

Abstract
Background
Canine mammary tumors (CMTs) are the most common type of cancer found in female dogs. Establishment and evaluation of tumor cell lines can facilitate investigations of the biological mechanisms of cancer. Different cell models are used to investigate genetic, epigenetic, and cellular pathways, cancer progression, and cancer therapeutics. Establishment of new cell models will greatly facilitate research in this field. In the present study, we established and characterized two new CMT cell lines derived from a single CMT.
Results
We established two cell lines from a single malignant CMT specimen: DTK-E and DTK-SME. Morphologically, the DTK-E cells were large, flat, and epithelial-like, whereas DTK-SME cells were round and epithelial-like. Doubling times were 24 h for DTK-E and 18 h for DTK-SME. On western blots, both cell lines expressed cytokeratin AE1, vimentin, cytokeratin 7 (CK7), and heat shock protein 27 (HSP27). Moreover, investigation of chemoresistance revealed that DTK-SME was more resistant to doxorubicin-induced apoptosis than DTK-E was. After xenotransplantation, both DTK-E and DTK-SME tumors appeared within 14 days, but the average size of DTK-SME tumors was greater than that of DTK-E tumors after 56 days.
Conclusion
We established two new cell lines from a single CMT, which exhibit significant diversity in cell morphology, protein marker expression, tumorigenicity, and chemoresistance. The results of this study revealed that the DTK-SME cell line was more resistant to doxorubicin-induced apoptosis and exhibited higher tumorigenicity in vivo than the DTK-E cell line. We anticipate that the two novel CMT cell lines established in this study will be useful for investigating the tumorigenesis of mammary carcinomas and for screening anticancer drugs.
Keywords: Canine mammary tumor, Cell line, Heat shock protein 27, Chemoresistance, Tumorigenicity
Background
Mammary gland carcinomas are the most common neoplasms in both women and female dogs [1,2]. Clinical evidence has shown that approximately 50% of canine mammary tumors (CMTs) are diagnosed as malignant in dogs [3]. CMTs possess epidemiological, clinicopathological, and biochemical characteristics similar to those of human breast carcinomas [4,5]. Therefore, CMTs have been considered an appropriate and valid model for human breast cancer studies for more than three decades [6].
Tumor cell lines have been widely used for in vitro research, and have proven to be a particularly useful tool for genetic analyses. Previous characterizations of tumor cell lines have shown that they are also excellent models for studying the biological mechanisms underlying cancer [7]. The use of tumor cell lines has increased our knowledge of the deregulated genes and signaling pathways involved in cancer [8,9]. Furthermore, original cell models have been developed to test anticancer drugs [8,10–12]. Tumor cell lines continue to be used in the development of new therapies [7,11,13], and also provide an alternative to direct transplantation of tumors in animals for testing chemotherapeutics [14].
The use of an appropriate in vitro model is crucial in cancer research. Different cell models are used to investigate genetic, epigenetic, and cellular pathways [7]; proliferation deregulation, apoptosis, and cancer progression [9]; and identification of potential molecular markers [15], as well as to screen and characterize cancer therapeutics [11,16]. The findings reported from tumor cell lines are typically extrapolated to in vivo human tumors [15]. Thus, many biomedical and pharmaceutical companies have recognized the importance of tumor cell lines as models for drug testing and translational studies [8].
In the present study, we established and characterized two new CMT cell lines, termed DTK-E and DTK-SME. These two cell lines exhibited significant diversity in cell morphology, protein marker expression, tumorigenicity, and chemoresistance. Our results suggest that the two established cell lines might constitute a useful experimental model for investigating the tumorigenesis of mammary carcinomas and for screening potential anticancer drugs.
Results
Morphological analysis of established cell lines
After over 100 passages, two CMT cell lines, DTK-E and DTK-SME, were established from canine mammary carcinomas. When adhered to a culture plate, DTK-E cells exhibited a large, flat, epithelial-like morphology (Figure 1A), while DTK-SME cells presented a rounded, epithelial-like morphology (Figure 1B). Of the two established cell lines, only DTK-SME cells possessed pile up (Figure 1B). Transmission electron micrographs showed that both DTK-E and DTK-SME cells exhibited high nuclear-cytoplasmic ratios and large nucleoli (Figure 2).
Figure 1.
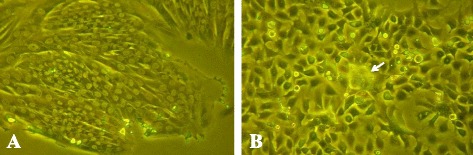
Phase contrast micrographs of CMT cells derived from malignant tumor tissue. (A) DTK-E cells displayed mostly large, flat, epithelial-like characteristics; (B) DTK-SME cells displayed rounded, epithelial-like features and possessed pile up. Arrowhead indicates position of pile up.
Figure 2.
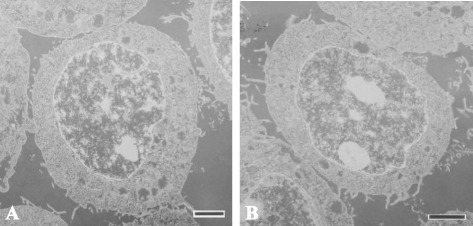
Transmission electron micrographs of CMT cell lines. Intracellular morphology of (A) DTK-E and (B) DTK-SME cells. Images are representative of 20 random sections per cell line. Scale bars = 2 μm.
