

Abstract
Global change is expected to modify the frequency and magnitude of defoliating insect outbreaks in forest ecosystems. Bats are increasingly acknowledged as effective biocontrol agents for pest insect populations. However, a better understanding is required of whether and how bat communities contribute to the resilience of forests to man- and climate-driven biotic disturbances. We studied the responses of forest insectivorous bats to a major pine defoliator, the pine processionary moth pityocampa, which is currently expanding its range in response to global warming. We used pheromone traps and ultrasound bat recorders to estimate the abundance and activity of moths and predatory bats along the edge of infested pine stands. We used synthetic pheromone to evaluate the effects of experimentally increased moth availability on bat foraging activity. We also evaluated the top-down regulation of moth population by estimating T. pityocampa larval colonies abundance on the same edges the following winter. We observed a close spatio-temporal matching between emergent moths and foraging bats, with bat activity significantly increasing with moth abundance. The foraging activity of some bat species was significantly higher near pheromone lures, i.e. in areas of expected increased prey availability. Furthermore moth reproductive success significantly decreased with increasing bat activity during the flight period of adult moths. These findings suggest that bats, at least in condition of low prey density, exhibit numerical and functional responses to a specific and abundant prey, which may ultimately result in an effective top-down regulation of the population of the prey. These observations are consistent with bats being useful agents for the biocontrol of insect pest populations in plantation forests.
Introduction
Predator-prey relationships are shaped by the functional and numerical responses of the predators to prey density [1]. The numerical response involves predator density being adjusted to prey abundance through changes in reproduction, dispersal and foraging behaviour leading to aggregative patterns in habitat patches with large feeding resources [2]. The functional response is the adjustment of the predator consumption rate to the abundance or the biomass of its feed so that the consumption rate is a major determinant of the top-down regulation of prey population by predators [3], [4]. Insect pests are a major threat to forests worldwide and it is expected that climate change will further enhance insect herbivory, due to positive response of forest insects to warmer and drier conditions [5], [6]. Climate warming is also likely to trigger both more intense and severe insect outbreaks, and range expansion northwards and upwards [7]–[9]. The pine processionary moth Thaumetopoea pityocampa Denis & Schiffermüller (Lepidoptera: Notodontidae) is the main defoliator of pines (Pinus spp) in the western part of the Mediterranean Basin, including Southern Europe, the Balkans and North Africa. Defoliation by this species significantly reduces tree growth, and severe or repeated defoliation can lead to tree death [10]. The distribution range of T. pityocampa is currently expanding towards higher latitudes and elevations as a result of the release of thermal constraints allowing improved winter survival and feeding activity [5], [11].
Predators and parasitoids contribute to shaping the cyclic population dynamics of T. pityocampa [12], [13]. However, recent studies have also suggested that climate warming leads to more stable top down regulation involving generalist predators rather than specialist predators [1], [14], [15]. Birds have long been considered as the only predatory vertebrates that are efficient pest regulators, although recent comparative studies in tropical forests have shown that insect predation by bats may be more significant than predation by birds [16]–[18]. Using mostly echolocation, insectivorous bats are nocturnal predators preying on invertebrates both at forest edges and within forest stands. Using diverse hunting techniques they can catch a wide range of arthropod prey and consume over half of their body mass in insects nightly [19]. Given that many herbivorous insects are mainly active at night [20], it is therefore likely that chiropterans contribute greatly to pest regulation [21]. However, the pest regulation service provided by bats remains to be quantified, notably for the control of those forest pests that represent a growing concern under climate change [18]. A pioneer review on the biological control of forest insects by vertebrates pointed out the lack of information on numerical response of predators to prey density, a key issue to better estimate predation effectiveness [22]. Since this seminal work, few if any studies on vertebrate predators have managed to fully address these critical questions [21]. Even if they revealed a significant impact of bat predation on insect population, all previous studies have failed to identify which prey insect species was actually concerned [21].
Species-specific prey consumption and foraging observations have been performed to study bats as insect predators but these two approaches have never been used simultaneously. Few studies to date have actually considered bat activity as a numerical response to insect prey availability [23]–[25]. Even fewer studies have investigated the functional responses of bats to insect abundance via dietary and faeces analyses [26], [27]. Here, in an innovative experimental approach, we used synthetic sex pheromone lures to manipulate the local availability of a specific prey species, the pine processionary moth (Thaumetopoea pityocampa), without modifying local environmental conditions, by contrast with the use of exclosures [21] or light attraction [28]. This method allowed investigating, in natural field conditions, the responses of bats to experimentally controlled prey availability. In this manipulative experiment, we formulated the following three hypotheses for bat predation on T. pityocampa: (i) the occurrence and abundance of predatory bats will match in time and space with those of the main insect prey species; (ii) predatory bats will increase their foraging activities when and where prey is more abundant, i.e. exhibiting positive numerical and functional responses to prey density; and (iii) insect prey abundance will decrease at the next generation where bat foraging activity has been higher, due to consumption of adult moths and consequent reduction of reproductive success.
