

Abstract
A series of multifunctional directed 3-arylcoumarin-tetracyclic tacrine derivatives was designed and synthesized for the treatment of Parkinson’s disease (PD). A number of derivatives (18, 19, 20, 21, and 24) demonstrated significant reduction of aggregation of “human” alpha-synuclein (α-synuclein) protein, expressing on transgenic Caenorhabditis elegans (C. elegans) model NL5901. Moreover, compounds 16, 18, and 24 also exhibited good antioxidant properties and significantly increased the dopamine (DA) content in N2 and NL5901 strains of C. elegans. Interestingly, the protective efficacy of these hybrids seems to be mediated via activation of longevity promoting transcription factor DAF-16. In addition, molecular modeling studies have evidenced the exquisite interaction of most active compounds 18 and 24 with α-synuclein protein. Taken together, the data indicate that the derivatives may be useful leads against aging and age associated PD.
Keywords: 3-Arylcoumarin, tetracyclic tacrine, Parkinson’s disease, C. elegans, α-synuclein, reactive oxygen species
Parkinson’s disease (PD) is an irreversible neurodegenerative brain disorder, due to diminution of dopaminergic neurons in the region of substantia nigra.1 The main pathological hallmark of PD is the presence and accumulation of a protein known as alpha-synuclein (α-synuclein).2 The accumulation induces dopaminergic nerve terminal degeneration, which eventually leads to loss of dopamine-producing cells in the mid-brain.3 In a disease condition, the gradual loss of dopamine levels leads to motor and nonmotor dysfunctions.4l-Dopa (levodopa) is the most potent medication used for over 30 years, which is the precursor of dopamine.5 It effectively relieves the dopaminergic deficit; however, the disease persists and further uncontrolled symptoms leads to several complications like disturbing dyskinesias or unconditional movements.6
Presently, there is no cure for PD and the drugs used for treatment are mainly dopamine agonists (pramipexole and ropinirole) and monoamine oxidase inhibitors (rasagiline and selegiline) (Figure 1A), which provide only symptomatic relief.7 Therefore, novel treatment and prevention approaches are urgently needed for PD therapy.
Figure 1.

(A) Chemical structure of some potent anti-Parkinson’s drugs. (B) Chemical structure of acetylcholinesterase inhibitors and some potent neuroprotective tetracyclic tacrine derivatives.
Because of the complex etiology found in PD, scientists are now turning to the design of multitarget-directed ligands (MTDLs) that should be able to interact with the multiple targets.8 In the design of MTDLs, molecular hybridization strategy could be utilized, wherein two compounds binding to their targets are used as the starting points and their structural features are fused to incorporate activity at both targets into a single hybrid molecule.9
Considerable interest has been focused on the coumarin structure, which has been known to possess a broad spectrum of biological activities.10 Recent evidence has suggested that 3-aryl coumarin nucleus constitutes a promising scaffold for inhibition of acetylcholinesterase (AChE) and AChE-induced β-amyloid aggregation.11 Recently, Abdelhafez et al. reported that some of the 3-substituted coumarin derivatives potentially act as a dopamine releasing agent.12 Other relevant reports have also suggested that these derivatives protect the neurons from the oxidative disorders and effectively enhance the dopamine biosynthesis.13,14 However, the AChE inhibitors like donepezil and tacrine derivatives (Figure 1B) have shown beneficial effects in the treatment of PD.15 Marco-Contelles et al. reported the neuroprotective activity of tetracyclic tacrine derivatives.16 Some of recent reports have also underscored the importance of tacrine containing derivatives that could exhibit neuroprotective effects in an neurodegenerative condition.17 The increase of the dopamine levels and decrease of ROS and neuroprotective properties are crucial to overcome PD. The concept of MTDL approach prompted us to synthesize a novel class 3-aryl coumarin and tetracyclic-tacrine hybrids that inhibit α-synuclein aggregation and increase availability of dopamine levels, thereby halting the progression of the disease.
