

Abstract
The structure of the adduct formed in the reaction between Aubipyc, a cytotoxic organogold(III) compound, and the model protein hen egg white lysozyme (HEWL) has been solved by X-ray crystallography. It emerges that Aubipyc, after interaction with HEWL, undergoes reduction of the gold(III) center followed by detaching of the cyclometalated ligand; the resulting naked gold(I) ion is found bound to the protein at Gln121. A direct comparison between the present structure and those previously solved for the lysozyme adducts with other gold(III) compounds demonstrates that coordinated ligands play a key role in the protein–metallodrug recognition process. Structural data support the view that gold(III)-based antitumor prodrugs are activated through metal reduction.
Keywords: Gold-based drugs, metallodrugs, protein−ligand interactions
Knowledge of the interactions of medicinal gold compounds with proteins is crucial to define their mechanism of action; indeed these compounds are believed to be activated upon protein interaction1 and then to exert their biological actions by hitting and selectively damaging a few protein targets. Despite numerous papers reporting on the interactions of gold compounds with proteins, only few structural studies have appeared so far, and very few Au binding sites have been characterized in proteins.2−5
By using hen egg white lysozyme (HEWL) and bovine pancreatic ribonuclease (RNase A) as model proteins, we previously solved the X-ray structures of a few protein adducts with gold compounds of medicinal interest.6,7 These two proteins have often been used to study protein modifications induced by metal-based bioactive agents.6−15 We found that the reaction of three structurally different medicinal gold compounds, i.e., AuSac2, Auoxo6, and Au2phen (Figure 1), with HEWL invariantly leads to the formation of the same type of metal–protein adduct bearing a gold(I) ion tightly anchored to His15 with Cl– as the second ligand.6 In the same work, we tried to crystallize the complex between HEWL and Aubipyc (Figure 1), i.e., [(bipydmb-H)Au(OH)][PF6] (where bipydmb-H = deprotonated 6-(1,1-dimethylbenzyl)-2,2′-bipyridine), a cyclometalated gold(III) complex exhibiting remarkable anticancer properties in vitro,16 but attempts to obtain a stable adduct in the solution used to grow HEWL–Aubipyc crystals (1.0 M NaCl and 0.050 M sodium acetate, pH 4.0–4.4) failed.6 Notably, Au(I) ions bind to His residues even in the complex formed between RNase A and Auoxo6.7
Figure 1.

Structure of AuSac2, Auoxo6, Au2phen, and Aubipyc.
Here we report the crystal structure of the HEWL–gold(I) adduct, solved using X-ray diffraction data collected on crystals of HEWL obtained in the presence of Aubipyc (1:10 protein to metal ratio) via the hanging drop method. These crystals have been grown using a reservoir solution of 0.6 M NaNO3, 0.1 M sodium acetate pH 4.4, and 20% ethylene glycol.
We have also obtained crystals of HEWL in the presence of Au2phen and Auoxo6 exactly under the same experimental conditions, but inspection of electron density map did not reveal the formation of protein–gold adducts (data not shown).
Crystals have been flash-cooled in a cold gaseous nitrogen stream, without any cryoprotectants, after mother liquor removal (Figure 2). This methodology can yield improved low-temperature crystalline order,17 higher resolution of X-ray diffraction,18 and reduced mosaicity.
Figure 2.

Crystal of HEWL–Aubipyc mounted in a nylon loop and flash-cooled in the absence of mother liquid and cryoprotectants.
A data set from crystals of HEWL–Au(I) has been collected at 1.80 Å resolution at Institute of Biostructures and Bioimages, Naples, Italy. Each diffraction image was taken by oscillating 1.0°; the integrated intensities were merged and scaled using HKL2000.19 The results of the data collection are summarized in Table S1, Supporting Information. The structure was solved by difference Fourier method using PDB 4J1A(20) without water and ligands as starting model and subjected to several cycle of restrained refinement using Refmac5.721 and rebuilding in Coot.22
The final model, which includes 1235 non-hydrogen atoms, refines to an R factor of 0.181 (Rfree = 0.244). The average B factor for all atoms of the structure is 31.2 Å2. A full list of the refinement statistics is reported in Table S2, Supporting Information. This model has been deposited in the Protein Data Bank under the accession code 4OOT.
