

Abstract
In recent times, human cell-based assays are gaining attention in assessments of immunomodulatory effects of chemicals. In the study here, the possible effects of cypermethrin and mancozeb on lymphocyte proliferation and proinflammatory (tumor necrosis factor (TNF-) α) and immunoregulatory cytokine (interferon- (IFN-) γ, interleukins (IL) 2, 4, 6, and 10) formation in vitro were investigated. Human peripheral blood mononuclear cells (PBMC) were isolated and exposed for 6 hr to noncytotoxic doses (0.45–30 µM) of cypermethrin or mancozeb in the presence of activating rat S9 fraction. Cultures were then further incubated for 48 or 72 hr in fresh medium containing phytohemagglutinin (10 µg/mL) to assess, respectively, effects on cell proliferation (BrdU-ELISA method) and cytokine formation (flow cytometric bead immunoassays). Mancozeb induced dose-dependent increases in lymphocyte proliferation, inhibition of production of TNFα and the TH2 cytokines IL-6 and IL-10, and an increase in IFNγ (TH1 cytokine) production (at least 2-fold compared to control); mancozeb also induced inhibition of IL-4 (TH2) and stimulated IL-2 (TH1) production, albeit only in dose-related manners for each. In contrast, cypermethrin exposure did not cause significant effects on proliferation or cytokine profiles. Further studies are needed to better understand the functional significance of our in vitro findings.
1. Introduction
Pesticide-associated immune dysfunction has gained regulatory and public attention in the past 20 years due to the wide use of these agents in agriculture, industries, and domestic purposes. Chronic exposure to pesticides increases the risk of immunomodulation [1–5] and the onset of lymphoid neoplasms [6] and leukemias [7]. Cypermethrin and mancozeb are widely used in agriculture, households, and industries due to their “low” toxicity in mammals and short environmental persistence [8–11]. Earlier studies have indicated the immunomodulatory effects such as decrease in antiovalbumin titer of blood sera, autologous rosette formation of T-lymphocytes [12–14], lymphocyte transformation rate, and an increase in neutrophil phagocytosis rate [15] from exposure to cypermethrin in animal models. Similarly, occupational exposure to mancozeb had shown significant increase in T-cell functional response such as mitogen-induced proliferation and a decrease in TNF-alpha [16, 17]. Although, this information is limited to few functional properties, it clearly indicates that the proliferation and TH1/TH2 cytokine production, the key indicators, play a major role in etiology of several immunological disorders.
There is a wide range of experimental protocols and guidelines that have been validated for assessing the chemical induced immune dysfunctions. These assays include local lymph node assay (LLNA), guinea pig maximization test (GPMT) for the dermal allergic potential, T-cell dependent antibody response (TDAR) assay for immunosuppression, and popliteal lymph node assay (PLNA) for autoimmune reactions. Nevertheless, these assays are based on animal models and having limitations such as extrapolating the animal data to humans, false positives or negatives, and several others in evaluating the immune system. Furthermore, the 3Rs (reduce, refine, and replace) strongly recommend using alternative approaches for evaluating the immunocompetence of chemicals and xenobiotics.
Proliferation and cytokine production are two key functions of immune cells and, as such, are important endpoints to examine during any evaluation of immunotoxic potential of a given xenobiotic [18–20]. Mitogen-induced proliferation of lymphocytes is often a preferable assay that correlates with the status of cell-mediated immunity in a host [21] after exposure to a xenobiotic. Similarly, cytokines, as regulators of immune function, are sensitive indicators of immunomodulation in an exposed host [22–25].
As immune functions are mediated by several cytokines/chemokines (that, in turn, are influenced by bacterial or viral infections, drugs, and/or exposure to environmental or workplace agents), it is necessary to analyze a panel of cytokines to better understand effects of a given toxicant on host immunocompetence [26]. Evaluations of proinflammatory and immunoregulatory agents also provide valuable information during assessments of chemical induced immunomodulation. In addition, because maintaining homeostasis of T-helper (TH) type 1 and TH2 cytokines is critical to immunocompetence [27, 28], deviations in levels of either of these or the balance between these two can provide strong evidence to understand immunopathologies induced by chemical exposures [24] and also help in developing better testing regimens for evaluating the immune system.
The present study was designed to evaluate the potential effects of two widely used pesticides, cypermethrin and mancozeb, on functional properties of immune system through lymphocyte proliferation and TH1/TH2 cytokine production in human PBMC.
2. Materials and Methods
2.1. Chemicals
Cypermethrin (>99%) and mancozeb (>95%) were purchased from Sigma (St. Louis, MO). Stock solutions of these chemicals were made in dimethyl sulfoxide (DMSO; Sigma) and stored at −80°C. Stock solutions were diluted in DMSO at the desired concentrations before further dilution in culture medium. Final culture levels of DMSO never exceeded 0.1%.