Materials and Methods
Study area and site selection
The study area was located in the Landes de Gascogne forest (N44 45.000, W0 50.00), in south-western France; this forest is the largest forest plantation in Europe with approximately one million ha of pure stands of the maritime pine Pinus pinaster. The climate is thermo-Atlantic and the soil is podzolic. We selected 23 pine plantations of the same age (30 years old) with similar edges, at least 200 m long, and separated by at least 900 m. Edges in front of clearcuts or very young stands were avoided. The Office National des Forêts and Groupama provided us special authorizations to access their private stands and conduct the study.
Moth surveys
The field experiment was conducted in July 2012 during the period of peak T. pityocampa moth emergence [29]. Synthetic sex pheromone trapping is considered to be an efficient method to monitor T. pityocampa populations [30]. It relies on a positive relationship between pheromone trap captures and local population density estimated through counts of larval colonies [30]. After two nights of bat sampling (Figure 1), we replaced bat detectors with pheromone-baited traps at the same sites along the selected forest edges. These traps were 30×30 cm, plate sticky traps with both sides covered with glue, baited with 0.5 mg of pityolure [30]. Traps were activated for six consecutive nights. At the end of the trapping session, the sum of male captures was recorded for each trap to estimate the abundance of T. pityocampa in each plot.
Figure 1. Experimental set up established in a subset of 12 pine plantation edges.

In late February 2013, we estimated the density of T. pityocampa larval colonies along the same sampled stand edges in order to quantify the effect of adult moth predation by bats on the abundance of prey offspring. We counted all larval colonies on all pine trees along 100 m of stand edge, focusing on the first two rows of the stand where most of T. pityocampa larval colonies are concentrated [31]. We calculated the ratio of the number of larval colonies per stand edge divided by the number of male catches on the same edge as an indicator of moth reproductive success (because the sex-ratio is 1∶1 in T. pityocampa, the number of male captures approximated the number of pairs). Two stands were discarded because they had been thinned in between male trapping and larval colonies counting (for a total of N = 21 stands). Time patterns of T. pityocampa moth activity per night period were adapted from the detailed biological study of Demolin (1969).
Bat surveys
Along the edge of each stand, one automatic ultrasound bat detector system (Sound Meter SM2BAT, Wildlife Acoustics) fitted with multidirectional microphones (SMX-US weatherproof ultrasonic microphone, Wildlife Acoustics) was installed to record night bat activity. Detectors were calibrated to detect all bat calls and programmed to record from one hour before sunset to one hour after sunrise. Each edge was sampled for two consecutive nights between July 9 and July 27, 2012. Recordings were included in the analysis only for nights without rain, when the wind was <30 km/h and the ambient temperature above 10°C [32]. In addition, we studied a subset of 12 forest edges where two bat detectors, separated by 100 m, were deployed: each of the two detectors was equipped with a T. pityocampa sex pheromone dispenser on alternate nights (Figure 1). The synthetic pheromone was used to attract and increase availability of flying male moths around the microphone of the bat detector.
Bat calls were identified by one trained operator (YC) using Batsound 4.1. We used existing identification keys and published data [33]–[35]. All sequences were analyzed and identified to the finest taxonomic level possible: species level identification was feasible for various kinds of calls, but there were large overlaps between some species making species level identification impossible. In the study area Pipistrellus kuhlii and P. nathusii calls were really similar. However, in our case, they were only allocated to Pipistrellus kuhlii because this species is very common and widespread within the study area, whereas P. nathusii is very rare according to regional atlas data [36] and it was never detected by diagnostic calls in our own survey. By contrast, using a conservative approach, we decided not to discriminate Eptesicus serotinus and Nyctalus leisleri and classified them as a single sonotype [37]. Both species were recorded with certainty (5012 calls for E. serotinus and 459 calls for N. leisleri) and both species are commonly distributed in the study area. Nevertheless it seems that serotine bats E. serotinus are far more abundant than lesser noctules N. leisleri in pine plantation forests at the regional level [36].
We assessed bat activity levels using the number of search phase sequences for all species or sonotype. These sequences were composed of two or more pulse calls separated from other calls by one second or more [37], [38]. To better evaluate bat activity, continuous sequences longer than 5 s were scored as two sequences. We also assessed bat feeding activity by counting the number of feeding buzz calls indicating active prey capture attempts by a foraging individual bat. Compared to regular calls, feeding buzz calls were defined as more steeply frequency-modulated with pulse intervals gradually decreasing [39].