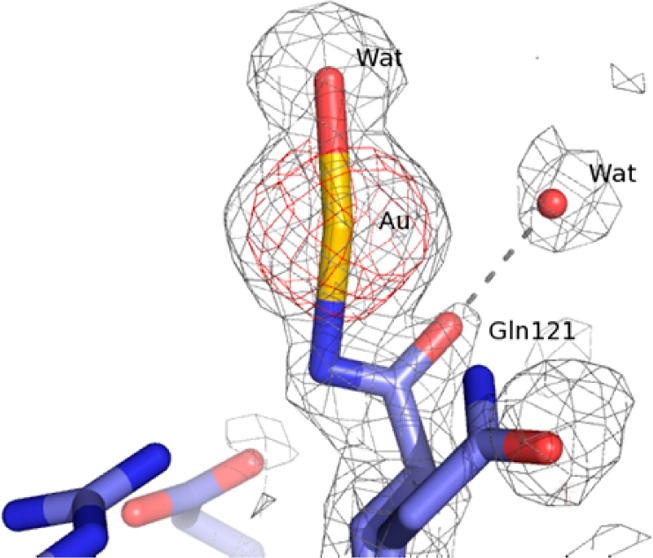
The overall conformation of the protein in the complex (Figure 3) is very similar to that of the metal-free protein (CA root-mean-square deviation from the HEWL structure (PDB code 2VB1)23 is 0.79 Å) implying that the structural effects of gold coordination are highly localized. The gold atom (occupancy 0.6) is clearly detected by inspection of the electron density maps (Figure 4).
Figure 3.
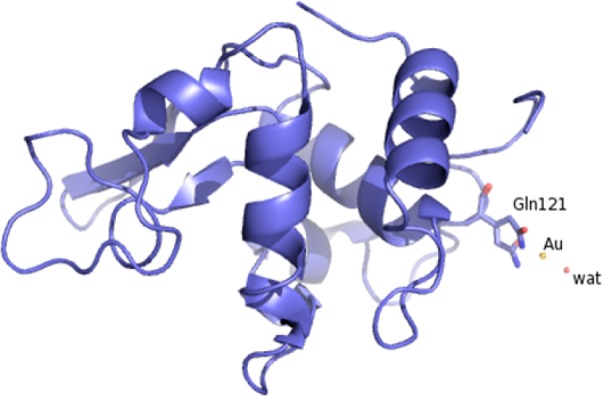
Ribbon diagram of HEWL with gold ion bound to Gln121.
Figure 4.
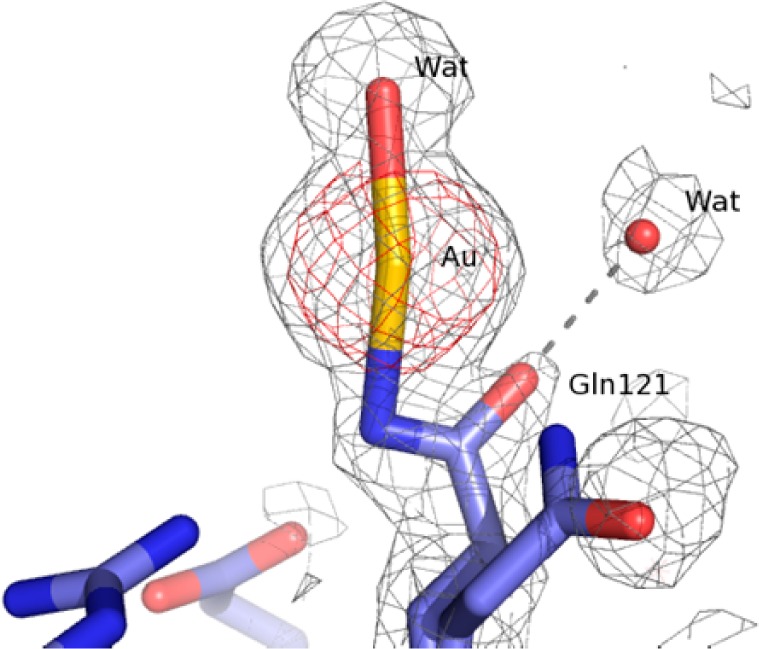
Structural details of the gold center in HEWL–Aubipyc. 2Fo – Fc electron density map is contoured at 2σ (red) and 0.5σ (gray). Gln121 adopts two distinct conformations.