2.2. Peripheral Blood Mononuclear Cells
Upon obtaining informed consent, peripheral blood was collected from individual male volunteers 26–35 years of age. Selection of volunteers was based on the criteria listed by Indian Council of Medical Research (ICMR-2004), briefly, only those volunteers who are nonsmokers, are nonalcoholics, and have had no recent history of illness as certified by a medical practitioner. The study was approved by the Institutional Ethics Committee of the International Institute of Biotechnology and Toxicology, in its meeting held on April 2, 2011.
Blood was collected into heparinized tubes and mononuclear cells were isolated using Ficoll-Hypaque (ρ = 1.077 g/mL) density gradient centrifugation at 400 ×g for 30 min. The buffy coat containing mononuclear cells was isolated, transferred to a fresh centrifuge tube, and washed twice with PBS (using ≈3 vol of collected buffy coat each time). The final cell pellet containing peripheral blood mononuclear cells (PBMC) was resuspended to a final level of 1-2 × 106 cells/mL in RPMI 1640 medium supplemented with L-glutamine, 10% heat-inactivated fetal bovine serum, 100 U penicillin/mL, and 0.1 mg streptomycin/mL (all Gibco, Paisley, UK).
2.3. Cytotoxicity Assessment
Preliminary cytotoxicity studies were performed using PBMC from 2 donors/test agent to assess biovariance. PBMC (105 cells/well, 24-well plate) were exposed for 24 hr to serial doses of cypermethrin or mancozeb in the presence of a metabolic activator (i.e., rat liver S9 fraction (Moltox, Boone, NC)). Immediately before use, 10% S9 mix containing 15% S9 fraction was added to the reaction medium. Based on trypan blue (Gibco, Paisley, UK) dye exclusion, doses that caused >10% of cytotoxicity were excluded from further analysis.
2.4. Culture Set-Up/PBMC Exposure
To assess biological variance, assays were performed (in triplicate) using PBMC from three donors with each chemical separately. Cultures of PBMC (105 cells/well, 24-well plate) were exposed to noncytotoxic doses of cypermethrin (1.87, 3.75, 7.50, 15, or 30 μM) or mancozeb (0.45, 0.93, 1.87, 3.75, or 7.50 μM) or to medium containing solvent (DMSO) or medium alone for 6 hr. All cultures contain freshly prepared S9 mix. At the end of the exposure, the medium was removed from each well and fresh medium containing phytohemagglutinin (10 μg PHA/mL; Gibco) mitogen was added. The cells were then incubated at 37°C under 5% CO2 and 95% humidity for 48 or 72 hr to assess, respectively, cell proliferation and cytokine release.
2.5. Cell Proliferation-BrdU ELISA Method
Cell proliferation was measured upon completion of the 48 hr incubation period as noted above using a BrdU-ELISA kit (Roche Diagnostics, Mannheim, Germany) as per manufacturer instructions. In brief, kit-provided BrdU labeling solution (20 μL) was added to each well and the plate was incubated at 37°C overnight. Thereafter, the cells were centrifuged at 300 ×g for 10 min, the labeling solution was removed, and the plate then was dried at 60°C for 1 hr before the cells were fixed by addition of FixDenat (200 μL/well) solution and incubation at room temperature for 15 min. Antibody conjugate (anti-BrdU-POD solution, 100 μL/well) was then added and the plate was incubated at room temperature for 90 min. The cells were then washed twice with PBS (200 μL) and kit-provided substrate solution (100 μL) was added to each well. The plates were left at room temperature for 20 min and the absorbance in each well was then measured at 370 nm in an automated plate reader (Awareness Technology, Inc., Palm city, FL). Sets of blank (100 μL culture medium alone) and control wells were included in each experiment.
2.6. TH1 and TH2 Cytokine Analysis
After the 72 hr incubation with PHA, culture supernatants from each well were harvested and stored at −80°C until analysis. Levels of select proinflammatory (tumor necrosis factor (TNF)-α) and immunoregulatory (TH1/TH2) cytokines (e.g., interferon- (IFN-) γ, interleukin (IL-) 2, IL-4, IL-6, and IL-10) in samples were estimated using a flow cytometric bead immunoassay (according to manufacturer protocols) in a FACSAria-II system (BD Biosciences, San Jose, CA). The cytometer was calibrated using standard beads before analysis and standard curves were generated for each cytokine using known concentrations (provided in kit) ranging from 20 to 5000 pg/mL.
2.7. Data Analysis
Cell proliferation was expressed in terms of percentage growth, with 100% corresponding to the values seen with the control wells. Triplicate data was averaged and expressed as mean ± SE for three individual experiments (n = 3) conducted from PBMC of three different donors for each compound. Cytokine analyses were performed using FCAP Array software (version 3.0). The concentration of each analyte in a sample was extrapolated from a calibration curve generated in parallel and that was modeled by a five-parameter log-logistic curve (5PL) for each analyte, against log-transformed median fluorescence intensity (MFI) versus concentrations.