Statistical analyses
We used Generalized Linear Mixed Models (GLMM) [40] implemented in R-package lme4 [41] to analyze the complete data set of 23 edges during all sampled nights and thereby assess the effects of T. pityocampa abundance on bat activity. The response variable was the number of call sequences during each sampled night. The explanatory variable was T. pityocampa abundance. We accounted for the hierarchical structure of the data by adding nested random effects of night, plot and hour to the model intercept. Residual repartition and deviation from normality (normal Q–Q) interpretation plots were used for model validation. We also tested the individual effect of the presence of a T. pityocampa pheromone dispenser on bat activity, prey capture attempts and bat species richness. We paired echolocation data recorded with and without a pheromone lure, during the same night along the same edge in 12 forest stands, using a permutation t-test implemented in the R-package Deducer [42]. We log-transformed the ratio of larval colonies/male moth captures to meet the assumption that residuals are normally distributed. Thereby we used linear models to test for the effect of overall bat activity or bat feeding activity on moth reproductive success.
Results
Temporal patterns of bat and moth activity
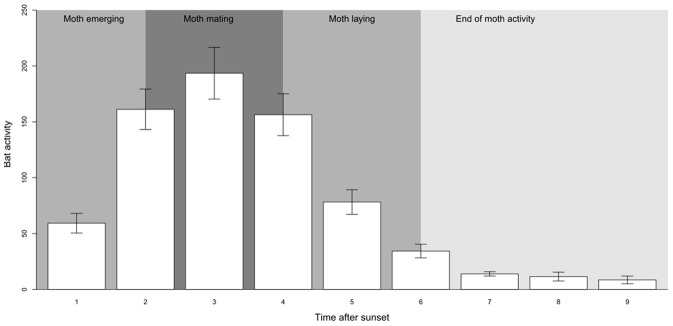
We recorded 49271 passes identifiable to species or sonotype level. They include 27997 (56.9% of the identifiable passes) P. kuhlii passes, 17300 (35.1%) E. serotinus - N. leisleri sonotype passes, and 3015 (6.1%) P. pipistrellus passes. The other 1.8% were assigned to Myotis spp (341 passes), Plecotus spp (299), Nyctalus noctula (109), Barbastella barbastellus (94), Nyctalus lasiopterus (13) and Rhinolophus ferrumequinum (1). The maximum activity of bats along the 23 sampled edges matched the period of pine processionary moth mating activities, which occur during the 4 hours after sunset (Figure 2). Total bat activity and total feeding buzzes per night were significantly correlated (R2 = 0.47; P<0.0001) and these two variables showed the same temporal patterns throughout the night. Finally, bat activity rhythm, all species together, matched the described time pattern activity of T. pityocampa.
Figure 2. Compared periods of nocturnal activities for bats and pine processionary moths (adapted from Demolin 1969) along the 23 sampled forest edges.
Bat response to moth abundance
We caught 409 male moths in the 46 traps (mean 9 moths per trap). The mean capture per trap and per plot ranged from 1 to 22 suggesting that our experimental design fitted a range of T. pityocampa densities. For bat data analysis, we only used data from the first 4 hours of the night, i.e. when bat activity was maximal, in order to limit bias and to avoid zero-inflated counts. Total bat activity significantly increased with increasing moth abundance on forest edge (z = 17.27; P<0.0001) (Table 1). The activity of several bat species also significantly increased with local moth abundance: P. kuhlii (z = 18.41; P<0.0001), E. serotinus - N. leisleri sonotype (z = 4.154; P<0.0001) and P. pipistrellus (z = 7.684; P<0.0001) (Table 1). There was no effect of moth abundance on bat species diversity (z = 1.438; P = 0.151).
Table 1. Results of Poisson GLMMs linking total bat activity and individual species activities to pine processionary moth abundance.
| Bat species | Estimates | SE | z-value | P |
| Total bat activity | 0.0611 | 0.0035 | 17.27 | <0.0001 |
| Pipistrellus kuhlii | 0.0973 | 0.0052 | 18.41 | <0.0001 |
| Eptesicus serotinus - Nyctalus leisleri | 0.0215 | 0.0051 | 4.15 | <0.0001 |
| Pipistrellus pipistrellus | 0.0909 | 0.0118 | 7.68 | <0.0001 |
| Plecotus spp | −0.0580 | 0.0426 | −1.36 | 0.1732 |
| Barbastella barbastellus | 0.0127 | 0.0691 | 0.18 | 0.8550 |
Bat response to moth aggregates
For a given edge within a given night, we did not observe any significant effect of the presence of pheromone lure on total bat activity (P = 0.558), or on the activities of P. kuhlii (P = 0.871), E. serotinus - N. leisleri sonotype (P = 0.422) and P. pipistrellus (P = 0.510). Artificial moth aggregates, resulting from pheromone attraction, had also no significant effect on bat species richness (P = 0.856). However, even if bat activities remained the same along the edge, there was a significant increase in bat feeding activity near the pheromone lure, i.e. close to artificial moth aggregates (P<0.001; mean of differences = −13.58, see Figure 3). The largest bat species emitted significantly more feeding buzzes in the presence of pheromone lure (E. serotinus - N. leisleri sonotype; P = 0.013; mean of differences = −2.08; and P. kuhlii; P = 0.001; mean of differences = −9.62), while it was not the case for the smallest species P. pipistrellus (P = 0.214).
