provide a biological link between cancer and the socioeconomic and environmental factors that drive cancer health disparity. It occurs during normal metabolism and is the non-enzymatic glycosylation of sugar moieties to biological macromolecules such as protein and DNA. Glycation results in the accumulation of reactive metabolites known as advanced glycation end products (AGE’s) (9). Cellular clearance of AGE metabolites is inefficient and they accumulate in our tissues and organs as we grow older with pathogenic effects. AGEs contribute to diseases such as diabetes, cardiovascular disease, arthritis, and neurodegenerative disorders (9). Failure to remove these highly reactive metabolites can lead to protein damage, aberrant cell signaling, increased inflammatory responses, increased oxidative damage, changes in extracellular matrix composition and decreased genetic fidelity (10, 11). AGE functions as a ligand activator for RAGE which is overexpressed in a variety of tumor types including prostate (11). AGE activation of RAGE is functionally linked to an increased recruitment of immune cells in a broad range of diseases (11, 12). Its stimulation by AGE induces the transcriptional activation of NFkB, STAT3 and HIF1α which results in the expression of immune cytokines such as IL1, IL6 and TNFα (11, 12). Studies support a direct link between RAGE activation and proliferation, survival, migration, and invasion of tumor cells (11, 12). Blockade of RAGE suppresses tumor growth in two independent mouse models (11, 12) and loss of RAGE in inflammatory mouse models confers resistance to induced skin carcinogenesis (11, 12). In castrate sensitive and castrate resistant prostate cancer cell lines AGE was identified as the ligand for RAGE interactions but other ligands (S100B or amphoterin) were not (13). Significantly, apart from their endogenous production during normal metabolism, AGE accumulation is also associated with the socioeconomic and environmental factors that drive cancer health disparity. A sedentary lifestyle and a poor/unhealthy diet all contribute to health disparity, increase cancer risk and contribute to the AGE accumulation pool (14, 15). Consuming foods that are high in sugar/fat, heavily cooked or highly processed substantially increases the level of AGEs within our bodies (14, 15). AGE content in the so-called Western Diet has steadily increased over the last 50 years and it is now a significant contributor to the AGE accumulation pool and to the progression of many chronic diseases (14, 15). Additionally, lifestyle choices such as alcohol consumption and smoking also elevate AGE levels and human (16) and animal studies (17) show physical activity interventions can help maintain a stable AGE level or may even reduce it.
Due to the common links between the factors that drive cancer health disparity and the increased accumulation of AGE metabolites we examined AGE levels in serum and prostate tumor specimens and identified a potential race specific, tumor dependent pattern of AGE accumulation which may be indicative of disease progression.
Materials and Methods
Biological samples
Upon IRB approval, serum and tissue samples were obtained from the Hollings Cancer Center Tissue Biorepository here at the Medical University of South Carolina. Serum samples consisted of a cohort of 26 samples: 14 patients (7 African American, 7 European American) with low grade prostate cancer (Gleason 4-6) and 12 patients (5 African American, 7 European American) with high grade prostate cancer (Gleason 7-9). A subset of matching tissue sections (N=8, 2 per dataset) were also obtained for immunohistochemical (IHC) and immunofluorescent (IF) studies. Gleason grade was confirmed and IHC examined by a qualified pathologist.
AGE ELISA
To examine the levels of AGE in serum, 96-well format Oxi-select ELISA’s (Cell Biolabs, San Diego, CA) were used as directed by the manufacturer. All samples were normalized to total protein concentration in 1x phosphate buffered saline (PBS).
Immunohistochemistry and immunofluorescent (IF) tissue staining
All specimens were formalin-fixed and paraffin-embedded. Deparaffinized tissue sections were rehydrated, and antigen retrieval performed by heating in a vegetable steamer in 10 mM citrate, pH 6.0 for 30mins. Endogenous peroxidase activity was blocked using 0.3% H2O2 in methanol for 30mins. Sections were washed and nonspecific binding was blocked using 2.5% horse serum and then incubated overnight at 4°C with target-specific primary antibody at a 1:100 dilution. CML primary antibody (ab27684) was from Abcam (Cambridge, MA). RAGE primary antibody (AB5601) was from Millipore (Billerica, MA). Sections were fixed in Permount (Invitrogen) and mounted on slides. All sections were examined using an Olympus BX50 microscope and pictures were taken using an Olympus DP 70 camera connected to DP Controller software (Olympus, Center Valley, PA). IHC scores were calculated using the formula: intensity X % positive tumor cells. Intensity staining was scored: 1 – weak, 2 – intermediate, 3 – strong and 4 - very strong. % positive tumor cells were scored: 1 – less than 10%, 2 – 10 to 30%, 3 – 30 to 60% and 4 – 60 to 100% positive cells. Fluorescence was quantified using image J.