Cytokine quantities obtained with the FCAP Array software were compared with concurrent solvent controls and statistical analysis performed by two-way ANOVA followed by a Newman-Keuls Test post hoccomparison using Prism software (v6.03 for Windows, GraphPad, San Diego, CA). For all of the comparisons, a 0.05 α-type error (P < 0.05) was considered as significant.
3. Results
3.1. Effects on Proliferation
A dramatic effect on PHA-induced lymphocyte proliferation was observed with mancozeb; the percentage proliferation (compared to that seen with the solvent control) was increased in a dose-dependent manner at the doses tested (0.45–7.5 μM; Figure 1(a)). At 7.5 μM, the average value (as percent of control) had reached 174.5 (±5.2)% (Table 1). In contrast, cypermethrin caused nominal nonsignificant changes in proliferation at the doses tested (1.87–30 μM; Figure 1(b)) relative to the control values.
Figure 1.

Proliferation-BrdU ELISA assay. Proliferation rate in mitogen (PHA-) stimulated PBMC cultures. (a) Mancozeb; (b) cypermethrin. ∗P < 0.05; ∗∗P < 0.01 versus zero control. For each donor, statistical analyses performed across the doses showed that values were consistently significantly different as a function of dose in the mancozeb study.
Table 1.
Effect on lymphocyte proliferation (%) and TH1/TH2 cytokine production (pg/mL) in PBMC cultures. Values shown are average ± SD from three volunteers (each was conducted in triplicate).
(a).
| Mancozeb (µM) | Proliferation | TNF-α | IFN-γ | IL-2 | IL-10 | IL-6 | IL-4 |
|---|---|---|---|---|---|---|---|
| Solv. | 100.0 ± 0.0 | 696.4 ± 60.2 | 1303.4 ± 175.1 | 56.6 ± 10.9 | 37.7 ± 7.2 | 597.8 ± 63.6 | 19.7 ± 3.1 |
| 0.45 | 105.4 ± 2.5 | 657.4 ± 69.3 | 1912.3 ± 127.6 | 109.4 ± 12.9 | 41.0 ± 7.3 | 628.8 ± 69.1 | 22.2 ± 4.9 |
| 0.9 | 119.9 ± 4.5 | 621.1 ± 66.9 | 2401.0 ± 113.0 | 138.1 ± 23.5 | 39.8 ± 8.3 | 551.2 ± 86.1 | 19.6 ± 4.6 |
| 1.87 | 138.4 ± 6.8 | 424.7 ± 21.7 | 2617.8 ± 165.6 | 152.9 ± 22.2 | 33.6 ± 8.7 | 390.6 ± 100.4 | 17.4 ± 3.1 |
| 3.75 | 162.5 ± 4.6 | 238.6 ± 35.8 | 2974.6 ± 411.6 | 163.7 ± 26.6 | 24.2 ± 9.6 | 217.5 ± 72.2 | 14.5 ± 2.7 |
| 7.5 | 174.5 ± 5.2 | 129.1 ± 25.2 | 3539.2 ± 103.7 | 167.3 ± 28.6 | 15.1 ± 4.6 | 107.7 ± 34.9 | 10.0 ± 1.7 |
(b).
| Cypermethrin (µM) | Proliferation | TNF-α | IFN-γ | IL-2 | IL-10 | IL-6 | IL-4 |
|---|---|---|---|---|---|---|---|
| Solv. | 100.0 ± 0.0 | 426.6 ± 44.4 | 2555.9 ± 281.0 | 127.4 ± 6.2 | 31.9 ± 5.8 | 614.5 ± 90.8 | 17.9 ± 3.2 |
| 1.87 | 102.7 ± 1.4 | 412.0 ± 22.8 | 2450.4 ± 352.0 | 138.4 ± 8.9 | 28.9 ± 4.9 | 629.0 ± 31.0 | 16.2 ± 2.8 |
| 3.75 | 102.7 ± 3.4 | 409.6 ± 54.3 | 2421.8 ± 244.5 | 148.6 ± 9.8 | 25.9 ± 4.4 | 601.2 ± 27.0 | 15.3 ± 2.5 |
| 7.5 | 101.1 ± 1.2 | 397.8 ± 59.0 | 2316.2 ± 310.4 | 145.3 ± 7.9 | 25.9 ± 2.8 | 565.9 ± 30.4 | 13.6 ± 1.8 |
| 15 | 100.0 ± 1.1 | 382.4 ± 57.0 | 2222.3 ± 336.4 | 149.6 ± 13.4 | 24.3 ± 3.2 | 538.2 ± 33.9 | 13.0 ± 1.4 |
| 30 | 98.0 ± 0.8 | 365.1 ± 42.5 | 2134.9 ± 364.2 | 159.6 ± 12.8 | 23.0 ± 3.3 | 527.3 ± 34.1 | 12.1 ± 1.2 |
3.2. Effect on Proinflammatory TNFα Production
Dose-dependent decreases in PHA-stimulated TNFα release were evident in the mancozeb-exposed cultures (Figure 2(a)). Maximal inhibition of TNFα production was seen at the highest test dose and 7.5 μM (average of the three donor populations = 129.1 (±25.2) pg/mL; Table 1) was more than 2-fold below the control culture levels (average = 696.4 (±60.2) pg/mL; Table 1). Although slight changes were observed in the cypermethrin-exposed cultures, these were very modest even at the highest test dose, 30 μM (average = 365.1 (±42.5) pg/mL; Table 1) relative to the control value (average = 426.6 (±44.4) pg/mL; Table 1) (Figure 2(b)).
