

Abstract
The coordination between nuclear and organellar genes is essential to many aspects of eukaryotic life, including basic metabolism, energy production, and ultimately, organismal fitness. Although nuclear genes are biparentally inherited, mitochondrial and chloroplast genes are almost exclusively maternally inherited, and this asymmetry may lead to a bias in the chromosomal distribution of nuclear genes whose products act in the mitochondria or chloroplasts. In particular, because X-linked genes have a higher probability of cotransmission with organellar genes (2/3) compared with autosomal genes (1/2), selection for coadaptation has been predicted to lead to an overrepresentation of nuclear-mitochondrial and nuclear-chloroplast genes on the X chromosome relative to autosomes. In contrast, the occurrence of sexually antagonistic organellar mutations might lead to selection for movement of cytonuclear genes from the X chromosome to autosomes to reduce male mutation load. Recent broad-scale comparative studies of N-mt distributions in animals have found evidence for these hypotheses in some species, but not others. Here, we use transcriptome sequences to conduct the first study of the chromosomal distribution of cytonuclear interacting genes in a plant species with sex chromosomes (Rumex hastatulus; Polygonaceae). We found no evidence of under- or overrepresentation of either N-mt or N-cp genes on the X chromosome, and thus no support for either the coadaptation or the sexual-conflict hypothesis. We discuss how our results from a species with recently evolved sex chromosomes fit into an emerging picture of the evolutionary forces governing the chromosomal distribution of nuclear-mitochondrial and nuclear-chloroplast genes.
Keywords: cytonuclear interactions, coadaptation, sexual conflict, gene transfer
Introduction
The intimate relationships between nuclear and organellar genomes in eukaryotes represent some of the most striking examples of coevolved mutualisms (Gillham 1994; Lane 2005; Aanen et al. 2014). The long coevolutionary history of nuclear and mitochondrial genomes is perhaps best illustrated by the finding that the vast majority of mitochondrial genes in animals have been transferred to the nuclear genome (Adams and Palmer 2003; Rand et al. 2004; Burt and Trivers 2006). Indeed, animal mitochondria now encode only a few proteins after having lost the majority of their original genes (Berg and Kurland 2000; Ridley 2000; Bar-Yaacov et al. 2012). Moreover, almost one-fifth of the Arabidopsis thaliana nuclear genome is of chloroplast origin (Martin 2003), suggesting that organellar-to-nuclear gene movement has played a crucial role in the evolution of plant genetic systems.
The evolution of cytonuclear interactions and the chromosomal distribution of the genes involved should be influenced by the contrasting modes of inheritance of organellar genes (maternal inheritance) and autosomal genes (biparental inheritance). This difference may, for example, result in conflict between nuclear and organellar genes over sex determination and sex ratio (Cosmides and Tooby 1981; Werren and Beukeboom 1998), and several mitochondrial genes in plants are known to cause male sterility (Burt and Trivers 2006; Touzet and Meyer 2014). In systems with XY sex determination, where males are the heterogametic (XY) and females the homogametic sex (XX), genes on the X-chromosome spend two-thirds of their time in females (Rand et al. 2001) and therefore share a female-biased inheritance pattern relative to Y-linked or autosomal genes, which may result in intergenomic coadaptation or conflict.
A potential consequence of intergenomic conflict or coadaptation between nuclear genes, whose products interact with mitochondrial or chloroplasts (mitonuclear and cytonuclear genes, respectively) and other regions of the genome, is a shift in the chromosomal location of such genes, either becoming more or less abundant on the X chromosome. Several molecular mechanisms have been suggested to be involved in driving gene movement, including gene duplication followed by fixation and subsequent gene loss (Wu and Yujun Xu 2003), and autosomal gene duplications followed by the evolution of sex-biased gene expression (Connallon and Clark 2011). The evolutionary mechanisms of this gene movement have also been explored by several recent studies (Drown et al. 2012; Hill and Johnson 2013; Dean et al. 2014; Rogell et al. 2014), and two main processes have been proposed to account for the movement of genes to or from the X chromosome. The coadaptation hypothesis predicts that the cotransmission of X-linked and organellar genes should result in their coadaptation, in which selection on beneficial epistatic interactions results in an overrepresentation of cytonuclear genes on the X chromosome relative to autosomes (Rand et al. 2004; Drown et al. 2012). In contrast, the sexual conflict hypothesis predicts the opposite chromosomal distribution, with more cytonuclear genes occurring on autosomes to alleviate mutation load in males. To date, empirical evidence for these hypotheses are mixed. Drown et al. (2012) used previously published reference genomes to examine the chromosomal distribution of N-mt genes in 16 vertebrates and found a strong underrepresentation of such genes on the X chromosomes relative to autosomes in 14 mammal species, but not in two avian species with ZW sex determining systems; note that the coadaptation hypothesis does not predict that ZW systems should show a bias in the distribution of cytonuclear genes. Dean et al. (2014) included seven additional species in their analysis with independently derived sex chromosomes and found that the underrepresentation of N-mt genes on the X chromosome was restricted to therian mammals and Caenorhabditis elegans.