Biological analysis of established cell lines
Alterations in the biological properties of the two CMT cell lines were monitored during their establishment. During the first 10 passages, 50% fetal bovine serum (FBS) was required to maintain cell growth. After 10 passages, the percentage of FBS in the medium was gradually reduced. After 100 passages, 5% FBS was sufficient to support the growth of both CMT cell lines. In addition, contact inhibition was only significant during the first 35 passages, and was completely lost after 50 passages (data not shown). During establishment of the two CMT cell lines, we also determined the doubling time. The final doubling times for DTK-E and DTK-SME cells were 24 h and 18 h, respectively (Figure 3).
Figure 3.
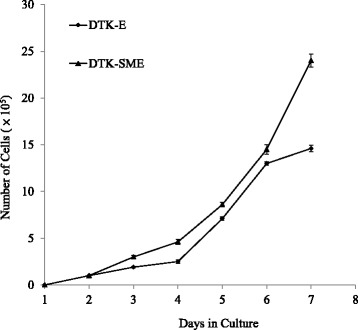
Growth curves of DTK-E and DTK-SME cell lines. Symbols represent the mean of triplicate samples and the data are expressed as the mean ± SE.
Expression profile of CMT genes and proteins
We used western blot analysis with specific antibodies to investigate the individual protein expression profiles of the two CMT cell lines (Table 1). Both CMT cell lines expressed the low-molecular weight cytokeratin AE1 and vimentin, cytokeratin 7 (CK7), and heat shock protein 27 (HSP27) (Figure 4A, B). In addition, DTK-SME exhibited a higher level of HSP27 expression than DTK-E cells. In reverse transcription-polymerase chain reaction (RT-PCR) assays, the CK7 and HSP27 gene expression profiles were the same as their respective protein expression profiles; both DTK-E and DTK-SME cells expressed CK7 and HSP27 genes (Figure 4C).
Table 1.
Overview of the sources and dilutions of the primary antibodies used in this study
| Antibody | Source | Clone | Dilution |
|---|---|---|---|
| AE1 | NeoMarkers | AE1 (mouse monoclonal) | 1:1000 |
| Vimentin | NeoMarkers | V-9 (mouse monoclonal) | 1:1000 |
| CK 7 | Santa Cruz | 5F282 (mouse monoclonal) | 1:1000 |
| Hsp27 | GeneTex | GTX25579 (rabbit polyclonal) | 1:1000 |
| Caspase 3 | NeoMarkers | CPP (rabbit polyclonal) | 1:1000 |
| β-actin | Sigma | AC-15 (rabbit polyclonal) | 1:5000 |
Figure 4.
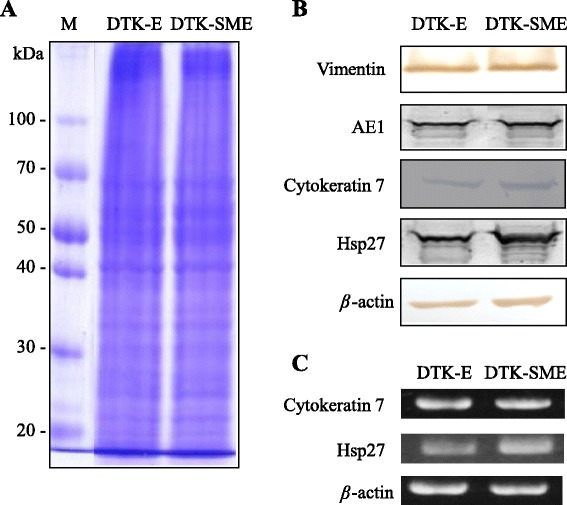
Expression profiles of protein markers in the DTK-E and DTK-SME cell lines. (A) Western blot analysis of total proteins in cell lysates. Lane 1, pre-stained protein marker; Lane 2, DTK-E; Lane 3, DTK-SME. (B) Protein expression in DTK-E and DTK-SME cell lines detected with antibodies to vimentin, cytokeratin 7 (CK7), low-molecular weight cytokeratin AE1, and heat shock protein 27 (HSP27). (C) RT-PCR analysis of CK7 and HSP27 gene expression in DTK-E and DTK-SME cell lines. β-actin served as an internal control.
Tumorigenicity of established CMT cell lines
We performed xenografts to examine the tumorigenicity of the two CMT cell lines. At passage 100, each cell line was injected subcutaneously into five 8-week-old athymic mice (BALB/c nu/nu). A tumor mass was first observed at the injection site in all mice injected with DTK-E and DTK-SME cells at 1 week post-injection; at 8 weeks, the DTK-SME and DTK-E groups exhibited masses of 1.8–2.2 cm and 0.5–0.7 cm in diameter, respectively (Figure 5).
Figure 5.
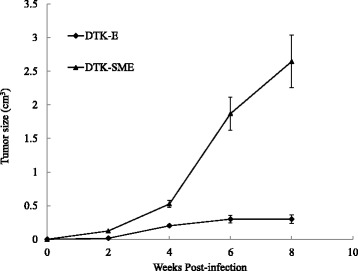
Tumor growth curves of DTK-E and DTK-SME cells in tumorigenicity tests. Cells were subcutaneously injected into nude mice and monitored for 8 weeks. The tumor volumes were estimated every 2 weeks and calculated, and the data are expressed as the mean ± SE.
