

Abstract
Morphogenetic transitions of the opportunistic fungal pathogen Candida albicans are influenced by temperature changes, with induction of filamentation upon a shift from 30 to 37°C. Hsp90 was identified as a major repressor of an elongated cell morphology at low temperatures, as treatment with specific inhibitors of Hsp90 results in elongated growth forms at 30°C. Elongated growth resulting from a compromised Hsp90 is considered neither hyphal nor pseudohyphal growth. It has been reported that ascorbic acid (vitamin C) interferes with the yeast-to-hypha transition in C. albicans. In the present study, we show that ascorbic acid also antagonizes the morphogenetic change caused by hampered Hsp90 function. Further analysis revealed that Upc2, a transcriptional regulator of genes involved in ergosterol biosynthesis, and Erg11, the target of azole antifungals, whose expression is in turn regulated by Upc2, are required for this antagonism. Ergosterol levels correlate with elongated growth and are reduced in cells treated with the Hsp90 inhibitor geldanamycin (GdA) and restored by cotreatment with ascorbic acid. In addition, we show that Upc2 appears to be required for ascorbic acid-mediated inhibition of the antifungal activity of fluconazole. These results identify Upc2 as a major regulator of ascorbic acid-induced effects in C. albicans and suggest an association between ergosterol content and elongated growth upon Hsp90 compromise.
INTRODUCTION
Candida albicans is among the few fungal species that are frequently encountered as clinical pathogens (1). C. albicans is a frequent commensal of healthy individuals, but when the host immune system is weakened, it can become a deadly pathogen (2). Its pathogenic nature relies on a multitude of factors, including the ability to grow at 37°C, to produce adhesins and hydrolases, and to change between different morphological forms, including yeast cells, hyphae, pseudohyphae, chlamydospores, opaque cells, and the recently described GUT morphology (3–5). It is generally assumed that the yeast form is crucial for fungal dissemination throughout the body via the bloodstream (6), while hyphae, on the other hand, are required for the production of extracellular enzymes and invasive growth (7).
The yeast-to-hypha transition of C. albicans can be triggered by environmental cues, such as pH, serum, elevated temperature, and CO2 (4, 8). In addition, several small molecules, such as autoregulatory molecules, cell cycle inhibitors, and histone deacetylase inhibitors, are also capable of modulating morphogenetic responses (9, 10). Most environmental triggers and small molecules function through activation of the cyclic AMP (cAMP)-protein kinase A (PKA) or mitogen-activated protein kinase (MAPK) pathway via their downstream transcription factors, Efg1 and Cph1, respectively (4, 11, 12). Over the past few years, several reports have appeared that showed that geldanamycin (GdA), a benzoquinone ansamycin antibiotic, is another small molecule that strongly affects the cell shape of C. albicans at 30°C, as its addition results in elongated cells. GdA inhibits the function of Hsp90 by binding to its ADP/ATP binding pocket (10, 13), indicating that Hsp90 prevents an elongated cell shape of C. albicans at lower temperatures. This morphogenetic process also involves the Ras1-cAMP-PKA signaling pathway, but it seems to do this independently of the downstream transcription factor Efg1, suggesting the involvement of other transcriptional regulators (13). Screening of a transcription factor (TF) deletion collection for mutants impaired in the Hsp90-mediated elongated-cell-shape phenotype resulted in the identification of Hms1 (14). Two upstream regulators of this TF, the cyclin-dependent kinase Pho85 and the cyclin Pcl1, are also required for elongated cell shape (14). As Hms1 is not a component of the cAMP-PKA pathway, the link between Hsp90-dependent elongated cell shape and the Ras1-PKA signaling pathway remained to be identified. Our aim in this work was to identify transcription factors that upon overexpression affect the Hsp90-dependent elongated cell shape. For this, we screened a TF overexpression library and found that overexpression of Upc2 prevents the elongated-growth phenotype when GdA is added. The Zn2Cys6 transcriptional regulator Upc2 is important for the regulation of the ergosterol biosynthetic pathway in response to commonly used antifungals (15). The transcription factor can act as a repressor or an activator depending on its target and on the initiating conditions and performs its function by binding to sterol response elements (SREs) (15–17). Gain-of-function mutations in UPC2 have been associated with resistance to antifungal treatments (18–21), while strains devoid of Upc2 are hypersusceptible to azole antifungals (22). One of the genes regulated by Upc2 is Erg11, which is a key enzyme in the biosynthesis of ergosterol, and it is the target of the azole antifungal drugs. Overexpression of Erg11 has been shown to cause tolerance for these drugs.
In order to find components linking the GdA-induced elongated-growth phenotype and the block of this phenotype upon overexpression of UPC2, we tested a number of compounds that were shown to mediate morphogenesis in C. albicans. Ascorbic acid (vitamin C) was previously shown to play a role in morphogenesis, as it blocks serum-initiated hypha formation, a process mediated by adenylate cyclase (23, 24). How ascorbic acid affects morphogenesis is not clear, and we propose that it may function by modulating the role of Hsp90 in cell shape formation. In this paper, we describe a previously unreported negative effect of ascorbic acid on the Hsp90-dependent cell shape. Focusing on the mode of action of ascorbic acid, we provide evidence that it requires Upc2 and Erg11 to perform its function. We further demonstrate that intracellular ergosterol levels play a role in the ascorbic acid-mediated effect on cell shape. In addition, we show that Upc2 is also required for the antagonistic effect of ascorbic acid on fluconazole toxicity. Together, these results show that ascorbic acid inhibits Hsp90-mediated cell shape transition via the transcriptional regulator Upc2.
MATERIALS AND METHODS
Strains and growth conditions.
The strains used in this study are listed in Table 1. C. albicans strains were grown overnight in 3 ml of YPD medium (1% yeast extract, 2% bacteriological peptone, and 2% glucose) at 30°C. The cells were subsequently diluted to an optical density at 600 nm (OD600) of 0.2 and cultured for the indicated times at 30°C with the indicated treatments (4 μM or 10 μM geldanamycin, 2.5 mM ascorbic acid, l-cysteine, d-cysteine, glutathione, dithiothreitol [DTT], or 0.1 μg/ml doxycycline). When ascorbic acid was added, the medium was buffered to pH 7. All chemicals were purchased from Sigma-Aldrich.
TABLE 1.
Strains used in this study
| Strain | Description | Genotype | Source |
|---|---|---|---|
| SC5314 | Wild type | 70 | |
| CAI4 | ura3Δ::imm434/ura3Δ::imm434 iro1Δ/iro1Δ::imm343 | 71 | |
| AFA60a | CAI4 transformed with URA3 vector | ura3Δ::imm434/ura3Δ::imm434 iro1Δ/iro1Δ::imm343 RPS10::CIp10 | 42 |
| AFA59b | CAI4 carrying extra copy of CaERG11 under the control of ACT1 promoter | ura3Δ::imm434/ura3Δ::imm434 iro1Δ/iro1Δ::imm343 RPS10::pAFC89b | 42 |
| DSY448 | cdr1Δ/cdr1Δ mutant | ura3Δ::imm434/ura3Δ::imm434 iro1Δ/iro1Δ::imm343 cdr1Δ::hisG-URA3-hisG/cdr1Δ::hisG | 72 |
| DSY465 | mdr1Δ/mdr1Δ mutant | ura3Δ::imm434/ura3Δ::imm434 iro1Δ/iro1Δ::imm343 benΔ::hisG-URA3-hisG/benΔ::hisG | 72 |
| CMDR1E2A and -B | CAI4 carrying CaMDR1 under the control of ADH1 promoter | ura3Δ::imm434/ura3Δ::imm434 iro1Δ/iro1Δ::imm343 ADH1/adh1::PADH1-MDR1-URA3 | 39 |
| SN152 | Background strain TF wild type and TF077 | arg4Δ/arg4Δ leu2Δ/leu2Δ his1Δ/his1Δ URA3/ura3Δ::imm434 IRO1/iro1Δ::imm343 | 73 |
| TF wild type | Control wild-type strain (paired “wild-type” TF077) | arg4Δ/arg4Δ leu2Δ/LEU2 his1Δ/HIS1 URA3/ura3Δ::imm434 IRO1/iro1Δ::imm343 | 31 |
| TF077 | upc2Δ/upc2Δ mutant | As SN152, but upc2Δ::HIS1/upc2Δ::LEU2 | 31 |
| BWP17 (TW14901) | Background strain D-6 and EC-7 | ura3Δ::imm434/ura3Δ::imm434 iro1/iro1Δ::imm434 his1Δ::hisG/his1Δ::hisG arg4Δ/arg4Δ | 22 |
| D-6 (TW14903) | upc2Δ/upc2Δ mutant | As BWP17 but upc2Δ::URA3/upc2Δ::ARG4 | 22 |
| EC-7 (TW14904) | Reconstituted strain | As D-6 but upc2Δ::URA3/upc2Δ::UPC2-HIS1 | 22 |
| pTET-ERG11 | CAI4 with doxycycline-repressible ERG11 as the only ERG11 allele | As CAI4, but tetO-ERG11/erg11Δ | 41 |
| CaLC1411 (CaLC436) | Strain with doxycycline-repressible HSP90 as the only HSP90 allele | arg4Δ/arg4Δ his1Δ/his1Δ URA3/ura3Δ::imm434 IRO1/iro1Δ::imm434 HIS1/his1Δ::TAR-FRT hsp90Δ::CdHIS1/his1Δ::TAR-FRT | 74 |
| SC2H3 | One- and two-hybrid reporter strain in the SN152 background | Like SN152, but 5 × LexAOp-ADH1b/HIS1 5 × LexAOp-ADH1b/lacZa | 27 |
| OX control | Strain carrying empty vector under MET3 promoter | As SC2H3, but MET3pro::lexA | Stynen et al., unpublished |
| Upc2 OX | Strain carrying extra copy of UPC2 under MET3 promoter | As SC2H3, but MET3pro::lexA::UPC2 | Stynen et al., unpublished |
| Gcn4 OX | Strain carrying extra copy of GCN4 under MET3 promoter | As SC2H3, but MET3pro::lexA::GCN4 | Stynen et al., unpublished |
| Cap1 OX | Strain carrying extra copy of CAP1 under MET3 promoter | As SC2H3, but MET3pro::lexA::CAP1 | Stynen et al., unpublished |
| Dot6 OX | Strain carrying extra copy of DOT6 under MET3 promoter | As SC2H3, but MET3pro::lexA:DOT6 | Stynen et al., unpublished |
| Rtg3 OX | Strain carrying extra copy of RTG3 under MET3 promoter | As SC2H3, but MET3pro::lexA::RTG3 | Stynen et al., unpublished |
This is SN152, but it contains two reporter genes (HIS1 and LacZ), and the expression of these reporter genes is controlled by a promoter element consisting of 5 times the LexA operator sequence and the ADH1 basic promoter sequence.
