

Abstract
Background and Aims
Parasitic plants obtain nutrients from their hosts through organs called haustoria. The hyaline body is a specialized parenchymatous tissue occupying the central parts of haustoria in many Orobanchaceae species. The structure and functions of hyaline bodies are poorly understood despite their apparent necessity for the proper functioning of haustoria. Reported here is a cell wall-focused immunohistochemical study of the hyaline bodies of three species from the ecologically important clade of rhinanthoid Orobanchaceae.
Methods
Haustoria collected from laboratory-grown and field-collected plants of Rhinanthus minor, Odontites vernus and Melampyrum pratense attached to various hosts were immunolabelled for cell wall matrix glycans and glycoproteins using specific monoclonal antibodies (mAbs).
Key Results
Hyaline body cell wall architecture differed from that of the surrounding parenchyma in all species investigated. Enrichment in arabinogalactan protein (AGP) epitopes labelled with mAbs LM2, JIM8, JIM13, JIM14 and CCRC-M7 was prominent and coincided with reduced labelling of de-esterified homogalacturonan with mAbs JIM5, LM18 and LM19. Furthermore, paramural bodies, intercellular deposits and globular ergastic bodies composed of pectins, xyloglucans, extensins and AGPs were common. In Rhinanthus they were particularly abundant in pairings with legume hosts. Hyaline body cells were not in direct contact with haustorial xylem, which was surrounded by a single layer of paratracheal parenchyma with thickened cell walls abutting the xylem.
Conclusions
The distinctive anatomy and cell wall architecture indicate hyaline body specialization. Altered proportions of AGPs and pectins may affect the mechanical properties of hyaline body cell walls. This and the association with a transfer-like type of paratracheal parenchyma suggest a role in nutrient translocation. Organelle-rich protoplasts and the presence of exceptionally profuse intra- and intercellular wall materials when attached to a nitrogen-fixing host suggest subsequent processing and transient storage of nutrients. AGPs might therefore be implicated in nutrient transfer and metabolism in haustoria.
Keywords: Haustorium, hyaline body, arabinogalactan proteins, AGPs, rhinanthoid Orobanchaceae, cell wall, hemiparasite, parasitic plant, immunocytochemistry, paramural body, globular ergastic body, Rhinanthus minor, Odontites vernus, Melampyrum pratense
INTRODUCTION
Parasitic plants are defined as plants that possess haustoria, i.e. grafting organs allowing attachment to host plant organs and infiltration of their vasculature for nutrients (Kuijt, 1969; Heide-Jørgensen, 2008). Several types of haustoria have evolved in different lineages of parasitic plants but in all cases they are primarily parenchymatous organs traversed by a central strand of xylem connecting the vascular systems of the parasite and the host.
Haustorial structure has been very well researched in the Orobanchaceae, the largest monophyletic clade of parasitic plants (Těšitel et al., 2010), having ∼2060 species of facultative and obligatory hemiparasites and holoparasites and one non-parasitic genus, Lindenbergia (McNeal et al., 2013). In some species within the family, the central haustorial region surrounding the xylem bridge is occupied by a distinctive type of parenchymatous tissue known as the hyaline body, alternatively referred to as the central parenchymatous core (Musselman and Dickison, 1975) or haustorial nucleus (Nwoke, 1982). The hyaline body is absent from some species within the Orobanchaceae, e.g. Striga gesneroides (Ba, 1983) and Rhamphicarpa fistulosa (Neumann et al., 1999). The total number of Orobanchaceae species with hyaline bodies has not been determined but the species that are currently known to possess them are distributed across different major clades within the family. Outside the Orobanchaceae, hyaline bodies have been reported from Thesium chinense (Suetsugu et al., 2008) and Santalum album (Tennakoon and Cameron, 2006), both members of the Santalaceae.
Some of the special characteristics of the hyaline body include a relatively small cell size and denser, darkly stained cytoplasm (Visser et al., 1984; Gurney et al., 2003; Ndambi et al., 2011), large nuclei (Visser et al., 1984; Riopel and Timko, 1995; Ndambi et al., 2011; Brokamp et al., 2012) and cell wall modifications in the form of various types of thickenings (Dobbins and Kuijt, 1973; Renaudin and Cheguillaume, 1977; Visser et al., 1984; Mallaburn and Stewart, 1987). Some of these features have often led to the tissue being described as highly metabolically active (Visser et al., 1984; Gurney et al., 2001; Rümer et al., 2007).
The hyaline body has long been presumed to be an important part of the haustorial machinery. However, there is insufficient structural, and practically no functional, evidence to support any specific roles. Only one previous publication deals specifically with the structure of the hyaline body and describes organelle-rich cytoplasm and extensive paramural deposits in Alectra orobanchoides (Visser et al., 1984). Some detailed anatomical accounts of the tissue have also been provided by other authors who discussed the wider haustorial structure. A collenchyma core was described in the upper haustorium of Castilleja sp. (Dobbins and Kuijt, 1973), where cell wall thickenings and paramural bodies were found in the centrally located tissue, later described as collenchyma-containing central parenchymatous core by Musselman and Dickison (1975). The collenchymatous character of these modifications is questionable and similar structures were subsequently found in the hyaline bodies of Thesium humifusum (Renaudin and Cheguillaume, 1977) and Striga sp. (Mallaburn and Stewart, 1987).
While no experimental evidence is available to demonstrate the functions of the hyaline body, roles in nutrient transfer, processing and storage have been proposed (Renaudin and Cheguillaume, 1977; Visser et al., 1984; Mallaburn and Stewart, 1987; Riopel and Timko, 1995; Runo et al., 2012). In Rhinanthus minor (yellow rattle), the hyaline body has been suggested to synthesize abscisic acid, which is found abundantly in this tissue and has been proposed to regulate haustorial cell wall thickening and lignification (Jiang et al., 2004; Rümer et al., 2007). A role in starch storage and protein synthesis by the hyaline bodies of Striga asiatica was implied based on the abundant rough endoplasmic reticulum and amyloplasts respectively (Mallaburn and Stewart, 1987). Modification of nutrients derived from the host is in agreement with the observation that the proportion of amino acids to inorganic nitrogen in the haustorial xylem fluid is higher than in the penetrated host xylem fluid (Govier et al., 1967).
It has been demonstrated that the structure of the hyaline bodies in Striga hermonthica (witch weed) (Arnaud et al., 1999; Gurney et al., 2003, 2006) and R. minor (Cameron et al., 2006; Cameron and Seel, 2007; Rümer et al., 2007) is strongly affected by the level of compatibility with the host, whereby the tissue develops poorly on incompatible hosts. Differences in the development of haustoria on hosts and non-hosts have, in turn, important implications for agrosystems and the development of crop resistance as well as for seminatural ecosystems where the parasites occur. Parasitic plants that are agricultural weeds effect huge crop and financial losses, with S. hermonthica alone costing over $1 billion annually (Parker, 2009, 2012). In non-agricultural ecosystems, parasitic plants can have a pronounced effect on community structure through mediating competitive balance between hosts and non-hosts (Press and Phoenix, 2005). The most researched example is the effect of R. minor on grassland productivity and biodiversity. Rhinanthus minor preferentially parasitizes grasses and legumes, leaving most dicots unaffected and with comparatively increased competitive ability (Gibson and Watkinson, 1991, 1992; Davies, 1997; Ameloot et al., 2005; Ameloot, 2007). This ultimately leads to increased floral diversity as many dicots, which would otherwise be outcompeted by grasses, can colonize and exist in a Rhinanthus-rich grassland (Davies, 1997; Pywell et al., 2004; Bullock and Pywell, 2005; Westbury et al., 2006). Previous histochemical studies of R. minor haustoria attached to non-hosts show that, in addition to lacking xylem bridges, they possess poorly developed hyaline bodies (Cameron et al., 2006; Cameron and Seel, 2007; Rümer et al., 2007). However, the tissue has not been characterized in detail in this species. As the hyaline body is one of the most strongly affected elements of the haustorium during incompatible reactions, its biology might provide key insights into parasite–host interactions and their large scale consequences.
This article focuses on the structure and composition of hyaline body cell walls in three species of the rhinanthoid Orobanchaceae (Rhinantheae), a clade of the Orobanchaceae grouping ecologically important genera (Těšitel et al., 2010a; Těšitel, 2011). We investigated the hyaline bodies of R. minor, Odontites vernus (red bartsia) and Melampyrum pratense (common cow wheat). Based on the first extensive immunohistochemical in situ screen of the hyaline body in any parasitic species, we present a detailed characterization of its unusual features that may help direct future studies and form hypotheses about the functions of this tissue.