Doxorubicin (DOX)-induced dose-dependent growth suppression, apoptosis, and caspase-3 activation in CMT cells
Next, we investigated the susceptibility of the two CMT cell lines to DOX. Cells were treated with DOX and cell growth and apoptosis were analyzed at 24 h, 48 h, and 72 h. We found that DOX (1000 nM) was able to promote apoptosis and inhibit proliferation in a dose- and time-dependent manner in DTK-E cells, but not in DTK-SME cells (Figures 6 and 7). Further quantitative analysis revealed that 72-h exposures to 100 nM and 1000 nM DOX resulted in 2% and 58% mortality, respectively, in DTK-E cells (Figure 6). In addition, DOX (1000 nM) treatment caused DNA fragmentation in DTK-E cells at 72 h (Figure 7A). In contrast, no DNA fragmentation was observed at 72 h in DTK-SME cells. Because activation of caspase-3 is a hallmark of apoptosis, we examined caspase-3 activation in DOX-treated (1000 nM) CMT cells (Figure 7B). DOX induced the time-dependent cleavage of procaspase-3 into its active caspase-3 form in DTK-E cells, but not in DTK-SME cells.
Figure 6.
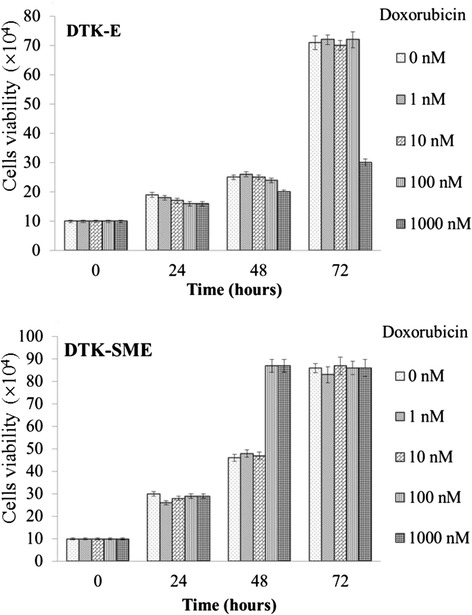
Effects of DOX on DTK-E and DTK-SME cell proliferation. Cells were treated with escalating doses of DOX (1, 10, 100, and 1000 nM) and incubated for 24 h, 48 h, and 72 h. Cell viability was determined every 24 h. Cell numbers were averaged over triplicate samples, and the data are expressed as the mean ± SE.
Figure 7.
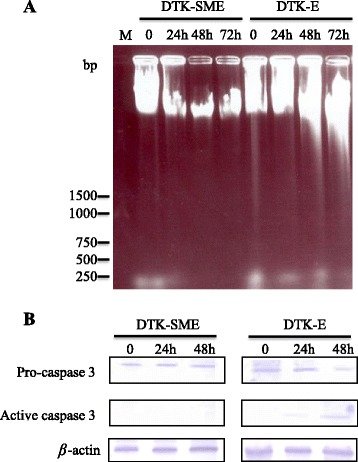
DOX-induced apoptosis and caspase-3 activation in DTK-E and DTK-SME cells. (A) Agarose gel electrophoresis of total cellular DNA showed DOX-induced DNA fragmentation (apoptosis) in CMT cells. Cells were incubated with 1000 mM DOX for 24 h, 48 h, and 72 h. M: DNA marker. (B) Western blots show DOX-induced activation of caspase-3 in CMT cells. Cells were incubated with 1000 nM DOX for 24 h and 48 h.
Discussion
In the present study, we established and characterized two new CMT cell lines derived from a single malignant tumor: DTK-E and DTK-SME. DTK-E cells showed a large, flat, epithelial-like morphology, while DTK-SME cells showed a rounded epithelial-like morphology. Both DTK-E and DTK-SME cells exhibited a high nuclear-cytoplasmic ratio, which is often associated with malignancy in tumor cells. Consistent with this finding, xenografts of injected DTK-E and DTK-SME cells developed into prominent solid tumors within 2 weeks, and the tumor mass of DTK-SME was larger than that of DTK-E after the 8-week study.
The human breast contains a branching ductal network composed of two epithelial cell types: an inner layer of polarized luminal epithelial cells and an outer layer of myoepithelial cells. Breast cancer arises mainly in the luminal epithelial compartment of the terminal duct lobular units [17,18]. Several proteins have been identified as important cell markers for investigating tumor pathophysiology. Cytokeratins (CKs) are critical markers of epithelial differentiation [19]. The distribution of CKs is tissue-specific, and this feature facilitates the determination of tumor origin [20–22]. One low-molecular weight cytokeratin subtype, CK7, is a marker of luminal epithelial cells in both human and canine mammary tissues [23]. In a study of 435 cases, expression of CK7 was associated with up to 96% of human breast epithelial neoplasms [24]. Other studies have also reported a high incidence of CK7 (56%) in canine mammary gland carcinomas [25]. In this study, we showed that DTK-E and DTK-SME expressed CK7 and possessed high tumorigenicity. These results suggest that the DTK-E and DTK-SME cell lines could have originated directly from the tumor cells in the surgical specimen.