Transcription factor overexpression strains were grown in complete supplement mixture minus methionine (CSM−Met medium) [0.073% CSM−Met−Ura, 0.17% yeast nitrogen base (YNB) without amino acids and (NH4)2SO4, 0.5% (NH4)2SO4, pH 5.5] supplemented with 0.1% uridine and the indicated treatment (10 μM GdA or 2.5 or 10 mM methionine).
Microscopy.
Imaging of the cells was done by differential interference contrast microscopy using the Zeiss Axioplan 2 microscope. Images were obtained with a Zeiss Axiocam MRc5 camera using Axiovision software 3.0 (Carl Zeiss, Inc., NY).
Cell sedimentation assay.
The cell sedimentation rate was quantified using the assay described by Eboigbodin and Biggs (25) and Fu et al. (26). Cells were grown in YPD liquid medium with or without 0.1 μg/ml doxycycline (for transcriptional repression) and 2.5 mM l-cysteine or ascorbic acid. The OD600 of the cells in the upper part of the glass tube was determined at the indicated time points, as sediments settle to the bottom of the glass tube. The formula (ODI − ODt/ODI) × 100, where ODI is the initial OD taken at time zero and ODt is the OD taken at the indicated time point, was utilized to calculate the percentage of sedimented cells.
Upc2 overexpression strain: transcription factor library.
The UPC2 overexpression strain used in this study is part of a transcription factor overexpression library constructed in our laboratory on the basis of the previously developed C. albicans two-hybrid system (27). Briefly, the nuclear localization sequence (NLS) was removed from the one-hybrid plasmid pC2HB, resulting in pC1H. Subsequently, genes encoding known and putative transcription factors were successfully cloned in fusion with the DNA-binding domain of LexA in the plasmid to create the pC1H-PTF library. The resulting plasmids were integrated into the two-hybrid reporter strain S2CH3 (27) between XOG1 and HOL1 loci on chromosome 1 after linearization at the NotI restriction site. Selection was performed on CSM−Leu medium. The complete library consists of ∼200 strains, each carrying an ectopically expressed protein under the control of an inducible MET3 promoter (B. Stynen and P. Van Dijck, unpublished data). The use of the inducible MET3 promoter allows conditional expression, as it is repressed in the presence of methionine and cysteine (28).
Ergosterol extraction and quantification.
Ergosterol extraction and quantification were performed as reported by Arthington-Skaggs et al. (29), with minor modifications. Briefly, cells were grown overnight in 4 ml YPD medium at 30°C. The cells were diluted to an OD600 of 0.2 and grown in YPD medium supplemented with 10 μM GdA, 2.5 mM ascorbic acid, or a combination of both. After the indicated time, 220 ODs of cells at 600 nm was used in the heptane extraction. The same procedure was used for all experimental conditions. Cells were harvested by centrifugation at 3,000 rpm for 5 min and washed once with 10 ml sterile water. Three milliliters of 25% alcoholic potassium hydroxide solution (25 g KOH and 35 ml sterile H2O brought to 100 ml with 100% ethanol) was added to each pellet and vortexed for 1 min. The cell suspensions were heated in an 85°C water bath for 1 h and then allowed to cool down to room temperature. Sterols were extracted with a mixture of 1 ml of sterile water and 3 ml of n-heptane, followed by mixing for 3 min. The organic layer was transferred to a clean glass tube and stored at −20°C for 24 h. Prior to the spectrophotometrical scan at between 230 and 300 nm using a Shimadzu UV-1650PC spectrophotometer, the sterol extracts were diluted 5-fold in 100% ethanol. A dilution of heptane and ethanol was used as the blank. Ergosterol levels were subsequently calculated as a percentage of the wet weight, as described previously (29).
Quantitative real-time PCR.
Overnight cultures of C. albicans strains were diluted to an OD600 of 0.2, and cultures were grown at 30°C for the indicated times in the presence and absence of either doxycycline (0.1 μg/ml), ascorbic acid (2.5 mM), or GdA (10 μM) before RNA extraction. RNA was DNase treated prior to cDNA synthesis with the Promega A3500 reverse transcription kit. Quantitative real-time PCR was performed using the GoScript Reverse Transcription System (Promega) on a StepOne Plus real-time PCR system (Applied Biosystems). Reactions were performed in triplicate, with oligonucleotides CaERG11up and CaERG11down, UPC2_fw and UPC2_rv, HSP90_fw and HSP90_rv, and TEF1a-fw and TEF1a-rv as the primer pairs (Table 2). The degree of regulation was determined by the ΔΔCT method, using expression of TEF1 to normalize the data.
TABLE 2.
Primers used in this study
| Primer | Sequence | Source |
|---|---|---|
| CaERG11up | TTACCTCATTATTGGAGACGTGATG | 42 |
| CaERG11down | CACGTTCTCTTCTCAGTTTAATTTCTTTC | 42 |
| TEF1a_fw | CCACTGAAGTCAAGTCCGTTGA | 42 |
| TEF1a-rv | CACCTTCAGCCAATTGTTCGT | 42 |
| UPC2_fw | GGATGCTCGACATGCAAAAAG | This study |
| UPC2_rv | TGCCACATACAGGTCTCTGTTCA | This study |
| HSP90_fw | CCATCTGATATCACTCAAGATG | This study |
| HSP90_rv | AGTGATAAACACTCTACGGACG | This study |
Checkerboard assay.
A checkerboard assay was performed on 96-well plates by combining different concentrations of ascorbic acid and fluconazole, and the fractional inhibitory concentration index (FICI) was determined. The assay was performed with both the wild type and the UPC2 deletion mutant to verify whether the effect of ascorbic acid on fluconazole susceptibility is exerted via Upc2.
The concentrations of ascorbic acid ranged from 0.078 mM to 5 mM for both the wild type and the UPC2 deletion mutant. The concentration of fluconazole ranged from 0.004 μg/ml to 2 μg/ml for the UPC2 deletion mutant and 0.125 μg/ml to 64 μg/ml for the wild type. The FICI was calculated according to the following formula: FICindex = FICA + FICB = (MICA comb/MICA alone) + (MICB comb/MICB alone), where MICA alone and MICB alone are the MICs of ascorbic acid and fluconazole alone and MICA comb and MICB comb are the MICs of ascorbic acid and fluconazole in combination. Interactions were referred to as synergistic when the FICindex value was ≤0.5 and antagonistic when the FICindex value was >4. FICindex values that were >0.5 or ≤4 indicated no interaction between ascorbic acid and fluconazole (30). There was no difference in FIC indexes observed between 24 and 48 h.
Oxidative-stress sensitivity assay.
The wild type and the two independent UPC2 deletion mutants of the Homann collection (31) were grown to mid-log phase in liquid YPD medium. The cells were diluted to an OD600 of 1, and 10-fold serial dilutions were spotted on YPD plates containing the indicated concentrations of H2O2. The plates were photographed after 24-h incubation at 37°C.
Propidium iodide staining.
Viability staining was performed by incubating a subset of cells (taken at the indicated time points of the experiments) for 20 min in the dark with the fluorescent stain propidium iodide (PI) (46 mM; Sigma-Aldrich). Prior to PI fluorescence analysis (excitation and emission maxima, 535 and 617 nm), the cells were washed and resuspended in phosphate-buffered saline (PBS).
Statistical analysis.