Figure 2.
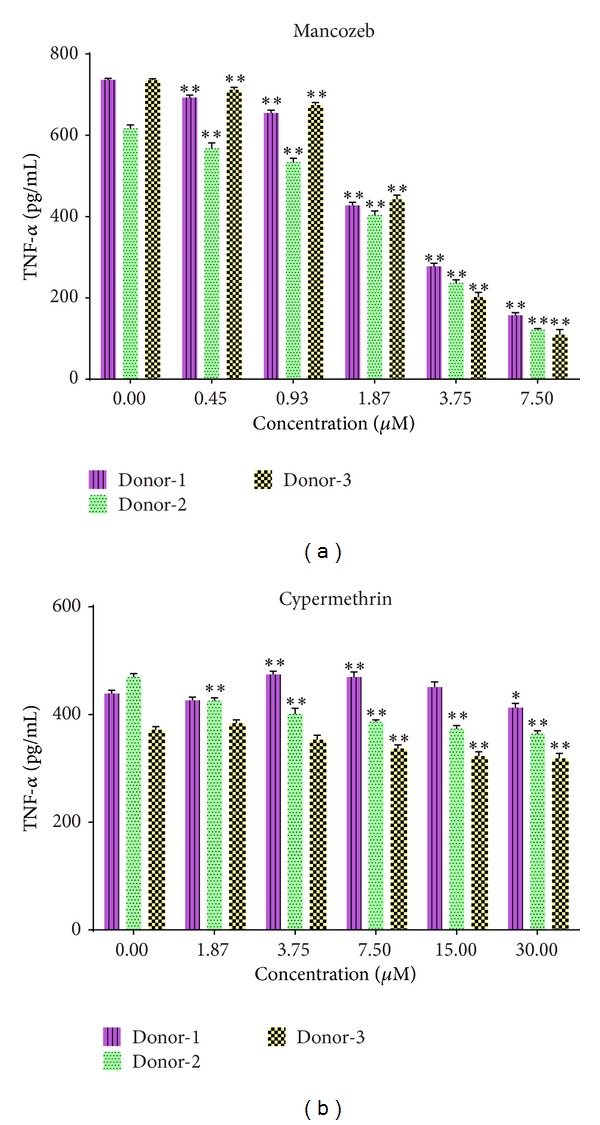
Proinflammatory response. Effects on levels of proinflammatory cytokine TNFα in culture supernatants of PBMC. (a) Mancozeb. (b) Cypermethrin. Values shown are mean (±SE) from triplicate samples conducted with PBMC from three volunteers. ∗P < 0.05; ∗∗P < 0.01 versus zero control. For each donor, statistical analyses performed across the doses showed that values were consistently significantly different as a function of dose in the mancozeb study.
3.3. Effect on TH1 (IL-2 and IFNγ) Cytokines
Mancozeb induced formation and release of both IL-2 and IFNγ by the PBMC. Maximal induction of cytokine production (a 2-fold change) was observed at highest concentration, that is, 7.5 μM. IFNγ production increased dose-dependently to (average) 3539.2 (±103.7) pg/mL from 1303.4 (±175.1) pg/mL for the control cells (Table 1, Figure 3(a)); IL-2 levels increased (albeit in non-dose-dependent manner) to 167.4 (±28.6) pg/mL from 56.6 (±10.9) pg/mL (Table 1, Figure 3(c)). Changes in levels of these TH1 cytokines in the cypermethrin-exposed cultures were weak relative to control cell values. Interestingly, IFNγ production seemed to decrease, albeit insignificantly, from (average) 2555.9 (±281.0) pg/mL down to 2134.9 (±364.2) pg/mL at the 30 μM dose (Table 1, Figure 3(b)); IL-2 production increased from 127.4 (±6.2) pg/mL to 159.6 (±12.8) pg/mL (Table 1, Figure 3(d)).
Figure 3.
