

Abstract
Increased carbon emissions from fossil fuels are increasing the pCO2 of the ocean surface waters in a process called ocean acidification. Elevated water pCO2 can induce physiological and behavioural effects in teleost fishes, although there appear to be large differences in sensitivity between species. There is currently no information available on the possible responses to future ocean acidification in elasmobranch fishes. We exposed small-spotted catsharks (Scyliorhinus canicula) to either control conditions or a year 2100 scenario of 990 μatm pCO2 for four weeks. We did not detect treatment effects on growth, resting metabolic rate, aerobic scope, skin denticle ultrastructure or skin denticle morphology. However, we found that the elevated pCO2 group buffered internal acidosis via 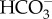 accumulation with an associated increase in Na+, indicating that the blood chemistry remained altered despite the long acclimation period. The elevated pCO2 group also exhibited a shift in their nocturnal swimming pattern from a pattern of many starts and stops to more continuous swimming. Although CO2-exposed teleost fishes can display reduced behavioural asymmetry (lateralization), the CO2-exposed sharks showed increased lateralization. These behavioural effects may suggest that elasmobranch neurophysiology is affected by CO2, as in some teleosts, or that the sharks detect CO2 as a constant stressor, which leads to altered behaviour. The potential direct effects of ocean acidification should henceforth be considered when assessing future anthropogenic effects on sharks.
accumulation with an associated increase in Na+, indicating that the blood chemistry remained altered despite the long acclimation period. The elevated pCO2 group also exhibited a shift in their nocturnal swimming pattern from a pattern of many starts and stops to more continuous swimming. Although CO2-exposed teleost fishes can display reduced behavioural asymmetry (lateralization), the CO2-exposed sharks showed increased lateralization. These behavioural effects may suggest that elasmobranch neurophysiology is affected by CO2, as in some teleosts, or that the sharks detect CO2 as a constant stressor, which leads to altered behaviour. The potential direct effects of ocean acidification should henceforth be considered when assessing future anthropogenic effects on sharks.
Keywords: behaviour, respirometry, aerobic scope, blood chemistry, carbon dioxide, Davenport diagram
1. Introduction
Since the beginning of the industrial revolution, anthropogenic use of fossil carbon resources has increased the atmospheric CO2 concentration, reaching 400 μatm pCO2 as of 2013 [1]. The atmospheric CO2 concentration will continue to increase, possibly reaching 940 μatm by 2100 according to the RCP8.5 high-emission scenario [2]. The increase in atmospheric pCO2, with the concurrent rise in surface water pCO2, is shifting the oceanic carbonate ion balance and lowering the pH (i.e. ocean acidification) [3]. Elevated water pCO2 affects a range of organisms, and their responses are highly varied [4].
Teleost fishes can show behavioural disturbances when exposed to elevated CO2 levels. Behaviours such as activity, boldness, lateralization, and responses to olfactory, auditory and visual cues are reported to be altered in some species [5–8], whereas other species appear less sensitive [9,10]. Very little is known about possible effects of ocean acidification on elasmobranchs. It was recently reported that 5 days of exposure to elevated CO2 altered the response to olfactory cues in smooth dogfish (Mustelus canis) [11]. Shark populations are currently in sharp decline globally, and further negative effects of ocean acidification could therefore lead to an increased risk of shark species extinctions as well as reduced ecosystem function [12].
The small-spotted catshark (Scyliorhinus canicula) is a benthic temperate shark with a wide distribution in a broad range of habitats [13]. Here, we tested the hypothesis that elevated CO2 levels affect the behaviour and physiology of S. canicula, similar to the effects reported in teleost fishes. We exposed sharks to either control or a high pCO2 water treatment for 4 weeks, and subsequently examined the behaviour and physiology of the sharks.