MATERIALS AND METHODS
Plant material
Rhinanthus minor (seeds collected in Roscahill, County Galway and Lusmagh, County Offaly, Ireland) and Odontites vernus (collected in the Burren, County Clare and in Galway City, Ireland) were grown in the laboratory with compatible hosts Arrhenatherum elatius ssp. bulbosum, referred to as Arrhenatherum elatius throughout the article (collected at Inis Mór, County Clare, Ireland), and Vicia sepium (collected in the Burren, County Clare, Ireland). Rhinanthus minor was also paired with non-hosts Plantago lanceolata and Prunella vulgaris (collected in Roscahill, County Galway, Ireland). Melampyrum pratense haustoria were harvested from plants growing in Carron, County Clare, Ireland. Because haustoria were dislodged from the host roots during collection, host species could not be determined.
Seeds of R. minor and O. vernus were sterilized by immersion in 30 % v/v thick Domestos bleach in distilled water (working concentration of sodium hypochlorite 1·35 % w/v) for 10 min, washed in several changes of distilled water and placed in Petri dishes on filter paper (Whatman No. 1) moistened with 0·1 % Bravo500 fungicide (Syngenta) in distilled water at a density of 25 seeds per dish for R. minor and 50 seeds for O. vernus. Seeds were cold-treated for 8–12 weeks in the dark at a constant temperature of 4 °C (in a fridge) or under a 4 °C/0 °C regime (12 h/12 h) in a Binder growth chamber model KBW720 (www.binder-world.com).
Host seeds were sterilized as above. Bare caryopses of A. elatius ssp. bulbosum and seeds of P. lanceolata were placed on moist filter paper and germinated abundantly within several days. A germination rate of 30 % after 3 weeks was reached by P. vulgaris using the same method. Scarification for 30 min in concentrated sulphuric acid (10276, BDH) was necessary for the germination of V. sepium.
Sample preparation
Haustoria were fixed immediately after collection in 2 % v/v formaldehyde and 2·5 % glutaraldehyde in PEM buffer [50 mm piperazine-N, N′-bis(2-ethane-sulphonic acid) (PIPES), 5 mm ethylene glycol bis(β-aminoethylether)-N, N, N′, N′-tetraacetic acid (EGTA), 5 mm MgSO4, pH 6·9] for 2 h with the initial 30 min of vacuum infiltration. Samples were washed in three changes of phosphate-buffered saline (PBS) for 5 min each wash with some samples additionally post-fixed in 1 % w/v osmium tetroxide in PEM followed by washing in PBS. Osmicated and non-osmicated material was dehydrated at 4 °C in a graded ethanol series (30, 50, 70, 90 and 100 %) with an overnight step in 70 % ethanol and all other steps at 1 h each. The samples were subsequently infiltrated in London Resin medium grade (Agar Scientific) with 30, 50, 70, 90 and 100 % steps (1–2 h per step) and an overnight step in 100 % resin. Resin (100 %) was subsequently changed and samples left for up to several days. Samples were then polymerized in gelatin capsules at 60 °C for 2 d.
Immunohistochemistry
For immunofluorescence, non-osmicated samples were sectioned with a histo diamond knife (Diatome) at 0·5 μm and mounted on polylysine-coated or Vectabond-treated slides. Samples were immunolabelled with an array of 32 monoclonal antibodies (mAbs) to cell wall glycan epitopes. Sections were blocked for 30 min in a 1 % w/v milk protein or bovine serum albumin solution in PBS buffer, washed two times in PBS buffer (5 min per step) and incubated for 1·5 h in 1:10 dilutions of the primary antibodies in the blocking solution. Anti-callose antibody 400-2 (Biosupplies) was diluted at 8 μg ml–1. Following three washing steps in PBS (5 min per step), the sections were incubated in the secondary antibody (rat or mouse) linked to FITC fluorochrome for 1·5 h. Sections were then washed in PBS and post-stained in 0·2 % toluidine blue O in 1 % aqueous borax to eliminate intense autofluorescence. Finally, the samples were mounted in Citifluor AF2 antifade mounting reagent (Agar Scientific) and observed with an Olympus BX51 epifluorescence microscope with an X-Cite® 120Q mercury lamp. The images were acquired using an Olympus XC10 digital camera and Cell B software.
A number of samples were pretreated to remove pectins prior to immunolabelling to investigate the possibility of epitope masking (Jauh and Lord, 1996; Marcus et al., 2008; Hervé et al., 2011). Sections were incubated in 0·1 m sodium carbonate (S7795, Sigma) at pH 11·4 for 2 h to remove methyl groups and make the polymers more vulnerable to the action of unmasking enzymes. The slides were subsequently washed in PBS and treated with Pectinex® Ultra SPL (pectinase from Aspergillus aculeatus, P2611, Sigma) in N-cyclohexyl-3-aminopropanesulphonic acid (CAPS) buffer (50 mm CAPS, 2 mm CaCl2, pH 10) at 1000 units of enzyme ml–1 or pectate lyase in CAPS buffer at 10 mg mL–1 (1 in 75 v/v) for 2 h.
Electron microscopy
For transmission electron microscopy (TEM), non-osmicated and osmicated material was used. Sections of silver to silvery-gold colour (∼60–80 μm) were collected on Formvar-coated nickel slot grids. Samples were optionally stained with lead citrate and uranyl acetate. However, as the contrast of samples was generally good, this procedure was often eliminated to minimize the occurrence of precipitates. TEM images were taken using a Zeiss Libra® 120 (120 kV) with a slow-scan CCD digital camera (Albert Tröndle Restlichtverstärkersysteme) at Skidmore Microscopy and Imaging Center (USA) as well as a Hitachi H7000 (75 kV working voltage) with a Hamamatsu 1 K digital camera and AMT image acquisition software at the Centre for Microscopy and Imaging, NUI Galway.
RESULTS
Hyaline body development and cell wall epitope composition
Clearly marked hyaline bodies with distinctive cell wall architecture developed in haustoria of R. minor and O. minor attached to compatible hosts and in M. pratense haustoria collected in the field from unknown hosts. Hyaline bodies in haustoria of R. minor attached to incompatible hosts P. lanceolata and P. vulgaris were poorly developed and much more difficult to distinguish than in haustoria attached to the compatible hosts. The results presented in the main body of this article refer to the compatible pairings, while a short account of hyaline body structure and cell wall architecture in association with non-hosts is provided in Supplementary Data Fig. S1.
Figure 1 shows a diagram and micrographs illustrating compatible pairings and hyaline body location in a longitudinal cross-section, i.e. when the haustorium is sectioned longitudinally (parallel to the secondary xylem axis) and the host root transversely. The hyaline body occupied the central part of the haustorium and was ovoid to spherical in R. minor (Fig. 1A, B) and M. pratense (Fig. 1D). Hyaline bodies of O. vernus (Fig. 1C) were more cylindrical and often made up of particularly small and regularly stacked cells of exceptionally dense (darkly stained) appearance. The hyaline body in all compatible pairings was composed of isodiametric parenchyma cells, with large nuclei and dense cytoplasm often being prominent (Fig. 1A). However, in some mature haustoria large central vacuoles filled most of the cell volume, pushing the nuclei against the cell walls. In haustoria attached to compatible hosts, a layer of paratracheal parenchyma developed between the hyaline body and the xylem bridge. It abutted the xylem bridge along the hyaline body-traversing part and above and was characterized by prominent cell wall thickenings in contact with the xylem (see the lower micrograph detail in Fig. 1A). A range of more elaborate thickenings were occasionally found and are described in detail in Supplementary Data Fig. S2.
Fig. 1.

Overview of hyaline body location and structure. (A) Diagram illustrating the position of the hyaline body (HB) in a mature haustorium of R. minor. The hyaline body is centrally located and is composed of relatively small, isodiametric cells characterized by large nuclei (black) and dense cytoplasm (light grey). Cortical parenchyma (COR) is strongly vacuolated and therefore the nuclei and protoplasts are not apparent. The difference between the hyaline body and cortical parenchyma is clearly visible in the micrographic detail. A strand of paratracheal parenchyma (PP) with thickened cell walls (marked with dark grey in the diagram and an asterisk symbol in the micrographic detail) is associated with the haustorial xylem where it traverses the hyaline body and above. No cells except xylem are shown in the host root. HX, host xylem; CF, clasping fold; END, endophyte; XYL, xylem bridge; PP, paratracheal parenchyma; INT, interface between the haustorium and the host root. (B, C, D) Micrographs showing outlines (dashed line) of hyaline bodies in R. minor (B), O. vernus (C) and M. pratense (D). Longitudinal cross-sections are stained with toluidine blue O. (A, C, E) 10 μm sections from Steedman's wax-embedded samples.