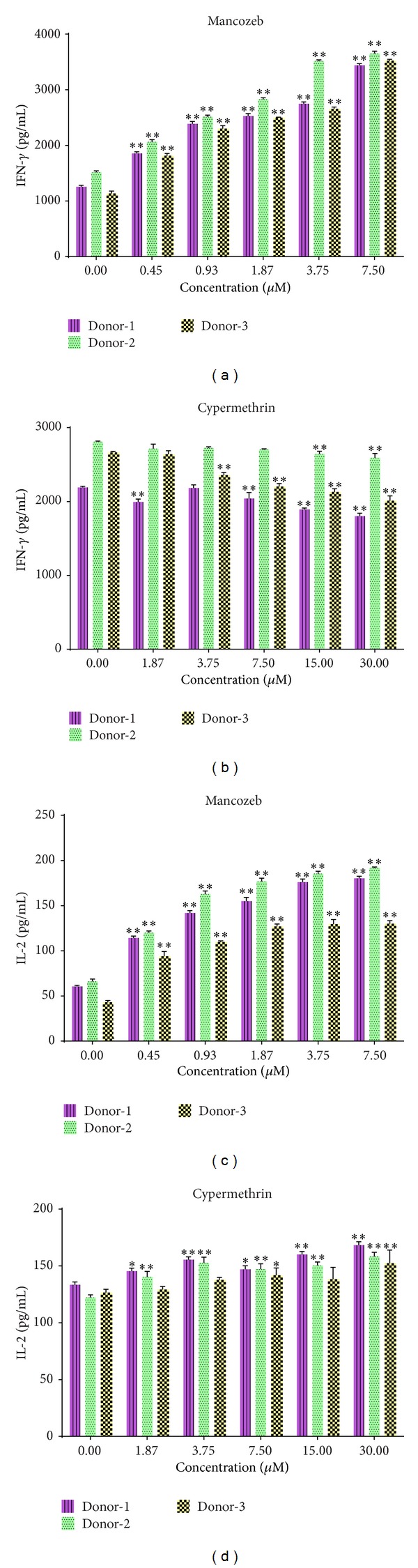
TH1 cytokine profiles. Effect on TH1 cytokines IFNγ and IL-2 production in PBMC cultures. (a) Mancozeb IFNγ. (b) Cypermethrin IFNγ. (c) Mancozeb IL-2. (d) Cypermethrin IL-2. Values shown are mean (±SE) from triplicate samples conducted with PBMC from three volunteers. ∗P < 0.05; ∗∗P < 0.01 versus zero control. For each donor, statistical analyses performed across the doses showed that resultant IFNγ values were consistently significantly different as a function of dose in the mancozeb study.
3.4. Effect on TH2 (IL-4, IL-6, and IL-10) Cytokines
TH2 cytokine production by PBMC was significantly inhibited with increasing doses of mancozeb. Maximal inhibition of cytokine production was observed at highest 7.5 μM concentration, with IL-4 production dropping to (average) 10.0 (±1.7) pg/mL from 19.7 (±3.1) pg/mL (Figure 4(a), Table 1), IL-6 (dose-dependently) to 107.7 (±34.9) pg/mL from 597.8 (±63.6) pg/mL (Figure 4(b), Table 1), and IL-10 (dose-dependently) to 15.1 (±4.6) pg/mL from 37.8 (±7.2) pg/mL (Figure 4(c), Table 1). On the other hand, while there were changes in IL-4 levels with at least two donor PBMC populations exposed to cypermethrin, the average values at 30 μM were comparable to the controls, though the levels had declined from 17.9 pg IL-4/mL (±3.2) to 12.1 (±1.2) pg/mL (Figure 4(d), Table 1). While the impact from this pesticide on IL-6 production by the cells was slight IL-6 levels dropped from 614.5 (±90.8) pg/mL to 527.3 (±34.1) pg/mL; (Figure 4(e), Table 1), a clear trend toward inhibition of IL-10 formation was evident for at least two donor PBMC populations. However, the average values at 30 μM were not significantly different from the control (31.9 (±5.8) pg/mL versus 23.0 (±3.3) pg/mL; Figure 4(f), Table 1).
Figure 4.

TH2 cytokine profiles. Effect on TH2 cytokines IL-4, IL-6, and IL-10 production in PBMC cultures. Mancozeb: (a) IL-4; (b) IL-6; and (c) IL-10. Cypermethrin: (d) IL-4; (e) IL-6; and (f) IL-10. Values shown are mean (±SE) from triplicate samples conducted with PBMC from three volunteers. ∗P < 0.05; ∗∗P < 0.01 versus zero control. For each donor, statistical analyses performed across the doses showed that resultant IL-6 values were consistently significantly different as a function of dose in the mancozeb study. Further, in that same study, IL-10 values were also consistently significantly different as a function of dose for doses of ≥0.93 μM.
4. Discussion
Exposures to pesticides as environmental contaminants or in the workplace elicit a wide variety of adverse effects on the human immune system and often lead to various immune-based/related disorders [2, 29]. Despite the number of studies available for assessing the immune functions, in recent years, there has been a growing interest in considering the role of proinflammatory and immunoregulatory cytokines in toxic responses to chemical exposure; this has, in turn, resulted in an increased consideration in measures of these parameters when developing risk assessment strategies [30]. Homeostasis of the levels of TH1 and TH2, as well as proinflammatory, cytokines is crucial. Deviations in levels of either of these or the balance between these two can provide strong evidence to understand immunopathologies induced by chemical exposures [24, 31].