Here, we use sex-linked and autosomal transcriptome sequences to investigate the chromosomal distributions of cytonuclear interactions in the dioecious annual plant Rumex hastatulus (Polygonaceae). Examining cytonuclear interactions within a plant species is of interest for several reasons (see Sloan 2014). First, plants carry an additional maternally inherited organellar genome that is absent in animals, the chloroplast genome. This provides an opportunity to compare the chromosomal distribution of two independent kinds of cytonuclear interacting genes: Nuclear-mitochondrial and nuclear-chloroplast. Second, whereas animal sex chromosomes evolved hundreds of millions of years ago (180 Ma in mammals and 140 Ma in birds; Cortez et al. 2014), the origin of plant sex chromosomes is a more recent event (Charlesworth 2013). In R. hastatulus, sex chromosomes are thought to have evolved approximately 15–16 Ma (Navajas-Perez et al. 2005) and genes on the Y chromosome show evidence of degeneration, resulting in a considerable proportion of genes that are hemizygous on the X chromosome (Hough et al. 2014). Rumex hastatulus therefore provides an opportunity to test whether the early changes involved in sex chromosome evolution are associated with a concomitant shift in the chromosomal location of N-mt or N-cp genes. Moreover, the presence in this system of X-linked genes that have recently become hemizygous provides an opportunity to compare the chromosomal distributions of X-linked genes that are hemizygous versus those that have retained Y-linked alleles (X/Y genes). Hemizygous genes are particularly good candidates for evaluating evidence for coadaptation and/or sexual conflict because of their relatively older age (Hough et al. 2014), and because beneficial mutations in such genes are exposed to positive selection regardless of dominance and may therefore spread more rapidly.
Results and Discussion
It has been suggested that cytonuclear genes may be either over or underrepresented on the X chromosome compared with autosomes, depending on whether their interactions are driven by coadaptation or sexual conflict (Rand et al. 2001; Drown et al. 2012; Hill and Johnson 2013; Dean et al. 2014; Rogell et al. 2014). We annotated sex-linked and autosomal transcriptome sequences to test these predictions in the dioecious plant R. hastatulus. We found that neither mitochondria- or chloroplast-interacting nuclear genes were under or overrepresented on the X chromosome (Fisher’s exact test, P = 0.4947 and P = 0.3074, respectively; fig. 1). This pattern indicates that neither the coadaptation nor the sexual conflict hypothesis alone is sufficient to explain the chromosomal distribution of cytonuclear genes in R. hastatulus.
Fig. 1.—
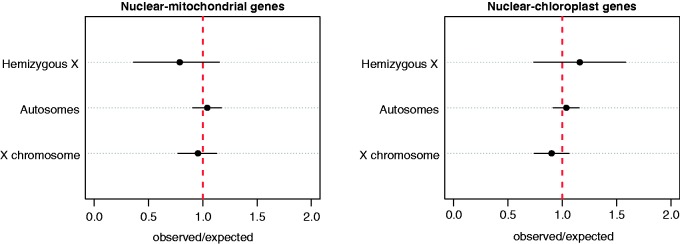
Representation of the chromosomal location of cytonuclear genes in R. hastatulus. Dots represent the observed to expected ratio of mitonuclear (N-mt) and chloronuclear (N-cp) genes on autosomes, the X chromosome, and hemizygous X-genes, with the 95% confidence intervals estimated by bootstrapping (10,000 replicates). The vertical dotted line at one represents no over or underrepresentation.