The hyaline body of R. minor was not visible as a distinct structure in the pre-infiltration developmental stages (Fig. 2A, B). It was, however, clearly marked in haustoria during the penetration of host cortex (Fig. 2E), as the cells in the hyaline body region were tightly packed and possessed large nuclei with distinctive nucleoli (Fig. 2F). Numerous ribosomes could be observed in the cytosol and cell walls were thin (Fig. 2G). During the penetration of host endodermis but prior to haustorial xylem differentiation (Fig. 2H), relatively large intercellular spaces filled with wall material became apparent (Fig. 2I, J). The structure of the hyaline body became more clearly defined while the protoplast remained dense. This dense cytoplasm was also present in samples where xylem bridges were largely differentiated but presumably not yet functional, i.e. where the lumina were filled with what appeared to be protoplast remnants following programmed cell death (Fig. 2K–M). Intercellular spaces were prominent and clearly visible at light microscopy level (Fig. 2L).
Fig. 2.

Anatomy of key developmental stages of the hyaline body in R. minor. (A, B) Early morphological differentiation in the haustorial initial (pre-attachment to early contact stage). (C, D) Large nuclei and dense cytoplasm become apparent in the host-facing side of a more advanced haustorial initial. (E, F, G) A distinctively ovoid, ‘dense’ region is apparent in a haustorium that has begun penetrating host cortex but has not reached the endodermis (end). The cells are small and tightly packed, with numerous vacuoles (v) and large nuclei ( ). Cross-cell walls characteristic of recent cell divisions are present (
). Cross-cell walls characteristic of recent cell divisions are present ( ). TEM detail (G) shows that cell walls (cw) are very thin and ribosomes are very abundant in the cytoplasm. (H, I, J) Ovoid tissue region in a haustorium penetrating host endodermis (end), but prior to xylem development, is further differentiated, with prominent intercellular spaces filled with deposits (black arrows). (K, L, M) Hyaline body cells during xylem bridge (x) maturation (remnants of cytoplasm in the lumina) show more extensive vacuolation. The cytoplasm remains dense with e.g. the endoplasmic reticulum (ER) being still abundant.
). TEM detail (G) shows that cell walls (cw) are very thin and ribosomes are very abundant in the cytoplasm. (H, I, J) Ovoid tissue region in a haustorium penetrating host endodermis (end), but prior to xylem development, is further differentiated, with prominent intercellular spaces filled with deposits (black arrows). (K, L, M) Hyaline body cells during xylem bridge (x) maturation (remnants of cytoplasm in the lumina) show more extensive vacuolation. The cytoplasm remains dense with e.g. the endoplasmic reticulum (ER) being still abundant.
Immunodetection with a set of 32 cell wall-specific monoclonal antibodies showed that throughout development the composition of hyaline body cell walls varied from that of the surrounding parenchyma. Table 1 summarizes the distribution of cell wall components within the hyaline body and adjacent tissues. The most distinctive labelling patterns are illustrated in the provided micrographs. None of the monoclonal antibodies used generated non-specific binding to the sections. Therefore, all immunofluorescence images show specific staining with monoclonal antibodies. Control images are presented in Supplementary Data Fig. S3. Images stained with toluidine blue O are included as an anatomical reference and are marked with the same letter as the corresponding epifluorescence micrographs and a prime (′).
Table 1.
Summary of immunolabelling results for R. minor and O. vernus hyaline bodies in mature haustoria attached to compatible hosts
| Specificity | Antibody | Cortex | Hyaline body |
Paratracheal parenchyma |
|||
|---|---|---|---|---|---|---|---|
| Cell walls | Paramural bodies | Ergastic globules | Wall thickenings | Non-thickened walls | |||
| Extensins | LM1 | – | + | – | + | – | – |
| JIM12 | – | – | – | – | – | – | |
| JIM13 | – | – | – | – | – | – | |
| JIM19 | – | – | – | – | – | – | |
| JIM20 | – | + | – | + | – | – | |
| AGP glycans | LM2 | – | ++ | ++ | ++ | – | –/+ |
| JIM8 | –/+ | ++ | ++ | ++ | – | –/+ | |
| JIM13 | –/+ | ++ | ++ | ++ | – | –/+ | |
| JIM14 | –/+ | ++ | ++ | ++ | – | –/+ | |
| CCRC-M7 | + | ++ | + | + | – | –/+ | |
| JIM4 | – | – | – | – | – | – | |
| JIM15 | – | – | – | – | – | – | |
| JIM16 | – | – | – | – | – | – | |
| MAC207 | – | – | – | – | – | – | |
| Pectins | |||||||
| de-esterified HG | JIM5 | ++ | –/+ | ++ | ++ | + (thin layers) | –/+ |
| de-esterified HG | LM18 | ++ | –/+ | ++ | ++ | + (thin layers) | –/+ |
| de-esterified HG | LM19 | ++ | –/+ | ++ | ++ | + (thin layers) | –/+ |
| esterified HG | JIM7 | ++ | ++ | ++ | ++ | + (thick bands) | ++ |
| esterified HG | LM20 | ++ | ++ | ++ | ++ | – | – |
| RG-I backbone | RU-I | –/+ | –/+/++ | – | – | –/+/++ | + |
| RG-I backbone | RU-II | –/++ | –/+/++ | – | – | –/+/++ | +, ++ |
| RG-I galactan side chain | LM5 | –/+/++ | ++ | ++ | ++ | ++ | ++ |
| RG-I arabinan side chain | LM6 | – | –/+ | – | +/++ | –/+ | – |
| Xyloglucan | LM15 | –/+ | –/+ | + | + | + | |
| LM24 | – | –/+ | + | + | –/+ (thin layers) | – | |
| LM25 | – | –/+ | + | + | + (thin layers) | + | |
| CCRC-M1 | – | – | – | – | – | – | |
| Mannan | LM21 | – | – | – | – | – | – |
| LM22 | – | – | – | – | – | – | |
| Xylan | LM10 | – | – | – | – | – | – |
| LM11 | – | – | – | – | – | – | |
EXT, extensin; AGP, arabinogalactan protein; HG, homogalacturonan; RG-I, rhamnogalacturonan-I, XG, xyloglucan.
– no labelling; –/+ no labelling or weak labelling; + weak labelling; ++ strong labelling.
Pectic homogalacturonans (HGs) and arabinogalactan proteins (AGPs) were the most strongly labelled cell wall polymers in haustoria. HGs with high levels of methyl-esterification were most ubiquitous and detected with mAbs JIM7 and LM20 at the same intensity in all investigated developmental stages in the cortex and the hyaline body (Fig. 3C, F, I, L). However, a striking negative correlation between de-esterified homogalacturonan and AGP epitopes was observed in the hyaline body (Figs 3 and 4). The hyaline body cell walls and intercellular spaces became enriched in AGP epitopes detected by antibodies LM2 (Figs 3D, G, J and 4A), JIM8, JIM13 and JIM14 (Table 1) during early developmental stages, after the penetration of cortex had begun. This enrichment in AGPs corresponded with greatly reduced detection of de-esterified HG epitopes in the hyaline body (Figs 3E, H, K and 4B). When the interface between the hyaline body and the cortical parenchyma was observed at high magnification, a very unusual polarized distribution of AGP and de-esterified pectic HG epitopes could be observed in the outermost cell layer of the hyaline body (Fig. 4). AGP labelling gradually decreased towards, and eventually disappeared within, the outermost cell walls of this cell layer (Fig. 4A). The labelling of de-esterified pectins displayed the opposite pattern (Fig. 4B). Therefore, the detection of AGPs was complementary to the detection of de-esterified HG, i.e. it disappeared precisely where the labelling of de-esterified HG began (Fig. 4C). This biochemical polarity was often not associated with any apparent structural differences. AGP labelling of the hyaline body in general was very consistent and allowed precise identification of the extent of the tissue when no obvious anatomical features were present. Enzymatic removal of pectic HG from sections did not alter the detection of AGPs (data not shown).
Fig. 3.

Relationship between homogalacturonan and AGP components of hyaline body cell walls in R. minor at different developmental stages. The image illustrates immunofluorescence of haustoria during penetration of host cortex (A–C) and endodermis (D–F), during maturation of xylem (x) bridge (G–I) and at full maturity (J–L), as presented in Fig. 2. Anatomy reference images of toluidine blue O staining photographed in bright field are marked with the same letter as the corresponding fluorescence images and a prime (′). Sections were immunolabelled with mAbs LM2 (β-linked GalA epitope of AGPs), JIM5 (de-esterified pectins) and JIM7 (esterified HG), as indicated in the top right corner of each immunofluorescence image. The hyaline body (hb) becomes strongly enriched in AGPs approximately around the time of endodermal penetration. Concurrently, detection of de-esterified HG in the hyaline body becomes much reduced in comparison with the cortex (cor). Esterified pectins remain labelled equally in the cortex and hyaline body throughout development.
Fig. 4.