Statistical analysis
For statistical testing, two-sided Student’s t-tests were done using an Excel spreadsheet. p < 0.05 was considered statistically significant. Pearson Correlation analysis was used to examine the correlation between circulating AGE levels and those found in the tumor as well as between tumor AGE levels and RAGE expression in the tumor.
Results
Circulating AGE levels in prostate cancer patients
Levels of the AGE metabolite carboxymethyl-lysine (CML) were measured in serum and tissue from prostate cancer patients by ELISA. CML is extensively studied in animal models of disease, in epidemiological studies, and with regard to food content(18). It is often used as a marker of total AGEs in biological systems (14). Our analysis (Fig 1A) shows that circulating CML levels are significantly higher (p=0.0016) in serum from high grade prostate cancer patients (Gleason grade 7 to 10 (N=12)) compared to that observed in low grade prostate cancer patients (Gleason grade 4 to 6 (N=14)). African American men in the US are 2.5 times more likely to die of prostate cancer than their Caucasian counterparts(2). When the data was further stratified by self-reported race/ethnicity we found that compared to the European American (EA) prostate cancer patients (N=14), circulating AGE metabolite levels were significantly higher in serum from African American (AA) prostate cancer patients (N=12) (Fig 1B). This was observed in both the low grade (p=0.0003) and high grade (p=0.0015) serum (Fig 1C).
Figure 1. AGE levels are elevated in prostate cancer.
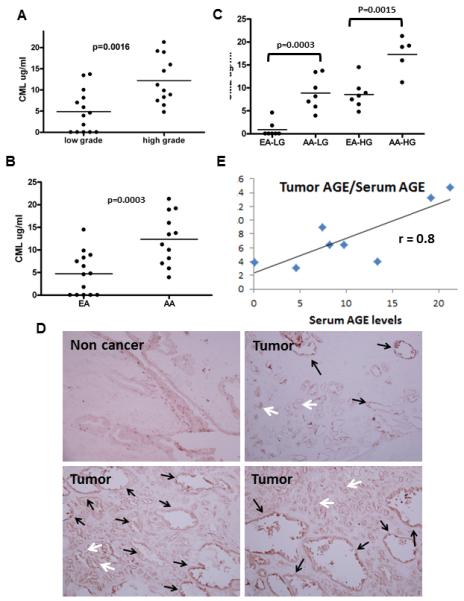
Quantification of circulating AGE (CML) levels, analyzed by ELISA, in serum from prostate cancer patients separated by A. grade (low = Gleason 4-6, high = Gleason 7-9) B. race (African American (AA) and European American (EA)) and C. race and grade. D. IHC staining (10X magnification) of AGE with an antibody raised against CML in tumor tissue (N=8) compared to non-cancer prostate tissue (N=4). Black arrows indicate the benign tissue and white arrows indicate the malignant tissue. Insets show 100 x magnifications. E. Pearson correlation analysis of tumor AGE and serum AGE levels.
Tumor AGE levels in prostate cancer patients
AGEs accumulate in our tissues as we grow older and promote numerous disease phenotypes (9, 19). To characterize AGE accumulation in prostate tumors we examined AGE levels in normal (N=4) and prostate cancer tissue (N=8) by IHC. Compared to the staining in the non-cancer tissue which represents AGE accumulation levels as a consequence of growing older, AGE metabolite staining was significantly higher in the benign (black arrows Fig 1D) and malignant (white arrows Fig 1D) tumor tissue. We compared AGE levels observed in the circulation to that observed in the tumor. In matched serum (N=4) and tumor (N=4) samples, analysis using the Pearson correlation (Fig 1E) shows a strong positive correlation between circulatory AGE levels and those observed in the tumor (r = 0.8).