Immunomodulatory effects of two widely used pesticides, that is, cypermethrin and mancozeb, in PBMC were evaluated in the present study. The in vitro data here indicated that mancozeb exerted potential immunomodulatory effects characterized by (i) increased cell proliferative responses to PHA stimulation; (ii) significant reductions in PHA-induced TNFα release; and (iii) significant alterations in TH1 and TH2 cytokine profiles. These outcomes reflect the same types of effects of mancozeb on mitogen-induced proliferation and on TNFα and IL-2 levels that had been reported earlier [16, 17, 32, 33]. On the other hand, here, cypermethrin only caused slight changes in the cytokine profiles or on mitogen-induced proliferation.
Numerous in vivo studies have reported dose-dependent decreases in hematologic endpoints, such as erythrocyte counts and packed cell volumes that were accompanied by significant changes in lymphocyte, monocyte, and total leukocyte counts as a result of exposure to cypermethrin [34–36] or mancozeb [17, 37]. The results in the present in vitro study correlated fairly well with those in vivo findings and indicated that the mancozeb directly affected immune cells. However, cypermethrin exposure did not show any effect on lymphocyte proliferation and cytokine production. This clearly shows that the pesticides will act differently on immune cells and have different effects.
The range of in vitro doses tested here was also in accordance with those earlier in vivo studies. In those studies, significant effects on hemato-/immunologic parameters were seen with mancozeb doses that ranged from 250 to 1500 mg/kg BW [38] or with cypermethrin (through oral or intraperitoneal routes) at doses ranging from 25 to 300 mg/kg BW [34, 35]. Still, while these in vitro studies demonstrated effects from direct exposure of PBMC to each pesticide, questions about relevance of the doses used (0.45–30 μM of parent compounds) seem to persist. Occupational exposure to pyrethroids and ethylenebisdithiocarbamates often results in increases in body burdens of the toxicants that are, in turn, reflected at mg levels of select metabolites of the parent agent (for cypermethrin, 3-phenoxy-benzoate; for mancozeb, ethylene thiourea) in the blood/urine of the workers [17, 39, 40]. As such, at even just a single mg of each metabolite, this would yield corresponding levels of 4.7 (3-phenoxybenzoate) and 9.8 (ethylene thiourea) μM in the human blood/urine of the exposed workers. Thus, the doses of cypermethrin or mancozeb used in the current experiment were likely to have been on par with levels of the parent + metabolite found in workers routinely exposed to either pesticide. Nevertheless, the true relationship between cumulative quantities of metabolites produced by continuous exposures to these pesticides in workers can and should be estimated by others using mathematical toxicokinetic modeling to better estimate at what doses these pesticides would be unquestionably relevant in this type of in vitro studies performed here.
5. Conclusions
The present study demonstrates that pesticides that are indiscriminately used might increase the risk for immunomodulation in an exposed host. As such, this also reflects the need for continued evaluation of the immunotoxic potentials of these and other types of commonly encountered occupational/environmental chemicals. As the data here also show, in vitro studies using freshly obtained human cells (i.e., PBMC) to assess agent-induced changes in endpoints like lymphocyte proliferation and TH1/TH2 cytokine production will certainly contribute to a better understanding of the effects of xenobiotics on immune function and also increase the arsenal that investigators can use when designing/developing testing regimens to assess such effects.
Acknowledgments
This work was supported by the International Institute of Biotechnology And Toxicology. The authors thank the management of International Institute of Biotechnology And Toxicology for funding and supporting the research and the staff for donating blood samples. This work would have not been possible without the moral support from Dr. Srivatsa Prakhya; special thanks are due to him.
Conflict of Interests
The authors declare no conflict of interests. The authors alone are responsible for the content of this paper.