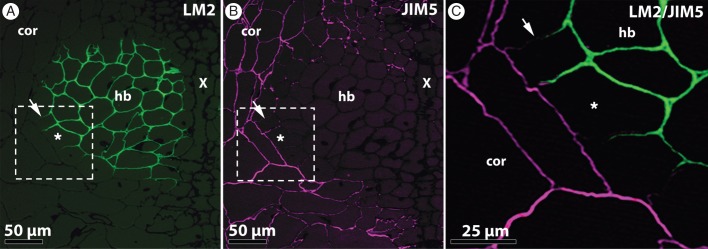
AGP and de-esterified homogalacturonan transition at the hyaline body (hb)–cortex (cor) boundary of R. minor. The magnified region of the samples illustrated in Fig. 3J, K shows that the gradual enrichment in AGPs coincides precisely ( ) with a strong decrease in de-esterified HG labelling. (A) Detail of immunolabelling with LM2 (AGPs), showing polarized heterogeneity of epitope distribution in the outermost cell layer of the hyaline body (hb). X, xylem bridge. (B) Labelling of the same region as in (A) (serial section) with JIM5 (de-esterified HG). Fake magenta colour was applied. (C) Stacked image of (A) and (B). Asterisks indicate a reference cell .
) with a strong decrease in de-esterified HG labelling. (A) Detail of immunolabelling with LM2 (AGPs), showing polarized heterogeneity of epitope distribution in the outermost cell layer of the hyaline body (hb). X, xylem bridge. (B) Labelling of the same region as in (A) (serial section) with JIM5 (de-esterified HG). Fake magenta colour was applied. (C) Stacked image of (A) and (B). Asterisks indicate a reference cell .
Differences between the hyaline body and haustorial cortex were also found in the distribution of pectic rhamnogalacturonan-I (RG-I) backbone and side chain epitopes. However, the consistency of these differences was considerably lower than in the case of HG and AGPs (Table 1). Examples of backbone and side chain epitope detection are illustrated in Supplementary Data Fig. S4. Detection of xyloglucans was generally very low in all haustorial cell walls (Table 1) even after pretreatment with pectate lyase. LM15- and LM25-detected epitopes were in a vast majority of samples confined to paramural bodies and ergastic inclusions described in the next section.
Callose was detected only in the newly deposited walls of the cortex-penetrating haustoria (data not shown), highlighting the meristematic character of the immature hyaline body region during this developmental stage.
Structure and composition of paramural bodies and globular ergastic inclusions
As exemplified in the micrograph detail in Fig. 1A, the hyaline body cell walls in many cases appeared anatomically similar to those of the outer parenchyma. However, a diverse array of cell wall modifications and intracellular globules composed of cell wall material in the cytoplasm (globular ergastic bodies) were common in the hyaline bodies of all three species investigated. In R. minor, discrete paramural bodies and globular inclusions were abundantly found in compatible pairings, particularly with the legume V. sepium (Fig. 5A), but also in some haustoria attached to A. elatius ssp. bulbosum (Fig. 5B–D). Paramural bodies stained darker purple with toluidine blue O than the remaining parts of the walls and the globular structures (Fig. 5A) and originated in a membrane system of endoplasmic reticulum-like tubules connected to the plasma membrane (Fig. 5C). The globular inclusions were, on the other hand, most likely derived from vesicles as similar bodies surrounded by a single membrane were identified in osmicated material. However, it could not be determined whether these vesicles originated at the Golgi network or the endoplasmic reticulum.
Fig. 5.

Structure and major epitopes of paramural bodies and globular ergastic inclusions in hyaline bodies of R. minor. (A, E–J) Sections from the same haustorium attached to a root of V. sepium. Differences in colouration are a result of toluidine blue O dye washing out of the tissues during observations. (B–D) TEM micrographs of an osmicated haustorium attached to A. elatius. (A) The bulk of the cell volume is occupied by vacuoles (v) and large nuclei (n), with marked nucleoli. Paramural bodies ( ) and ergastic globular bodies (arrowheads) within the cytoplasm are abundant. Some material is deposited intercellularly (
) and ergastic globular bodies (arrowheads) within the cytoplasm are abundant. Some material is deposited intercellularly ( ). (B) Ultrastructure of the hyaline body. Numerous mitochondria (m) and dictyosomes (Golgi stacks) (d) can be observed in the cells with abundant paramural deposits. (C) Detail showing deposition of paramural deposits, which appear to be accreted in layers (1) and discrete, ovoid ‘packets’ (2), which continue to be coated by more deposits (3). The packets originate in inflated sections of tubular membranous structures continuous with the plasma membrane. (D) Detail of the ultrastructure of globular bodies showing that they are bound by a single membrane. (E–J) Immunodetection of the most abundant classes of epitopes in the paramural bodies and globular ergastic bodies. Anatomy reference images of toluidine blue O staining photographed in bright field are marked with the same letter as the corresponding fluorescence images and a prime symbol (′). Antibodies: LM19, de-esterified HG; LM25, xyloglucan; LM2, JIM8, JIM13 and CCRC-M7, AGPs. LM2, JIM8 and JIM13 produce the most uniform and strong labelling of the globules. Pectin and xyloglucan are detected in patches while CCRC-M7-specific epitopes concentrate at the surface of the ergastic globules. Labelling with LM2 also appears stronger on the outer surface of the granules. Note that the strong labelling of paramural bodies with LM19 contrasts with lack of detection in the hyaline body cell walls.
). (B) Ultrastructure of the hyaline body. Numerous mitochondria (m) and dictyosomes (Golgi stacks) (d) can be observed in the cells with abundant paramural deposits. (C) Detail showing deposition of paramural deposits, which appear to be accreted in layers (1) and discrete, ovoid ‘packets’ (2), which continue to be coated by more deposits (3). The packets originate in inflated sections of tubular membranous structures continuous with the plasma membrane. (D) Detail of the ultrastructure of globular bodies showing that they are bound by a single membrane. (E–J) Immunodetection of the most abundant classes of epitopes in the paramural bodies and globular ergastic bodies. Anatomy reference images of toluidine blue O staining photographed in bright field are marked with the same letter as the corresponding fluorescence images and a prime symbol (′). Antibodies: LM19, de-esterified HG; LM25, xyloglucan; LM2, JIM8, JIM13 and CCRC-M7, AGPs. LM2, JIM8 and JIM13 produce the most uniform and strong labelling of the globules. Pectin and xyloglucan are detected in patches while CCRC-M7-specific epitopes concentrate at the surface of the ergastic globules. Labelling with LM2 also appears stronger on the outer surface of the granules. Note that the strong labelling of paramural bodies with LM19 contrasts with lack of detection in the hyaline body cell walls.
The biochemical composition of globular ergastic bodies and paramural bodies was similar. The main epitopes detected were those of AGPs, homogalacturonan, galactan and xyloglucan (Fig. 5E–J, Table 1). The globules additionally contained weakly labelled arabinan and extensin epitopes. The pattern of homogalacturonan and xyloglucan labelling was patchier in the globular ergastic bodies than in the paramural bodies. In fact, AGP glycans detected with LM2, JIM8, JIM13 and JIM14 were the most strongly and uniformly detected class of epitopes in the ergastic globules. However, CCRC-M7 labelled AGPs in a very different pattern, with fluorescence restricted to the globule surface (Fig. 5J).
Strong labelling with anti-AGP probes was a common feature of all observed cell wall modifications and ergastic granules in all three species investigated. Figure 6 summarizes a range of such modifications found in haustoria of O. vernus, M. pratense and R. minor, labelled with antibody LM2. Panels A–C in Fig. 6 illustrate the extremely abundant and large ergastic globules found in O. vernus. Cells packed with these membrane-bound structures, filled with grainy substances (Fig. 6B), labelled extremely strongly with LM2 (Fig. 6C), in sharp contrast with the surrounding cortical parenchyma. In M. pratense, a similar effect of AGP-‘filled’ cells was obtained due to the strong labelling of extremely extensive paramural deposits (Fig. 6D). AGP epitopes were also strongly labelled in the intercellular spaces in this species. This was also true for the extremely swollen middle lamellae occasionally found in the hyaline bodies of R. minor (Fig. 6E). AGPs were in fact the only abundantly detected cell wall molecules in the latter type of cell wall modification, in addition to very weak and scattered labelling of HG (data not shown).
Fig. 6.