2. Material and methods
See the supplementary material for details. In short, 20 small-spotted catsharks were exposed to either control pCO2 (401 μatm) or high pCO2 (993 μatm) conditions for 1 month prior to behavioural and physiological testing. The water pCO2 was measured using infrared (IR) absorbance [9]. The swimming pattern of the sharks was filmed overnight using IR light and IR cameras, and behaviour parameters were calculated and statistically analysed. Lateralization was measured in a double T-maze according to Jutfelt et al. [6] with 40 turning decisions per shark. Relative and absolute lateralization indexes were calculated [14]. Oxygen consumption rates (resting and maximum) were obtained using intermittent flow respirometry, and aerobic scope was calculated [15]. Blood pH was analysed immediately after collection by venepuncture, and plasma was analysed for total CO2. Plasma concentrations of K+, Na+, Ca2+ and Cl− were measured using an ion radiometer biosensor. Pectoral fin skin samples were analysed for denticle morphology and surface imperfections using scanning electron microscopy. The sharks used in this study were returned to the public aquarium from which they were borrowed. Data are presented as mean ± SE.
3. Results
(a). Physiology
Blood plasma values are provided in table 1. 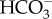 levels were significantly different (p < 0.0001) between control and CO2-exposed sharks (figure 1a). Plasma Na+ concentrations were elevated (p = 0.034) in the CO2 treatment. Growth, Fulton's condition and mortality did not differ significantly (table 2). The metabolic parameters (resting and maximum oxygen consumption, absolute and factorial aerobic scope) did not differ between treatments (figure 1b). The denticles lacked significant differences in length (p = 0.666) and width (p = 0.584) between the control and CO2-exposed animals (figure 1c). Females displayed significantly larger denticles than did the males (pwidth, length, ratio < 0.0001, mixed model analysis); and there was no correlation with shark weight. No ultrastructure damage or unusual growth of denticles was observed for any of the sharks.
levels were significantly different (p < 0.0001) between control and CO2-exposed sharks (figure 1a). Plasma Na+ concentrations were elevated (p = 0.034) in the CO2 treatment. Growth, Fulton's condition and mortality did not differ significantly (table 2). The metabolic parameters (resting and maximum oxygen consumption, absolute and factorial aerobic scope) did not differ between treatments (figure 1b). The denticles lacked significant differences in length (p = 0.666) and width (p = 0.584) between the control and CO2-exposed animals (figure 1c). Females displayed significantly larger denticles than did the males (pwidth, length, ratio < 0.0001, mixed model analysis); and there was no correlation with shark weight. No ultrastructure damage or unusual growth of denticles was observed for any of the sharks.
Table 1.
Effects of CO2 on blood parameters of S. canicula after 4 weeks of treatment. Control n = 6, CO2 n = 6. Italic numbers indicate a significant difference (p < 0.05).
| blood parameter | control | CO2 | significance |
|---|---|---|---|
| pH | 7.82 ± 0.04 | 7.75 ± 0.06 | p = 0.313 |
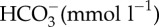 |
4.70 ± 0.39 | 7.68 ± 0.13 | p < 0.0001 |
| K+ (mmol l−1) | 3.28 ± 0.13 | 3.27 ± 0.25 | p = 0.969 |
| Ca2+ (mmol l−1) | 2.89 ± 0.02 | 2.88 ± 0.04 | p = 0.804 |
| Cl− (mmol l−1) | 236.28 ± 3.67 | 243.33 ± 1.74 | p = 0.113 |
| Na+ (mmol l−1) | 257.39 ± 2.29 | 266.17 ± 2.74 | p = 0.034 |
| haemoglobin (g l−1) | 29.11 ± 1.69 | 27.75 ± 0.93 | p = 0.534 |
| haematocrit | 15.58 ± 1.24 | 15.17 ± 0.28 | p = 0.740 |
| MCHC (g l−1) | 19.44 ± 2.05 | 18.34 ± 0.72 | p = 0.660 |
Figure 1.