Statistical significance was calculated using a Student t test analysis. All experiments were performed with at least three biological repeats.
RESULTS
Ascorbic acid interferes with the morphogenetic process governed by Hsp90.
The transition of C. albicans from yeast to hyphae can be induced or repressed by a wide variety of triggers (32). Partial inhibition of Hsp90, obtained either by addition of GdA or by decreased transcription of the corresponding HSP90 gene, results in elongated cells, which are considered neither hyphae nor pseudohyphae (13). In order to understand the mechanism by which Hsp90 affects morphogenesis, we tested a number of compounds known to inhibit filamentation for interaction with GdA and determined the effect of ascorbic acid on the GdA-induced elongated growth. Addition of ascorbic acid, which was previously reported to interfere with the yeast-to-hypha transition (24), resulted in a strong reduction of the GdA-induced elongated cell shape (Fig. 1A). Similar results were obtained using radicicol, an Hsp90 inhibitor structurally distinct from GdA (reference 33 and data not shown). Since Hsp90 is an essential chaperone, cell viability was assessed with PI, a dye that is excluded from viable cells. As shown in Fig. 1B, the viability of GdA-treated cells is similar to that of untreated cells, ruling out possible effects due to elevated cell mortality.
FIG 1.
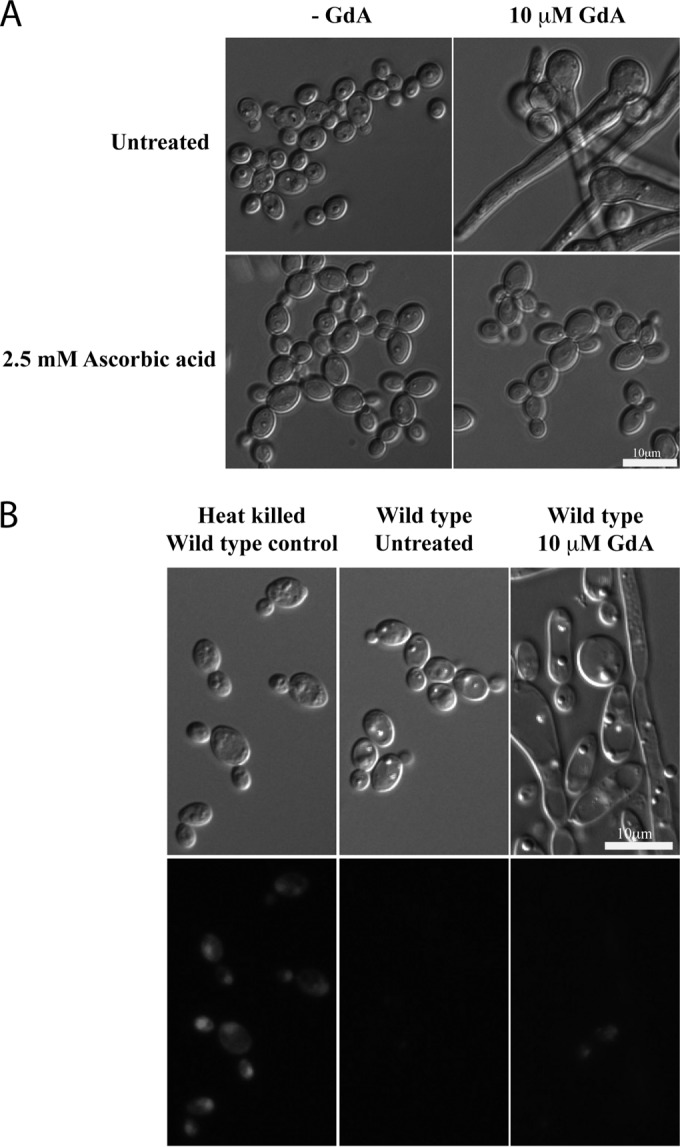
Ascorbic acid abrogates GdA-induced elongated growth. (A) Inhibiting effect of ascorbic acid on elongated growth induced by 10 μM GdA. Wild-type SC5314 cells were grown at 30°C in liquid rich medium containing GdA (10 μM) with 2.5 mM ascorbic acid. The images were taken after 8 h of growth. (B) GdA-mediated pharmacological inhibition of Hsp90 for extended periods is not lethal. Propidium iodide-stained heat-killed wild-type cells (15 min at 80°C) clearly illustrate a complete lethal phenotype, as indicated by the staining (which is excluded from viable cells), while the GdA-treated cells closely resemble the untreated wild-type cells. Top, DIC images; bottom, propidium iodide fluorescence images. The images were taken after 24 h.
To test whether the observed effect of ascorbic acid was dependent on Hsp90 or the result of chemical-chemical interference with GdA, similar experiments were conducted using a strain whose Hsp90 content can be modulated via the use of a tetracycline-repressible promoter (13). As expected, lowering the expression of HSP90 with doxycycline in this strain resulted in elongated growth (Fig. 2A). Cell viability tests were also performed and indicated that the cells were alive (Fig. 2B). Addition of ascorbic acid to cells genetically depleted of Hsp90 greatly increased the percentage of yeast cells (75%) compared to the control conditions without the addition of ascorbic acid (32%) (Fig. 2C). The effect of ascorbic acid was also determined using a sedimentation assay. Addition of ascorbic acid to cells in a liquid culture resulted in a lower sedimentation rate, indicating a higher proportion of yeast cells in these cultures than in non-ascorbic-acid-treated cells (Fig. 2D). Since transcription of HSP90 in the tetO-HSP90 strain is reduced in the presence of doxycycline, the possibility of a doxycycline-ascorbic acid chemical interaction was ruled out in experiments with wild-type control cells. Here, the concentration of doxycycline used in our experiments was demonstrated not to interfere with the effect of ascorbic acid on the GdA-dependent elongated cell shape described above (see Fig. S1A in the supplemental material). We also confirmed that doxycycline-mediated repression of Hsp90 was not significantly altered in the presence of ascorbic acid (see Fig. S1B in the supplemental material). Taken together, these results indicate that ascorbic acid has a negative effect on elongated cell shape formation resulting from lower Hsp90 activity or expression.
FIG 2.

Ascorbic acid influences phenotypes caused by Hsp90 genetic depletion. (A) Ascorbic acid (2.5 mM) counteracts elongated growth upon genetic depletion of Hsp90 (0.1 μg/ml doxycycline [Dox]). The images were taken after 24 h. (B) Genetic depletion of Hsp90 for extended periods in the presence of 0.1 μg/ml doxycycline is not completely lethal. Propidium iodide-stained heat-killed wild-type cells (15 min at 80°C) clearly illustrate a complete lethal phenotype, as indicated by the staining, while cells genetically depleted of Hsp90 closely resemble the untreated wild-type cells. The images were taken at 24 h. (C) The percentage of yeast cells grown for 24 h in the presence of ascorbic acid was significantly (**, P < 0.001) higher than under the untreated conditions. (D) The sedimentation rate of cultures of a Candida strain in which the remaining Hsp90 was under the control of a tetracycline-repressible promoter was monitored in liquid YPD rich medium supplemented with 0.1 μg/ml Dox (◼) and 0.1 μg/ml Dox plus 2.5 mM ascorbic acid (●). The error bars indicate standard deviations.
Upc2 is required for the ascorbic acid-mediated effect.
A transcription factor deletion collection was previously used to characterize the regulatory system mediating Hsp90-regulated elongated growth (14). In a complementary approach, we screened a transcription factor overexpression library to identify TFs that upon overexpression prevent GdA-induced morphogenesis. The library consists of ∼200 strains, each carrying an ectopically expressed TF under the control of the inducible MET3 promoter (Stynen et al., unpublished). Methionine only was used to repress transcription, as cysteine was shown to interfere with elongation even in a wild-type strain due to its antioxidant properties (see Discussion and Fig. S2 in the supplemental material). Several transcriptional regulators whose overexpression resulted in blocked or decreased elongated growth in the presence of GdA were identified (Table 3). We continued with the strain overexpressing UPC2 (Fig. 3A), as it showed the strongest effect, based on semiquantitative observations. Expression of UPC2 under inducing conditions was ∼4.5-fold higher than that of an isogenic control strain carrying an empty plasmid (Fig. 3A). As expected, the characteristic elongated growth form was visible when the MET3 promoter was repressed by the addition of methionine, confirming that an increased dosage of Upc2 is responsible for the loss of elongation in the presence of inactive Hsp90 (Fig. 3B).
TABLE 3.
Transcription factors that upon overexpression block GdA-induced elongated growth
| TF | Descriptiona |
|---|---|
| Upc2 | Zn2-Cys6 transcriptional regulator of ergosterol biosynthetic genes and sterol uptake |
| Cap1 | AP-1 family bZIP transcription factor involved in drug resistance and oxidative-stress regulation |
| Dot6 | Uncharacterized ORF encoding a protein with a predicted role in telomeric gene silencing and filamentation |
| Rtg3 | Uncharacterized ORF encoding a putative transcription factor with a bZIP DNA-binding motif |
| Gcn4 | bZIP transcription factor involved in amino acid control response |
ORF, open reading frame.