Diversity of AGP-rich structures in the hyaline bodies of O. vernus, M. pratense and R. minor. Arabinogalactan epitopes were found in all cell wall modifications and ergastic granules in all three species and in all cases they were very strongly detected with LM2. (A) Membrane-bound globular ergastic bodies (arrowhead) of O. vernus were considerably bigger than in R. minor and often occupied most of the cell volume, resulting in sharp contrast with the cortical parenchyma (cor). (B) TEM detail of (A). The bodies differed in the density of their content, which appeared to be granular. They were often associated with dense, osmiophilic bodies (ob). Large vacuoles (v) were filled with looser grainy material. Deposits of similar grainy substance were also found intercellularly ( ). Dictyosomes (d) were present in the scarce cytoplasm (c). Cell wall is indicated by cw. (C–E) Immunolabelling of various cell wall modifications in hyaline bodies with antibody LM2 recognizing AGP glycans. Anatomy reference images of toluidine blue O staining photographed in bright field are marked with the same letter as the corresponding fluorescence images and a prime symbol (′). (C) Hyaline body (hb) of O. vernus attached to A. elatius, with strongly labelled cell walls and ergastic globule-rich cell content. Note that the paratracheal parenchyma (pp) abutting the xylem bridge (xb) did not label. (D) Hyaline body of M. pratense attached to an unknown host, with LM2 labelling of very extensive paramural bodies (
). Dictyosomes (d) were present in the scarce cytoplasm (c). Cell wall is indicated by cw. (C–E) Immunolabelling of various cell wall modifications in hyaline bodies with antibody LM2 recognizing AGP glycans. Anatomy reference images of toluidine blue O staining photographed in bright field are marked with the same letter as the corresponding fluorescence images and a prime symbol (′). (C) Hyaline body (hb) of O. vernus attached to A. elatius, with strongly labelled cell walls and ergastic globule-rich cell content. Note that the paratracheal parenchyma (pp) abutting the xylem bridge (xb) did not label. (D) Hyaline body of M. pratense attached to an unknown host, with LM2 labelling of very extensive paramural bodies ( ), extracellular deposits and, in some cells, the cytoplasm. n, nucleus. (E) Labelling of extremely expanded intercellular spaces found in some hyaline bodies of R. minor attached to V. sepium.
), extracellular deposits and, in some cells, the cytoplasm. n, nucleus. (E) Labelling of extremely expanded intercellular spaces found in some hyaline bodies of R. minor attached to V. sepium.
Cell wall composition in paratracheal parenchyma
As shown in Figs 3J and 6C and Table 1, AGP immunofluorescence was generally absent from or very weak in the cell walls of paratracheal parenchyma. Furthermore, all three classes of cell wall polysaccharides detected in paratracheal parenchyma thickenings, namely homogalacturonans, rhamnogalacturonans and xyloglucans, displayed radial heterogeneity, i.e. they appeared to be concentrated in distinctive layers of varying thickness (Fig. 7B–D). In the case of RG-I backbone and de-esterified HG epitopes, bands of labelling typically correlated with darker toluidine blue O staining (Fig. 7B, C). An exception to the layered pattern was the distribution of RG-I galactan epitope (LM5), which was the only epitope labelled uniformly throughout the thickened walls (Fig. 7A).
Fig. 7.

Immunocytochemistry of paratracheal parenchyma cell walls. Arrows indicate the cytoplasm-facing surface of cell wall thickenings that develop in contact with haustorial xylem (xyl). Panels A, B, C and D show immunofluorescence micrographs. Anatomy reference images of toluidine blue O staining photographed in bright field are marked with the same letter as the corresponding fluorescence images and a prime symbol (′). (A) LM5-detected galactan of RG-I is the only epitope labelled throughout the entire thickness of all cell walls in paratracheal parenchyma. Other cell wall epitopes are detected in discrete bands in the thickenings. (B) RG-I backbone epitopes are detected with monoclonal antibody RU-II in, typically, xylem-facing layers. Note that thickenings that stain more weakly with toluidine blue O do not label. (C) De-esterified pectins are detected with LM19 in the non-thickened walls, while within the thickenings they occur in thin layers that stain much more darkly with toluidine blue O. (D) Xyloglucan detection with mAb LM15 is mostly confined to narrow bands within the thickened portions of the cell walls.
DISCUSSION
In this paper we demonstrate for the first time a range of unusual cell wall modifications found in the haustorial hyaline bodies of three rhinanthoid Orobanchaceae species: R. minor, O. vernus and M. pratense. In addition to the paramural and extracellular structures similar to those previously described in A. orobanchoides (Visser et al., 1984), globular bodies of cell wall material (globular ergastic bodies) were observed in the cytoplasm. We present the first immunocytochemical study of the hyaline body cell walls and globular ergastic bodies, showing enrichment in a set of AGP glycan epitopes. These modifications were common in all three species examined and might be a widespread feature essential for the optimal functioning of haustoria within the Orobanchaceae.
The most striking feature observed during this study was the extent of AGP labelling in the hyaline bodies, the epitopes being very strongly detected in the cell walls as well as in the cell contents. AGPs are the most glycosylated of all plant proteins, possessing an arabinose- and galactose-rich carbohydrate moiety of up to 98 % w/w that is attached to a hydroxyproline-rich protein domain (Seifert and Roberts, 2007). While the immense biochemical diversity of AGPs poses many challenges to their full characterization (Tan et al., 2013), they are believed to be involved in many fundamental reproductive and developmental processes in plants (Seifert and Roberts, 2007; Ellis et al., 2010; Nguema-Ona et al., 2012). The general complexity of AGPs, their structural diversity and the variation in the developmental stages at which they were found in this study make interpretation of their functions a challenging task, further complicated by some of the very unusual and intriguing features we observed in haustoria. However, we have included discussion of the possible implications of AGPs for hyaline body function based on current knowledge of these molecules.
The apparent dominance of AGP epitopes over other epitopes of cell wall matrix glycans has not been observed in any other immunohistochemical study of cell walls. In parasitic plant haustoria specifically, AGPs have previously been immunodetected only in the searching cells of Cuscuta pentagona (Vaughn, 2003). Furthermore, the complementary pattern of intense AGP labelling and lack of de-esterified HG detection in the hyaline body appears to have no counterparts in the literature. Surprisingly, no differences in the distribution of esterified and de-esterified HG have been found in two species from the same family, Buchnera hispida and Striga hermonthica (Neumann et al., 1999), even though they possess distinctive hyaline bodies.
Homogalacturonan is secreted into the cell wall in a highly methyl-esterified form and later de-esterified by pectin methylesterases (Jarvis, 1984; Willats et al., 2001; Wolf et al., 2009). Cell walls surrounding young cells are therefore often rich in highly methyl-esterified HG. Since hyaline body cells often retain a ‘young’, protoplasm-dense appearance longer than the cells of the surrounding parenchyma, it is logical to hypothesize that labelling of de-esterified HG is low as a consequence of the tissue being developmentally younger. However, as the reduced labelling is maintained throughout haustorial maturity, other explanations might be more plausible. Considering the apparently high amount of AGP glycan chains, which are known to be bulky (Seifert and Roberts, 2007), de-esterified HG might be masked through interaction with AGP glycan moieties immediately after de-methylation. It has been shown that AGPs form what appear to be covalent linkages with galacturonic acid residues of HG (Immerzeel et al., 2006) and with other pectic components (Tan et al., 2013). More specifically, ester linkages with type II arabinogalactan moieties were suggested, as was the possibility of the two polymers interacting during vesicular transport to the wall. This is interesting considering that the globular ergastic bodies, which contain both HG and AGPs, appear to be membrane-bound, vesicle-like structures. Masking of HG by AGPs is difficult to investigate in situ as no effective methods for the removal of AGPs from sections are currently available. Preliminary treatments of sections with trypsin, which might dislodge the protein domain of AGPs, produced no change in labelling (data not shown). This is not unexpected as the AGP-directed mAbs used detect epitopes present in the glycan chains, which are bulky and possibly intertwined with other cell wall molecules to form a complex network. Precipitation of AGPs from fresh sections with Yariv reagent is a possibility, although the resulting reagent–AGP complexes might continue to obscure pectin. Furthermore, when hypothesizing a masking physical interaction that hampers detection of de-esterified HG in the cell wall, it is difficult to explain detection of de-esterified HG alongside AGPs in the globular ergastic bodies as well as the paramural bodies. Another possibility is that, rather than being masked by AGP glycans, HG is deposited in lower amounts in the AGP-enriched walls and the portion of HG that is present occurs mainly in the esterified form. The resulting overall reduction in HG content is plausible as hyaline body cell walls often stain lighter purple with toluidine blue O than the paramural bodies and cortical cell walls. This indicates that they most likely contain less pectins (O'Brien et al., 1964). A potential link between AGPs and pectins in the hyaline body could be investigated using methods that require small amounts of material, such as epitope detection chromatography (Cornuault et al., 2014), which utilizes antibodies used for in situ detection while eliminating the problems associated with epitope masking (Marcus et al., 2008).