Figure 3. Comparison of prey capture attempts (mean number of buzzes+SE) for total and bat species groups in the presence (black bars) vs. in absence (white bars) of a T. pityocampa sex pheromone lure.

Bat predation effect on moth demography
There was a positive correlation between male moth abundance as estimated by pheromone trap catches and larval colonies abundance the next year along the same forest edges (R2 = 0.245; P = 0.013). More interestingly, the ratio of larval colonies per trapped males, a measure of fertility, significantly decreased with increasing total bat activity (R2 = 0.335; P = 0.003, see Figure 4) and the activity of the P. kuhlii (R2 = 0.166; P = 0.047) and the E. serotinus - N. leisleri sonotype (R2 = 0.418; P<0.001). By contrast, the activity of P. pipistrellus had no significant effect on moth abundance (R2 = 0.014; P = 0.270). Bat feeding activity did not show a significant effect on prey reproductive success (R2 = 0.102; P = 0.087). The ratio of larval colonies per trapped males significantly decreased with increasing E. serotinus - N. leisleri sonotype feeding activity (R2 = 0.391; P = 0.001).
Figure 4. Effect of total bat activity on prey reproductive success (ratio of larval colonies per male moth captured) the following summer along 21 sampled edges.

Discussion
The magnitude of arthropod consumption by bats changes along their reproductive cycle. In Europe, most of the bat species give birth in late spring or early summer [43]. In common with other mammals, lactation required a substantial energy expenditure [27], [44]. Female bats therefore have to optimize their hunting activities when lactating. In the study area, bats lactate in late June and July, and this is also the period when the emergence of T. pityocampa peaks. In early summer, T. pityocampa is the most abundant moth species in pine plantation forests (Charbonnier unpub. data), moreover showing a circadian rhythm in tight coincidence with bat activity (Figure 2). Furthermore, this temporal synchronization may be coupled with spatial matching due to the forest edge preference of both bats [23], [45], [46] and T. pityocampa adult moths [47]. Previous studies have shown that insectivorous bat activities are strongly correlated with arthropod abundance, suggesting that bats actively search for areas of concentrated prey resources [23], [25], [48]. In our study, we found that bat activity increased with prey availability (moth abundance, Table 1). This finding provides support for our initial hypothesis of significant bat numerical responses to T. pityocampa abundance. Our results are consistent with previous studies showing that insectivorous bats are able to adjust their predatory activity to prey availability [26], [27]. This behaviour may favour optimal foraging on resources that are unpredictable in time and space, such as local outbreaks of T. pityocampa within large pine plantations. Moreover, as the period of T. pityocampa emergence coincides with the period of highest energy requirements for female bats, it is possible that bats have also a demographic response to the moth. Further experiments are needed to test this hypothesis, as observed for another predatory vertebrate, the Eurasian hoopoe Upupa epops [49].
McCracken et al. [27], by including comparisons of bats’ diet vs. potential prey abundance in a large study area, suggested that there were bat functional responses to particular types of prey at the landscape scale. Here, without changing environmental conditions, we artificially increased the availability of one prey species in its favoured habitat using specific sexual pheromones. These lures may have several effects besides local attraction of males by increasing either abundance or flight activity in intensity and time. Introducing synthetic lures can shift or extend the flight period of male moths, so that increased bat capture attempts could result from the longer time window when bats are able to find the moths. This increase of prey availability, due to an increasing encounter probability, actually resulted in a significantly higher bat feeding activity (Fig. 2). This suggests that the more T. pityocampa moths are present on a given edge, the more foraging bats are able to feed on them. Without changing their overall flight activity at the local scale of a given forest edge, but only by enhancing prey capture attempts in relation to prey availability, bats seemed to display a functional response to this specific insect prey. We were unable to determine the exact shape of the functional response curve, because we used a proxy of feeding activity (buzz numbers) rather than true prey capture attempts per individual bat. Previous studies showed that Holling type II (cyrtoid) responses are the most frequent for vertebrates [50]. Nevertheless, bats can travel long distances, use different foraging tactics and feed on several prey, so it is more likely that the bat functional response is a Holling type III (sigmoid) response [51]. This type of functional response is mostly associated with generalist predators because they are able to switch between alternative prey items according to availability. Therefore, these bat populations may remain fairly abundant when T. pityocampa availability declines, and respond rapidly when the pine defoliator build up its populations.