FIG 3.
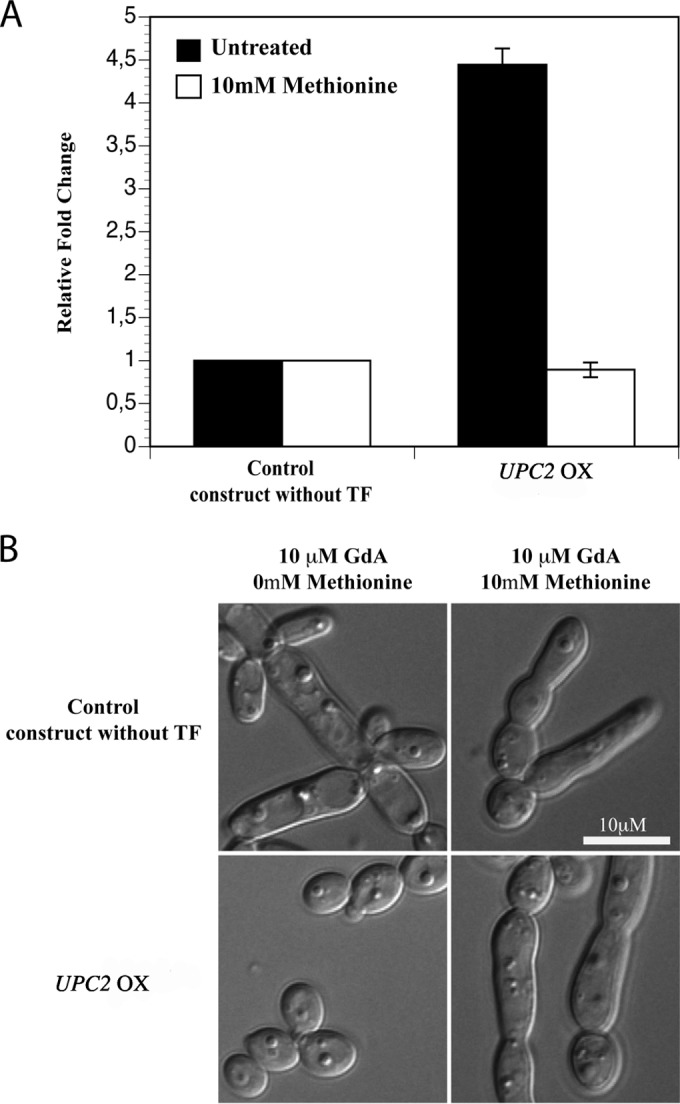
Overexpression of UPC2 represses GdA-induced elongated growth. (A) Quantitative real-time PCR of UPC2 expression under the control of the inducible MET3 promoter in the UPC2 overexpression (OX) strain and the control strain carrying an identical construct with the exception of the transcription factor. Promoter-repressing conditions in the presence of 10 mM methionine are represented by the white bars, while the black bars represent inducing conditions. The graph shows mean values with standard deviations from two independent experiments. (B) The inducible MET3 promoter allows regulated expression. Overexpression of UPC2 in the presence of GdA (10 μM) blocks the rise of the characteristic elongated growth form. The yeast form is reversed to the elongated structures under promoter-repressing conditions (represented by addition of 10 mM methionine). The control strain, carrying an identical expression construct except for the transcriptional regulator, displays a filamentous growth form under both promoter-repressing and -inducing conditions. The images were taken after 8 h of growth at 30°C in CSM−Met medium.
To confirm the role of Upc2 in the Hsp90-mediated inhibition of elongated growth forms, we tested two independent strains in which the endogenous UPC2 genes were deleted (22, 31), as such strains are expected to be hypersusceptible to GdA. Treatment of the wild-type strain with a low concentration of GdA (4 μM) has no effect on morphogenesis of wild-type cells. However, the same concentration was sufficient to induce elongated growth in the upc2Δ/upc2Δ strains, confirming their hypersusceptibility (strain D-6 is shown in Fig. 4A). In the complemented strain, elongation was impaired (Fig. 4A). Taken together, these observations indicate that Upc2 plays a role in the GdA-associated elongation process and confirm the previously observed genetic interaction between Upc2 and Hsp90 (34).
FIG 4.

The UPC2 deletion strain is hypersusceptible to GdA and impairs ascorbic acid-associated blockage of the elongated cell shape. (A) The UPC2 deletion strain is hypersusceptible to GdA. The upc2Δ/upc2Δ (D-6) strain was tested for its hypersusceptibility to GdA by treating it with 4 μM GdA, a concentration that is unable to induce elongated growth in the corresponding wild type. The UPC2 reintegrant strain (EC-7) (upc2Δ/upc2Δ + UPC2) displays a phenotype similar to that of the wild type. The images were taken after 8 h of growth at 30°C in YPD medium. (B) Deletion of UPC2 impairs ascorbic acid-associated blockage of the elongated cell shape. Deletion strains were grown at 30°C in YPD rich medium supplemented with either 10 μM GdA, 2.5 mM l-ascorbic acid, or a combination of the two. While ascorbic acid blocks the GdA-induced filamentation of the wild-type strain, it is unable to do so in the upc2Δ/upc2Δ mutants D-6 and TF077, while the reconstitution of UPC2 restores the yeast growth response. The images were taken after 8 h.
The obvious question, then, was whether Upc2 is required for the effect that ascorbic acid has on Hsp90-mediated cell elongation. Addition of ascorbic acid to wild-type cells prevents GdA-induced cell elongation. In the absence of Upc2 (in both independent deletion strains D-6 and TF077), GdA-induced cell elongation was not affected, whereas the UPC2 reconstituted strain EC-7 resulted in a phenotype similar to that of the wild-type strain (Fig. 4B). These results indicate that the effect of ascorbic acid on the morphological response upon inhibition of Hsp90 is Upc2 dependent.
Hsp90 inhibition or depletion results in reduced intracellular ergosterol levels.
To determine the mechanism by which ascorbic acid and Upc2 affect elongated cell growth, we tested the roles of genes whose expression is regulated by Upc2, such as the drug efflux pump genes CDR1 and MDR1 (17, 35). First, we hypothesized that ascorbic acid could increase the efflux of GdA, which was reported to be a substrate of the human ATP-binding cassette (ABC) transporter efflux pump, Mdr1 (36, 37). If this hypothesis were valid in C. albicans, one would expect mutants lacking Cdr1, a fungal ABC multidrug transporter (38), to elongate in the presence of both GdA and ascorbic acid. In addition, one would expect a strain devoid of this efflux pump to be hypersusceptible to GdA. However, neither hypersusceptibility to GdA nor impairment of the morphogenetic effect of ascorbic acid was observed for the cdr1Δ/cdr1Δ mutant (data not shown).
We also investigated Mdr1, a member of the multidrug resistance 1 (MDR1) major facilitator family. The ascorbic acid-mediated effect was not affected in strains overexpressing MDR1 (39) or with MDR1 deleted, and they were not hypersusceptible to GdA (data not shown). Together, these results rule out the hypothesis that an ascorbic acid-mediated increased efflux of GdA causes the ascorbic acid-mediated phenotype.
In addition to CDR1 and MDR1, expression of ERG11, a key gene in the ergosterol biosynthesis pathway, is also under the control of Upc2 (15, 22, 40). We therefore reasoned that levels of Erg11 might play an instrumental role in the morphogenetic transition regulated by Hsp90. The role of Erg11 in ascorbic-acid-induced inhibition was investigated using an engineered strain in which the only ERG11 allele is under the control of a tetracycline-repressible promoter (41). Cells in which the transcription of ERG11 was repressed by the addition of doxycycline but that were still viable (see Fig. S3 in the supplemental material) were tested for GdA-induced elongation in the presence of ascorbic acid. As shown in Fig. 5A (compare images a, c, g, and h), Erg11 expression is required for the ascorbic acid-induced inhibition of cell elongation upon GdA treatment. We also tested an ERG11 overexpression strain (∼3.5-fold higher expression than the control strain) (42), but as shown in Fig. 5B, such increased dosage of ERG11 was not able to suppress the elongated phenotype upon Hsp90 inhibition, demonstrating that increased dosage of Erg11 is not sufficient to block GdA-induced elongation. We also investigated whether ascorbic acid induces ERG11 expression in the presence and absence of Hsp90 inhibitors. In the wild-type strain, there is a significant drop in ERG11 expression upon GdA treatment, while simultaneous addition of ascorbic acid to the treatment leads to increased transcription of ERG11 (Fig. 5C). This increase is absent in a upc2Δ/upc2Δ mutant, suggesting a role for Upc2 as an important regulator.
FIG 5.