The synthesis of target and intermediate compounds is represented in Scheme 1. The Duff formylation on different ortho-substituted phenols (1 and 2) in the presence of hexamethylenetetraamine (HMTA) and trifluoroacetic acid (TFA) at 120 °C gave aromatic dicarbaldehyde intermediates (3 and 4).18 These dicarbaldehydes were then engaged with different substituted phenylacetic acids in the presence of cyanuric chloride and N-methyl morpholine (NMM) in DMF for 1 h, resulting in a good yield of respective 3-aryl coumarin aldehydes (5–9).19 Further, these substrates (5–9) underwent the multicomponent reaction with malononitrile and different 1,3-cyclohexadiones in the presence of dimethylaminopyridine (DMAP) as a catalyst afforded coumarin-4H-chromene intermediates (10–15).20 Finally, the Friedlander reaction between these intermediates (10–15)21 and different cyclic ketones (cyclopentanone and cyclohexanone) in the presence of aluminum trichloride (AlCl3) as a catalyst furnished the target 3-arylcoumarin-tetracylic tacrine derivatives (16–24) in good yields.
Scheme 1. Synthesis of 3-Arylcoumarin-tetracyclic Tacrine Derivatives.

Reagents and conditions: (i) HMTA, TFA, 120 °C, 4 h. (ii) aq. H2SO4, 100 °C, 2 h. (iii) Appropriate phenylacetic acid, cyanuric chloride, NMM, DMF, 110 °C, 30–90 min. (iv) Malononitrile, different 1,3-cyclohexadiones, DMAP, EtOH, reflux, 0.5 h. (v) Different cyclohexanones, AlCl3, dichloroethane, reflux, 6 h.
In the present study, we utilized the transgenic C. elegans model to evaluate the effect of these compounds (racemic mixtures) on PD. The transparent anatomy of the nematode readily helps in monitoring the aggregation of the neural protein α-synuclein.22 Interestingly, 60–80% of C. elegans genes are homologous to humans.23
We employed transgenic C. elegans model NL5901 expressing human α-synuclein protein tagged with yellow fluorescent protein (NL5901 (Punc-54::α- synuclein::YFP+unc-119)) in the body wall muscle.24 We explored the role of different 3-arylcoumarin-tetracyclic tacrine derivatives on α-synuclein aggregation and compared it with the known positive control Bacopa monneiri (BM).25 Worms treated with different derivatives were analyzed using fluorescent microscopy. We further quantified the images for fluorescence intensity of α-synuclein aggregation using ImageJ software. Out of nine compounds tested, five compounds exhibited significant reduction of aggregation of α-synuclein protein along with the positive control BM. In the case of worms treated with test compounds, 18, 19, 20, 21, and 24 showed mean fluorescence intensity 2.94 ± 0.24, 3.82 ± 0.25, 4.42 ± 0.15, 5.73 ± 0.40, and 3.76 ± 0.23 arbitrary units, respectively, thereby showing potential reduction by 2.6 (p < 0.001), 2.0 (p < 0.01), 1.76 (p < 0.001), 1.35(p < 0.05), and 2.1 (p < 0.001) folds, when compared to untreated worms, whereas BM exhibited 1.25-fold reduction of α-synuclein aggregation (p < 0.001) (Figure 2). The mean fluorescence intensity for control group was 7.76 ± 0.03 arbitrary units. However, worms treated with compounds 16, 17, and 23 exhibited no effect on α-synuclein aggregation and mean fluorescence intensity 8.63 ± 0.10 arbitrary units. From the results, it is clear that compounds 18, 19, 20, 21, and 24 reduced the α-synuclein protein aggregation, suggesting their therapeutic potential against PD.
Figure 2.
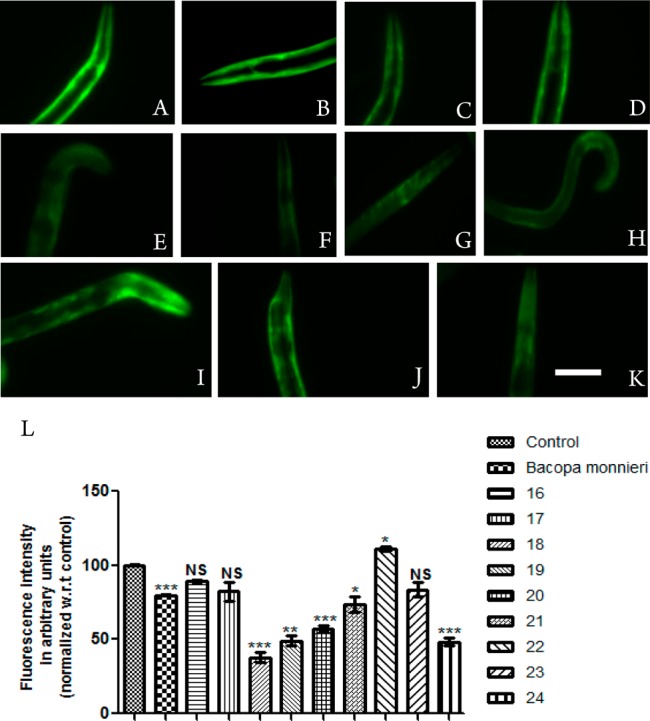
α-Synuclein aggregation in NL5901 strain of C. elegans treated with OP50 (A), Bacopa monnieri (B), 16 (C), 17 (D), 18 (E), 19 (F), 20 (G), 21(H), 22 (I), 23 (J), and 24 (K). Scale bar, 50 μm. (L) Graphical representation for fluorescence intensity of aggregation of the nematodes as quantified using ImageJ software. *p < 0.05, **p < 0.01, ***p < 0.001, and NS, not significant.