Effects of pCO2 on the physiology of S. canicula. (a) Davenport diagram showing plasma 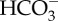 levels, pCO2 and pH after 4 weeks of exposure to control water (400 µatm CO2) or ocean acidification conditions (990 µatm CO2). Asterisks indicate a significant difference (p < 0.05). (b) Oxygen consumption rates. Control n = 7, CO2
n = 9. Resting metabolic rate (RMR), maximum metabolic rate (MMR) and aerobic scope are indicated by brackets. Denticle scanning electron micrographs showing control (c) and CO2-exposed (d) pectoral fin denticles from adults. Image (e) shows a single dorsal denticle from the CO2-exposed group with no visible damage or divergent growth. (Online version in colour.)
levels, pCO2 and pH after 4 weeks of exposure to control water (400 µatm CO2) or ocean acidification conditions (990 µatm CO2). Asterisks indicate a significant difference (p < 0.05). (b) Oxygen consumption rates. Control n = 7, CO2
n = 9. Resting metabolic rate (RMR), maximum metabolic rate (MMR) and aerobic scope are indicated by brackets. Denticle scanning electron micrographs showing control (c) and CO2-exposed (d) pectoral fin denticles from adults. Image (e) shows a single dorsal denticle from the CO2-exposed group with no visible damage or divergent growth. (Online version in colour.)
Table 2.
Effects of CO2 on growth and mortality of S. canicula. Control n = 6, CO2 n = 8 for standard growth rates (SGR; W, weight; L, length; BL, body length) and Fulton's condition. Control n = 10, CO2 n = 10 for mortality measurements.
| parameter | control | CO2 | significance |
|---|---|---|---|
| SGR W (% W d−1) | 0.21 ± 0.04 | 0.40 ± 0.08 | p = 0.098 |
| SGR L (% BL d−1) | 0.04 ± 0.02 | 0.11 ± 0.04 | p = 0.090 |
| mortality (%) | 20 | 10 | p = 0.343 |
| Fulton's condition | 0.30 ± 0.01 | 0.30 ± 0.01 | p = 0.768 |
(b). Behaviour
The absolute lateralization was significantly different (p < 0.05) between the control and CO2-exposed sharks (figure 2a). The CO2-exposed animals expressed a longer swimming time per swimming event (figure 2b), with significantly fewer swimming events compared with the control animals (p = 0.014, mixed model analysis). Sharks in the CO2-exposed group had a tendency towards a higher proportion of swimming events reaching the surface (p = 0.082, mixed model analysis).
Figure 2.
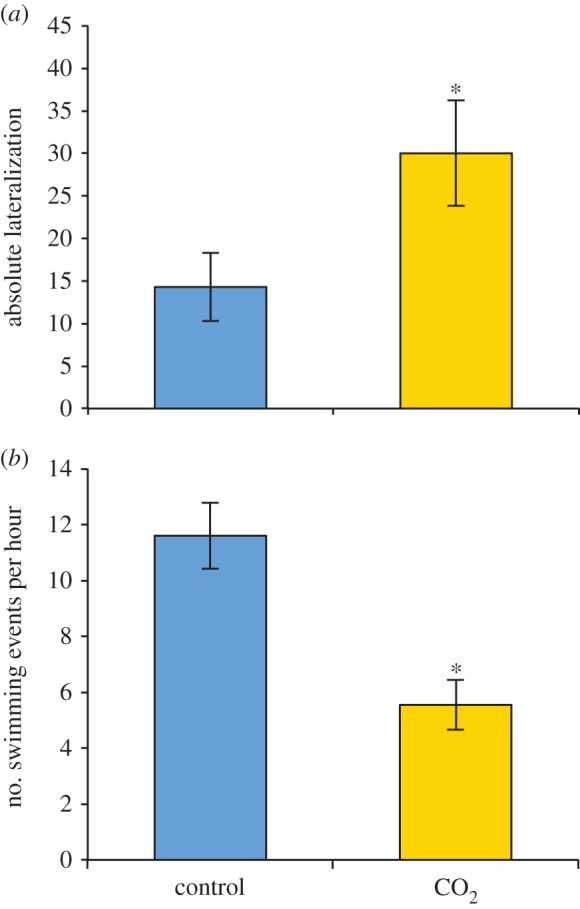
Effects of pCO2 on the behaviour of S. canicula. (a) Absolute lateralization. Control n = 7, CO2 n = 10. The total average number of swimming events during the dark period is shown in figure (b). Asterisks indicate a significant difference (p < 0.05). Control n = 6, CO2 n = 5. (Online version in colour.)