Vimentin is an intermediate filament protein expressed in mesenchymal cells [26]. Vimentin expression in invasive breast carcinomas is generally considered to indicate the epithelial-to-mesenchymal transition or myoepithelial histogenesis. Vimentin-positive breast tumors have been described as mostly malignant, highly invasive, and chemoresistant [27]. Interestingly, the DTK-E and DTK-SME cell lines established in this study were vimentin-positive and displayed tumorigenicity in mice.
HSP27 is a member of the small heat shock protein family of molecular chaperones. It plays an essential role in protein folding and protects cells from stress-induced damage [28,29]. In general, the healthy canine (and human) mammary gland exhibits little or no HSP27 expression [30–32]. In human breast cancer tumor cells, overexpression of HSP27 was associated with accelerated cancer progression [33], increased anchorage-independent growth [34], increased invasiveness [35,36], and resistance to chemotherapeutic drugs [34,37–39]. In patients, elevated expression of HSP27 in human breast cancer tissues has been shown to indicate poor prognosis and a low survival rate [40,41]. Therefore, increased expression of HSP27 in human breast cancers might affect the prognosis and treatment outcome [42].
DOX is an antitumor drug that can promote tumor cell apoptosis. Currently, DOX is widely used to treat a broad spectrum of cancers. In vitro studies of cancer cells have shown that HSP27 expression protects against apoptosis and confers resistance to DOX [38,43,44]. Interestingly, we also observed an association between increased HSP27 expression and resistance to DOX (Figures 6 and 7). Indeed, HSP27 expression was elevated in both the DTK-E and DTK-SME cell lines, although the expression level was higher in DTK-SME cells. Accordingly, DTK-SME cells were more resistant to DOX-mediated apoptosis and tumor masses were larger than those of DTK-E cells. The correlation between HSP27 and CMT malignancy currently remains unclear; thus, these two CMT cell lines with different tumorigenicities may serve as a valuable cell model for future studies of the roles of HSP27 in tumorigenicity and chemoresistance.
Conclusion
We established and characterized two new CMT cell lines with diverse tumorigenic properties. DTK-E cells were large, flat, and epithelial-like, while DTK-SME cells showed a rounded epithelial-like morphology and possessed pile up. Both DTK-E and DTK-SME cells were vimentin+/AE1+/CK7+/HSP27+ and tumorigenic in nude mice. These two cell lines also exhibited differential susceptibility to DOX-mediated apoptosis. Based on their diverse malignancy, tumorigenicity, protein marker profiles, and drug susceptibility, these newly established CMT cell lines represent useful cell models for basic tumor biology studies and for developing antitumor drugs to treat both human and canine breast carcinomas.
Methods
Tumor specimen
The tumor tissues used in this study were obtained from client-owned dogs with the consents of the owners acquired prior to sample collection. The handling of animals was practiced following a high standard of veterinary care. A tumor specimen was surgically excised from the mammary tissue of an 8-year-old mixed-breed female dog. Fresh neoplastic masses from the specimen were placed in primary culture, and the remaining samples were fixed in 10% formalin for histopathology. A histological examination confirmed the diagnosis of mammary carcinoma. Tumor specimens consisted of highly proliferating, luminal epithelial and fibroblastic stromal cells; they also had glandular structures with irregular lumens embedded in fibrous stroma.
Establishment of cell lines
A single tumor specimen was washed with RPMI 1640 medium (Sigma; St. Louis, MO, USA) that contained 500 IU/mL penicillin and streptomycin. The tissue was minced with scissors and transferred to a 60-mm-diameter tissue culture dish (Nunc; Waltham, MA USA) containing 10 mL of 0.25% trypsin solution (0.25% trypsin and 0.2% ethylenediaminetetraacetic acid [EDTA] in phosphate-buffered saline [PBS]). The tissue suspension was gently agitated with a magnetic stir bar at 4°C for 1 h. The resulting supernatant was transferred to a new tube. An equal volume of complete RPMI 1640 medium, containing 10% FBS (Hyclone Laboratories; Logan, UT, USA) and 100 IU/mL penicillin and streptomycin was added and mixed well. The mixture was centrifuged at 180 × g for 5 min, and the pelleted cells were resuspended in fresh, complete medium. The cells were then seeded into 25-cm2 culture flasks and incubated with complete medium at 37°C in a humidified atmosphere of 95% air and 5% CO2. Observations were performed daily with a phase-contrast microscope. When cultured cells reached 90% confluence, they were washed with 2 mM EDTA in PBS, dispersed in the 0.25% w/v trypsin solution, and placed in a new flask at a density of 1 × 105 cells/mL. After 20 passages, cells were stored in RPMI 1640 medium containing 20% FBS and 10% dimethyl sulfoxide (Sigma). In addition, at passage 20, different cell types were subcloned from the parental cell line with a limiting-dilution method.
Electron microscopy
After 100 passages, cultured CMT cells were harvested, fixed with 2.5% glutaraldehyde in 0.1 M cacodylate buffer, and placed at 4°C overnight. After fixation, the cells were treated with 1% osmium tetroxide at 4°C for 2 h, washed in cacodylate buffer, dehydrated in graded acetone solutions, and embedded in Spur’s low-viscosity resin. The embedded specimen was sectioned with a Reichert-Jung Ultracut E ultramicrotome (Nussloch, Germany), and the sections were stained with 2% uranyl acetate (EMS; Fort Washington, PA, USA) in distilled water for 1 min. Images were acquired with a JEOL JEM 2000 EXII transmission electron microscope (JEOL Ltd.; Tokyo, Japan).