Pathogenesis of PD is intimately associated with oxidative stress, which is due to ROS generated by dopamine metabolism, mitochondrial dysfunction, and neuroinflammation.26 Since the reduction of oxidative stress is another crucial aspect in designing agents for PD treatment, we examined antioxidative properties of our hybrids by using a nonfluorescent dye dichlorodihydrofluorescein diacetate (H2DCFDA) assay, in the wild-type N2 strain of C. elegans.27 Interestingly, compounds 16, 18, and 24 showed significant reduction in oxidative stress to that of untreated worms, suggesting that the hybrids have the potential to be potent multifunctional agents (Supporting Information Figure S1).
Since 3-arylcoumarin is a promising scaffold to inhibit AChE and AChE-induced β-amyloid aggregation,11 we further examined the AChE inhibition property of these hybrids. In this study, we have employed an indirect method by the quantification of ACh level using amplex red assay kit. Worms treated with compounds 16, 17, 18, 19, 20, 22, 23, and 24 except 21 exhibited a significant increase in the ACh level, indicating that the test compounds were significantly inhibiting AChE via increasing ACh level (Supporting Information Figure S2).
Among the various test compounds, 18 and 24 displayed the best effects in both reducing the α-synuclein aggregate and exhibiting antioxidant properties; therefore, 18 and 24 were taken further to analyze their effects on dopamine signaling. For the quantification of dopamine (DA) content, nonanol assay was employed, which gives us indirect readings of the DA contents as any deviation from the normal DA levels affects worm response time to volatile repellent nonanol.28 Worms with higher DA content responds early by moving back, and the worm with decreased DA content takes longer time to respond toward nonanol. We quantified the effects of compounds 18 and 24 on the DA content along with BM in N2 and NL5901 strains of C. elegans.29 After treatment of the worms with compounds BM, 18, and 24, the response time was significantly decreased by 1.63 (p < 0.05), 2.6 (p < 0.01), and 1.8 (p < 0.05) folds in N2 and 3.25 (p < 0.01), 1.75 (p < 0.05), and 1.9 (p < 0.05) folds in NL5901, respectively, as compared to untreated worms indicating that there was increase in DA content after treatment (Figure 3a). Thus, the DA content was improved considerably after exposure with 18 and 24 compounds in both the strains N2 and NL5901 of C. elegans.
Figure 3.
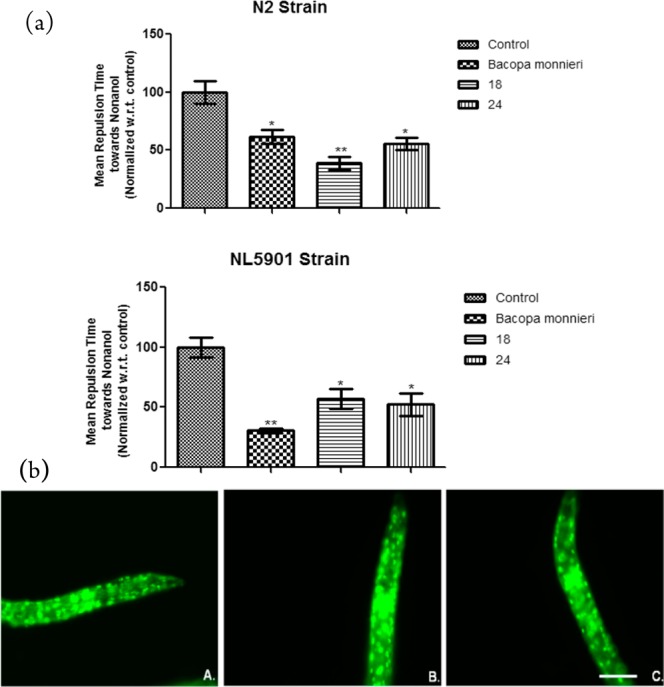
(a) Effect of active compounds on dopamine content as estimated by nonanol repulsion assay in N2 and NL5901 strain of C. elegans. *p < 0.05, **p < 0.01, NS, not significant. (b) DAF-16 expression in TJ356 strain of C. elegans fed with OP50 (A) and treated with compounds 18 (B) and 24 (C). Scale bar, 50 μm.