Only the largest bat species showed a significant functional response. The E. serotinus - N. leisleri and P. kuhlii displayed significantly more prey capture attempts in the presence of higher moth availability. By contrast, the smaller P. pipistrellus did not produce more buzzes in the presence of a pheromone dispenser and thus moth availability. This species may be too small to feed on T. pityocampa and, according to the optimal foraging theory, would spend too much time handling this kind of prey [52]. Various other bat species specialize on moths, such as Barbastella barbastellus [53], [54] or Plecotus spp [55], but they did not show either numerical or functional responses to higher densities of moths in our study. These species can fly slowly and accurately to glean foliage-resting moths in dense vegetation [56] within old pine and mixed forests [57]; this may explain their lack of response to the experimentally generated flying moth along forest edges. Another explanation could be the low level of T. pityocampa populations during the experiment. Generalist species are known to have greater impact on prey populations at low density whereas specialized predators are more effective with high prey density [58], [59]. Here, moth density may have been too low to trigger a functional response by a bat specialist. In addition, if fruit bats commonly use olfactory cues to find their food, such behaviour is much less frequent in insectivorous bats, which use odour for social interactions rather than for foraging [60], [61]. It is therefore unlikely that the higher bat feeding activity recorded near sex pheromone lures was a direct response to prey odour (kairomonal attraction). Bats are polyphagous predators and there is no evidence that any are sufficiently specialized to have evolved the capacity to smell insect pheromones.
Jactel et al. [30] found a positive relationship between the abundance of T. pityocampa male moths trapped and the numbers of larval colonies in the next generation. In our study, we observed a significant and negative correlation between general bat activity during the flight period of male processionary moths (time period of the experiment) and the ratio of larval colonies (the next winter) per male moth trapped along the same forest edge. This finding clearly suggests that bat numerical and functional responses to moth density resulted in less offspring per adult moth. It is therefore most likely that bats were feeding on T. pityocampa moths, reducing the reproductive success of the insect species. This prediction is strengthened by low predation and parasitism rates of egg masses in the study area [62], which is the only intermediate life stage between laying moths and larval colonies. Nevertheless, these results are preliminary and would require a longer period of monitoring to be generalized. Several records of bats caught in pine processionary pheromone traps (Martin, unpublished data) and visual observation of bats feeding (e.g. [63]; Serra-Cobo, unpublished data) also suggest that several bat species can actively prey on T. pityocampa moths. However only an in depth study of bats diet in pine forests, for example based on DNA analysis of droppings, will allow to formally confirm this assumption. Moreover, monitoring the effects of bat predation on moth demography during an entire epidemic cycle would allow testing the hypothesis that top down regulation is less effective during pest outbreaks [58], [59]. Such an impact of predation on adult moths is critical for the effectiveness of top down regulation of the pest insect because this stage of the biological cycle is the smallest in size [64]. In T. pityocampa, many other biological control agents including predatory vertebrates (birds) have been described for all larval and pupal development stages, but few before for adult moths [12], [13].
It is increasingly acknowledged that bats may contribute substantially to pest regulation in temperate agro-ecosystems [21]. However, the reasons for this effectiveness are not fully understood. Insectivorous bats are an example of generalist predators that maintain constant vital rates and stable populations by shifting to alternative prey [1], [14]. Even if our study was limited to a single year, our new experimental approach based on artificial increase in prey availability using pheromone lures revealed a facilitating mechanism of foraging plasticity: the ability of bats to detect and concentrate on local aggregates of the most abundant prey species. These numerical and functional responses of generalist bat species may result in a negative effect on the reproductive success of the prey and then in a reduction of the population growth rate, at least in condition of low prey density. These various characteristics make bats potential biological control agents that could contribute to regulate T. pityocampa populations.
Acknowledgments
We are grateful to Groupama and the Office National des Forêts for the permission to access their sites. We thank Michel Renou and Aaron J. Corcoran for their helpful comments on earlier versions of the manuscript. We also acknowledge Inge van Halder, Julien Nezan, Fabrice Vetillard, Pierre Gaüzère, Jean-Yves Barnagaud, Bastien Castagneyrol, Dominique Piou, Eliomys and the Groupe Chiroptères d’Aquitaine for their helps and advices.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.
Funding Statement
The research reported here was conducted as part of the European FunDiv EUROPE project, which received funding from the European Commission’s Seventh Framework Program under the grant agreement no 265171. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Bretagnolle V, Gillis H (2010) Predator-prey interactions and climate change. Effects of Climate Change on Birds. Oxford University Press. 227–247.
- 2. Goss-Custard JD (1970) The responses of redshank (Tringa totanus (L.)) to spatial variations in the density of their prey. Journal of Animal Ecology 39: 91–113 10.2307/2891 [DOI] [Google Scholar]
- 3. Abrams PA, Ginzburg LR (2000) The nature of predation: prey dependent, ratio dependent or neither? Trends in Ecology & Evolution 15: 337–341 10.1016/S0169-5347(00)01908-X [DOI] [PubMed] [Google Scholar]
- 4.Glen DM (2004) Birds as predators of lepidopterous larvae. Insect and bird interactions. Andover, Hampshire, UK: Intercept.