Growth assay
To investigate the growth of CMT cells, after 100 passages, 1 × 105 cells were seeded in 25-cm2 tissue culture flasks and incubated in complete medium at 37°C. After the cells were detached with trypsinization, they were stained with trypan blue in triplicate flasks and counted with a hemocytometer. The average number of cells was calculated and recorded.
Western blot analysis
CMT cells were collected and lysed with 100 μL sample buffer that contained 100 mM Tris–HCl buffer, pH 6.8, 4% sodium dodecyl sulfate (SDS), 0.07% β-mercaptoethanol, 20% glycerol, and 0.2% bromophenol blue. The crude cell lysate proteins were electrophoresed on a 12% SDS-acrylamide gel and then stained with Coomassie Brilliant Blue R-250 for protein profiling or transferred to polyvinylidene fluoride membranes for western blot analysis. The membranes were blocked with 5% skim milk, incubated with primary antibodies (Table 1) for 2 h, and subsequently incubated with alkaline phosphatase-conjugated secondary antibodies. Membranes were developed with 5-bromo-4-chloro-3-indolyl phosphate and 4-nitro-blue tetrazolium chloride as substrates.
Total RNA isolation and RT-PCR
Total RNA was isolated from the cells using TRIzol reagent (Invitrogen; Paisley, Scotland, UK) according to the manufacturer’s protocol. After digestion with RNase-free DNase (New England Biolabs; Beverly, MA, USA), 2 μg of total RNA was reverse-transcribed into first-strand cDNA using random primers according to the Reverse Transcriptase Kit (Roche; Penzberg, Germany) protocol. The PCR was carried out in a volume of 50 μL containing 2 μL cDNA, 0.5 μM forward primer, 0.5 μM reverse primer, 2.5 μM dNTP, 1× PCR buffer, and 2.5 U Taq DNA polymerase (Viogene; Taipei, Taiwan). The primers for CK7 were as follows: Forward-5′- CAGGTGCGCCTGAGCTCG-3′, Reverse-5′-GCGGTAGGTGGCGATCTC-3′. PCR was carried out under the following conditions: 10 min at 95°C; 35 cycles of 1 min at 95°C, 1 min at 56°C, and 1 min at 72°C; and 10 min at 72°C. The primers for HSP27 were as follows: Forward-5′-ATGACCGAGCGCCGAGTGCCC-3′, Reverse-5′-CTTGGCTCCAGACTGCTCCGA-3′. PCR was carried out under the following conditions: 5 min at 95°C; 35 cycles of 45 s at 95°C, 45 s at 54°C, and 1 min at 72°C; and 10 min at 72°C. The primers for β-actin were as follows: Forward-5′-CTGGGACGACATGGAGAA-3′, Reverse-5′-GAGTACTTGCGCTCAGG-3′. PCR was carried out under the following conditions: 5 min at 95°C; 35 cycles of 45 s at 95°C, 45 s at 55°C, and 45 s at 72°C; and 10 min at 72°C. The PCR products were electrophoresed on a 1.2% agarose TAE-buffered gel and stained with ethidium bromide.
Tumorigenicity
Animal handling and experimental procedures were approved by the Institutional Animal Care and Use Committee of National Ilan University (Reference number: No. 103-10). After 100 passages, CMT cells (1 × 107) were subcutaneously inoculated into the backs of 8-week-old athymic BALB/c nude mice. Five nude mice were used to test each cell line. Physiological data of the mice were recorded weekly, and tumor volumes were estimated every 14 days and calculated as described previously [45]. Mice were sacrificed 56 days after inoculation. Results are expressed as the mean tumor volume.
Susceptibility to doxorubicin
After 100 passages, CMT cells (1 × 105) were seeded into 25-cm2 tissue culture flasks in triplicate. After 24-h incubation, cells were treated with the antitumor drug DOX for 72 h. Mock control cells were treated with water. After the treatment, cells were trypsinized, harvested, stained with 0.4% trypan blue, and counted with a hemocytometer under a light microscope. The average number of cells in triplicate flasks was calculated and recorded.
DNA ladder assay
After DOX (1000 nM) treatment, cells were collected and lysed at 55°C for 1 h in 500 μL lysis buffer that contained 100 mM NaCl, 10 mM Tris (pH 8.0), 25 mM EDTA, 0.5% SDS, 200 μg/mL DNase-free proteinase K (Sigma), and 50 μg/mL DNase-free RNase (Sigma). DNA was extracted with phenol/chloroform, precipitated with 100% ethanol, and resuspended in double-distilled water. DNA samples were then separated on l.6% agarose gels and stained with ethidium bromide.
Acknowledgements
This study was supported by grant nos. NSC 91-2320-B-001-049-B and 94-2815-C-197-013-B from the National Science Council, Taiwan.
Abbreviations
- CMT
Canine mammary tumor
- CK
Cytokeratin
- CK7
Cytokeratin 7
- HSP27
Heat shock protein 27
- DOX
Doxorubicin
- FBS
Fetal bovine serum
- EDTA
Ethylenediaminetetraacetic acid
- PBS
Phosphate-buffered saline
- SDS
Sodium dodecyl sulfate
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
TZH, cell lines establishment, growth assay; CJL, western blot analysis; YLH and YHC, susceptibility to doxorubicin assay; CTL, tumor specimen and electron microscopy; CYC and YSL, establishment of cell lines, tumorigenicity and apoptosis assays, statistical analysis, manuscript and figure preparation. All authors read and approved the final manuscript.