ERG11 is required for the ascorbic acid-mediated effect. (A) A tetO-ERG11/erg11Δ strain was grown at 30°C in YPD rich medium supplemented with either 10 μM GdA, 2.5 mM l-ascorbic acid, 0.1 μg/ml doxycycline, or a combination of these substances. While ascorbic acid blocks elongation upon GdA treatment of the tetO-ERG11/erg11Δ strain in the absence of doxycycline, compromising ERG11 expression is able to overcome this ascorbic acid-associated blockage. The images were taken after 8 h. (B) Overexpression of ERG11 is not sufficient to block elongated growth caused by Hsp90 inhibition. C. albicans strains AFA60 (control) and AFA59b (ACT1p-CaERG11) were grown at 30°C in YPD rich medium supplemented with 10 μM GdA. GdA-induced elongation of both the control and the ACT1p-CaERG11 strains shows that an increased dosage of Erg11 cannot suppress elongation. The images were taken after 8 h. (C) There is a significant drop (*, P < 0.05) in ERG11 expression upon GdA treatment in the wild type, while simultaneous addition of ascorbic acid during this treatment leads to an increase. Interestingly, the latter observation is absent in the upc2Δ/upc2Δ mutant, suggesting a role for Upc2. The data are the means ± standard deviations of triplicate experiments.
Erg11's eminent role in the regulation of sterol biosynthesis (15, 22, 43) and ascorbic acid-mediated effects on the Hsp90 morphogenetic circuitry (Fig. 1A and 2A) suggest a possible relationship between the ergosterol content and ascorbic acid-induced inhibition of cell elongation in the presence of GdA. To verify the impact of impaired Hsp90 function on ergosterol levels, we determined ergosterol levels in the wild-type and upc2Δ/upc2Δ strains upon treatment with GdA (10 μM) and/or ascorbic acid (2.5 mM). As shown in Fig. 6A, a significant decrease in ergosterol levels can be observed after addition of GdA to the wild-type strain (P < 0.05) and the upc2Δ/upc2Δ mutant (P < 0.001) compared to the corresponding untreated control strains. Whereas ergosterol levels are restored when ascorbic acid (2.5 mM) is added together with GdA to the wild-type strain, addition of both compounds to upc2Δ/upc2Δ cells failed to show a similar restoration. The difference in ergosterol levels compared to the untreated upc2Δ/upc2Δ mutant remained significant (P < 0.05). As can be seen, there was also a detectable decrease in ergosterol levels when ascorbic acid alone was administered, but this reduction was not significant (P > 0.05). These results point to a correlation between ergosterol levels and Hsp90-governed elongated growth. To determine whether reduced ergosterol levels were the result of oxidative stress caused by the absence of Upc2, as was previously claimed (44), we tested the two independent upc2 mutant strains from the Homann collection (31) that we used throughout our study for their susceptibility to oxidative stress. Figure S4 in the supplemental material shows that the tolerance for H2O2 of the mutants is absolutely comparable to that of the corresponding isogenic wild-type strain, thus ruling out the hypothesis that deletion of UPC2 could lower the anti-oxidative-stress potential of the cells. Different response of other upc2 mutants to oxidative stress may have different causes, such as a different genetic background (44).
FIG 6.
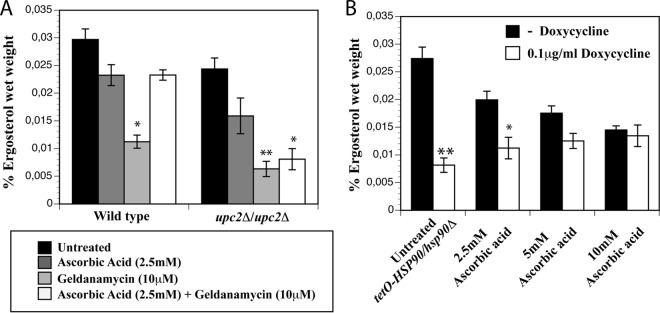
Hsp90 inhibition or depletion results in reduced intracellular ergosterol levels. (A) Ergosterol measurements in the wild type and the upc2Δ/upc2Δ mutant show a significant decrease in ergosterol content after pharmacological inhibition of Hsp90 in comparison with the representative untreated control (*, P < 0.05; **, P < 0.001). Addition of ascorbic acid under these conditions was reflected by an increase of ergosterol content only in the wild type. The results are means ± standard deviations of at least three independent measurements. (B) Ergosterol measurements in the tetO-HSP90/hsp90Δ strain show a similar significant decrease in ergosterol content in the presence of doxycycline (0.1 μg/ml) compared to the untreated control (*, P < 0.05; **, P < 0.001). This drop in ergosterol levels has great similarities to pharmacological inhibition by GdA. Addition of ascorbic acid is reflected by an increase in ergosterol content in the presence of doxycycline, while a general decrease is noticeable in its absence. The data are means ± standard deviations of triplicate experiments.
The results obtained with pharmacological inhibition of Hsp90 were confirmed using genetic depletion of Hsp90. The tetO-HSP90/hsp90Δ strain was grown in the presence and absence of doxycycline, and ergosterol levels were determined. As shown in Fig. 6B, depletion of Hsp90 results in a significant decrease in ergosterol levels (P < 0.001). Similar to pharmacological inhibition, addition of ascorbic acid results in restoration of ergosterol levels, with higher ergosterol levels in the presence of higher ascorbic acid levels. To rule out a more indirect effect of ascorbic acid on ergosterol levels via its antioxidant effect, we analyzed reactive oxygen species (ROS) production by way of rhodamine fluorescence in the wild-type strain and in the Tet0-HSP90/hsp90Δ strain in the presence or absence of doxycycline and with or without ascorbic acid. The fluorescence of doxycycline-treated cells, as well as of doxycycline- and ascorbic acid-treated cells, was comparable to that of untreated cells. H2O2-treated cells were used as the positive control (data not shown).
These results suggest that decreased ergosterol levels caused by Hsp90 inhibition (Fig. 6A and B) promote elongated growth. However, Fig. 5A (image d) shows that a reduction of ergosterol levels (by downregulating ERG11 expression) was not sufficient to induce elongation, since elongated growth forms still required Hsp90 inhibition.
To our knowledge, this is the first time that a clear correlation between impaired/decreased Hsp90 expression and ergosterol levels has been observed. Moreover, addition of ascorbic acid restores ergosterol levels to wild-type levels, and this is clearer upon pharmacological inhibition than upon genetic depletion of Hsp90.
Upc2 is required for other ascorbic acid-mediated effects.
It has been reported in previous studies that ascorbic acid reduces the antifungal effect of fluconazole, possibly via its antioxidant properties (45). The authors suggested that the antioxidant properties of ascorbic acid counteract fluconazole-induced reactive oxygen species, resulting in an effect similar to that obtained with other antioxidants, such as glutathione (45). In order to establish a general role for Upc2 in orchestrating ascorbic acid-mediated phenomena, we verified a possible relationship between ascorbic acid and fluconazole in a upc2Δ/upc2Δ strain by means of checkerboard assays, two-dimensional tests designed to measure drug-drug interactions. Fluconazole and ascorbic acid display antagonistic activities, as determined via the calculation of the FICI (FICI, 33) (Table 4). This kind of interaction is no longer detectable when the checkerboard is performed using cells of the upc2Δ/upc2Δ strain (FICI, 0.75) (Table 4). Thus, the combination of ascorbic acid with fluconazole no longer displayed antagonistic activity in cells lacking the Upc2 transcription factor.
TABLE 4.
Antagonistic effect of ascorbic acid on fluconazolea
| Strain | MICFLC (μg/ml) | MICFLC/AA (μg/ml) | MICAA (μg/ml) | MICAA/FLC (μg/ml) | FIC indexb |
|---|---|---|---|---|---|
| Wild type | 2 | >64 | 1761.2 (10 mM) | 1761.2 (10 mM) | 33 |
| upc2Δ/upc2Δ | 0.25 | 0.125 | 1761.2 (10 mM) | 440.3 (2.5 mM) | 0.75 |
The antagonistic effect of ascorbic acid on fluconazole is not present in a upc2Δ/upc2Δ mutant. MICFLC and MICAA are the MICs of fluconazole and ascorbic alone, while MICFLC/AA and MICAA/FLC are the MICs of ascorbic acid and fluconazole in combination.
Interpretation of the FIC index was performed as previously described (30).
DISCUSSION
A decreased dosage of Hsp90, as well as its pharmacologic depletion, results in a morphology change of C. albicans at low temperature from yeast to elongated cells (13). The molecular mechanism of this morphogenetic transition has been studied extensively over the last few years (46, 47). We contribute to this characterization by showing that reduced levels of ergosterol result from GdA-mediated impairment of Hsp90 and that these changes correlate with elongated growth. The effect of GdA can be suppressed by addition of ascorbic acid, which again increased levels of ergosterol to normal and resulted in cell growth in the yeast form. Ascorbic acid suppression did not depend on its antioxidant activity but on the transcriptional regulator Upc2, as in a upc2Δ/upc2Δ mutant ascorbic acid cannot restore ergosterol levels to normal upon GdA treatment. In summary, ascorbic acid can block GdA-dependent elongated growth by restoring normal ergosterol levels in a Upc2-dependent fashion.