- 5. Robinet C, Roques A (2010) Direct impacts of recent climate warming on insect populations. Integrative Zoology 5: 132–142 10.1111/j.1749-4877.2010.00196.x [DOI] [PubMed] [Google Scholar]
- 6. Jactel H, Petit J, Desprez-Loustau M-L, Delzon S, Piou D, et al. (2012) Drought effects on damage by forest insects and pathogens: a meta-analysis. Global Change Biology 18: 267–276 10.1111/j.1365-2486.2011.02512.x [DOI] [Google Scholar]
- 7. Battisti A, Stastny M, Buffo E, Larsson S (2006) A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Global Change Biology 12: 662–671 10.1111/j.1365-2486.2006.01124.x [DOI] [Google Scholar]
- 8. Hódar JA, Zamora R, Cayuela L (2012) Climate change and the incidence of a forest pest in Mediterranean ecosystems: can the North Atlantic Oscillation be used as a predictor? Climatic Change 113: 699–711 10.1007/s10584-011-0371-7 [DOI] [Google Scholar]
- 9.Klapwik MJ, Ayres MP, Battisti A, Larsson S (2012) Assessing the impact of climate change on outbreak potential. Insect Outbreaks Revisited. John Wiley & Sons. 429–450.
- 10. Jacquet J–S, Orazio C, Jactel H (2012) Defoliation by processionary moth significantly reduces tree growth: a quantitative review. Annals of Forest Science 69: 857–866 10.1007/s13595-012-0209-0 [DOI] [Google Scholar]
- 11. Battisti A, Stastny M, Netherer S, Robinet C, Schopf A, et al. (2005) Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol Appl 15: 2084–2096 10.1890/04-1903 [DOI] [Google Scholar]
- 12. Battisti A (1989) Field studies on the behaviour of two egg parasitoids of the pine processionary mothThaumetopoea pityocampa. Entomophaga 34: 29–38 10.1007/BF02372584 [DOI] [Google Scholar]
- 13. Barbaro L, Battisti A (2011) Birds as predators of the pine processionary moth (Lepidoptera: Notodontidae). Biological Control 56: 107–114 10.1016/j.biocontrol.2010.10.009 [DOI] [Google Scholar]
- 14. De Sassi C, Tylianakis JM (2012) Climate change disproportionately increases herbivore over plant or parasitoid biomass. PLoS ONE 7: e40557 10.1371/journal.pone.0040557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Twardochleb LA, Novak M, Moore JW (2012) Using the functional response of a consumer to predict biotic resistance to invasive prey. Ecol Appl 22: 1162–1171. [DOI] [PubMed] [Google Scholar]
- 16. Kalka MB, Smith AR, Kalko EKV (2008) Bats limit arthropods and herbivory in a tropical forest. Science 320: 71–71 10.1126/science.1153352 [DOI] [PubMed] [Google Scholar]
- 17. Williams-Guillen K, Perfecto I, Vandermeer J (2008) Bats limit insects in a neotropical agroforestry system. Science 320: 70–70 10.1126/science.1152944 [DOI] [PubMed] [Google Scholar]
- 18. Böhm SM, Wells K, Kalko EKV (2011) Top-Down Control of Herbivory by Birds and Bats in the Canopy of Temperate Broad-Leaved Oaks (Quercus robur). PLoS ONE 6: e17857 10.1371/journal.pone.0017857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kalka M, Kalko E (2006) Gleaning bats as underestimated predators of herbivorous insects: diet of Micronycteris microtis (Phyllostomidae) in Panama. J Trop Ecol 22: 1–10 10.1017/S0266467405002920 [DOI] [Google Scholar]
- 20. Rydell J, Entwistle A, Racey PA (1996) Timing of foraging flights of three species of bats in relation to insect activity and predation risk. Oikos 76: 243–252 10.2307/3546196 [DOI] [Google Scholar]
- 21. Kunz TH, Braun de Torrez E, Bauer D, Lobova T, Fleming TH (2011) Ecosystem services provided by bats. Annals of the New York Academy of Sciences 1223: 1–38 10.1111/j.1749-6632.2011.06004.x [DOI] [PubMed] [Google Scholar]
- 22. Buckner CH (1966) The Role of Vertebrate Predators in the Biological Control of Forest Insects. Annual Review of Entomology 11: 449–470 10.1146/annurev.en.11.010166.002313 [DOI] [PubMed] [Google Scholar]
- 23. Müller J, Mehr M, Bässler C, Fenton M, Hothorn T, et al. (2012) Aggregative response in bats: prey abundance versus habitat. Oecologia 169: 673–684 10.1007/s00442-011-2247-y [DOI] [PubMed] [Google Scholar]
- 24. Dodd LE, Chapman EG, Harwood JD, Lacki MJ, Rieske LK (2012) Identification of prey of Myotis septentrionalis using DNA-based techniques. Journal of Mammalogy 93: 1119–1128 10.1644/11-MAMM-A-218.1 [DOI] [Google Scholar]