Since DAF-16 is considered as a major transcription factor required for the fundamental lifespan extension and longevity, its translocation into nucleus was examined by using a transgenic GFP expressing strain TJ356 of C. elegans.30 An increase in GFP intensity was observed in the case of worms treated with compound 18 when compared to the worms raised on normal E. coli-OP50 diet. Figure 3b depicts the representative images for control (Figure 3b(A)), compound 18 treated worms (Figure 3b(B)), and compound 24 treated worms (Figure 3b(C)). However, no significant effect was observed in compound 24 treated worms as compared to the control. The present experiment suggested that the neuroprotective effects of compound 18 might be associated with DAF-16 signaling pathway.
Next, since tacrine derivatives are known to exhibit hepatotoxicity in liver, we examined the most active compounds for their cytotoxicities in Hep-G2 and HEK-293 cell lines. Interestingly, both the active compounds 18 and 24 possessed higher safety profile than tacrine (Supporting Information Figures S3 and S4).
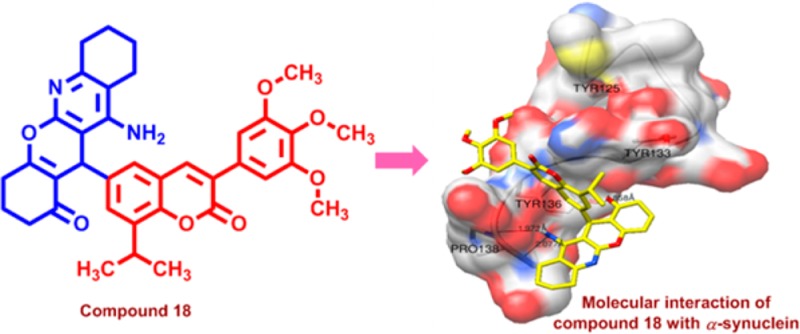
The preceding experimental evidence on α-synuclein aggregation of these derivatives motivated us to probe the molecular interactions with α-synuclein protein for the most active compounds 18 and 24. In the literature, docking studies of α-synuclein were successfully reported using Autodock 4.2 (Supporting Information Figure S5).31 Autodock studies revealed that S-conformation of compound 18 (18-S) formed large clusters of 25 poses having least inhibitory constant (Ki) of 207.96 nM, binding energy of −9.12 kcal/mol, and displaying strong hydrogen bond by 1.858 Å with Tyr 136 and 1.973 and 2.077 Å hydrogen bonds with Pro138, indicating interaction of ligand with tyrosine triad (Supporting Information Figure S6a). Thus, it can be used as an inhibitor for preventing α-synuclein aggregation and intracellular accumulation of misfolded protein. In the case of S-conformation of compound 24 (24-S), a cluster of 13 poses was formed having least Ki of 263 nM and binding energy of −8.98 kcal/mol. Compound 24-S, in this lowest energy pose, shows a strong hydrogen bond by 1.806 Å with Tyr 136 and two hydrogen bonds of the order 1.845 and 2.078 Å with Pro138 (Supporting Information Figure S6b). Slight increase in Ki value of compound 24-S compared to compound 18-S can be attributed to its distorted trimethoxybenzene ring due to the presence of an extra methyl group, which is near to hydrophobic Tyr 133 (Figure 4A). Less number of poses in the least energy cluster size may possibly account for a slight decrease in activity of compound 24-S compared to that of active compound 18-S. Docking studies of the R-conformation of compounds 18 (18-R) and 24-R reveal a potential binding site with lowest Ki value of 1.57 and 1.97 μM, respectively, both having hydrogen bond with Glu126 residue of the order 3.005 and 3.095 Å, respectively (Figure 4B and Supporting Information Figure S6c,d). In this study, the number of conformations in a simple cluster of compounds 18-R and 24-R were 45 and 28, respectively. Additionally, the molecular dynamics studies performed by Gromacs 4.532 indicated a stable binding of both isomers of 18 and 24 with root-mean-square deviation (rmsd) less than 0.3 nm over a time period of 2 ns (Figure 4C).33
Figure 4.