References
- 1.Repetto R, Baliga S. Pesticides and the Immune System; Public Health Risks. Washington, DC, USA: World Resources Institute; 1996. Trends and patterns of pesticide use; pp. 3–8. [Google Scholar]
- 2.Vial T, Nicolas B, Descotes J. Clinical immunotoxicity of pesticides. Journal of Toxicology and Environmental Health. 1996;48(3):215–229. doi: 10.1080/009841096161294. [DOI] [PubMed] [Google Scholar]
- 3.Stiller-Winkler R, Hadnagy W, Leng G, Straube E, Idel H. Immunological parameters in humans exposed to pesticides in the agricultural environment. Toxicology Letters. 1999;107(1–3):219–224. doi: 10.1016/s0378-4274(99)00050-8. [DOI] [PubMed] [Google Scholar]
- 4.Vine MF, Stein L, Weigle K, et al. Effects on the immune system associated with living near a pesticide dump site. Environmental Health Perspectives. 2000;108(12):1113–1124. doi: 10.1289/ehp.001081113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nishimoto S, Kanda K, Okabe M, Akiyama K, Kakinuma Y. Abnormal response induced by pesticides on mammalian immune system. Interdisciplinary Studies on Environ-mental Chemistry-Environmental Research in Asia. 2009:211–217. [Google Scholar]
- 6.Zakerinia M, Namdari M, Amirghofran S. The relationship between exposure to pesticides and the occurrence of lymphoid neoplasm. Iranian Red Crescent Medical Journal. 2012;14(6):337–344. [PMC free article] [PubMed] [Google Scholar]
- 7.Ma X, Buffler PA, Gunier RB, et al. Critical windows of exposure to household pesticides and risk of childhood leukemia. Environmental Health Perspectives. 2002;110(9):955–960. doi: 10.1289/ehp.02110955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hayes WJ, Laws ER, editors. Handbook of Pesticide Toxicology. Vol. 3. New York, NY, USA: Academic Press; 1990. (Classes of Pesticides). [Google Scholar]
- 9.Lorgue G, Lechenet J, editors. Clinical Veterinary Toxicology. London, UK: Blackwell Science; 1996. [Google Scholar]
- 10.Descotes J. Immunotoxicology of Drugs and Chemicals. An Experimental and Clinical Approach. Vol. 1. Principles and Methods of Immunotoxicology. Amsterdam, The Netherlands: Elsevier Science; 2004. Regulatory aspects of immunotoxicity evaluation; pp. 257–268. [Google Scholar]
- 11.Hasan HO. Fungicides and Their Effects on Animals, Fungicides. 2010. [Google Scholar]
- 12.Desi I, Varga L, Dobronyi I, Szklenarik G. Immunotoxicological investigation of the effects of a pesticide; cypermethrin. Archives of Toxicology. 1985;8:305–309. doi: 10.1007/978-3-642-69928-3_58. [DOI] [PubMed] [Google Scholar]
- 13.Desi I, Dobronyi I, Varga L. Immuno-, neuro-, and general toxicologic animal studies on a synthetic pyrethroid: cypermethrin. Ecotoxicology and Environmental Safety. 1986;12(3):220–232. doi: 10.1016/0147-6513(86)90014-x. [DOI] [PubMed] [Google Scholar]
- 14.WHO (World Health Organization) International Program on Chemical Safety: WHO Task Group Meeting on Environmental Health Criteria for Cypermethrin . Cypermethrin. Vol. 82. Geneva, Switzerland: United Nations Environment Program, the International Labor Organization, and World Health Organization; 1989. (Environmental Health Criteria). [Google Scholar]
- 15.Liu P, Song X, Yuan W, et al. Effects of cypermethrin and methyl parathion mixtures on hormone levels and immune functions in Wistar rats. Archives of Toxicology. 2006;80(7):449–457. doi: 10.1007/s00204-006-0071-7. [DOI] [PubMed] [Google Scholar]
- 16.Colosio C, Barcellini W, Maroni M, et al. Immunomodulatory effects of occupational exposure to mancozeb. Archives of Environmental Health. 1996;51(6):445–451. doi: 10.1080/00039896.1996.9936044. [DOI] [PubMed] [Google Scholar]
- 17.Corsini E, Birindelli S, Fustinoni S, et al. Immunomodulatory effects of the fungicide Mancozeb in agricultural workers. Toxicology and Applied Pharmacology. 2005;208(2):178–185. doi: 10.1016/j.taap.2005.02.011. [DOI] [PubMed] [Google Scholar]
- 18.Koeper L-M, Vohr H-W. Functional assays are mandatory for a correct prediction of immunotoxic properties of compounds in vitro . Food and Chemical Toxicology. 2009;47(1):110–118. doi: 10.1016/j.fct.2008.10.025. [DOI] [PubMed] [Google Scholar]
- 19.Lankveld DP, van Loveren H, Baken KA, Vandebriel RJ. In vitro testing for direct immunotoxicity: state of the art. Methods in Molecular Biology. 2010;598:401–423. doi: 10.1007/978-1-60761-401-2_26. [DOI] [PubMed] [Google Scholar]
- 20.Luebke R. Immunotoxicant screening and prioritization in the 21st century. Toxicologic Pathology. 2012;40(2):294–299. doi: 10.1177/0192623311427572. [DOI] [PubMed] [Google Scholar]