- 25. Fukui D, Murakami M, Nakano S, Aoi T (2006) Effect of emergent aquatic insects on bat foraging in a riparian forest. Journal of Animal Ecology 75: 1252–1258 10.1111/j.1365-2656.2006.01146.x [DOI] [PubMed] [Google Scholar]
- 26. Agosta SJ, Morton D, Kuhn KM (2003) Feeding ecology of the bat Eptesicus fuscus: “preferred” prey abundance as one factor influencing prey selection and diet breadth. J Zool 260: 169–177 10.1017/S0952836903003601 [DOI] [Google Scholar]
- 27. McCracken GF, Westbrook JK, Brown VA, Eldridge M, Federico P, et al. (2012) Bats track and exploit changes in insect pest populations. PLoS ONE 7: e43839 10.1371/journal.pone.0043839 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Stone EL, Jones G, Harris S (2009) Street Lighting Disturbs Commuting Bats. Current Biology 19: 1123–1127 10.1016/j.cub.2009.05.058 [DOI] [PubMed] [Google Scholar]
- 29. Démolin G (1969) Comportement des adultes de Thaumetopoea pityocampa Schiff. Dispersion spatiale, importance écologique. Annales des Sciences Forestières 26: 81–102 10.1051/forest/19690104 [DOI] [Google Scholar]
- 30. Jactel H, Menassieu P, Vétillard F, Barthélémy B, Piou D, et al. (2006) Population monitoring of the pine processionary moth (Lepidoptera: Thaumetopoeidae) with pheromone-baited traps. Forest Ecology and Management 235: 96–106 10.1016/j.foreco.2006.08.002 [DOI] [Google Scholar]
- 31. Dulaurent A–M, Porté AJ, van Halder I, Vétillard F, Menassieu P, et al. (2011) A case of habitat complementation in forest pests: Pine processionary moth pupae survive better in open areas. Forest Ecology and Management 261: 1069–1076 10.1016/j.foreco.2010.12.029 [DOI] [Google Scholar]
- 32.Parsons K, Bat Conservation Trust (2007) Bat surveys: good practice guidelines. London: Bat Conservation Trust.
- 33.Barataud M, Tupinier Y (2012) Écologie acoustique des chiroptères d’Europe identification des espèces, étude de leurs habitats et comportements de chasse. Mèze; Paris: Biotope; Muséum national d’histoire naturelle.
- 34.Russ J (2012) British bat calls: a guide to species identification. Exeter: Pelagic Pub.
- 35. Russo D, Jones G (2002) Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls. Journal of Zoology 258: 91–103 10.1017/S0952836902001231 [DOI] [Google Scholar]
- 36.Ruys T, Bernard Y (2014) Atlas des mammifères sauvages d’Aquitaine - Les Chiroptères. C. Nature. Le Haillan. 256 p. [Google Scholar]
- 37. Jung K, Kaiser S, Böhm S, Nieschulze J, Kalko EKV (2012) Moving in three dimensions: effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. Journal of Applied Ecology 49: 523–531 10.1111/j.1365-2664.2012.02116.x [DOI] [Google Scholar]
- 38. Kalcounis MC, Hobson KA, Brigham RM, Hecker KR (1999) Bat activity in the boreal forest: Importance of stand type and vertical strata. J Mammal 80: 673–682 10.2307/1383311 [DOI] [Google Scholar]
- 39. Schnitzler H–U, Kalko EKV (2001) Echolocation by Insect-Eating Bats. BioScience 51: 557–569 10.1641/0006-3568(2001)0510557:EBIEB2.0.CO2 [DOI] [Google Scholar]
- 40. Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, et al. (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology & Evolution 24: 127–135 10.1016/j.tree.2008.10.008 [DOI] [PubMed] [Google Scholar]
- 41.Bates D, Maechler M, Bolker B (2012) Linear mixed model implementation in lme4.
- 42.Fellows I (2012) Package “Deducer.”
- 43.Dietz C, Nill D, Helversen O von (2009) Bats of Britain, Europe and Northwest Africa. London: A & C Black.
- 44.Kunz T, Stern A (1995) Maternal investment and post-natal growth in bats London: Zoological Society of London and the Mammal Society.
- 45. Morris AD, Miller DA, Kalcounis-Rûpell MC (2010) Use of forest edges by bats in a managed pine forest landscape. The Journal of Wildlife Management 74: 26–34 10.2193/2008-471 [DOI] [Google Scholar]
- 46.Jantzen MK, Fenton MB (2013) The depth of edge influence among insectivorous bats at forest-field interfaces. Canadian Journal of Zoology: 130311043958007. doi:10.1139/cjz-2012-0282.