4. Discussion
Here, we show that pCO2 levels possible for the end of the century disturb the behaviour of small-spotted catsharks. The control sharks showed a nocturnal swimming pattern characterized by many starts and stops, whereas sharks in the high CO2-exposed group displayed more continuous swimming with fewer but longer swimming bouts. It is therefore possible that behavioural disturbance could become a consequence of future ocean acidification. The increased swimming duration mimics the altered behaviour that has been previously described in some teleost fishes, as hyperactivity has been recorded as one common behavioural symptom after exposure to elevated CO2 levels [5,8,16]. In teleosts, behavioural disturbances from CO2 exposure are hypothesized to be caused by a dysfunction of GABAergic neurons in the central nervous system (CNS) [17]. This change in behaviour may be caused by a shift in plasma Cl− and 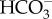 ion concentrations, which leads to reduced hyperpolarization of neurons of high CO2-exposed fish. The GABAA receptor is present in all vertebrates, and the altered shark behaviour could therefore be caused by reduced inhibition of CNS motor regions. The plasma levels of
ion concentrations, which leads to reduced hyperpolarization of neurons of high CO2-exposed fish. The GABAA receptor is present in all vertebrates, and the altered shark behaviour could therefore be caused by reduced inhibition of CNS motor regions. The plasma levels of 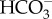 were dramatically elevated in the sharks, whereas the Cl− levels were not significantly affected, suggesting that
were dramatically elevated in the sharks, whereas the Cl− levels were not significantly affected, suggesting that 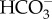 is responsible for the majority of the behavioural shift. However, an alternative explanation for the altered swimming behaviour is that the sharks were continuously detecting the high CO2 levels and altered their swimming behaviour to search for better water quality. Such CO2 avoidance behaviour has recently been described in Atlantic cod [9].
is responsible for the majority of the behavioural shift. However, an alternative explanation for the altered swimming behaviour is that the sharks were continuously detecting the high CO2 levels and altered their swimming behaviour to search for better water quality. Such CO2 avoidance behaviour has recently been described in Atlantic cod [9].
Asymmetric behaviour, where an individual prefers one side, is common in vertebrates and teleost fishes exposed to elevated CO2 typically show reduced lateralization [6,14]. The sharks displayed the opposite effect, with significantly increased lateralization. This effect could be due to a disruption of the almost perfect behavioural symmetry observed in the control group, leading to a random distribution of behavioural asymmetry in the elevated CO2 group.
The sharks appeared physiologically robust and were able to maintain normal oxygen consumption at high pCO2, both at rest and after a swim challenge, and the aerobic scope was not affected. In teleost fishes, the aerobic scope can be reduced [18], unaffected [19] or higher [20,21] in high pCO2 water, and it is possible that there could be similar species differences within elasmobranchs. Denticles consist of hydroxylapatite, which has a low solubility and is not likely affected by weak acids [22]. The larger denticle size of females is likely for protection from male aggression during courtship [23].
Sharks, like teleosts [24], counter acidosis by gill H+/Na+ and 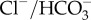 counter-exchange, as previously demonstrated for sharks after short-term exposure to very high pCO2 levels [25]. The significant increases in plasma Na+ and
counter-exchange, as previously demonstrated for sharks after short-term exposure to very high pCO2 levels [25]. The significant increases in plasma Na+ and 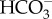 concentrations in this study show that the same mechanism is responsible for pH regulation under near-future CO2 levels and that the ion concentrations remain altered after long-term exposure. It is possible that the new plasma ion levels are within the tolerance range for the sharks, but sub-lethal long-term impacts of altered ion levels cannot be excluded.
concentrations in this study show that the same mechanism is responsible for pH regulation under near-future CO2 levels and that the ion concentrations remain altered after long-term exposure. It is possible that the new plasma ion levels are within the tolerance range for the sharks, but sub-lethal long-term impacts of altered ion levels cannot be excluded.