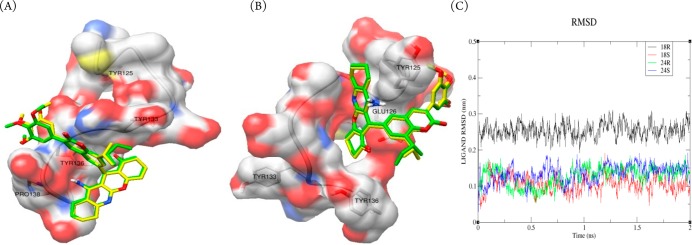
(A) Molecular interaction of compound 18-S (yellow) and compound 24-S (green) with α-synuclein. (B) Compound 18-R (yellow) and compound 24-R (green) with α-synuclein. (C) Temporal molecular dynamics studies of compounds 18-R, 18-S, 24-R, and 24-S for 2 ns.
In terms of preliminary structure–activity relationships of 3-arylcoumarin-tetracylic tacrine hybrids, compounds containing trimethoxy groups (18, 24, 20, and 21) and alkyl substitution (sec-butyl and isopropyl group) at C8 position of 3-aryl coumarin ring are optimal for activity. However, on the tetracyclic tacrine core the presence of unsubstituted cyclohexanone and cyclohexane is preferred over substituted cyclohexanone and cyclopentane, respectively (16 and 17), in inhibiting the α-synuclein aggregation.
In conclusion, a series of novel multifunctional 3-arylcoumarin-tetracyclic tacrine derivatives have been synthesized and evaluated for PD therapy. Compounds 18 and 24 were identified as molecules of interest as they potentially inhibited the aggregation of α-synuclein protein and exhibited promising antioxidant properties. Interestingly, they also enhanced the dopamine content, which is crucial and beneficial for transporting signals to the nerve cells, in the condition of PD. Mechanistically, the protective efficacy of these derivatives seems to be mediated via signaling the expression of DAF-16 protein. Molecular modeling studies further corroborated and confirmed that compounds 18 and 24 strongly bind with the α-synuclein protein and efficiently inhibit the aggregation. Results presented herein not only reveal the structural and mechanistic insight of these novel hybrids but also provide impetus to synthesize enantiomerically pure molecules, which is ongoing in our lab.
Acknowledgments
Instrumentation facilities from SAIF and CDRI are gratefully acknowledged.
Supporting Information Available
Experimental procedures, characterization of new compounds, and modeling and assay protocols. This material is available free of charge via the Internet at http://pubs.acs.org.
R.K.M. and P.J. are thankful to CSIR, New Delhi, India for financial support. Nematode strains used in this work were provided by the C. elegans Genetics Center (CGC), University of Minnesota, MN, USA, which is funded by the NIH National Center for Research Resources (NCRR). This is CSIR-CDRI communication number 8763.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- Obeso J. A.; Rodriguez-Oroz M. C.; Goetz C. G.; Marin C.; Kordower J. H.; Rodriguez M.; Hirsch E. C.; Farrer M.; Schapira A. H.; Halliday G. Missing pieces in the Parkinson’s disease puzzle. Nat. Med. 2010, 16, 653–661. [DOI] [PubMed] [Google Scholar]
- Spillantini M. G.; Schmidt M. L.; Lee V. M.; Trojanowski J. Q.; Jakes R.; Goedert M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [DOI] [PubMed] [Google Scholar]
- Lee F. J.; Liu F.; Pristupa Z. B.; Niznik H. B. Direct binding and functional coupling of alpha-synuclein to the dopamine transporters accelerate dopamine-induced apoptosis. FASEB J. 2001, 15, 916–926. [DOI] [PubMed] [Google Scholar]
- Olanow C. W.; Obeso J. A.; Stocchi F. Continuous dopamine-receptor treatment of Parkinson’s disease: scientific rationale and clinical implications. Lancet Neurol. 2006, 5, 677–687. [DOI] [PubMed] [Google Scholar]