To more readily quantify AGE levels in prostate tumors we examined CML accumulation in the same normal and prostate cancer tissue using IF staining. Compared to the levels observed in non-cancer prostate tissue, higher CML staining in the tumor tissue was confirmed (Fig 2A). Stratification of fluorescent intensity by Gleason grade and self-reported race shows that AAs with either low grade (AA-LG) or high grade (AA-HG) prostate cancer had significantly higher (p=0.005) CML levels than their European American counterparts (EA-LG; EA-HG) (Fig 2B). While AGE staining was evident in both the stromal and epithelial compartments, closer analysis of the fluorescent intensity shows that highest AGE accumulation was observed in the tumor epithelial cells with particular high intensity occurring at the cell membrane (Fig 2C).
Figure 2. AGE levels are highest in the epithelial tissue of high grade African American prostate cancer patients.

A. IF staining (10X magnification) of AGE with an antibody raised against CML in tumor tissue (N=8) compared to non-cancer prostate tissue (N=4). B. Quantification of the fluorescent intensity observed in 5 independent microscope fields of European American (EA) and African American (AA) patients with either low grade (LG) or high grade (HG) prostate cancer. C. Higher magnification (40x magnification) of the immunofluorescent staining in the tumor epithelial cells and the stroma.
RAGE expression in prostate cancer tissue
AGEs function as ligand activators for the transmembrane receptor for AGE (RAGE) which is overexpressed in a variety of tumor types (11). In diabetes and cardiovascular disease, RAGE stimulation by AGE induces the transcriptional activation of factors critical for immune response. Silencing of RAGE expression reduces prostate specific antigen expression and inhibits cell proliferation in prostate cancer cell lines and tumor growth in NUDE mice (20). Studies show that the V-domain of RAGE preferentially interacts with AGEs on prostate cancer cells (13) and that AGE treatment of prostate cancer cells induced both cell growth and invasion (21).
To characterize RAGE protein levels in normal (N=4) and prostate cancer tissue (N=8) we used IHC staining. Like AGE, compared to non-cancer prostate tissue, RAGE was significantly higher in the benign (black arrows Fig 3A) and malignant (white arrows Fig 3A) tumor tissue. Quantification of fluorescent staining shows that RAGE expression was higher in low grade versus high grade prostate cancer confirming published literature and that AAs with either low grade (LG) or high grade (HG) prostate cancer had significantly higher (p=0.005) RAGE levels than their European American counterparts (EA-LG; EA-HG) (Fig 3B). Upon analysis by Pearson correlation, AGE tumor levels showed a strong correlation (r = 0.6) to that of RAGE in the same tissue (Fig 3C).
Figure 3. RAGE levels are elevated in high grade and African American prostate cancer tissue.

A. IHC staining (10x magnification) of RAGE in prostate tumor (N=8) compared to non-cancer (N=4) tissue. B. IF staining and quantitation of RAGE expression in tumor tissue (N=8) compared to non-cancer prostate tissue (N=4). C. Pearson correlation analysis of tumor RAGE and tumor AGE levels.
Discussion
In a limited data set, our studies provide initial evidence that AGE accumulation may represent a novel metabolic susceptibility marker which may result from a mechanistic link between inflammation, cancer and factors promoting cancer disparity.
Both AGE accumulation and the Western lifestyle that contributes to the AGE accumulation pool (14, 15) are intrinsically linked with chronic systemic inflammation (11, 12). Experimental, clinical and epidemiological studies demonstrate that chronic inflammation can predispose to different types of cancer and contribute to its progression (22). A recent article in this journal reports that chronic inflammation in low risk benign prostate tissue was positively correlated with increased prostate cancer, especially in high-grade patients (23). The inflammatory environment caused by AGE mediated activation of the RAGE pathway could facilitate tumor formation and progression by promoting cancer associated processes such as ECM remodeling, angiogenesis and metastasis (11, 12). The significance of AGE accumulation as an inflammatory mediator that contributes to the excessive risk and mortality among African-American men may arise from the fact that poor diet and a lack of exercise are lifestyle factors particularly prevalent within African American communities who are also known to have and increased burden of inflammation associated chronic disease. Significantly, differential cytokine expression associated with an increased immune response was found to be a predominant pathway increased in African American prostate cancer patients (1). A similar race specific increase in immune response gene copy number and gene expression was also observed in matched radical prostatectomy tissues (24) and in Gleason 6 prostate tumors (4).