In conclusion, we demonstrated that plasma pH during long-term exposure to ocean acidification conditions was maintained via chronic elevation of 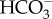 concentrations, with an associated decrease in Na+. Furthermore, shark behaviour was altered and resulted in increased lateralization and a shift in swimming from a pattern of many starts and stops to more continuous swimming. Elasmobranchs may therefore be susceptible to the effects of future ocean acidification.
concentrations, with an associated decrease in Na+. Furthermore, shark behaviour was altered and resulted in increased lateralization and a shift in swimming from a pattern of many starts and stops to more continuous swimming. Elasmobranchs may therefore be susceptible to the effects of future ocean acidification.
Supplementary Material
Acknowledgements
We are grateful to Emma Nohrén and Helle Andersen at Havets Hus, Lysekil, Sweden, for lending us the sharks. We also thank Isabel Casties and Charlotte Alvord for excellent technical assistance. We are grateful to Paolo Domenici for constructive discussions regarding our data.
Funding statement
The Swedish Research Council Formas funded the study.
References
- 1.Keeling R. 2014. A daily record of atmospheric carbon dioxide from Scripps Institution of Oceanography at UC San Diego See keelingcurve.ucsd.edu.
- 2.Stocker TF, et al. 2013. Climate Change 2013: The physical basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK.
- 3.Caldeira K, Wickett ME. 2003. Oceanography: anthropogenic carbon and ocean pH. Nature 425, 365 ( 10.1038/425365a) [DOI] [PubMed] [Google Scholar]
- 4.Kroeker KJ, Kordas RL, Crim R, Hendriks IE, Ramajo L, Singh GS, Duarte CM, Gattuso J-P. 2013. Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming. Glob. Change Biol. 19, 1884–1896. ( 10.1111/gcb.12179) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Forsgren E, Dupont S, Jutfelt F, Amundsen T. 2013. Elevated CO2 affects embryonic development and larval phototaxis in a temperate marine fish. Ecol. Evol. 3, 3637–3646. ( 10.1002/ece3.709) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Jutfelt F, de Souza KB, Vuylsteke A, Sturve J. 2013. Behavioural disturbances in a temperate fish exposed to sustained high-CO2 levels. PLoS ONE 8, e65825 ( 10.1371/journal.pone.0065825) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Munday PL, Donelson JM, Dixson DL, Endo GGK. 2009. Effects of ocean acidification on the early life history of a tropical marine fish. Proc. R. Soc. B 276, 3275–3283. ( 10.1098/rspb.2009.0784) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Munday PL, Pratchett MS, Dixson DL, Donelson JM, Endo GGK, Reynolds AD, Knuckey R. 2012. Elevated CO2 affects the behavior of an ecologically and economically important coral reef fish. Mar. Biol. 160, 2137–2144. ( 10.1007/s00227-012-2111-6) [DOI] [Google Scholar]
- 9.Jutfelt F, Hedgärde M. 2013. Atlantic cod actively avoid CO2 and predator odour, even after long-term CO2 exposure. Front. Zool. 10, 81 ( 10.1186/1742-9994-10-81) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Maneja RH, et al. 2012. The swimming kinematics of larval Atlantic cod, Gadus morhua L., are resilient to elevated seawater pCO2. Mar. Biol. 160, 1963–1972. ( 10.1007/s00227-012-2054-y) [DOI] [Google Scholar]
- 11.Dixson DL, Jennings AR, Atema J, Munday PL. In press. Odor tracking in sharks is reduced under future ocean acidification conditions. Glob. Change Biol. ( 10.1111/gcb.12678) [DOI] [PubMed] [Google Scholar]