At variance with our previous studies,6 here the gold atom is bound to Gln121 side chain. The coordination geometry of the metal is completed by a water molecule. The geometry is nearly linear, strongly indicating that the gold atom is in the oxidation state +1 (Figures 3 and 4).
Since gold(I) has a preference for soft ligands such as sulfur and phosphorus,4 secondary Au sites were searched close to Cys or Met residues. Yet, there is no evidence of gold atoms bound to these residues. Similarly, no evidence is obtained for the formation of colloidal gold.
Reduction of gold(III) to gold(I) appears to be a common feature in the behavior of medicinal gold(III) complexes, as already observed in many cases,6,7 although not a general rule. Indeed, in some cases, gold(III) complexes, particularly of polydentate ligands, have been reported to maintain their oxidation state, provided the availability in the protein of coordination environments that favor a higher oxidation state.24 Aubipyc was shown to manifest a reasonable stability of its gold(III) chromophore in aqueous solutions (at physiological pH)16 and to display a conspicuous resistance toward reduction.25 It was also found that Aubipyc retains its cyclometalated ligand upon interaction with HEWL, at least in ammonium acetate solutions.26 In line with these previous observations, we propose that Aubipyc could react with the Gln121 amidic side chain to give the gold(III) adduct [Au(bipydmb-H){HN(CO)R}]+ (R = protein). This hypothesis is strongly supported by the recent isolation of the acetamido complex [Au(bipydmb-H){NH(CO)CH3}](PF6), obtained from reaction of Aubipyc with CH3(CO)NH2.27 Upon binding to Gln121, the gold ion may then be reduced by ethylene glycol, thus allowing the detaching of the terdentate ligand. In fact, it is known that this latter solvent is capable of providing reducing species, without the need for a separate reducing agent.28
X-ray absorption near edge structure (XANES) and extended X-ray absorption fine structure (EXAFS) studies carried out on AubipyOH ([Au(2,2′-bipyridine)(OH)2](PF6)), in complex with bovine serum albumin,29,30 showed that the protein-bound gold center manifests a lower coordination number than in AubipyOH alone, strongly suggesting that gold(III) to gold(I) reduction has occurred upon protein interaction.29 Data are also in agreement with the fact that gold(I) centers are anchored to the protein mainly through nitrogen donors.30 Aubipyc reduction and gold(I) binding to proteins is consistent with the observation that this molecule is able to inhibit mitochondrial thioredoxin reductase, producing apoptotic cell death.31
In a more general framework, these data reveal that the formed HEWL–gold adducts are distinct when different gold-based bioactive agents react with the same protein (compare data reported in ref (6) and the present work), although the metallodrug–protein reactions invariantly lead to formation of Au(I) ions.
The higher redox stability of Aubipyc, imparted by the cyclometalated ligand, with respect to Auoxo6 and Au2phen, may account for the different coordination site.
In this respect, it is worth reminding that the interactions of ruthenium complexes with proteins critically depend on the studied model protein (lysozyme or cyt c): significant changes in reactivity were indeed highlighted for different ruthenium compounds and for different proteins.14,20,32 The same may hold for gold complexes.
The understanding of the factors that drive formation of protein adducts with gold compounds is important for the biopharmaceutical development of this class of metal-based agents. To this end, we have been investigating the reactions of a variety of gold compounds with the model protein hen egg white lysozyme to examine the effects of the carrier ligand on the formation of gold–protein complexes.
Upon analyzing the structure of the HEWL–gold adduct arising from Aubipyc and comparing this with those previously solved,6,7 we may conclude the following:
-
(a)
In spite of its higher redox stability, Aubipyc undergoes reduction of the gold(III) center after reaction with HEWL, losing its original ligands. These observations indicate that, similar to other cytotoxic gold(III) agents, Aubipyc likely behaves as a redox activated species.