There are several factors that are expected to be important in determining cytonuclear gene distributions, and these may explain the lack of bias in R. hastatulus. For example, under both the coadaptation and sexual conflict hypotheses, the age of the sex chromosomes will determine the extent to which selection (either for coadaptation, or sexual antagonism) has had time to operate, which depends on the rate of gene movement onto and off of the sex chromosomes. Although previous studies of cytonuclear genes in animals have focused almost exclusively on ancient sex chromosome systems (Drown et al. 2012; Dean et al. 2014; Rogell et al. 2014), our study focused on a dioecious plant species in which sex chromosomes evolved more recently (∼15 Ma; Navajas-Perez et al. 2005), and many genes likely stopped recombining much more recently (Hough et al. 2014). The lack of bias in the chromosomal distribution of cytonuclear genes may therefore reflect the recent timescale of sex chromosome evolution rather than the absence of biased gene movement. The relatively young age of sex chromosomes may also have played a role in the lack of bias reported in the sex and neo-sex chromosomes in three-spined stickleback, which evolved approximately 10 Ma (Kondo et al. 2004) and approximately 2 Ma, respectively (Natri et al. 2013). Comparative studies of sex chromosomes of different age will be central for understanding the rate at which organellar gene movement occurs.
In addition to being evolutionarily older, X-linked hemizygous genes are expected to show a greater effect of over or underrepresentation than genes with both X- and Y-alleles because recessive mutations (involved in either coadaptation or sexual conflict) will be exposed to selection instead of masked by an alternate allele in a heterozygous genotype. We detected a slightly greater underrepresentation of X-hemizygous N-mt genes compared with autosomes or X-genes with retained Y-alleles, but the effect was not statistically significant (P = 0.4947). The opposite pattern was evident for N-cp genes, which were slightly overrepresented on hemizygous genes, but again this effect was not significant (P = 0.3074). A larger sample of hemizygous genes would be required to more confidently assess whether such genes are in fact more often involved in cytonuclear interactions than other genes on the X chromosome, and to test whether the opposite pattern for N-mt and N-cp hemizygous genes is a result of a different rate of nuclear gene transfer between mitochondrial and chloroplast genomes. In particular, the smaller number of hemizygous X-linked genes in our data set implies that power was reduced for this comparison, such that a 5% difference could only be detected with approximately 60% power (see supplementary material S1, Supplementary Material online).
Another factor that will affect the chromosomal distribution of cytonuclear genes is the number of N-mt and N-cp genes that were located on the autosome from which the sex chromosomes evolved. Because the origins of mitochondria and chloroplasts both vastly predate that of sex chromosomes (1.5–2 Ba compared with <200 Ma; Dyall et al. 2004; Timmis et al. 2004; Cortez et al. 2014), gene transfer from organellar genomes to the nuclear genome began long before the evolution of sex chromosomes. A bias in the chromosomal distribution of cytonuclear genes in either direction may therefore arise if the ancestral autosome was particularly rich or poor in cytonuclear genes. Indeed, it is striking that autosomes in the animal species previously examined exhibited extensive variation in the relative number of N-mt genes (see Drown et al. 2012; Dean et al. 2014). That the ancestral number of N-mt and N-cp genes is likely to be important is highlighted by the fact that the majority of genes involved in mitochondrial DNA and RNA metabolism in A. thaliana are found on chromosome 3 (Elo et al. 2003). If such a biased autosomal distribution of organellar variation is representative of the ancestral sex chromosomes, the X chromosome could carry significantly more N-mt or N-cp genes because of this ancestral gene number rather than a biased rate of gene movement. This effect is likely exacerbated in early sex chromosome systems, where the majority of genes may not have experienced opportunities for movement. Genetic mapping and comparative genomic studies of genes that have transferred from organellar genomes after the origin of sex chromosomes may provide a means to control for ancestral differences in gene number and provide a better test of biases in organellar-nuclear gene movement.
To conclude, our study is the first investigation of the extent to which coadaptation and sexual conflict may have shaped the chromosomal distribution of cytonuclear genes in a plant species with sex chromosomes. We found no sign of under or overrepresentation of either N-mt or N-cp genes on the X chromosome, implying that neither coadaptation nor sexual conflict alone can explain the chromosomal distributions of these genes. Instead, we suggest that additional factors, including the age of sex chromosomes and the time that has elapsed since X–Y recombination became suppressed, are likely to have been important determinants of the patterns we observed. To determine whether the lack of underrepresentation of mitonuclear genes on the X chromosome reflects an absence of gene movement, future studies should focus on quantifying rates of gene movement after sex chromosome origination, and consider the extent to which neutral processes including the number of mitonuclear genes on ancestral sex chromosomes have played an important role in shaping the current chromosomal distributions of such genes. Cytonuclear conflict and coevolution have undoubtedly played a major role in many aspects of genome evolution in both plant and animal systems, and the previously reported evidence from therian mammals and C. elegans (Drown et al. 2012; Dean et al. 2014) suggests that sexual conflict and coadaptation might represent important mechanisms driving chromosomal gene movement; however, it remains unclear whether these processes have also shaped the chromosomal distribution of cytonuclear genes in plants.