- 12.Ferretti F, Worm B, Britten GL, Heithaus MR, Lotze HK. 2010. Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 13, 1055–1071. ( 10.1111/j.1461-0248.2010.01489.x) [DOI] [PubMed] [Google Scholar]
- 13.Ellis JR, Shackley SE. 1997. The reproductive biology of Scyliorhinus canicula in the Bristol Channel, UK. J. Fish Biol. 51, 361–372. ( 10.1111/j.1095-8649.1997.tb01672.x) [DOI] [Google Scholar]
- 14.Domenici P, Allan B, McCormick MI, Munday PL. 2012. Elevated carbon dioxide affects behavioural lateralization in a coral reef fish. Biol. Lett. 8, 78–81. ( 10.1098/rsbl.2011.0591) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Clark TD, Sandblom E, Jutfelt F. 2013. Aerobic scope measurements of fishes in an era of climate change: respirometry, relevance and recommendations. J. Exp. Biol. 216, 2771–2782. ( 10.1242/jeb.084251) [DOI] [PubMed] [Google Scholar]
- 16.Munday PL, Cheal AJ, Dixson DL, Rummer JL, Fabricius KE. 2014. Behavioural impairment in reef fishes caused by ocean acidification at CO2 seeps. Nat. Clim. Change 4, 487–492. ( 10.1038/nclimate2195) [DOI] [Google Scholar]
- 17.Nilsson GE, Dixson DL, Domenici P, McCormick MI, Sørensen C, Watson S-A, Munday PL. 2012. Near-future carbon dioxide levels alter fish behaviour by interfering with neurotransmitter function. Nat. Clim. Change 2, 201–204. ( 10.1038/nclimate1352) [DOI] [Google Scholar]
- 18.Munday PL, Crawley NE, Nilsson GE. 2009. Interacting effects of elevated temperature and ocean acidification on the aerobic performance of coral reef fishes. Mar. Ecol. Prog. Ser. 388, 235–242. ( 10.3354/meps08137) [DOI] [Google Scholar]
- 19.Melzner F, Göbel S, Langenbuch M, Gutowska MA, Pörtner HO, Lucassen M. 2009. Swimming performance in Atlantic cod (Gadus morhua) following long-term (4–12 months) acclimation to elevated seawater PCO2. Aquat. Toxicol. 92, 30–37. ( 10.1016/j.aquatox.2008.12.011) [DOI] [PubMed] [Google Scholar]
- 20.Gräns A, et al. 2014. Aerobic scope fails to explain the detrimental effects on growth resulting from warming and elevated CO2 in Atlantic halibut. J. Exp. Biol. 217, 711–717. ( 10.1242/jeb.096743) [DOI] [PubMed] [Google Scholar]
- 21.Rummer JL, Stecyk JAW, Couturier CS, Watson SA, Nilsson GE, Munday PL. 2013. Elevated CO2 enhances aerobic scope of a coral reef fish. Conserv. Physiol. 1, cot023 ( 10.1093/conphys/cot023) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen ZF, Darvell BW, Leung VWH. 2004. Hydroxyapatite solubility in simple inorganic solutions. Arch. Oral Biol. 49, 359–367. ( 10.1016/j.archoralbio.2003.12.004) [DOI] [PubMed] [Google Scholar]
- 23.Crooks N, Babey L, Haddon WJ, Love AC, Waring CP. 2013. Sexual dimorphisms in the dermal denticles of the lesser-spotted catshark, Scyliorhinus canicula (Linnaeus, 1758). PLoS ONE 8, e76887 ( 10.1371/journal.pone.0076887.t001) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Heuer RM, Grosell M. In press. Physiological impacts of elevated carbon dioxide and ocean acidification on fish. Am. J. Physiol. Regulatory, Integrative Comp. Physiol. ( 10.1152/ajpregu.00064.2014) [DOI] [PubMed] [Google Scholar]
- 25.Claiborne JB, Evans DH. 1992. Acid-base balance and ion transfers in the spiny dogfish (Squalus acanthias) during hypercapnia: a role for ammonia excretion. J. Exp. Zool. 261, 9–17. ( 10.1002/jez.1402610103) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.