-
(b)
The resulting gold(I) ions are bound to Gln121. Remarkably, although lysozyme has been very often used to study the interaction between metal-based agents and proteins,6,8−15 the binding site close to Gln121 has never been reported before for a metal compound.
-
(c)
In the conditions used to grow lysozyme–Aubipyc crystals, the reaction between Aubipyc and HEWL produces an adduct that is distinct from that produced in the reaction between the same protein and other cytotoxic Au(III) and Au(I) compounds.6 This result suggests that the nature of the final protein–gold adduct is strongly dependent on the nature of the starting gold compound and that the carrying ligand plays a major role in determining the type of protein–gold adduct.
As a final comment, we note that cytotoxic gold(III) compounds generally react with the model protein lysozyme upon metal center reduction. Although the in vivo relevance of our data is necessarily limited by the inevitable use of the chosen crystallization conditions (i.e., the chemicals and the pH), these findings suggest that the likely activation mechanism of these prodrugs is based on metal reduction. Gold(III) compounds may undergo redox reactions with proteins rich in sulfur containing residues, but they also interact with His or Gln. Further work on protein–gold(III) compound interactions will help to improve our understanding of the chemistry and reactivity of these candidate drugs.
Acknowledgments
The authors thank Giosuè Sorrentino and Maurizio Amendola for technical assistance.
Glossary
ABBREVIATIONS
- HEWL
hen egg white lysozyme
- Aubipyc
[(bipydmb-H)Au(OH)][PF6]
Supporting Information Available
Data collection and processing; structure solution and refinement. This material is available free of charge via the Internet at http://pubs.acs.org.
Author Contributions
The manuscript was written through contributions of all authors.
This work was supported by Beneficentia Stiftung (Vaduz, Liechtenstein), AIRC (IG-12085), and COST Action CM1105.
The authors declare no competing financial interest.
Supplementary Material
References
- Bertrand B.; Casini A. A golden future in medicinal inorganic chemistry: The promise of anticancer gold organometallic compounds. Dalton Trans. 2014, 43, 4209–4219. [DOI] [PubMed] [Google Scholar]
- Urig S.; Fritz-Wolf K.; Réau R.; Herold-Mende C.; Tóth K.; Davioud-Charvet E.; Becker K. Undressing of phosphine gold(I) complexes as irreversible inhibitors of human disulfide reductases. Angew. Chem., Int. Ed. 2006, 45, 1881–6. [DOI] [PubMed] [Google Scholar]
- Zou J.; Taylor P.; Dornan J.; Robinson S. P.; Walkinshaw M. D.; Sadler P. J. First crystal structure of a medicinally relevant gold protein complex: unexpected binding of [Au(PEt3)]+ to histidine. Angew. Chem., Int. Ed. 2000, 39, 2931–2934. [DOI] [PubMed] [Google Scholar]
- Bhabak K. P.; Bhuyan B. J.; Mugesh G. Bioinorganic and medicinal chemistry: aspects of gold(I)-protein complexes. Dalton Trans. 2011, 40, 2099–111. [DOI] [PubMed] [Google Scholar]
- Bertrand B.; Stefan L.; Pirrotta M.; Monchaud D.; Bodio E.; Richard P.; Le Gendre P.; Warmerdam E.; de Jager M. H.; Groothuis G. M.; Picquet M.; Casini A. Caffeine-based gold(I) N-heterocyclic carbenes as possible anticancer agents: Synthesis and biological properties. Inorg. Chem. 2014, 53, 2296–2303. [DOI] [PubMed] [Google Scholar]
- Messori L.; Scaletti F.; Massai L.; Cinellu M. A.; Gabbiani C.; Vergara A.; Merlino A. The mode of action of anticancer gold-based drugs: a structural perspective. Chem. Commun. 2013, 49, 10100–10102. [DOI] [PubMed] [Google Scholar]