Authors’ information
TZH: Master degree student (Department and Graduate Institute of Veterinary Medicine, National Taiwan University, Taipei, Taiwan). CJL: Professor (Department of Nursing, Chang Gung University of Science and Technology, Taoyuan, Taiwan). YLH: Master degree student (Department of Biotechnology and Animal Science, National Ilan University, Yilan, Taiwan).
YHC: Professor (Department of Biotechnology and Animal Science, National Ilan University, Yilan, Taiwan).
CTL: Professor (Department and Graduate Institute of Veterinary Medicine, National Taiwan University, Taipei, Taiwan).
CYC: Associate research fellow (Institute of Cellular and Organismic Biology, Academia Sinica, Taipei, Taiwan).
YSL: Associate professor (Department of Biotechnology and Animal Science, National Ilan University, Yilan, Taiwan).
Contributor Information
Yen-Ling Hsiao, Email: mock_ring@yahoo.com.tw.
Tai-Zu Hsieh, Email: aooqoo10336@hotmail.com.
Chian-Jiun Liou, Email: ccliu@gw.cgust.edu.tw.
Yeong-Hsiang Cheng, Email: yhcheng@niu.edu.tw.
Chung-Tien Lin, Email: ctlin@ntu.edu.tw.
Chi-Yao Chang, Email: ychang@gate.sinica.edu.tw.
Yu-Shen Lai, Email: yslai@niu.edu.tw.
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Misdorp W. Tumors of the Mammary Gland. In: Meuten DJ, editor. Tumors in Domestic Animals. 4. Iowa, USA: Iowa State Press; 2002. pp. 575–606. [Google Scholar]
- 3.Brodey RS, Goldschmidt MH, Roszel JR. Canine mammary gland neoplasms. J Am Anim Hosp Assoc. 1983;19:61–90. [Google Scholar]
- 4.Martin PM, Cotard M, Mialot JP, Andre F, Raynaud JP. Animal models for hormone-dependent human breast cancer. Relationship between steroid receptor profiles in canine and feline mammary tumors and survival rate. Cancer Chemother Pharmacol. 1984;12:13–17. doi: 10.1007/BF00255902. [DOI] [PubMed] [Google Scholar]
- 5.Nieto A, Pena L, Perez-Alenza MD, Sanchez MA, Flores JM, Castano M. Immunohistologic detection of estrogen receptor alpha in canine mammary tumors: clinical and pathologic associations and prognostic significance. Vet Pathol. 2000;37:239–247. doi: 10.1354/vp.37-3-239. [DOI] [PubMed] [Google Scholar]
- 6.MacEwen EG. Spontaneous tumors in dogs and cats: models for the study of cancer biology and treatment. Cancer Metastasis Rev. 1990;9:125–136. doi: 10.1007/BF00046339. [DOI] [PubMed] [Google Scholar]
- 7.Louzada S, Adega F, Chaves R. Defining the sister rat mammary tumor cell lines HH-16 cl.2/1 and HH-16.cl.4 as an in vitro cell model for Erbb2. PLoS One. 2012;7:e29923. doi: 10.1371/journal.pone.0029923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gazdar AF, Girard L, Lockwood WW, Lam WL, Minna JD. Lung cancer cell lines as tools for biomedical discovery and research. J Natl Cancer Inst. 2010;102:1310–1321. doi: 10.1093/jnci/djq279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Vargo-Gogola T, Rosen JM. Modelling breast cancer: one size does not fit all. Nat Rev Cancer. 2007;7:659–672. doi: 10.1038/nrc2193. [DOI] [PubMed] [Google Scholar]
- 10.Chang CY, Chiou PP, Chen WJ, Li YH, Yiu JC, Cheng YH, Chen SD, Lin CT, Lai YS. Assessment of the tumorigenesis and drug susceptibility of three new canine mammary tumor cell lines. Res Vet Sci. 2010;88:285–293. doi: 10.1016/j.rvsc.2009.08.006. [DOI] [PubMed] [Google Scholar]
- 11.Nakatsu N, Yoshida Y, Yamazaki K, Nakamura T, Dan S, Fukui Y, Yamori T. Chemosensitivity profile of cancer cell lines and identification of genes determining chemosensitivity by an integrated bioinformatical approach using cDNA arrays. Mol Cancer Ther. 2005;4:399–412. doi: 10.1158/1535-7163.MCT-04-0234. [DOI] [PubMed] [Google Scholar]
- 12.Ruhe JE, Streit S, Hart S, Wong CH, Specht K, Knyazev P, Knyazeva T, Tay LS, Loo HL, Foo P, Wong W, Pok S, Lim SJ, Ong H, Luo M, Ho HK, Peng K, Lee TC, Bezler M, Mann C, Gaertner S, Hoefler H, Iacobelli S, Peter S, Tay A, Brenner S, Venkatesh B, Ullrich A. Genetic alterations in the tyrosine kinase transcriptome of human cancer cell lines. Cancer Res. 2007;67:11368–11376. doi: 10.1158/0008-5472.CAN-07-2703. [DOI] [PubMed] [Google Scholar]
- 13.Pfragner R, Behmel A, Hoger H, Beham A, Ingolic E, Stelzer I, Svejda B, Moser VA, Obenauf AC, Siegl V, Haas O, Niederle B. Establishment and characterization of three novel cell lines - P-STS, L-STS, H-STS - derived from a human metastatic midgut carcinoid. Anticancer Res. 2009;29:1951–1961. [PubMed] [Google Scholar]