In the course of our work, we found that several antioxidants interfered with the activity of GdA. Apart from its known inhibition of Hsp90, GdA is also involved in the generation of superoxide radicals, which is attributed to the presence of its quinone group (48). The presence of molecules with antioxidant properties alleviates the detrimental effects of oxidative stress imposed by free radicals. A variety of molecules, such as reduced glutathione (GSH) and its derivatives, cysteine, dithiothreitol, and ascorbic acid, are able to fulfill these protective requirements (49). It is possible that the superoxide radicals produced in the presence of GdA could affect elongated cell shape formation, and addition of ascorbic acid would then block this through its antioxidant activities. This would fit with data obtained by Patterson et al. (50), who showed that H2O2-mediated activation of Cap1 was a prerequisite for yeast-to-hypha transition.
However, the fact that the antagonistic effect of ascorbic acid on morphology is observed on cells genetically depleted of Hsp90, as well as on cells whose Hsp90 is inhibited by GdA, suggests that ascorbic acid acts directly or indirectly on Hsp90 or Hsp90-mediated processes. Other authors have reported an influence of ascorbic acid or its analogues on morphogenetic transitions in C. albicans (23, 24, 51). Nasution and colleagues (23) reported that addition of ascorbic acid not only lowered the intracellular concentration of reactive oxygen species, but also inhibited H2O2 and serum-initiated hyphal differentiation. Ojha and coworkers (24) similarly demonstrated inhibition of serum-induced hyphal formation by ascorbic acid and suggested a mode of action by interruption of the hyphal formation signal of C. albicans.
During the TF overexpression screening, we observed that cysteine directly affected the activity of GdA, as no elongated growth could be observed in the wild-type strain. This could be explained by earlier reports describing that thiol-containing antioxidants (e.g., cysteine) could physically interact with Hsp90 inhibitors, such as GdA and radicicol, and render them inactive as a consequence of the thiol-mediated interaction (52–57). This means that one has to be careful when using the MET3 promoter in combination with GdA or radicicol, as in this case cysteine, which is used to repress the promoter, will affect the activity of GdA and radicicol. This is the reason why in our experiments we used only methionine to repress the MET3 promoter.
One of the TFs that inhibited GdA-induced morphogenesis was Upc2, a Zn2Cys6 transcription factor, a key regulator of drug efflux pump expression and ergosterol biosynthesis (15, 35). Here, we show that Upc2 is required for ascorbic acid inhibition of cell elongation upon compromised Hsp90 function. Previously, it was shown that Upc2 is a key regulator of the ascorbic acid-mediated effect on fluconazole, as it decreases the antifungal effect of fluconazole, thus displaying a protective role in fungi (45). Together, these data show that Upc2 plays a central role as the transcriptional regulator in ascorbic acid-associated phenomena. Ascorbic acid is very important for human life because of its antioxidant properties, protecting cells from oxidative stress (58). The use of ascorbic acid is even clinically relevant for the treatment of a variety of diseases, such as respiratory tract infections (59, 60).
One of the genes that is regulated by Upc2 is ERG11, a gene involved in ergosterol biosynthesis and the molecular target of the azole antifungals (15, 22, 43, 61, 62). Previous work demonstrated that lower ergosterol levels, caused by ergosterol biosynthesis inhibitors, such as azoles, have an inhibitory effect on hypha formation (63, 64). However, elongated growth forms resulting from impaired Hsp90 function are not considered true hyphae (14, 65, 66). Consistent with the relationship between ergosterol depletion and C. albicans morphogenesis mentioned by Victoria et al. (67), we report a correlation between ergosterol content and elongated growth caused by impairment of Hsp90 function. The morphogenetic switch to elongated growth forms by either pharmacological inhibition or genetic depletion is accompanied with a significant decrease in ergosterol content. One could question whether this drop in ergosterol is really the cause or rather a consequence of the morphogenetic transition upon Hsp90 inhibition. The formation of elongated growth structures under low ergosterol levels could be interpreted as a cellular stress response caused by the disruption of sterol homeostasis. However, this seems unlikely, since our results argue against an elongation-inducing role for low ergosterol levels. First, we demonstrated that reduced levels of Erg11 are not sufficient to promote elongation (Fig. 5A). In addition, it was found that increasing concentrations of ascorbic acid reduced ergosterol content without initiating elongated growth (Fig. 6A and B). The latter data are consistent with previous observations by Singh et al. (68), who reported low ergosterol levels in C. albicans cells when they were grown in the presence of ascorbic acid.
On the other hand, the increase of ergosterol to wild-type levels in the presence of GdA upon addition of ascorbic acid is absent in a upc2Δ/upc2Δ mutant, suggesting the importance of Upc2 as a transcriptional regulator of Erg11 to overcome the drop in ergosterol levels caused by impaired Hsp90 (Fig. 6A). As mentioned above, we showed that an increased dosage of ERG11 alone was not sufficient to overcome the induction of elongated growth upon pharmacological inhibition of Hsp90. This could indicate that ergosterol biosynthesis is not the most important Upc2-mediated effect in response to ascorbic acid or that the expression of other ERG genes could be regulated by Upc2. We are currently identifying other Upc2-regulated genes by performing chromatin immunoprecipitation-sequencing (ChIP-seq) and RNA-seq analyses. We cannot exclude a possible connection with the previously established pathways in the Hsp90-dependent morphogenetic circuitry, such as the cAMP-PKA and cell cycle pathways or the Pho85-Pcl1-Hms1 module (13, 14, 46, 65, 69) or a novel, yet-to-be-determined pathway. Ascorbic acid or Upc2 may be required for the modulation of an inhibitor of these Hsp90 signaling modules.
In conclusion, our study elucidates the molecular circuitry through which ascorbic acid influences Hsp90-dependent C. albicans morphogenesis, involving the transcriptional regulator Upc2. We found that ascorbic acid can block GdA-dependent elongated growth by restoring normal ergosterol levels in a Upc2-dependent fashion. We suggest that influencing ergosterol biosynthesis via Erg11 is not the primary mode of action of Upc2-mediated effects in response to ascorbic acid, as overexpression of ERG11 did not block elongation upon GdA treatment. Further research is required to explore the interesting relationship between Hsp90, Upc2, Erg11, and the morphogenetic machinery.
Supplementary Material
ACKNOWLEDGMENTS
We are grateful to L. Cowen, O. Homann, J. Morschhäuser, D. Sanglard, T. White, and T. Roemer for providing strains. We especially thank Ilse Palmans for excellent technical assistance.
This work was supported by a grant from the Flemish Institute for Science and Technology (IWT) to F.V.H. and by the Fund for Scientific Research Flanders (FWO) and the Research Fund of KU Leuven.
Footnotes
Published ahead of print 1 August 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/EC.00096-14.