In addition to the tissue-level relationship between de-esterified HG and AGPs, the polarized distribution of AGPs and de-esterified HG in the individual cells of the outermost hyaline body cell layer is very unusual as it was not always paralleled by structural heterogeneity. To the best of our knowledge, polar immunolocalization patterns in cells without polar heterogeneity in cell wall structure have been previously observed only in relation to developmental directionality in embryogenic or differentiating cells (McCabe et al., 1997; Motose et al., 2004). Labelling with JIM8 and JIM15 in pre-division state B cells of carrot in an embryogenic suspension culture demonstrated planar polarity of AGP epitope distribution that was unrelated to differences in cell wall structure but determined cell fate during somatic embryogenesis (McCabe et al., 1997). The labelled and non-labelled ‘halves’ of B cells gave rise to non-embryogenic (state F) and embryogenic (state C) cells, respectively. The other example of polarity related to development comes from differentiating xylem elements of Zinnia elegans, in which, in vitro and in planta, xylogen (belonging to the non-classical AGPs) concentrates in the cell walls at the tips of differentiating tracheary elements (Motose et al., 2004). This polarity mediates inductive interactions between differentiating cells, allowing structural continuity of the xylem strand. In our study, polarity in AGP/de-esterified HG distribution in the outer cells of the hyaline body was not related to proliferation or differentiation. These biochemically distinctive walls were in contact only with each other, and not with the walls of the cortex. Such physical separation of biochemically distinct walls might also mean that the apoplastic system within the hyaline body is functionally isolated from that of the cortex.
Modification of the apoplastic system of the hyaline body is a plausible explanation behind the altered composition, as AGPs are believed to enhance cell wall rheology by increasing pectin porosity (Lamport and Kieliszewski, 2005; Lamport et al., 2006). Furthermore, as the degree of esterification has implications for pectin gelling properties in that de-esterified pectins are very prone to calcium cross-linking and gel formation (Jarvis, 1984), a lower content of de-esterified HG could increase the fluidity of the pectin matrix in the hyaline body cells. The intramolecular Ca2+ binding ability of AGPs might further prevent calcium cross-linking of HG and allow both HG and AGPs in the wall to remain fluid (Lamport and Várnai, 2012). AGPs can also act to loosen the pectin network by disrupting molecule alignment and cross-linking (Lamport et al., 2006). Therefore, our findings suggest that, in comparison with the surrounding tissues, wall permeability might be altered in the hyaline body. If this is the case, it might have important consequences for apoplastic transport and, therefore, nutrient translocation through the haustorium. However, the plasticizing function of AGPs is typically possible at a relatively low AGP:pectin ratio, estimated at 1:50–1:100 (Lamport et al., 2006). This leaves unexplained the apparently extremely high amounts of AGPs in the hyaline body cell walls. Another possibility is that the AGP-rich cell walls and their paramural extensions function as water-holding reservoirs, as previously suggested for very similar structures in the hyaline bodies of T. humifusum (Renaudin and Cheguillaume, 1977). It is possible that the bulky carbohydrate moieties of AGPs co-act with pectins in maintaining optimal hydration levels in the apoplast, contributing to osmotic homeostasis of the entire organ. AGPs have been, indeed, implicated in preventing the negative effects of repeated hydration and rehydration events on cell wall structure in resurrection plants (Moore et al., 2013). Their role in cell wall hydration might therefore be very important.
The role of the hyaline body as a nutrient processing and storage site is highly plausible considering the large amount of cell wall material deposited in the intercellular spaces, paramural bodies and ergastic inclusions. The fact that attachment to incompatible hosts resulted in poorly differentiated hyaline bodies with no such modifications lends further support to this hypothesis. The reason for potential storage of wall materials in the hyaline body is not known. It might be simply that carbohydrates and proteins are deposited when the parasite's demand for carbon is lower than the amounts taken up from the host, or that they form part of a local system of osmotic gradient regulation important in the flow of substances into the parasite (Jiang et al., 2010). Although hemiparasites photosynthesize and access only host xylem, but not phloem, abstraction of xylem-mobile organic carbon from the host has been documented for both O. vernus (Govier et al., 1967) and R. minor (Těšitel et al., 2010b). The latter has been shown to gain as much as 50 % of its carbon from the host (Těšitel et al., 2010b), abstracted mainly as amino acids (Jiang et al., 2010) and very little carbohydrate (Irving and Cameron, 2009). Dominance of amino acids in the abstracted carbon was also the case for Odontites (Govier et al., 1967). If the carbohydrate- and protein-based structures seen in this study are a means of storing host-derived carbon and nitrogen, processing must occur somewhere between abstraction and deposition and this is most likely achieved within the metabolically active hyaline body.
Flow of abstracted nutrients through the hyaline body is further supported by its association with the specialized type of paratracheal parenchyma, which does not occur below the hyaline body. It appears to be of the same structure as that found in Orobanche crenata (Dörr and Kollmann, 1976), Euphrasia cuneata (Fineran, 1987) or Triphysaria versicolor (Heide-Jørgensen, 1995). The main similarities are uniform wall thickening in contact with xylem, as well as organelle-rich cytoplasm. Additionally, we demonstrate that the thickenings are composed of biochemically distinct layers, highlighting their specialized character. A transfer function by forming a pathway of symplastic transport to parasite phloem has been suggested for these cells (Heide-Jørgensen, 1995). In addition to cells similar to those described in the main body of our article, several more elaborate forms of cell wall modifications suggestive of transfer functions are described and discussed in Supplementary Data Fig. S2. We therefore support the hypothesis that paratracheal parenchyma might participate in the unloading of xylem-mobile solutes into the hyaline body and their transport into the parasite's root phloem following processing and transient storage in the hyaline body.
Typical storage materials in plants are protein (Shewry, 1995), starch (Evert, 2006) and cell wall carbohydrates, including xyloglucan and mannan (Buckeridge, 2010). Furthermore, arabinogalactans, interpreted as pectic, have been reported from lupin cotyledons (Buckeridge, 2010) and abundant AGPs were found in mucilage ducts of corms of Colocasia esculenta (Jiang and Ramsden, 1999). It is therefore possible that AGPs have been previously overlooked as a class of storage molecules, and they might be an important element of nutrient metabolism in hyaline bodies. The storage function of the structures found in the hyaline bodies is supported by their large size and abundance, which in the case of R. minor is particularly pronounced in haustoria attached to the nutritious legume host V. sepium. The osmiophilic bodies often seen in association with globular ergastic inclusions (Fig. 6B) resemble precursors associated with protein storage vacuoles (Herman and Larkins, 1999). The size and quantities of globular ergastic bodies imply functions other than simply vesicular transport to the wall, in which they might, however, participate to some extent. To elucidate whether they represent early transport forms of paramural bodies, the composition and structure of both needs to be investigated in a higher-resolution developmental context. High-pressure freezing could prove useful in investigating the ontogeny of these structures, in particular their relationship with the endomembrane system.
To investigate the possibility of host solute incorporation into hyaline body structures more directly, carefully designed tracer and radioactive marker experiments could prove useful. To visualize incorporation suggestive of AGPs, either 14C- or 3H-arabinose could be introduced into host xylem and detected either via scintillation counting or autoradiography (against 2,5-diphenyloxazole-pretreated X-ray film in the case of tritiated arabinose). However, as the radioactivity will be incorporated into pentose residues (arabinase, xylose and 2-O-methyl xylose), other wall components, such as xyloglucan, would also be labelled, necessitating extraction and more detailed characterization of the radiolabelled wall components (Fry, 2001). Another approach would be to label the smaller protein moiety. However, in this case other proteins would be labelled, requiring further determination of the labelled compounds. Furthermore, identification of relevant molecular aspects of hyaline body biology could be achieved through transcriptomics using tissue fragments isolated with laser microdissection tools.
The architectural specialization of hyaline body cell walls, i.e. their distinctive composition and structure, is associated with cytological evidence of increased metabolic activity. It begins early during the differentiation of the hyaline body and is maintained throughout its maturity. This suggests that the hyaline body might aid the establishment of an interaction with a host as well as being a crucial element of its subsequent functioning. Future studies should focus on distinguishing between the cytological activity related to the formation of cell wall modifications and globular ergastic bodies and other functions of the hyaline body. Similarly, it will be important to distinguish between the developmental aspects of the presence of AGP in the hyaline body and the roles of these molecules in established haustoria. While the epitope structures detected and generally high abundance of AGPs were very consistent in this study, deviations from these patterns and variations in epitope proportions should be further researched across a wider taxonomic range and under various nutritional and environmental conditions.
Closing remarks
This is the first study that highlights the unusual character of hyaline body cell walls using immunocytochemical detection. The striking dominance of AGP epitopes suggests that they are central to the functioning and/or development of the hyaline body, most likely by regulating the rheological properties of the apoplast and contributing to nutrient metabolism. The unusual aspects of hyaline body cell wall architecture and intracellular cell wall materials identified in this study will contribute to the formation of further hypotheses about the purpose of this tissue, eventually tested by functional approaches.