- 21.Descotes J. Methods of evaluating immunotoxicity. Expert Opinion on Drug Metabolism and Toxicology. 2006;2(2):249–259. doi: 10.1517/17425255.2.2.249. [DOI] [PubMed] [Google Scholar]
- 22.Cohen MD, Schook LB, Oppenheim JJ, Freed BM, Rodgers KE. Symposium overview: alterations in cytokine receptors by xenobiotics. Toxicological Sciences. 1999;48(2):163–169. doi: 10.1093/toxsci/48.2.163. [DOI] [PubMed] [Google Scholar]
- 23.House RV. Theory and practice of cytokine assessment in immunotoxicology. Methods. 1999;19(1):17–27. doi: 10.1006/meth.1999.0823. [DOI] [PubMed] [Google Scholar]
- 24.Duramad P, Tager IB, Holland NT. Cytokines and other immunological biomarkers in children's environmental health studies. Toxicology Letters. 2007;172(1-2):48–59. doi: 10.1016/j.toxlet.2007.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Corsini E, House RV. Evaluating cytokines in immunotoxicity testing. Methods in Molecular Biology. 2010;598:283–302. doi: 10.1007/978-1-60761-401-2_20. [DOI] [PubMed] [Google Scholar]
- 26.Ringerike T, Ullerås E, Völker R, et al. Detection of immunotoxicity using T-cell based cytokine reporter cell lines (“Cell Chip”) Toxicology. 2005;206(2):257–272. doi: 10.1016/j.tox.2004.08.014. [DOI] [PubMed] [Google Scholar]
- 27.Romagnani S. T-cell subsets (Th1 versus Th2) Annals of Allergy, Asthma and Immunology. 2000;85(1):9–18. doi: 10.1016/S1081-1206(10)62426-X. [DOI] [PubMed] [Google Scholar]
- 28.Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annual Review of Immunology. 1989;7:145–173. doi: 10.1146/annurev.iy.07.040189.001045. [DOI] [PubMed] [Google Scholar]
- 29.Fritschi L, Benke G, Hughes AM, et al. Occupational exposure to pesticides and risk of non-Hodgkin’s lymphoma. The American Journal of Epidemiology. 2005;162(9):849–857. doi: 10.1093/aje/kwi292. [DOI] [PubMed] [Google Scholar]
- 30.Kooijman R, Devos S, Hooghe-Peters E. Inhibition of in vitro cytokine production by human peripheral blood mononuclear cells treated with xenobiotics: implications for the prediction of general toxicity and immunotoxicity. Toxicology in Vitro. 2010;24(6):1782–1789. doi: 10.1016/j.tiv.2010.07.007. [DOI] [PubMed] [Google Scholar]
- 31.Lesko LJ, Atkinson AJJ. Use of biomarkers and surrogate endpoints in drug development and regulatory decision making: criteria, validation, strategies. Annual Review of Pharmacology and Toxicology. 2001;41:347–366. doi: 10.1146/annurev.pharmtox.41.1.347. [DOI] [PubMed] [Google Scholar]
- 32.Chung A-H, Pyo M-Y. Effects of mancozeb on the activities of murine peritoneal macrophages in vitro and in vivo . Archives of Pharmacal Research. 2005;28(1):100–105. doi: 10.1007/BF02975143. [DOI] [PubMed] [Google Scholar]
- 33.Corsini E, Viviani B, Birindelli S, et al. Molecular mechanisms underlying mancozeb-induced inhibition of TNF-alpha production. Toxicology and Applied Pharmacology. 2006;212(2):89–98. doi: 10.1016/j.taap.2005.07.002. [DOI] [PubMed] [Google Scholar]
- 34.Sayim F, Karabay Yavaşoğlu NÜ, Uyanikgil Y, Aktuğ H, Yavaşoğlu A, Turgut M. Neurotoxic effects of cypermethrin in Wistar rats: a haematological, biochemical and histopathological study. Journal of Health Science. 2005;51(3):300–307. [Google Scholar]
- 35.Kamal S, M A, Rizvi F, Siddique M, Sadeeq-ur-Rehaman S. Effect of cypermethrin on clinico-haematological parameters in rabbits. Pakistan Veterinary Journal. 2007;27(4):171–175. [Google Scholar]
- 36.Nair RR, Abraham MJ, Nair ND, Lalithakunjamma CR, Aravindakshan CM. Hematological and biochemical profile in sublethal toxicity of cypermethrin in rats. International Journal of Biological and Medical Research. 2010;1(4):211–214. [Google Scholar]
- 37.Cox RH. HLA 417-416. Vienna, Va, USA: Hazleton Labs; 1986. Three-month dietary toxicity study in dogs with mancozeb. Submitted to WHO by Rohm and Haas Company, Spring House, Pa, USA (as Rohm and Haas Report No. 86RC-7) [Google Scholar]
- 38.Myoung-yun P, Cheong A. Effects of subacute oral administration of mancozeb on immunopathological parameters and splenocyte proliferation in mice. Environmental Health and Toxicology. 2004;19:367–373. [Google Scholar]
- 39.Leng G, Gries W. Methods in biotechnology. In: Martínez Vidal JL, Garrido Frenich A, editors. Pesticide Protocols. Vol. 19. Totowa, NJ, USA: Humana Press; 1999. [Google Scholar]
- 40.Lu D, Wang D, Feng C, et al. Urinary concentrations of metabolites of pyrethroid insecticides in textile workers, Eastern China. Environment International. 2013;60:137–144. doi: 10.1016/j.envint.2013.08.004. [DOI] [PubMed] [Google Scholar]