REFERENCES
- 1.Blackwell M. 2011. The fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 98:426–438. 10.3732/ajb.1000298 [DOI] [PubMed] [Google Scholar]
- 2.Cheng S-C, Joosten LAB, Kullberg B-J, Netea MG. 2012. Interplay between Candida albicans and the mammalian innate host defense. Infect. Immun. 80:1304–1313. 10.1128/IAI.06146-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lo HJ, Kohler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, Fink GR. 1997. Nonfilamentous C. albicans mutants are avirulent. Cell 90:939–949. 10.1016/S0092-8674(00)80358-X [DOI] [PubMed] [Google Scholar]
- 4.Biswas S, Van Dijck P, Datta A. 2007. Environmental sensing and signal transduction pathways regulating morphopathogenic determinants. Microbiol. Mol. Biol. Rev. 71:348–376. 10.1128/MMBR.00009-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pande K, Chen C, Noble SM. 2013. Passage through the mammalian gut triggers a phenotypic switch that promotes Candida albicans commensalism. Nat. Genet. 45:1088–1091. 10.1038/ng.2710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Saville SP, Lazzell AL, Monteagudo C, Lopez-Ribot JL. 2003. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentatous forms of Candida albicans during infection. Eukaryot. Cell 2:1053–1060. 10.1128/EC.2.5.1053-1060.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chandra J, Mukherjee PK, Ghannoum MA. 2008. In vitro growth and analysis of Candida biofilms. Nat. Protoc. 3:1909–1924. 10.1038/nprot.2008.192 [DOI] [PubMed] [Google Scholar]
- 8.Sudbery PE, Gow NA, Berman J. 2004. The distinct morphogenetic states of Candida albicans. Trends Microbiol. 12:317–324. 10.1016/j.tim.2004.05.008 [DOI] [PubMed] [Google Scholar]
- 9.Albuquerque P, Casadevall A. 2012. Quorum sensing in fungi: a review. Med. Mycol. 50:337–345. 10.3109/13693786.2011.652201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shareck J, Belhumeur P. 2011. Modulation of morphogenesis in Candida albicans by various small molecules. Eukaryot. Cell 10:1004–1012. 10.1128/EC.05030-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hogan DA, Sundstrom P. 2009. The Ras/cAMP/PKA signaling pathway and virulence in Candida albicans. Future Microbiol. 4:1263–1270. 10.2217/fmb.09.106 [DOI] [PubMed] [Google Scholar]
- 12.Hogan DA, Muhlschlegel FA. 2011. Candida albicans development regulation: adenylyl cyclase as a coincidence detector of parallel signals. Curr. Opin. Microbiol. 14:682–686. 10.1016/j.mib.2011.09.014 [DOI] [PubMed] [Google Scholar]
- 13.Shapiro RS, Uppuluri P, Zaas AK, Collins C, Senn H, Perfect JR, Heitman J, Cowen LE. 2009. Hsp90 orchestrates temperature-dependent Candida albicans morphogenesis via Ras1-PKA signaling. Curr. Biol. 19:621–629. 10.1016/j.cub.2009.03.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shapiro RS, Sellam A, Tebbji F, Whiteway M, Nantel A, Cowen LE. 2012. Pho85, Pcl1, and Hms1 signaling governs Candida albicans morphogenesis induced by high temperature or Hsp90 compromise. Curr. Biol. 22:461–470. 10.1016/j.cub.2012.01.062 [DOI] [PubMed] [Google Scholar]
- 15.MacPherson S, Akache B, Weber S, De Deken X, Raymond M, Turcotte B. 2005. Candida albicans zinc cluster protein Upc2p confers resistance to antifungal drugs and is an activator of ergosterol biosynthetic genes. Antimicrob. Agents Chemother. 49:1745–1752. 10.1128/AAC.49.5.1745-1752.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vik A, Rine J. 2001. Upc2p and Ecm22p, dual regulators of sterol biosynthesis in Saccharomyces cerevisiae. Mol. Cell. Biol. 21:6395–6405. 10.1128/MCB.21.19.6395-6405.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Znaidi S, Weber S, Al-Abdin OZ, Bomme P, Saidane S, Drouin S, Lemieux S, De Deken X, Robert F, Raymond M. 2008. Genomewide location analysis of Candida albicans Upc2p, a regulator of sterol metabolism and azole drug resistance. Eukaryot. Cell 7:836–847. 10.1128/EC.00070-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Heilmann CJ, Schneider S, Barker KS, Rogers PD, Morschhäuser J. 2010. An A643T mutation in the transcription factor Upc2p causes constitutive ERG11 upregulation and increased fluconazole resistance in Candida albicans. Antimicrob. Agents Chemother. 54:353–359. 10.1128/AAC.01102-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hoot SJ, Smith AR, Brown RP, White TC. 2011. An A643V amino acid substitution in Upc2p contributes to azole resistance in well-characterized clinical isolates of Candida albicans. Antimicrob. Agents Chemother. 55:940–942. 10.1128/AAC.00995-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Flowers SA, Barker KS, Berkow EL, Toner G, Chadwick SG, Gygax SE, Morschhäuser J, Rogers PD. 2012. Gain-of-function mutations in UPC2 are a frequent cause of ERG11 upregulation in azole-resistant clinical isolates of Candida albicans. Eukaryot. Cell 11:1289–1299. 10.1128/EC.00215-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Morio F, Pagniez F, Besse M, Gay-Andrieu F, Miegeville M, Le Pape P. 2013. Deciphering azole resistance mechanisms with a focus on transcription factor-encoding genes TAC1, MRR1 and UPC2 in a set of fluconazole-resistant clinical isolates of Candida albicans. Int. J. Antimicrob. Agents 42:410–415. 10.1016/j.ijantimicag.2013.07.013 [DOI] [PubMed] [Google Scholar]
- 22.Silver PM, Oliver BG, White TC. 2004. Role of Candida albicans transcription factor Upc2p in drug resistance and sterol metabolism. Eukaryot. Cell 3:1391–1397. 10.1128/EC.3.6.1391-1397.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nasution O, Srinivasa K, Kim M, Kim YJ, Kim W, Jeong W, Choi W. 2008. Hydrogen peroxide induces hyphal differentiation in Candida albicans. Eukaryot. Cell 7:2008–2011. 10.1128/EC.00105-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ojha R, Manzoor N, Khan LA. 2009. Ascorbic acid modulates pathogenicity markers of Candida albicans. Int. J. Micriobiol. Res. 1:19–24. 10.9735/0975-5276.1.1.19-24 [DOI] [Google Scholar]
- 25.Eboigbodin KE, Biggs CA. 2008. Characterization of the extracellular polymeric substances produced by Escherichia coli using infrared spectroscopic, proteomic, and aggregation studies. Biomacromolecules 9:686–695. 10.1021/bm701043c [DOI] [PubMed] [Google Scholar]
- 26.Fu MS, De Sordi L, Muhlschlegel FA. 2012. Functional characterization of the small heat shock protein Hsp12p from Candida albicans. PLoS One 7:e42894. 10.1371/journal.pone.0042894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stynen B, Van Dijck P, Tournu H. 2010. A CUG codon adapted two-hybrid system for the pathogenic fungus Candida albicans. Nucleic Acids Res. 38:e184. 10.1093/nar/gkq725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Care RS, Trevethick J, Binley KM, Sudbery PE. 1999. The MET3 promoter: a new tool for Candida albicans molecular genetics. Mol. Microbiol. 34:792–798. 10.1046/j.1365-2958.1999.01641.x [DOI] [PubMed] [Google Scholar]
- 29.Arthington-Skaggs BA, Jradi H, Desai T, Morrison CJ. 1999. Quantification of ergosterol content: novel method for determination of fluconazole susceptibility of Candida albicans. J. Clin. Microbiol. 37:3332–3337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Odds FC. 2003. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 52:1. 10.1093/jac/dkg301 [DOI] [PubMed] [Google Scholar]
- 31.Homann OR, Dea J, Noble SM, Johnson AD. 2009. A phenotypic profile of the Candida albicans regulatory network. PLoS Genet. 5:e1000783. 10.1371/journal.pgen.1000783 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sudbery P. 2011. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 9:737–748. 10.1038/nrmicro2636 [DOI] [PubMed] [Google Scholar]
- 33.Roe SM, Prodromou C, O'Brien R, Ladbury JE, Piper PW, Pearl LH. 1999. Structural basis for inhibition of the Hsp90 molecular chaperone by the antitumor antibiotics radicicol and geldanamycin. J. Med. Chem. 42:260–266. 10.1021/jm980403y [DOI] [PubMed] [Google Scholar]
- 34.Diezmann S, Michaut M, Shapiro RS, Bader GD, Cowen LE. 2012. Mapping the Hsp90 genetic interaction network in Candida albicans reveals environmental contingency and rewired circuitry. PLoS Genet. 8:e1002562. 10.1371/journal.pgen.1002562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Schubert S, Barker KS, Znaidi S, Schneider S, Dierolf F, Dunkel N, Aid M, Boucher G, Rogers PD, Raymond M, Morschhauser J. 2011. Regulation of efflux pump expression and drug resistance by the transcription factors Mrr1, Upc2, and Cap1 in Candida albicans. Antimicrob. Agents Chemother. 55:2212–2223. 10.1128/AAC.01343-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Huang Y, Blower PE, Liu R, Dai Z, Pham AN, Moon H, Fang J, Sadee W. 2007. Chemogenomic analysis identifies geldanamycins as substrates and inhibitors of ABCB1. Pharm. Res. 24:1702–1712. 10.1007/s11095-007-9300-x [DOI] [PubMed] [Google Scholar]
- 37.Workman P. 2003. Auditing the pharmacological accounts for Hsp90 molecular chaperone inhibitors: unfolding the relationship between pharmacokinetics and pharmacodynamics. Mol. Cancer Ther. 2:131–138 [PubMed] [Google Scholar]
- 38.Prasad R, De Wergifosse P, Goffeau A, Balzi E. 1995. Molecular cloning and characterization of a novel gene of Candida albicans, CDR1, conferring multiple resistance to drugs and antifungals. Curr. Genet. 27:320–329. 10.1007/BF00352101 [DOI] [PubMed] [Google Scholar]