- Lewitt P. A. Levodopa for the treatment of Parkinson’s disease. N. Engl. J. Med. 2008, 359, 2468–2476. [DOI] [PubMed] [Google Scholar]
- Abbott A. Levodopa: the story so far. Nature 2010, 466, S6–S7. [DOI] [PubMed] [Google Scholar]
- Meissner W. G.; Frasier M.; Gasser T.; Goetz C. G.; Lozano A.; Piccini P.; Obeso J. A.; Rascol O.; Schapira A.; Voon V.; Weiner D. M.; Tison F.; Bezard E. Priorities in Parkinson’s disease research. Nat. Rev. Drug Discovery 2011, 10, 377–393. [DOI] [PubMed] [Google Scholar]
- Morphy R.; Rankovic Z. Designed multiple ligands. An emerging drug discovery paradigm. J. Med. Chem. 2005, 48, 6523–6543. [DOI] [PubMed] [Google Scholar]
- Cavalli A.; Bolognesi M. L.; Minarini A.; Rosini M.; Tumiatti V.; Recanatini M.; Melchiorre C. Multi-target-directed ligands to combat neurodegenerative diseases. J. Med. Chem. 2008, 51, 347–372. [DOI] [PubMed] [Google Scholar]
- Venugopala K. N.; Rashmi V.; Odhav B. Review on natural coumarin lead compounds for their pharmacological activity. Biomed. Res. Int. 2013, 2013, 963248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piazzi L.; Rampa A.; Bisi A.; Gobbi S.; Belluti F.; Cavalli A.; Bartolini M.; Andrisano V.; Valenti P.; Recanatini M. 3-(4-[[Benzyl(methyl)amino]methyl]phenyl)-6,7-dimethoxy-2H-2-chr omenone (AP2238) inhibits both acetylcholinesterase and acetylcholinesterase-induced beta-amyloid aggregation: a dual function lead for Alzheimer’s disease therapy. J. Med. Chem. 2003, 46, 2279–2282. [DOI] [PubMed] [Google Scholar]
- Abdelhafez O. M.; Amin K. M.; Ali H. I.; Maher T. J.; Batran R. Z. Dopamine release and molecular modeling study of some coumarin derivatives. Neurochem. Int. 2011, 59, 906–912. [DOI] [PubMed] [Google Scholar]
- Matos M. J.; Pérez-Cruz F.; Vazquez-Rodriguez S.; Uriarte E.; Santana L.; Borges F.; Olea-Azar C. Remarkable antioxidant properties of a series of hydroxy-3-arylcoumarins. Bioorg. Med. Chem. 2013, 21, 3900–3906. [DOI] [PubMed] [Google Scholar]
- Yang Y. J.; Lee H. J.; Huang H. S.; Lee B. K.; Choi H. S.; Lim S. C.; Lee C. K.; Lee M. K. Effects of scoparone on dopamine biosynthesis and L-DOPA-induced cytotoxicity in PC12 cells. J. Neurosci. Res. 2009, 87, 1929–1937. [DOI] [PubMed] [Google Scholar]
- Van Laar T.; De Deyn P. P.; Aarsland D.; Barone P.; Galvin J. E. Effects of cholinesterase inhibitors in Parkinson’s disease dementia: a review of clinical data. CNS Neurosci. Ther. 2011, 17, 428–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marco-Contelles J.; León R.; de los Ríos C.; García A. G.; López M. G.; Villarroya M. New multipotent tetracyclic tacrines with neuroprotective activity. Bioorg. Med. Chem. 2006, 14, 8176–8185. [DOI] [PubMed] [Google Scholar]
- León R.; de los Ríos C.; Marco-Contelles J.; Huertas O.; Barril X.; Luque F. J.; López M. G.; García A. G.; Villarroya M. New tacrine-dihydropyridine hybrids that inhibit acetylcholinesterase, calcium entry, and exhibit neuroprotection properties. Bioorg. Med. Chem. 2006, 16, 7759–7769. [DOI] [PubMed] [Google Scholar]
- Sashidhara K. V.; Kumar M.; Khedgikar V.; Kushwaha P.; Modukuri R. K.; Kumar A.; Gautam J.; Singh D.; Sridhar B.; Trivedi R. Discovery of coumarin-dihydropyridine hybrids as bone anabolic agents. J. Med. Chem. 2013, 56, 109–122. [DOI] [PubMed] [Google Scholar]
- Sashidhara K. V.; Palnati G. R.; Avula S. R.; Kumar A. Efficient and general synthesis of 3-aryl coumarins using cyanuric chloride. Synlett 2012, 23, 611–621. [Google Scholar]