SUPPLEMENTARY DATA
ACKNOWLEDGEMENTS
We thank the Centre for Microscopy and Imaging at NUI Galway for access to microscopy facilities and we are particularly grateful to Mr Pierce Lalor for his technical support. We are grateful to Michael G. Hahn (Complex Carbohydrate Research Center, University of Athens, Georgia, USA) and Marie-Christine Ralet (INRA, Nantes, France) for the kind provision of the mAbs CCRC-M1 and CCRC-M7 and INRA-RU-II, respectively. Research presented in this article was funded by The Irish Research Council (EMBARK. RS/2009/1339) and the NUI Galway Thomas Crawford-Hayes Fund.
LITERATURE CITED
- Ameloot E. Impact of hemiparasitic Rhinanthus spp. on vegetation structure and dynamics of semi-natural grassland. Belgium: Katholieke Universiteit Leuven; 2007. PhD Thesis. [Google Scholar]
- Ameloot E, Verheyen K, Hermy M. Meta-analysis of standing crop reduction by Rhinanthus spp. and its effect on vegetation structure. Folia Geobotanica. 2005;40:289–310. [Google Scholar]
- Arnaud M-C, Véronési C, Thalouarn P. Physiology and resistance to Striga hermonthica in Sorghum bicolor var. Framida. Australian Journal of Plant Physiology. 1999;26:63–70. [Google Scholar]
- Ba AT. Evidence of enzyme activities in the haustorium of Striga gesneroides (Scrophulariaceae) In: Ramaiah KV, Vasudeva Rao MJ, editors. Proceedings of the Second International Workshop on Striga. Ouagadougou, Upper Volta: ICRISAT; 1983. pp. 39–42. [Google Scholar]
- Brokamp G, Dostert N, Cáceres-H F, Weigend M. Parasitism and haustorium anatomy of Krameria lappacea (Dombey) Burdet & B.B. Simpson (Krameriaceae), an endangered medicinal plant from the Andean deserts. Journal of Arid Environments. 2012;83:94–100. [Google Scholar]
- Buckeridge MS. Seed cell wall storage polysaccharides: models to understand cell wall biosynthesis and degradation. Plant Physiology. 2010;154:1017–1023. doi: 10.1104/pp.110.158642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bullock JM, Pywell RF. Rhinanthus: a tool for restoring diverse grassland? Folia Geobotanica. 2005;40:273–288. [Google Scholar]
- Cameron DD, Seel WE. Functional anatomy of haustoria formed by Rhinanthus minor: linking evidence from histology and isotope tracing. New Phytologist. 2007;174:412–419. doi: 10.1111/j.1469-8137.2007.02013.x. [DOI] [PubMed] [Google Scholar]
- Cameron DD, Coats AM, Seel WE. Differential resistance among host and non-host species underlies the variable success of the hemi-parasitic plant Rhinanthus minor. Annals of Botany. 2006;98:1289–1299. doi: 10.1093/aob/mcl218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornuault V, Manfield IW, Ralet MC, Knox JP. Epitope detection chromatography (EDC): a method to dissect the structural heterogeneity and interconnections of plant cell wall matrix glycans. Plant Journal. 2014;78:715–722. doi: 10.1111/tpj.12504. [DOI] [PubMed] [Google Scholar]
- Davies D. The impact of Rhinanthus spp. on sward productivity and composition: implications for the restoration of species-rich grasslands. Biological Conservation. 1997;82:87–93. [Google Scholar]
- Dobbins DR, Kuijt J. Studies on the haustorium of Castilleja (Scrophulariaceae). I. The upper haustorium. Canadian Journal of Botany. 1973;51:917–922. [Google Scholar]
- Dörr I, Kollmann R. Strukturelle Grundlagen des Parasitismus bei Orobanche. III. Die Differenzierung des Xylemanschlusses bei O. crenata. Protoplasma. 1976;89:235–249. [Google Scholar]
- Ellis M, Egelund J, Schultz CJ, Bacic A. Arabinogalactan-proteins: key regulators at the cell surface? Plant Physiology. 2010;153:403–419. doi: 10.1104/pp.110.156000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evert RF. Hoboken, NJ: John Wiley & Sons; 2006. Esau's Plant anatomy. [Google Scholar]
- Fineran BA. A structural approach towards investigating transport systems between host and parasite, as exemplified by some mistletoes and root parasites. In: Weber HC, Forstreuter W, editors. Proceedings of the 4th International Symposium on Parasitic Flowering Plants, Marburg 1987. Marburg: Philipps-Universität; 1987. pp. 201–220. [Google Scholar]
- Fry SC. The growing plant cell wall: chemical and metabolic analysis. Reprint Edition. The Caldwell, NJ: Blackburn Press; 2001. [Google Scholar]
- Gibson C, Watkinson A. Host selectivity and the mediation of competition by the root hemiparasite Rhinanthus minor. Oecologia. 1991;86:81–87. doi: 10.1007/BF00317393. [DOI] [PubMed] [Google Scholar]
- Gibson C, Watkinson A. The role of the hemiparasitic annual Rhinanthus minor in determining grassland community structure. Oecologia. 1992;89:62–68. doi: 10.1007/BF00319016. [DOI] [PubMed] [Google Scholar]
- Govier R, Nelson MD, Pate J. Hemiparasitic nutrition in angiosperms. I. The transfer of organic compounds from host to Odontites verna (Bell.) Dum. (Scrophulariaceae) New Phytologist. 1967;66:285–297. [Google Scholar]
- Gurney A, Grimanelli D, Kanampiu F, Hoisington D, Scholes J, Press M. Can wild relatives of cereals provide new sources of resistance to the parasitic angiosperm Striga hermonthica?; Proceedings of the Seventh International Parasitic Weed Symposium, Nantes, France, 4–8 June 2001; Nantes: Université de Nantes; 2001. p. 247. [Google Scholar]
- Gurney AL, Grimanelli D, Kanampiu F, Hoisington D, Scholes JD, Press MC. Novel sources of resistance to Striga hermonthica in Tripsacum dactyloides, a wild relative of maize. New Phytologist. 2003;160:557–568. doi: 10.1046/j.1469-8137.2003.00904.x. [DOI] [PubMed] [Google Scholar]
- Gurney AL, Slate J, Press MC, Scholes JD. A novel form of resistance in rice to the angiosperm parasite Striga hermonthica. New Phytologist. 2006;169:199–208. doi: 10.1111/j.1469-8137.2005.01560.x. [DOI] [PubMed] [Google Scholar]
- Heide-Jørgensen H. The haustorium of the root parasite Triphysaria (Scrophulariaceae), with special reference to xylem bridge ultrastructure. American Journal of Botany. 1995;82:782–797. [Google Scholar]
- Heide-Jørgensen HS. Parasitic flowering plants. Leiden: Brill; 2008. [Google Scholar]
- Herman EM, Larkins BA. Protein storage bodies and vacuoles. Plant Cell. 1999;11:601–613. doi: 10.1105/tpc.11.4.601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hervé C, Marcus SE, Knox JP. Monoclonal antibodies, carbohydrate-binding molecules, and the detection of polysaccharides in plant cell walls. In: Popper ZA, editor. The plant cell wall. Methods and protocols. New York: Humana Press; 2011. pp. 103–113. [DOI] [PubMed] [Google Scholar]
- Immerzeel P, Eppink MM, de Vries SC, Schols HA, Voragen AGJ. Carrot arabinogalactan proteins are interlinked with pectins. Physiologia Plantarum. 2006;128:18–28. [Google Scholar]
- Irving LJ, Cameron DD. You are what you eat: interactions between root parasitic plants and their hosts. Advances in Botanical Research. 2009;50:87–138. [Google Scholar]
- Jarvis MC. Structure and properties of pectin gels in plant cell walls. Plant, Cell and Environment. 1984;7:153–164. [Google Scholar]
- Jauh GY, Lord EM. Localization of pectins and arabinogalactan-proteins in lily (Lilium longiflorum L.) pollen tube and style, and their possible roles in pollination. Planta. 1996;199:251–261. [Google Scholar]
- Jiang F, Jeschke WD, Hartung W. Abscisic acid (ABA) flows from Hordeum vulgare to the hemiparasite Rhinanthus minor and the influence of infection on host and parasite abscisic acid relations. Journal of Experimental Botany. 2004;55:2323–2329. doi: 10.1093/jxb/erh240. [DOI] [PubMed] [Google Scholar]
- Jiang F, Jeschke WD, Hartung W, Cameron DD. Interactions between Rhinanthus minor and its hosts: a review of water, mineral nutrient and hormone flows and exchanges in the hemiparasitic association. Folia Geobotanica. 2010;45:369–385. [Google Scholar]