- 39.Hiller D, Sanglard D, Morschhauser J. 2006. Overexpression of the MDR1 gene is sufficient to confer increased resistance to toxic compounds in Candida albicans. Antimicrob. Agents Chemother. 50:1365–1371. 10.1128/AAC.50.4.1365-1371.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dunkel N, Liu TT, Barker KS, Homayouni R, Morschhauser J, Rogers PD. 2008. A gain-of-function mutation in the transcription factor Upc2p causes upregulation of ergosterol biosynthesis genes and increased fluconazole resistance in a clinical Candida albicans isolate. Eukaryot. Cell 7:1180–1190. 10.1128/EC.00103-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Becker JM, Kaufman SJ, Hauser M, Huang L, Lin M, Sillaots S, Jian B, Xu D, Roemer T. 2010. Pathway analysis of Candida albicans survival and virulence determinants in a murine infection model. Proc. Natl. Acad. Sci. U. S. A. 107:22044–22049. 10.1073/pnas.1009845107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fiori A, Van Dijck P. 2012. Potent synergistic effect of doxycycline with fluconazole against Candida albicans is mediated by interference with iron homeostasis. Antimicrob. Agents Chemother. 56:3785–3796. 10.1128/AAC.06017-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Song JL, Harry JB, Eastman RT, Oliver BG, White TC. 2004. The Candida albicans lanosterol 14-alpha-demethylase (ERG11) gene promoter is maximally induced after prolonged growth with antifungal drugs. Antimicrob. Agents Chemother. 48:1136–1144. 10.1128/AAC.48.4.1136-1144.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Dhamgaye S, Devaux F, Manoharlal R, Vandeputte P, Shah AH, Singh A, Blugeon D, Sanglard D, Prasad R. 2012. In vitro effect of malachite green on Candida albicans involves multiple pathways and transcriptional regulators UPC2 and STP2. Antimicrob. Agents Chemother. 56:495–506. 10.1128/AAC.00574-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wang Y, Jia XM, Jia JH, Li MB, Cao YY, Gao PH, Liao WQ, Cao YB, Jiang YY. 2009. Ascorbic acid decreases the antigungal effect of fluconazole in the treatment of candidiasis. Clin. Exp. Pharmacol. Physiol. 36:e40–e46. 10.1111/j.1440-1681.2009.05187.x [DOI] [PubMed] [Google Scholar]
- 46.Shapiro RS, Zaas AK, Betancourt-Quiroz M, Perfect JR, Cowen LE. 2012. The Hsp90 co-chaperone Sgt1 governs Candida albicans morphogenesis and drug resistance. PLoS One 7:e44734. 10.1371/journal.pone.0044734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Shapiro RS, Cowen LE. 2012. Thermal control of microbial development and virulence: molecular mechanisms of microbial temperature sensing. mBio 3:e00238–00212. 10.1128/mBio.00238-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dikalov S, Landmesser U, Harrison DG. 2002. Geldanamycin leads to superoxide formation by enzymatic and non-enzymatic redox cycling. Implications for studies of Hsp90 and endothelial cell nitric-oxide synthase. J. Biol. Chem. 277:25480–25485. 10.1074/jbc.M203271200 [DOI] [PubMed] [Google Scholar]
- 49.Deneke SM. 2000. Thiol-based antioxidants. Curr. Top. Cell Regul. 36:151–180 [DOI] [PubMed] [Google Scholar]
- 50.Patterson MJ, McKenzie CG, Smith DA, da Silva Dantas A, Sherston S, Veal EA, Morgan BA, MacCallum DM, Erwig LP, Quinn J. 2013. Ybp1 and Gpx3 signaling in Candida albicans govern hydrogen-peroxide-induced oxidation of the Cap1 transcription factor and macrophage escape. Antioxid. Redox Signal. 19:2244–2260. 10.1089/ars.2013.5199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Huh WK, Kim ST, Kim H, Jeong G, Kang SO. 2001. Deficiency of D-erythroascorbic acid attenuates hyphal growth and virulence of Candida albicans. Infect. Immun. 69:3939–3946. 10.1128/IAI.69.6.3939-3946.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Moulin E, Zoete V, Barluenga S, Karplus M, Winssinger N. 2005. Design, synthesis, and biological evaluation of HSP90 inhibitors based on conformational analysis of radicicol and its analogues. J. Am. Chem. Soc. 127:6999–7004. 10.1021/ja043101w [DOI] [PubMed] [Google Scholar]
- 53.Cysyk RL, Parker RJ, Barchi JJ, Jr, Steeg PS, Hartman NR, Strong JM. 2006. Reaction of geldanamycin and C17-substituted analogues with glutathione: product identifications and pharmacological implications. Chem. Res. Toxicol. 19:376–381. 10.1021/tx050237e [DOI] [PubMed] [Google Scholar]
- 54.Guo W, Reigan P, Siegel D, Ross D. 2008. Enzymatic reduction and glutathione conjugation of benzoquinone ansamycin heat shock protein 90 inhibitors: relevance for toxicity and mechanism of action. Drug Metab. Dispos. 36:2050–2057. 10.1124/dmd.108.022004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Samuni A, Goldstein S. 2012. Redox properties and thiol reactivity of geldanamycin and its analogues in aqueous solutions. J. Phys. Chem. B. 116:6404–6410. 10.1021/jp304206n [DOI] [PubMed] [Google Scholar]
- 56.Ross D. 1988. Glutathione, free radicals and chemotherapeutic agents. Mechanisms of free-radical induced toxicity and glutathione-dependent protection. Pharmacol. Ther. 37:231–249 [DOI] [PubMed] [Google Scholar]
- 57.Agatsuma T, Ogawa H, Akasaka K, Asai A, Yamashita Y, Mizukami T, Akinaga S, Saitoh Y. 2002. Holydrin and oxime derivatives of radicicol: synthesis and antitumor activities. Bioorg. Med. Chem. 10:3445–3454. 10.1016/S0968-0896(02)00260-2 [DOI] [PubMed] [Google Scholar]
- 58.Padayatty SJ, Katz A, Wang Y, Eck P, Kwon O, Lee JH, Chen S, Corpe C, Dutta A, Dutta SK, Levine M. 2003. Vitamin C as an antioxidant: evaluation of its role in disease prevention. J. Am. Coll. Nutr. 22:18–35. 10.1080/07315724.2003.10719272 [DOI] [PubMed] [Google Scholar]
- 59.Arroll B. 2005. Non-antibiotic treatments for upper-respiratory tract infections (common cold). Respir. Med. 99:1477–1484. 10.1016/j.rmed.2005.09.039 [DOI] [PubMed] [Google Scholar]
- 60.Stephensen CB, Marquis GS, Jacob RA, Kruzich LA, Douglas SD, Wilson CM. 2006. Vitamins C and E in adolescents and young adults with HIV infection. Am. J. Clin. Nutr. 83:870–879 [DOI] [PubMed] [Google Scholar]
- 61.Oliver BG, Song JL, Choiniere JH, White TC. 2007. cis-Acting elements within the Candida albicans ERG11 promoter mediate the azole response through transcription factor Upc2p. Eukaryot. Cell 6:2231–2239. 10.1128/EC.00331-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hoot SJ, Oliver BG, White TC. 2008. Candida albicans UPC2 is transcriptionally induced in response to antifungal drugs and anaerobicity through Upc2p-dependent and -independent mechanisms. Microbiology 154:2748–2756. 10.1099/mic.0.2008/017475-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ha KC, White TC. 1999. Effects of azole antifungal drugs on the transition from yeast cells to hyphae in susceptible and resistant isolates of the pathogenic yeast Candida albicans. Antimicrob. Agents Chemother. 43:763–768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Odds FC, Cockayne A, Hayward J, Abbott AB. 1985. Effects of imidazole- and triazole-derivative antifungal compounds on the growth and morphological development of Candida albicans hyphae. J. Gen. Microbiol. 131:2581–2589 [DOI] [PubMed] [Google Scholar]
- 65.Senn H, Shapiro RS, Cowen LE. 2012. Cdc28 provides a molecular link between Hsp90, morphogenesis, and cell cycle progression in Candida albicans. Mol. Biol. Cell 23:268–283. 10.1091/mbc.E11-08-0729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bachewich C, Nantel A, Whiteway M. 2005. Cell cycle arrest during S or M phase generates polarized growth via distinct signals in Candida albicans. Mol. Microbiol. 57:942–959. 10.1111/j.1365-2958.2005.04727.x [DOI] [PubMed] [Google Scholar]
- 67.Victoria GS, Yadav B, Lauhnar L, Jain P, Bhatnagar S, Komath SS. 2012. Mutual co-regulation between GPI-N-acetylglucosaminyltransferase and ergosterol biosynthesis in Candida albicans. Biochem. J. 443:619–625. 10.1042/BJ20120143 [DOI] [PubMed] [Google Scholar]
- 68.Singh M, Jayakumar A, Prasad R. 1979. The effect of altered ergosterol content on the transport of various amino acids in Candida albicans. Biochim. Biophys. Acta 555:42–55. 10.1016/0005-2736(79)90070-1 [DOI] [PubMed] [Google Scholar]
- 69.Serneels J, Tournu H, Van Dijck P. 2012. Tight control of trehalose content is required for efficient heat-induced cell elongation in Candida albicans. J. Biol. Chem. 287:36873–36882. 10.1074/jbc.M112.402651 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Gillum AM, Tsay EYH, Kirsch DR. 1984. Isolation of the Candida albicans gene for orotidine-5′-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 198:179–182. 10.1007/BF00328721 [DOI] [PubMed] [Google Scholar]
- 71.Fonzi WA, Irwin MY. 1993. Isogenic strain construction and gene mapping in Candida albicans. Genetics 134:717–728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sanglard D, Ischer F, Monod M, Bille J. 1996. Susceptibilities of Candida albicans multidrug transporter mutants to various antifungal agents and other metabolic inhibitors. Antimicrob. Agents Chemother. 40:2300–2305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Noble SM, Johnson AD. 2005. Strains and strategies for large-scale gene deletion studies of the diploid human fungal pathogen Candida albicans. Eukaryot. Cell 4:298–309. 10.1128/EC.4.2.298-309.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Cowen LE, Singh SD, Kohler JR, Collins C, Zaas AK, Schell WA, Aziz H, Mylonakis E, Perfect JR, Whitesell L, Lindquist S. 2009. Harnessing Hsp90 function as a powerful, broadly effective therapeutic strategy for fungal infectious disease. Proc. Natl. Acad. Sci. U. S. A. 106:2818–2823. 10.1073/pnas.0813394106 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.