Materials and Methods
Gene Identification and Functional Annotation
We used sex-linked and autosomal transcriptome sequence data for R. hastatulus reported in Hough et al. (2014; GenBank Sequence Read Archive accession no. SRP041588), and obtained three sets of genes with which to test for an over or underrepresentation of nuclear-mitochondrial- or nuclear-chloroplast genes. In total our analyses included 1,167 autosomal genes, 624 X-linked genes, and 107 hemizygous X-linked genes. The X-linked and hemizygous X-linked genes were shared between sex chromosome systems in this species (see Materials and Methods and SI Appendix of Hough et al. 2014 for full details regarding the identification of such genes from transcriptome sequence data). For autosomal genes, we included those previously identified as confidently autosomal in both R. hastatulus sex chromosome systems, as well as those uniquely identified in the XYY system. For each gene set, we queried the sequences translated in all reading frames against the A. thaliana protein database using the BLASTx homology search implemented in Blast2GO (Conesa et al. 2005), with a significance threshold (BLAST ExpectValue) of 1 × 10-3, above which matches were not reported. We limited our searches to the A. thaliana protein database because sequence matches to this database returned more detailed functional information than is available for most other species in the NCBI plant database. We obtained BLASTx results for 1,073 autosomal genes (90%), 567 X-linked genes (90%), and 95 hemizygous genes (89%). Gene ontology (GO) terms associated with the hits from BLASTx queries were then retrieved using the “Mapping” function in Blast2GO, which used BLAST accessions to link the queried sequences to functional information stored in the GO database (Gene Ontology Consortium 2008). Gene names were retrieved using NCBI mapping files “gene info” and “gene2accession,” and GO terms were assigned to query sequences using the “Annotation” function with an E Value-Hit-Filter of 1 × 10-6 and an annotation cut off of 55 (default parameters). Finally, we ran InterProScan (Quevillon et al. 2005) to retrieve sequence domain/motif information and merged the corresponding annotations with previously identified GO terms. This procedure generated output files containing GO ID’s and functional descriptions for each gene in our data set (files uploaded to GitHub). The numbers of genes in our final data set with functional annotations and N-mt and N-cp GO annotations are summarized in table 1.
Table 1.
Number of Genes in Our Data Set
| Gene Sets | Autosomal | X-Linked | X-Hemizygous |
|---|---|---|---|
| Original data set | 1,167 | 624 | 107 |
| With annotation | 1,073 | 567 | 95 |
| With N-mt GO ID | 194 | 94 | 13 |
| With N-cp GO ID | 222 | 102 | 22 |
Note.—Includes genes for which we obtained functional annotations (see Materials and Methods) and those with nuclear-mitochondrial and nuclear-chloroplast GO annotations.
Statistical Analyses
We used a similar approach to Drown et al. (2012) and Dean et al. (2014) and estimated the number of N-mt and N-cp genes on the X chromosome and autosomes, and then compared each of these estimates to an expected number. The expected number of N-mt genes was obtained by calculating the product of the proportion of all genes in the data set with mitochondrial annotations (matching GO:0005739) and the number of annotated genes in a given gene set. The expected numbers of N-cp genes were calculated similarly, using GO:0009507. We then calculated the ratios of the observed-to-expected numbers for both N-mt and N-cp genes in each gene set. The observed-to-expected ratio is expected to equal one when there is no under or overrepresentation, and greater than one when there is an overrepresentation. We note that, unlike for X-linked genes, we did not have information regarding the particular chromosome locations for autosomal genes, and therefore could not obtain the expected numbers of N-mt and N-cp genes per autosome as in previous studies (Drown et al. 2012; Dean et al. 2014). The expected numbers were thus calculated assuming that the set of autosomal genes represented a random sample of the autosomal chromosomes in this species, which is likely a valid assumption given that the sequences were obtained using whole transcriptome shotgun sequencing (Hough et al. 2014). Calculating the expected-to-observed ratios across X-linked, autosomal, and X-hemizygous genes thus allowed us to determine whether any of these gene sets contained an under or overrepresentation of N-mt and N-cp genes compared with the expectation based on the proportion of such genes in the full data set. We tested the significance of over or underrepresentation using Fisher’s exact tests, and calculated 95% confidence intervals for the numbers of N-mt or N-cp genes using 10,000 replicate bootstrapped samples. Given our sample sizes of genes with annotations (table 1), Fisher’s Exact Tests allowed us to test for differences in the proportions of cytonuclear genes on autosomes versus the X chromosome that were on the order of 5% with approximately 80% power, whereas power was reduced for smaller differences (supplementary material, Supplementary Material online). Similarly, for hemizygous X-linked genes, we calculate that differences of approximately 10% could be detected with approximately 80% power. All data analysis was done in R (R Development Core Team 2013; scripts are available for download from GitHub).