- 14.Shoemaker RH. The NCI60 human tumour cell line anticancer drug screen. Nat Rev Cancer. 2006;6:813–823. doi: 10.1038/nrc1951. [DOI] [PubMed] [Google Scholar]
- 15.Van Staveren WC, Solis DY, Hebrant A, Detours V, Dumont JE, Maenhaut C. Human cancer cell lines: experimental models for cancer cells in situ? For cancer stem cells? Biochim Biophys Acta. 2009;1795:92–103. doi: 10.1016/j.bbcan.2008.12.004. [DOI] [PubMed] [Google Scholar]
- 16.Kao J, Salari K, Bocanegra M, Choi YL, Girard L, Gandhi J, Kwei KA, Hernandez-Boussard T, Wang P, Gazdar AF, Minna JD, Pollack JR. Molecular profiling of breast cancer cell lines defines relevant tumor models and provides a resource for cancer gene discovery. PLoS One. 2009;4:e6146. doi: 10.1371/journal.pone.0006146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gudjonsson T, Adriance MC, Sternlicht MD, Petersen OW, Bissell MJ. Myoepithelial cells: their origin and function in breast morphogenesis and neoplasia. J Mammary Gland Biol Neoplasia. 2005;10:261–272. doi: 10.1007/s10911-005-9586-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ronnov-Jessen L, Petersen OW, Bissell MJ. Cellular changes involved in conversion of normal to malignant breast: importance of the stromal reaction. Physiol Rev. 1996;76:69–125. doi: 10.1152/physrev.1996.76.1.69. [DOI] [PubMed] [Google Scholar]
- 19.Abd El-Rehim DM, Pinder SE, Paish CE, Bell J, Blamey RW, Robertson JF, Nicholson RI, Ellis IO. Expression of luminal and basal cytokeratins in human breast carcinoma. J Pathol. 2004;203:661–671. doi: 10.1002/path.1559. [DOI] [PubMed] [Google Scholar]
- 20.Gupta RK, Naran S, Dowle C, Simpson JS. Coexpression of vimentin, cytokeratin and S-100 in monomorphic adenoma of salivary gland; value of marker studies in the differential diagnosis of salivary gland tumours. Cytopathology. 1992;3:303–309. doi: 10.1111/j.1365-2303.1992.tb00052.x. [DOI] [PubMed] [Google Scholar]
- 21.Harbaum L, Pollheimer MJ, Kornprat P, Lindtner RA, Schlemmer A, Rehak P, Langner C. Keratin 7 expression in colorectal cancer–freak of nature or significant finding? Histopathology. 2011;59:225–234. doi: 10.1111/j.1365-2559.2011.03694.x. [DOI] [PubMed] [Google Scholar]
- 22.Trask DK, Band V, Zajchowski DA, Yaswen P, Suh T, Sager R. Keratins as markers that distinguish normal and tumor-derived mammary epithelial cells. Proc Natl Acad Sci USA. 1990;87:2319–2323. doi: 10.1073/pnas.87.6.2319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Henderson D, Weber K. Immuno-electron microscopical identification of the two types of intermediate filaments in established epithelial cells. Exp Cell Res. 1981;132:297–311. doi: 10.1016/0014-4827(81)90106-3. [DOI] [PubMed] [Google Scholar]
- 24.Chu P, Wu E, Weiss LM. Cytokeratin 7 and cytokeratin 20 expression in epithelial neoplasms: a survey of 435 cases. Mod Pathol. 2000;13:962–972. doi: 10.1038/modpathol.3880175. [DOI] [PubMed] [Google Scholar]
- 25.Espinosa de los Monteros A, Fernandez A, Millan MY, Rodriguez F, Herraez P, Martin de las Mulas J. Coordinate expression of cytokeratins 7 and 20 in feline and canine carcinomas. Vet Pathol. 1999;36:179–190. doi: 10.1354/vp.36-3-179. [DOI] [PubMed] [Google Scholar]
- 26.Hellmen E, Lindgren A. The expression of intermediate filaments in canine mammary glands and their tumors. Vet Pathol. 1989;26:420–428. doi: 10.1177/030098588902600507. [DOI] [PubMed] [Google Scholar]
- 27.Korsching E, Packeisen J, Liedtke C, Hungermann D, Wulfing P, Van Diest PJ, Brandt B, Boecker W, Buerger H. The origin of vimentin expression in invasive breast cancer: epithelial-mesenchymal transition, myoepithelial histogenesis or histogenesis from progenitor cells with bilinear differentiation potential? J Pathol. 2005;206:451–457. doi: 10.1002/path.1797. [DOI] [PubMed] [Google Scholar]
- 28.De Maio A. Heat shock proteins: facts, thoughts, and dreams. Shock. 1999;11:1–12. doi: 10.1097/00024382-199901000-00001. [DOI] [PubMed] [Google Scholar]
- 29.Morimoto RI, Kline MP, Bimston DN, Cotto JJ. The heat-shock response: regulation and function of heat-shock proteins and molecular chaperones. Essays Biochem. 1997;32:17–29. [PubMed] [Google Scholar]