- Messori L.; Scaletti F.; Massai L.; Cinellu M. A.; Russo Krauss I.; di Martino G.; Vergara A.; Paduano L.; Merlino A. Interactions of gold-based drugs with proteins: crystal structure of the adduct formed between ribonuclease A and a cytotoxic gold(III) compound. Metallomics 2014, 6, 233–236. [DOI] [PubMed] [Google Scholar]
- Santos-Silva T.; Mukhopadhyay A.; Seixas J. D.; Bernardes G. J.; Romao C. C.; Romao M. J. CORM-3 reactivity toward proteins: the crystal structure of a Ru(II) dicarbonyl-lysozyme complex. J. Am. Chem. Soc. 2011, 133, 1192–5. [DOI] [PubMed] [Google Scholar]
- Helliwell J. R.; Tanley S. W. The crystal structure analysis of the relative binding of cisplatin and carboplatin in a mixture with histidine in a protein studied at 100 and 300 K with repeated X-ray irradiation. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2013, 69, 121–5. [DOI] [PubMed] [Google Scholar]
- Messori L.; Marzo T.; Gabbiani C.; Valdes A. A.; Quiroga A. G.; Merlino A. Peculiar features in the crystal structure of the adduct formed between cis-PtI2(NH3)2 and hen egg white lysozyme. Inorg. Chem. 2013, 52, 13827–13829. [DOI] [PubMed] [Google Scholar]
- Messori L.; Marzo T.; Fernandes Sanches R. N.; Rehman H.-U.; de Oliveira Silva D.; Merlino A. Unusual structural features in the lysozyme derivative of the tetrakis(acetato)chloridodiruthenium(II,III) complex. Angew. Chem., Int. Ed. 2014, 53, 6172–5. [DOI] [PubMed] [Google Scholar]
- Tanley S. W. M.; Schreurs A. M. M.; Helliwell J. R.; Kroon-Batenburg L. M. J. Experience with exchange and archiving of raw data: comparison of data from two diffractometers and four software packages on a series of lysozyme crystals. J. Appl. Crystallogr. 2013, 46, 108–119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanley S. W. M.; Diederichs K.; Kroon-Batenburg L. M. J.; Schreurs A. M. M.; Helliwell J. R. Experiences with archived raw diffraction images data: capturing cisplatin after chemical conversion of carboplatin in high salt conditions for a protein crystal. J. Synchrotron Radiat. 2013, 20, 880–883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messori L.; Merlino A. Ruthenium metalation of proteins: the X-ray structure of the complex formed between NAMI-A and hen egg white lysozyme. Dalton Trans. 2014, 43, 6128–31. [DOI] [PubMed] [Google Scholar]
- Messori L.; Merlino A. Cisplatin binding to proteins: molecular structure of the ribonuclease A adduct. Inorg. Chem. 2014, 53, 3929–31. [DOI] [PubMed] [Google Scholar]
- Marcon G.; Carotti S.; Coronnello M.; Messori L.; Mini E.; Orioli P.; Mazzei T.; Cinellu M. A.; Minghetti G. Gold(III) complexes with bipyridyl ligands: solution chemistry, cytotoxicity, and DNA binding properties. J. Med. Chem. 2002, 45, 1672–7. [DOI] [PubMed] [Google Scholar]
- Pellegrini E.; Piano D.; Bowler M. W. Direct cryocooling of naked crystals: are cryoprotection agents always necessary?. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2011, 67, 902–906. [DOI] [PubMed] [Google Scholar]
- Russo Krauss I.; Sica F.; Mattia C. A.; Merlino A. Increasing the X-ray diffraction power of protein crystals by dehydration: the case of bovine serum albumin and a survey of literature data. Int. J. Mol. Sci. 2012, 13, 3782–800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otwinowski Z.; Minor W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [DOI] [PubMed] [Google Scholar]