- Jiang G, Ramsden L. Characterisation and yield of the arabinogalactan-protein mucilage of taro corms. Journal of the Science of Food and Agriculture. 1999;79:671–674. [Google Scholar]
- Kuijt J. The biology of parasitic flowering plants. Berkeley: University of California Press; 1969. [Google Scholar]
- Lamport D, Kieliszewski M. Stress upregulates periplasmic arabinogalactan-proteins. Plant Biosystems. 2005;139:60–64. doi: 10.1111/j.1469-8137.2005.01591.x. [DOI] [PubMed] [Google Scholar]
- Lamport DTA, Várnai P. Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytologist. 2012;197:58–64. doi: 10.1111/nph.12005. [DOI] [PubMed] [Google Scholar]
- Lamport DTA, Kieliszewski MJ, Showalter AM. Salt stress upregulates periplasmic arabinogalactan proteins: using salt stress to analyse AGP function. New Phytologist. 2006;169:479–492. doi: 10.1111/j.1469-8137.2005.01591.x. [DOI] [PubMed] [Google Scholar]
- Mallaburn PS, Stewart GR. Haustorial function in Striga: comparative anatomy of S. asiatica (L.) Kuntz and S. hermonthica (Del.) Benth. (Scrophulariaceae) In: Weber HC, Forstreuter W, editors. Proceedings of the 4th International Symposium on Parasitic Flowering Plants, Marburg 1987. Marburg: Philipps-Universität; 1987. pp. 523–536. [Google Scholar]
- Marcus SE, Verhertbruggen Y, Hervé C, et al. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biology. 2008;8:60–72. doi: 10.1186/1471-2229-8-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCabe PF, Valentine TA, Forsberg LS, Pennell RI. Soluble signals from cells identified at the cell wall establish a developmental pathway in carrot. Plant Cell. 1997;9:2225–2241. doi: 10.1105/tpc.9.12.2225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNeal JR, Bennett JR, Wolfe AD, Mathews S. Phylogeny and origins of holoparasitism in Orobanchaceae. American Journal of Botany. 2013;100:971–983. doi: 10.3732/ajb.1200448. [DOI] [PubMed] [Google Scholar]
- Moore JP, Nguema-Ona EE, Vicré-Gibouin M, et al. Arabinose-rich polymers as an evolutionary strategy to plasticize resurrection plant cell walls against desiccation. Planta. 2013;237:739–54. doi: 10.1007/s00425-012-1785-9. [DOI] [PubMed] [Google Scholar]
- Motose H, Sugiyama M, Fukuda H. A proteoglycan mediates inductive interaction during plant vascular development. Nature. 2004;429:873–878. doi: 10.1038/nature02613. [DOI] [PubMed] [Google Scholar]
- Musselman LJ, Dickison WC. The structure and development of the haustorium in parasitic Scrophulariaceae. Botanical Journal of the Linnean Society. 1975;70:183–212. [Google Scholar]
- Ndambi B, Cadisch G, Elzein A, Heller A. Colonization and control of Striga hermonthica by Fusarium oxysporum f. sp. strigae, a mycoherbicide component: an anatomical study. Biological Control. 2011;58:149–159. [Google Scholar]
- Neumann U, Vian B, Weber HC, Sallé G. Interface between haustoria of parasitic members of the Scrophulariaceae and their hosts: a histochemical and immunocytochemical approach. Protoplasma. 1999;207:84–97. [Google Scholar]
- Nguema-Ona E, Coimbra S, Vicré-Gibouin M, Mollet J-C, Driouich A. Arabinogalactan proteins in root and pollen-tube cells: distribution and functional aspects. Annals of Botany. 2012;110:383–404. doi: 10.1093/aob/mcs143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nwoke FIO. The initiation of the secondary haustorium in Alectra vogelii Benth. Annals of Botany. 1982;49:669–676. [Google Scholar]
- O'Brien TP, Feder N, McCully ME. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma. 1964;59:368–373. [Google Scholar]
- Parker C. Observations on the current status of Orobanche and Striga problems worldwide. Pest Management Science. 2009;65:453–459. doi: 10.1002/ps.1713. [DOI] [PubMed] [Google Scholar]
- Parker C. Parasitic weeds: a world challenge. Weed Science. 2012;60:269–276. [Google Scholar]
- Press MC, Phoenix GK. Impact of parasitic plants on natural communities. New Phytologist. 2005;166:737–751. doi: 10.1111/j.1469-8137.2005.01358.x. [DOI] [PubMed] [Google Scholar]
- Pywell RF, Bullock JM, Walker KJ, Coulson SJ, Gregory SJ, Stevenson MJ. Facilitating grassland diversification using the hemiparasitic plant Rhinanthus minor. Journal of Applied Ecology. 2004;41:880–887. [Google Scholar]
- Renaudin S, Cheguillaume L. Sur quelques aspects de l'ultrastructure des suçoirs de Thesium humifusum L. Protoplasma. 1977;69:55–69. [Google Scholar]
- Riopel JL, Timko MP. Haustorial initiation and differentiation. In: Press MC, Graves JD, editors. Parasitic plants. London: Chapman and Hall; 1995. pp. 39–79. [Google Scholar]
- Rümer S, Cameron D, Wacker R, Hartung W, Jiang F. An anatomical study of the haustoria of Rhinanthus minor attached to roots of different hosts. Flora. 2007;202:194–200. [Google Scholar]
- Runo S, Macharia S, Alakonya A, Machuka J, Sinha N, Scholes J. Striga parasitizes transgenic hairy roots of Zea mays and provides a tool for studying plant-plant interactions. Plant Methods. 2012;8:20. doi: 10.1186/1746-4811-8-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seifert GJ, Roberts K. The biology of arabinogalactan proteins. Annual Review of Plant Biology. 2007;58:137–161. doi: 10.1146/annurev.arplant.58.032806.103801. [DOI] [PubMed] [Google Scholar]
- Shewry PR. Plant storage proteins. Biological reviews of the Cambridge Philosophical Society. 1995;70:375–426. doi: 10.1111/j.1469-185x.1995.tb01195.x. [DOI] [PubMed] [Google Scholar]
- Suetsugu K, Kawakita A, Kato M. Host range and selectivity of the hemiparasitic plant Thesium chinense (Santalaceae) Annals of Botany. 2008;102:49–55. doi: 10.1093/aob/mcn065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan L, Eberhard S, Pattathil S, et al. An Arabidopsis cell wall proteoglycan consists of pectin and arabinoxylan covalently linked to an arabinogalactan protein. Plant Cell. 2013;25:270–287. doi: 10.1105/tpc.112.107334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tennakoon KU, Cameron DD. The anatomy of Santalum album (sandalwood) haustoria. Canadian Journal of Botany. 2006;84:1608–1616. [Google Scholar]
- Těšitel J. Biology of the root-parasitic rhinanthoid Orobanchaceae. Czech Republic: University of South Bohemia in Českié Budějovice; 2011. PhD thesis. [Google Scholar]
- Těšitel J, Říha P, Svobodová Š, Malinová T, Štech M. Phylogeny, life history evolution and biogeography of the rhinanthoid Orobanchaceae. Folia Geobotanica. 2010a;45:347–367. [Google Scholar]
- Těšitel J, Plavcová L, Cameron DD. Heterotrophic carbon gain by the root hemiparasites, Rhinanthus minor and Euphrasia rostkoviana (Orobanchaceae) Planta. 2010b;231:1137–44. doi: 10.1007/s00425-010-1114-0. [DOI] [PubMed] [Google Scholar]
- Vaughn KC. Dodder hyphae invade the host: a structural and immunocytochemical characterization. Protoplasma. 2003;220:189–200. doi: 10.1007/s00709-002-0038-3. [DOI] [PubMed] [Google Scholar]
- Visser JH, Dörr I, Kollmann R. The ‘hyaline body’ of the root parasite Alectra orobanchoides Benth. (Scrophulariaceae)—its anatomy, ultrastructure and histochemistry. Protoplasma. 1984;121:146–156. [Google Scholar]
- Westbury DB, Davies A, Woodcock BA, Dunnett NP. Seeds of change: the value of using Rhinanthus minor in grassland restoration. Journal of Vegetation Science. 2006;17:435–446. [Google Scholar]
- Willats WG, Orfila C, Limberg G, et al. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls. Implications for pectin methyl esterase action, matrix properties, and cell adhesion. Journal of Biological Chemistry. 2001;276:19404–19413. doi: 10.1074/jbc.M011242200. [DOI] [PubMed] [Google Scholar]
- Wolf S, Mouille G, Pelloux J. Homogalacturonan methyl-esterification and plant development. Molecular Plant. 2009;2:851–860. doi: 10.1093/mp/ssp066. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.