Supplementary Material
Acknowledgments
The authors thank Rebecca Dean and Devin M Drown for comments on the manuscript. This research was supported by Discovery Grants to SCHB and SIW from the Natural Sciences and Engineering Council of Canada. J.H. was supported by an Ontario Graduate Fellowship and J.A.Å. by a Junior Fellowship from Massey College.
Literature Cited
- Aanen DK, Spelbrink JN, Beekman M. What cost mitochondria? The maintenance of functional mitochondrial DNA within and across generations. Philos Trans R Soc Lond B Biol Sci. 2014;369 doi: 10.1098/rstb.2013.0438. 2013043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adams KL, Palmer JD. Evolution of mitochondrial gene content: gene loss and transfer to the nucleus. Mol Phylogenet Evol. 2003;29:380–395. doi: 10.1016/s1055-7903(03)00194-5. [DOI] [PubMed] [Google Scholar]
- Bar-Yaacov D, Blumberg A, Mishmar D. Mitochondrial-nuclear co-evolution and its effects on OXPHOS activity and regulation. Biochim Biophys Acta. 2012;1819:1107–1111. doi: 10.1016/j.bbagrm.2011.10.008. [DOI] [PubMed] [Google Scholar]
- Berg OG, Kurland CG. Why mitochondrial genes are most often found in nuclei. Mol Biol Evol. 2000;17:951–961. doi: 10.1093/oxfordjournals.molbev.a026376. [DOI] [PubMed] [Google Scholar]
- Burt A, Trivers R. Genes in conflict: the biology of selfish genetic elements. Cambridge: Belknap Press of Harvard University Press; 2006. [Google Scholar]
- Charlesworth D. Plant sex chromosome evolution. J Exp Bot. 2013;64:405–420. doi: 10.1093/jxb/ers322. [DOI] [PubMed] [Google Scholar]
- Conesa A, et al. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics. 2005;21:3674–3676. doi: 10.1093/bioinformatics/bti610. [DOI] [PubMed] [Google Scholar]
- Connallon T, Clark AG. The resolution of sexual antagonism by gene duplication. Genetics. 2011;187:919–937. doi: 10.1534/genetics.110.123729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortez D, et al. Origins and functional evolution of Y chromosomes across mammals. Nature. 2014;508:488–493. doi: 10.1038/nature13151. [DOI] [PubMed] [Google Scholar]
- Cosmides LM, Tooby J. Cytoplasmic inheritance and intragenomic conflict. J Theor Biol. 1981;89:83–129. doi: 10.1016/0022-5193(81)90181-8. [DOI] [PubMed] [Google Scholar]
- Dean R, Zimmer F, Mank JE. The potential role of sexual conflict and sexual selection in shaping the genomic distribution of mito-nuclear genes. Genome Biol Evol. 2014;6:1096–1104. doi: 10.1093/gbe/evu063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drown DM, Preuss KM, Wade MJ. Evidence of a paucity of genes that interact with the mitochondrion on the X in mammals. Genome Biol Evol. 2012;4:763–768. doi: 10.1093/gbe/evs064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dyall SD, Brown MT, Johnson PJ. Ancient invasions: from endosymbionts to organelles. Science. 2004;304:253–257. doi: 10.1126/science.1094884. [DOI] [PubMed] [Google Scholar]
- Elo A, Lyznik A, Gonzalez DO, Kachman SD, Mackenzie SA. Nuclear genes that encode mitochondrial proteins for DNA and RNA metabolism are clustered in the Arabidopsis genome. Plant Cell. 2003;15:1619–1631. doi: 10.1105/tpc.010009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gene Ontology Consortium. The gene ontology project in 2008. Nucleic Acids Res. 2008;36:D440–D444. doi: 10.1093/nar/gkm883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillham NW. Organelle genes and genomes. New York: Oxford University Press; 1994. [Google Scholar]