While the Western lifestyle is an exogenous source of AGE metabolite, the oncogenic re-programming of cellular metabolism may be a significant endogenous source in tumor cells. Tumor cells up-regulate glucose metabolism in order to provide the energy required to sustain accelerated cell growth and progression (i.e. the Warburg effect) (25). Similar to diabetes, AGEs produced by the Warburg effect have the potential to alter cell signaling pathways to increase intrinsic inflammation and promote cancer associated processes. Significantly, studies indicate that abnormal glucose uptake and the Warburg effect occurs earlier in African American women at high-risk of breast cancer and there is a growing link between the Warburg effect and the cancer immune response.
A caveat to this study is the small cohort of samples but the results provide compelling preliminary evidence to rationalize large cohort studies to define the full potential of AGE accumulation as a molecular biomarker in cancer and to dissect its functional contribution to cancer onset and progression. Given the potential benefits of lifestyle changes and the role of dietary-AGE in promoting disease phenotypes, opportunities exist for impacting cancer prevention initiatives arising through health and nutritional education as well as reducing cancer disparity.
Acknowledgements
Supported in part by the shRNA Technology Shared Resource, the Biostatistics Shared Resource (P30 CA138313), and the Biorepository Shared Resource, Hollings Cancer Center, Medical University of South Carolina.
Grant Support
This work was/is supported by grants from the NIH (P20 CA157071 to M.E. Ford & J.D. Salley; R21 CA176135 to D.P. Turner.
Footnotes
Disclosure of Potential Conflicts of Interest.
The authors declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
- 1.Kinseth MA, Jia Z, Rahmatpanah F, Sawyers A, Sutton M, Wang-Rodriguez J, et al. Expression differences between African American and Caucasian prostate cancer tissue reveals that stroma is the site of aggressive changes. International journal of cancer Journal international du cancer. 2014;134:81–91. doi: 10.1002/ijc.28326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Martin DN, Starks AM, Ambs S. Biological determinants of health disparities in prostate cancer. Current opinion in oncology. 2013;25:235–41. doi: 10.1097/CCO.0b013e32835eb5d1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Powell IJ, Bollig-Fischer A. Minireview: the molecular and genomic basis for prostate cancer health disparities. Molecular endocrinology. 2013;27:879–91. doi: 10.1210/me.2013-1039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Reams RR, Agrawal D, Davis MB, Yoder S, Odedina FT, Kumar N, et al. Microarray comparison of prostate tumor gene expression in African-American and Caucasian American males: a pilot project study. Infectious agents and cancer. 2009;4(Suppl 1):S3. doi: 10.1186/1750-9378-4-S1-S3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Research. WCRFAIfC . Food, Nutrition, Physical Activity, and the Prevention of Cancer: a Global Perspective. AICR; Washington DC: 2007. [Google Scholar]
- 6.Avery KN, Donovan JL, Horwood J, Neal DE, Hamdy FC, Parker C, et al. The importance of dietary change for men diagnosed with and at risk of prostate cancer: a multi-centre interview study with men, their partners and health professionals. BMC family practice. 2014;15:81. doi: 10.1186/1471-2296-15-81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Patel AV, Rodriguez C, Jacobs EJ, Solomon L, Thun MJ, Calle EE. Recreational physical activity and risk of prostate cancer in a large cohort of U.S. men. Cancer epidemiology, biomarkers & prevention : a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive Oncology. 2005;14:275–9. [PubMed] [Google Scholar]
- 8.Kenfield SA, Stampfer MJ, Giovannucci E, Chan JM. Physical activity and survival after prostate cancer diagnosis in the health professionals follow-up study. Journal of clinical oncology : official journal of the American Society of Clinical Oncology. 2011;29:726–32. doi: 10.1200/JCO.2010.31.5226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ansari NA, Rasheed Z. [Non-enzymatic glycation of proteins: from diabetes to cancer] Biomed Khim. 2010;56:168–78. doi: 10.18097/pbmc20105602168. [DOI] [PubMed] [Google Scholar]