- 30.Ciocca DR, Oesterreich S, Chamness GC, McGuire WL, Fuqua SA. Biological and clinical implications of heat shock protein 27,000 (Hsp27): a review. J Natl Cancer Inst. 1993;85:1558–1570. doi: 10.1093/jnci/85.19.1558. [DOI] [PubMed] [Google Scholar]
- 31.Devaja O, King RJ, Papadopoulos A, Raju KS. Heat-shock protein 27 (HSP27) and its role in female reproductive organs. Eur J Gynaecol Oncol. 1997;18:16–22. [PubMed] [Google Scholar]
- 32.Romanucci M, Marinelli A, Sarli G, Della Salda L. Heat shock protein expression in canine malignant mammary tumours. BMC Cancer. 2006;6:171. doi: 10.1186/1471-2407-6-171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Love S, King RJ. A 27 kDa heat shock protein that has anomalous prognostic powers in early and advanced breast cancer. Br J Cancer. 1994;69:743–748. doi: 10.1038/bjc.1994.140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Oesterreich S, Weng CN, Qiu M, Hilsenbeck SG, Osborne CK, Fuqua SA. The small heat shock protein hsp27 is correlated with growth and drug resistance in human breast cancer cell lines. Cancer Res. 1993;53:4443–4448. [PubMed] [Google Scholar]
- 35.Lemieux P, Oesterreich S, Lawrence JA, Steeg PS, Hilsenbeck SG, Harvey JM, Fuqua SA. The small heat shock protein hsp27 increases invasiveness but decreases motility of breast cancer cells. Invasion Metastasis. 1997;17:113–123. [PubMed] [Google Scholar]
- 36.Grzegrzolka J, Kurnol K, Piotrow P, Pula B, Kobierzycki C, Piotrowska A, Jablonska K, Wojnar A, Rys J, Dziegiel P, Podhorska-Okolow M. Hsp-27 expression in invasive ductal breast carcinoma. Folia Histochem Cytobiol. 2012;50:527–533. doi: 10.5603/FHC.2012.0074. [DOI] [PubMed] [Google Scholar]
- 37.Diaz-Chavez J, Fonseca-Sanchez MA, Arechaga-Ocampo E, Flores-Perez A, Palacios-Rodriguez Y, Dominguez-Gomez G, Marchat LA, Fuentes-Mera L, Mendoza-Hernandez G, Gariglio P, López-Camarillo C. Proteomic profiling reveals that resveratrol inhibits HSP27 expression and sensitizes breast cancer cells to doxorubicin therapy. PLoS One. 2013;8:e64378. doi: 10.1371/journal.pone.0064378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hansen RK, Parra I, Lemieux P, Oesterreich S, Hilsenbeck SG, Fuqua SA. Hsp27 overexpression inhibits doxorubicin-induced apoptosis in human breast cancer cells. Breast Cancer Res Treat. 1999;56:187–196. doi: 10.1023/A:1006207009260. [DOI] [PubMed] [Google Scholar]
- 39.Shi P, Wang MM, Jiang LY, Liu HT, Sun JZ. Paclitaxel-doxorubicin sequence is more effective in breast cancer cells with heat shock protein 27 overexpression. Chin Med J (Engl) 2008;121:1975–1979. [PubMed] [Google Scholar]
- 40.Ciocca DR, Calderwood SK. Heat shock proteins in cancer: diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones. 2005;10:86–103. doi: 10.1379/CSC-99r.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Thanner F, Sutterlin MW, Kapp M, Rieger L, Morr AK, Kristen P, Dietl J, Gassel AM, Muller T. Heat shock protein 27 is associated with decreased survival in node-negative breast cancer patients. Anticancer Res. 2005;25:1649–1653. [PubMed] [Google Scholar]
- 42.Wei L, Liu TT, Wang HH, Hong HM, Yu AL, Feng HP, Chang WW. Hsp27 participates in the maintenance of breast cancer stem cells through regulation of epithelial-mesenchymal transition and nuclear factor-kappaB. Breast Cancer Res. 2011;13:R101. doi: 10.1186/bcr3042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Di X, Shiu RP, Newsham IF, Gewirtz DA. Apoptosis, autophagy, accelerated senescence and reactive oxygen in the response of human breast tumor cells to adriamycin. Biochem Pharmacol. 2009;77:1139–1150. doi: 10.1016/j.bcp.2008.12.016. [DOI] [PubMed] [Google Scholar]
- 44.Havasi A, Li Z, Wang Z, Martin JL, Botla V, Ruchalski K, Schwartz JH, Borkan SC. Hsp27 inhibits Bax activation and apoptosis via a phosphatidylinositol 3-kinase-dependent mechanism. J Biol Chem. 2008;283:12305–12313. doi: 10.1074/jbc.M801291200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Noel A, De Pauw-Gillet MC, Purnell G, Nusgens B, Lapiere CM, Foidart JM. Enhancement of tumorigenicity of human breast adenocarcinoma cells in nude mice by matrigel and fibroblasts. Br J Cancer. 1993;68:909–915. doi: 10.1038/bjc.1993.453. [DOI] [PMC free article] [PubMed] [Google Scholar]