- Hill GE, Johnson JD. The mitonuclear compatibility hypothesis of sexual selection. Proc R Soc B Biol Sci. 2013;280:20131314. doi: 10.1098/rspb.2013.1314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hough J, Hollister JD, Wang W, Barrett SCH, Wright SI. Genetic degeneration of old and young Y chromosomes in the flowering plant Rumex hastatulus. Proc Natl Acad Sci U S A. 2014;111:7713–7718. doi: 10.1073/pnas.1319227111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondo M, Nanda I, Hornung U, Schmid M, Schartl M. Evolutionary origin of the medaka Y chromosome. Curr Biol. 2004;14:1664–1669. doi: 10.1016/j.cub.2004.09.026. [DOI] [PubMed] [Google Scholar]
- Lane N. Power, sex, suicide: mitochondria and the meaning of life. Oxford: Oxford University Press; 2005. [Google Scholar]
- Martin W. Gene transfer from organelles to the nucleus: frequent and in big chunks. Proc Natl Acad Sci U S A. 2003;100:8612–8614. doi: 10.1073/pnas.1633606100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Natri HM, Shikano T, Merilä J. Progressive recombination suppression and differentiation in recently evolved neo-sex chromosomes. Mol Biol Evol. 2013;30:1131–1144. doi: 10.1093/molbev/mst035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navajas-Perez R, et al. The evolution of reproductive systems and sex-determining mechanisms within Rumex (Polygonaceae) inferred from nuclear and chloroplastidial sequence data. Mol Biol Evol. 2005;22:1929–1939. doi: 10.1093/molbev/msi186. [DOI] [PubMed] [Google Scholar]
- Quevillon E, et al. InterProScan: protein domains identifier. Nucleic Acids Res. 2005;33:W116–W120. doi: 10.1093/nar/gki442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Development Core Team . Vienna (Austria): R Foundation for Statistical Computing; 2013. R: a language and environment for statistical computing. [Google Scholar]
- Rand DM, Clark AG, Kann LM. Sexually antagonistic cytonuclear fitness interactions in Drosophila melanogaster. Genetics. 2001;159:173–187. doi: 10.1093/genetics/159.1.173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rand DM, Haney RA, Fry AJ. Cytonuclear coevolution: the genomics of cooperation. Trends Ecol Evol. 2004;19:645–653. doi: 10.1016/j.tree.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Ridley M. Mendel's demon: gene justice and the complexity of life. London: Weidenfeld & Nicolson; 2000. [Google Scholar]
- Rogell B, Dean R, Lemos B, Dowling DK. Mito-nuclear interactions as drivers of gene movement on and off the X chromosome. BMC Genomics. 2014;15:330. doi: 10.1186/1471-2164-15-330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sloan DB. Using plants to elucidate the mechanisms of cytonuclear co-evolution. New Phytol. 2014 doi: 10.1111/nph.12835. Advance Access published May 6, 2014, doi: 10.1111/nph.12835. [DOI] [PubMed] [Google Scholar]
- Timmis JN, Ayliffe MA, Huang CY, Martin W. Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes. Nat Rev Genet. 2004;5:123–135. doi: 10.1038/nrg1271. [DOI] [PubMed] [Google Scholar]
- Touzet P, Meyer EH. Cytoplasmic male sterility and mitochondrial metabolism in plants. Mitochondrion. 2014 doi: 10.1016/j.mito.2014.04.009. Advance Access published April 24, 2014, doi: 10.1016/j.mito.2014.04.009. [DOI] [PubMed] [Google Scholar]
- Werren JH, Beukeboom LW. Sex determination, sex ratios, and genetic conflict. Annu Rev Ecol Syst. 1998;29:233–261. [Google Scholar]
- Wu CI, Yujun Xu E. Sexual antagonism and X inactivation—the SAXI hypothesis. Trends Genet. 2003;19:243–247. doi: 10.1016/s0168-9525(03)00058-1. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.