- Khan A. T.; Lal M.; Ali S.; Khan M. M. One-pot three-component reaction for the synthesis of pyran annulated heterocyclic compounds using DMAP as a catalyst. Tetrahedron Lett. 2011, 52, 5327–5332. [Google Scholar]
- Marco-Contelles J.; Pérez-Mayoral E.; Samadi A.; Carreiras M. C.; Soriano E. Recent advances in the Friedländer reaction. Chem. Rev. 2009, 109, 2652–2671. [DOI] [PubMed] [Google Scholar]
- Bodhicharla R.; Nagarajan A.; Winter J.; Adenle A.; Nazir A.; Brady D.; Vere K.; Richens J.; O’Shea P.; Bell D. R.; de Pomerai D. Effects of α-synuclein overexpression in transgenic Caenorhabditis elegans strains. CNS Neurol. Disord. Drug Targets 2012, 11, 965–975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vistbakka J.; VanDuyn N.; Wong G.; Nass R. C. elegans as a genetic model system to identify Parkinson’s disease-associated therapeutic targets. CNS Neurol. Disord. Drug Targets 2012, 11, 957–964. [DOI] [PubMed] [Google Scholar]
- Shukla V.; Phulara S. C.; Yadav D.; Tiwari S.; Kaur S.; Gupta M. M.; Nazir A.; Pandey R. Iridoid compound 10-O-trans-p-coumaroylcatalpol extends longevity and reduces α synuclein aggregation in Caenorhabditis elegans. CNS Neurol. Disord. Drug Targets 2012, 11, 984–992. [DOI] [PubMed] [Google Scholar]
- Jadiya P.; Khan A.; Sammi S. R.; Kaur S.; Mir S. S.; Nazir A. Anti-Parkinsonian effects of Bacopa monnieri: insights from transgenic and pharmacological Caenorhabditis elegans models of Parkinson’s disease. Biochem. Biophys. Res. Commun. 2011, 413, 605–610. [DOI] [PubMed] [Google Scholar]
- Dias V.; Junn E.; Mouradian M. M. The role of oxidative stress in Parkinson’s disease. J. Parkinson’s Dis. 2013, 3, 461–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jadiya P.; Nazir A. Environmental toxicants as extrinsic epigenetic factors for Parkinsonism: Studies employing transgenic C. elegans model. CNS Neurol. Disord. Drug Targets 2012, 11, 976–983. [DOI] [PubMed] [Google Scholar]
- Bargmann C. I.; Hartwieg E.; Horvitz H. R. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 1993, 74, 515–527. [DOI] [PubMed] [Google Scholar]
- Kaur S.; Sammi S. R.; Jadiya P.; Nazir A. RNAi of cat-2, a putative tyrosine hydroxylase, increases alpha synuclein aggregation and associated effects in transgenic C. elegans. CNS Neurol. Disord. Drug Targets 2012, 11, 387–394. [DOI] [PubMed] [Google Scholar]
- Chiang W. C.; Tishkoff D. X.; Yang B.; Wilson-Grady J.; Yu X.; Mazer T.; Eckersdorff M.; Gygi S. P.; Lombard D. B.; Hsu A. L. C. elegans SIRT6/7 homolog SIR-2.4 promotes DAF-16 relocalization and function during stress. PLoS Genet. 2012, 8, e1002948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruzza P.; Siligardi G.; Hussain R.; Marchiani A.; Islami M.; Bubacco L.; Delogu G.; Fabbri D.; Dettori M. A.; Sechi M.; Pala N.; Spissu Y.; Migheli R.; Serra P. A.; Sechi G. Ceftriaxone blocks the polymerization of α-synuclein and exerts neuroprotective effects in vitro. ACS Chem. Neurosci. 2014, 5, 30–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hess B.; Kutzner C.; Van der Spoel D.; Lindahl E. GROMACS 4: algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [DOI] [PubMed] [Google Scholar]
- Huang H. J.; Chen H. Y.; Lee C. C.; Chen C. Y. Computational design of apolipoprotein e4 inhibitors for Alzheimer’s disease therapy from traditional chinese medicine. Biomed. Res. Int. 2014, 2014, 452625. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.