- Vergara A.; D’Errico G.; Montesarchio D.; Mangiapia G.; Paduano L.; Merlino A. Interaction of anticancer ruthenium compounds with proteins: high-resolution X-ray structures and Raman microscopy studies of the adduct between hen egg white lysozyme and AziRu. Inorg. Chem. 2013, 52, 4157–4159. [DOI] [PubMed] [Google Scholar]
- Murshudov G. N.; Skuba K. P.; Lebedev A. A.; Pannu N. S.; Steiner R. A.; Nicholls R. A.; Winn M. D.; Long F.; Vagin A. A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2011, 67, 355–367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emsley P.; Cowtan K. Coot: model-building tools for molecular graphics. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2004, 60, 2126–2132. [DOI] [PubMed] [Google Scholar]
- Wang J.; Dauter S. M.; Alkire R.; Joachimiak A.; Dauter Z. Triclinic lysozyme at 0.65 Å resolution. Acta Crystallogr., Sect. D: Biol. Crystallogr. 2007, 63, 1254–68. [DOI] [PubMed] [Google Scholar]
- Mendes F.; Groessl M.; Nazarov A. A.; Tsybin Y. O.; Sava G.; Santos I.; Dyson P. J.; Casini A. Metal-based inhibition of poly(ADP-ribose) polymerase: the guardina angel of DNA. J. Med. Chem. 2011, 54, 2196–2206. [DOI] [PubMed] [Google Scholar]
- Sanna G.; Pilo M. I.; Spano N.; Minghetti G.; Cinellu M. A.; Zucca A.; Seeber R. Electrochemical behaviour of cyclometallated gold(III) complexes. Evidences of transcyclometallation in the fate of electroreduced species. J. Organomet. Chem. 2001, 622, 47–53. [Google Scholar]
- Gabbiani C.; Massai L.; Scaletti F.; Michelucci E.; Maiore L.; Cinellu M. A.; Messori L. Protein metalation by metal-based drugs: reactions of cytotoxic gold compounds with cytochrome c and lysozyme. J. Biol. Inorg. Chem. 2012, 17, 1293–1302. [DOI] [PubMed] [Google Scholar]
- Results to be published. For N–H bond activation of aryl- and alkylamines by Aubipyc; seeCinellu M. A.; Minghetti G.; Pinna M. V.; Stoccoro S.; Zucca A. Synthesis and characterization of mononuclear amidogold(III) complexes. Crystal structure of [Au(N2C10H7(CMe2C6H4)-6](NHC6H3Me2-2,6)][PF6]. Oxidation of 4-methyl-aniline to azotoluene. Eur. J. Inorg. Chem. 2003, 2304–2310. [Google Scholar]
- Eustis S.; Hsu H. Y.; El-Sayed M. A. Gold nanoparticle formation from photochemical reduction of Au3+ by continuous excitation in colloidal solutions. A proposed molecular mechanism. J. Phys. Chem. B 2005, 109, 4811–4815. [DOI] [PubMed] [Google Scholar]
- Soldatov M. A.; Ascone I.; Congiu-Castellano A.; Messori L.; Cinellu M. A.; Balerna A.; Soldatov A. V.; Yalovega G. E. Potential antitumor gold drugs: DFT and XANES studies of local atomic and electronic structure. J. Phys.: Conf. Ser. 2009, 190, 1–4. [Google Scholar]
- Messori L.; Balerna A.; Ascone I.; Castellano C.; Gabbiani C.; Casini A.; Marchioni C.; Jaouen G.; Castellano A. C. X-ray absorption spectroscopy studies of the adducts formed between cytotoxic gold compounds and two major serum proteins. J. Biol. Inorg. Chem. 2011, 16, 491–499. [DOI] [PubMed] [Google Scholar]
- Rigobello M. P.; Messori L.; Marcon G.; Cinellu M. A.; Bragadin M.; Folda A.; Scutari G.; Bindoli A. Gold complexes inhibit mitochondrial thioredoxin reductase: consequences on mitochondrial functions. J. Inorg. Biochem 2004, 98, 1634–41. [DOI] [PubMed] [Google Scholar]
- Casini A.; Mastrobuoni G.; Terenghi M.; Gabbiani C.; Monzani E.; Moneti G.; Casella L.; Messori L. Ruthenium anticancer drugs and proteins: a study of the interactions of the ruthenium(III) complex imidazolium trans-[tetrachloro(dimethyl sulfoxide)(imidazole)ruthenate(III)] with hen egg white lysozyme and horse heart cytochrome c. J. Biol. Inorg. Chem. 2007, 12, 1107–17. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.