- 10.Duran-Jimenez B, Dobler D, Moffatt S, Rabbani N, Streuli CH, Thornalley PJ, et al. Advanced glycation end products in extracellular matrix proteins contribute to the failure of sensory nerve regeneration in diabetes. Diabetes. 2009;58:2893–903. doi: 10.2337/db09-0320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Riehl A, Nemeth J, Angel P, Hess J. The receptor RAGE: Bridging inflammation and cancer. Cell Commun Signal. 2009;7:12. doi: 10.1186/1478-811X-7-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rojas A, Gonzalez I, Morales E, Perez-Castro R, Romero J, Figueroa H. Diabetes and cancer: Looking at the multiligand/RAGE axis. World J Diabetes. 2011;2:108–13. doi: 10.4239/wjd.v2.i7.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Allmen EU, Koch M, Fritz G, Legler DF. V domain of RAGE interacts with AGEs on prostate carcinoma cells. Prostate. 2008;68:748–58. doi: 10.1002/pros.20736. [DOI] [PubMed] [Google Scholar]
- 14.Uribarri J, Woodruff S, Goodman S, Cai W, Chen X, Pyzik R, et al. Advanced glycation end products in foods and a practical guide to their reduction in the diet. J Am Diet Assoc. 2010;110:911–16. doi: 10.1016/j.jada.2010.03.018. e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Vlassara H. Advanced glycation in health and disease: role of the modern environment. Ann N Y Acad Sci. 2005;1043:452–60. doi: 10.1196/annals.1333.051. [DOI] [PubMed] [Google Scholar]
- 16.Yoshikawa T, Miyazaki A, Fujimoto S. Decrease in serum levels of advanced glycation end-products by short-term lifestyle modification in non-diabetic middle-aged females. Medical science monitor : international medical journal of experimental and clinical research. 2009;15:PH65–73. [PubMed] [Google Scholar]
- 17.Boor P, Celec P, Behuliak M, Grancic P, Kebis A, Kukan M, et al. Regular moderate exercise reduces advanced glycation and ameliorates early diabetic nephropathy in obese Zucker rats. Metabolism: clinical and experimental. 2009;58:1669–77. doi: 10.1016/j.metabol.2009.05.025. [DOI] [PubMed] [Google Scholar]
- 18.Ames JM. Determination of N epsilon-(carboxymethyl)lysine in foods and related systems. Annals of the New York Academy of Sciences. 2008;1126:20–4. doi: 10.1196/annals.1433.030. [DOI] [PubMed] [Google Scholar]
- 19.Singh R, Barden A, Mori T, Beilin L. Advanced glycation end-products: a review. Diabetologia. 2001;44:129–46. doi: 10.1007/s001250051591. [DOI] [PubMed] [Google Scholar]
- 20.Elangovan I, Thirugnanam S, Chen A, Zheng G, Bosland MC, Kajdacsy-Balla A, et al. Targeting receptor for advanced glycation end products (RAGE) expression induces apoptosis and inhibits prostate tumor growth. Biochem Biophys Res Commun. 2012;417:1133–8. doi: 10.1016/j.bbrc.2011.12.060. [DOI] [PubMed] [Google Scholar]
- 21.Ishiguro H, Nakaigawa N, Miyoshi Y, Fujinami K, Kubota Y, Uemura H. Receptor for advanced glycation end products (RAGE) and its ligand, amphoterin are overexpressed and associated with prostate cancer development. The Prostate. 2005;64:92–100. doi: 10.1002/pros.20219. [DOI] [PubMed] [Google Scholar]
- 22.Lu H, Ouyang W, Huang C. Inflammation, a key event in cancer development. Molecular cancer research : MCR. 2006;4:221–33. doi: 10.1158/1541-7786.MCR-05-0261. [DOI] [PubMed] [Google Scholar]
- 23.Gurel B, Lucia MS, Thompson IM, Jr., Goodman PJ, Tangen CM, Kristal AR, et al. Chronic inflammation in benign prostate tissue is associated with high-grade prostate cancer in the placebo arm of the prostate cancer prevention trial. Cancer Epidemiol Biomarkers Prev. 2014;23:847–56. doi: 10.1158/1055-9965.EPI-13-1126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rose AE, Satagopan JM, Oddoux C, Zhou Q, Xu R, Olshen AB, et al. Copy number and gene expression differences between African American and Caucasian American prostate cancer. Journal of translational medicine. 2010;8:70. doi: 10.1186/1479-5876-8-70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nat Rev Cancer. 2011;11:85–95. doi: 10.1038/nrc2981. [DOI] [PubMed] [Google Scholar]