- 47. Dulaurent A–M, Porté AJ, van Halder I, Vétillard F, Menassieu P, et al. (2012) Hide and seek in forests: colonization by the pine processionary moth is impeded by the presence of nonhost trees. Agricultural and Forest Entomology 14: 19–27 10.1111/j.1461-9563.2011.00549.x [DOI] [Google Scholar]
- 48. Hagen EM, Sabo JL (2012) Influence of river drying and insect availability on bat activity along the San Pedro River, Arizona (USA). Journal of Arid Environments 84: 1–8 10.1016/j.jaridenv.2012.03.007 [DOI] [Google Scholar]
- 49. Barbaro L, Couzi L, Bretagnolle V, Nezan J, Vetillard F (2008) Multi-scale habitat selection and foraging ecology of the eurasian hoopoe (Upupa epops) in pine plantations. Biodivers Conserv 17: 1073–1087 10.1007/s10531-007-9241-z [DOI] [Google Scholar]
- 50. Klinger R, Rejmánek M (2009) The numerical and functional responses of a granivorous rodent and the fate of neotropical tree seeds. Ecology 90: 1549–1563 10.1890/07-2146.1 [DOI] [PubMed] [Google Scholar]
- 51. Jeschke JM, Kopp M, Tollrian R (2004) Consumer-food systems: why type I functional responses are exclusive to filter feeders. Biological Reviews 79: 337–349 10.1017/S1464793103006286 [DOI] [PubMed] [Google Scholar]
- 52. Fossette S, Gleiss AC, Casey JP, Lewis AR, Hays GC (2012) Does prey size matter? Novel observations of feeding in the leatherback turtle (Dermochelys coriacea) allow a test of predator-prey size relationships. Biol Lett 8: 351–354 10.1098/rsbl.2011.0965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Sierro A, Arlettaz R (1997) Barbastelle bats (Barbastella spp.) specialize in the predation of moths: Implications for foraging tactics and conservation. Acta Oecol-Int J Ecol 18: 91–106 10.1016/S1146-609X(97)80067-7 [DOI] [Google Scholar]
- 54. Andreas M, Reiter A, Benda P (2011) Prey Selection and Seasonal Diet Changes in the Western Barbastelle Bat (Barbastella barbastellus). Acta Chiropterologica 14: 81–92 10.3161/150811012X654295 [DOI] [Google Scholar]
- 55. Ashrafi S, Beck A, Rutishauser M, Arlettaz R, Bontadina F (2011) Trophic niche partitioning of cryptic species of long-eared bats in Switzerland: implications for conservation. European Journal of Wildlife Research 57: 843–849 10.1007/s10344-011-0496-z [DOI] [Google Scholar]
- 56. Fenton MB (1990) The foraging behavior and ecology of animal-eating bats. Can J Zool-Rev Can Zool 68: 411–422 10.1139/z90-061 [DOI] [Google Scholar]
- 57. Sierro A (1999) Habitat selection by barbastelle bats (Barbastella barbastellus) in the Swiss Alps (Valais). Journal of Zoology 248: 429–432 10.1111/j.1469-7998.1999.tb01042.x [DOI] [Google Scholar]
- 58. Nixon AE, Roland J (2012) Generalist predation on forest tent caterpillar varies with forest stand composition: an experimental study across multiple life stages. Ecol Entomol 37: 13–23 10.1111/j.1365-2311.2011.01330.x [DOI] [Google Scholar]
- 59. Symondson WOC, Sunderland KD, Greenstone MH (2002) Can Generalist Predators Be Effective Biocontrol Agents? Annu Rev Entomol 47: 561–594 10.1146/annurev.ento.47.091201.145240 [DOI] [PubMed] [Google Scholar]
- 60. Bloss J (1999) Olfaction and the use of chemical signals in bats. Acta chiropterologica 1: 31–45. [Google Scholar]
- 61.Kunz TH, Fenton BB (2005) Bat ecology. Chicago, Ill.: University of Chicago Press.
- 62. Castagneyrol B, Jactel H, Charbonnier Y, Barbaro L, Dulaurent-Mercadal A-M (2014) Egg mortality in the pine processionary moth: habitat diversity, microclimate and predation effects. Agric For Entomol 16: 284–292 10.1111/afe.12056 [DOI] [Google Scholar]
- 63.Gonzales Cano JM (1981) Predacion de procesionaria del pino por vertebrados en la zona de Mora de Rubielos (Teruel). Boletin de la Estacion Central de Ecologia: 53–57.
- 64. Dempster JP (1983) The Natural Control of Populations of Butterflies and Moths. Biological Reviews 58: 461–481 10.1111/j.1469-185X.1983.tb00396.x [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.