

Abstract
AIM: To examine the effect of farnesoid X receptor (FXR) activation by GW4064 on endotoxin-induced hepatic inflammation in nonalcoholic fatty liver disease (NAFLD) and the underlying mechanism.
METHODS: Six-week-old male C57BL/6 mice were fed a normal diet or a high-fat (HF) diet for 8 wk. HF diet-fed mice were intraperitoneally injected with GW4064 (30 mg/kg) or DMSO (vehicle) once daily for a week and then sacrificed after lipopolysaccharide (LPS, 50 μg/mouse) administration. Hepatic inflammation, levels of the macrophage marker F4/80, and apoptosis were measured at the end of the study. Additionally, the expression of proinflammatory genes involved in NAFLD (interleukin-6, interleukin-1β, interferon-γ, MCP-1) were analyzed by real-time PCR in the murine macrophage cell line RAW 264.7 cultured with or without GW4064 (2 μmol/L) before treatment with LPS.
RESULTS: In patients with NAFLD, the expression of FXR was detected by immunohistochemical staining and the relation between FXR expression and NAFLD activity score (NAS) was analyzed. Activation of FXR by GW4064 alleviated hepatic inflammation induced by endotoxin in a murine NAFLD model fed an HF diet as reflected by reduced serum levels of aspartate aminotransferase and alanine aminotransferase. Apoptosis and proinflammatory cytokine levels in liver tissues were also reduced by GW4064, and GW4064 could reduce induction of proinflammatory cytokines by LPS in vitro. FXR levels were reduced in patients with non-alcoholic steatohepatitis compared with healthy controls and were negatively correlated with NAS.
CONCLUSION: FXR activation attenuates LPS-induced hepatic inflammation in murine NAFLD by reducing expression of proinflammatory cytokines in macrophages.
Keywords: Farnesoid X receptor, Nonalcoholic fatty liver disease, GW4064, Nonalcoholic fatty liver disease activity score, Lipopolysaccharide-induced hepatic inflammation, Macrophage activation
Core tip: We found that the synthetic farnesoid X receptor (FXR) agonist GW4064 attenuated lipopolysaccharide-induced hepatic injury in high-fat diet-fed mice by reducing proinflammatory cytokine expression in macrophages. Moreover, we found that FXR expression was decreased in patients with nonalcoholic steatohepatitis, the most extreme form of nonalcoholic fatty liver disease (NAFLD), and was inversely correlated with the NAFLD activity score. Together, these findings provide insights into the etiology of NAFLD and suggest that FXR activation with synthetic agonists may represent a potential therapeutic option for patients with NAFLD, which we believe will be of great interest to the readers of the journal.
INTRODUCTION
Nonalcoholic fatty liver disease (NAFLD) affects a large population worldwide[1-4]. NAFLD is frequently associated with metabolic syndrome including insulin resistance, diabetes, obesity, and hyperlipidemia[5-8]. The spectrum of NAFLD ranges from simple steatosis without inflammation to nonalcoholic steatohepatitis (NASH) and cirrhosis[9]. Currently, liver transplantation is the only therapeutic option for patients with end-stage NASH.
Farnesoid X receptor (FXR), a nuclear receptor for bile acids, is highly expressed in the liver, intestine, kidney, heart, and adipose tissue. FXR has pleiotropic functions in bile metabolism, immune modulation, and lipid metabolism[10-14] and plays a critical role in maintaining bile acid and cholesterol homeostasis by regulating cholesterol catabolism, bile acid synthesis, and absorption[15,16]. Recently, the role of FXR in lipid and glucose metabolism has been explored. FXR activation lowers plasma and liver triglyceride levels by enhancing plasma triglyceride clearance and repressing hepatic lipogenesis[17]. FXR-null mice fed a normal diet display elevated serum glucose and impaired insulin tolerance[18-20]. In contrast, FXR deficiency protects mice from high-fat diet-induced obesity and improves glucose homeostasis[21,22].
How FXR affects hepatic inflammation in NASH is not well understood. In this study, we examined the protective effects of FXR activation by GW4064 in mice with hepatic inflammation induced by lipopolysaccharide (LPS). We also investigated the possible mechanism of alleviation of hepatic inflammation by GW4064 in a murine model of NAFLD. Our data demonstrate that FXR activation suppresses hepatic inflammation and apoptosis by inhibiting secretion of proinflammatory cytokines by macrophages.
MATERIALS AND METHODS
Animals
Male wild-type C57BL/6 mice, aged 6-8 wk, were purchased from Shanghai Slac Laboratory Animal Co., Ltd.. All animals were housed in the animal house of the Central Laboratory of our hospital under pathogen-free conditions. There were five mice in each experimental group. Mice were fed diets (BioServ, Inc., Frenchtown, NJ) containing either high amounts of fat (HF, 59% kcal from fat, 25% total carbohydrate, 16% protein) or normal amounts of fat (ND, 12% kcal from fat, 59% total carbohydrate, 29% protein) for 8 wk. Mice received 30 mg/kg of the FXR agonist GW4064 (Cayman Chemical Co., Ann Arbor, MI) through daily intraperitoneal injection for 7 d after the mice had been on an HF diet for 7 wk. HF diet-fed mice were injected intraperitoneally with Escherichia coli LPS (50 μg/mouse, Sigma, St. Louis, MO) and sacrificed 6 h later to obtain serum and liver tissues. Body and liver weights were measured for all mice.
Human liver samples
Hepatic hemangioma samples and normal adjacent liver tissue were obtained upon surgical resection. The experiments were approved by the Ethics Committee of the First People’s Hospital of Zhenjiang and all patients signed an informed consent form.
Serum alanine aminotransferase and aspartate aminotransferase levels
Serum alanine aminotransferase (AST) and aspartate aminotransferase (ALT) levels were measured in the Clinical Chemistry Laboratory of the Department of Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences.
Histology and terminal deoxynucleotidyl transferase dUTP nick end labeling
All liver tissues were fixed in buffered formaldehyde (10%) overnight, embedded in paraffin, and cut into 4-μm sections. Liver tissue sections were stained with hematoxylin and eosin. The histology was evaluated by an experienced hepatologist to determine NAFLD activity score (NAS; Ref. 9) in a blinded fashion. A transferase dUTP nick end labeling (TUNEL) FITC kit (ShanghaRuian bioTechnologies Co., Ltd, Shanghai, China) was used to detect cell apoptosis in murine liver tissues according to the manufacturer’s instructions.
Immunohistochemistry
Slides were incubated with 3% H2O2 for 10 min after dewaxing, then washed in phosphate-buffered saline (PBS). Antigen retrieval was done in citrate buffer (pH 6.0) by microwave heating, and blocking was performed with 10% non-immune goat serum for 30 min after cooling. Slides were incubated with an anti-F4/80 antibody (Abcam, Cambridge, United Kingdom) overnight at 4 °C. After rinsing with PBS, sections were incubated with an HRP-conjugated secondary antibody (Changdao, Shanghai, China) for 30 min at room temperature and then stained with 3,3’-diaminobenzidine (DAB; Maixin-Bio, Guangzhou, China) for 3 min. Hematoxylin was applied as a counterstain. Five fields were randomly selected for quantification of positive cells in every sample.
Cell culture
The murine macrophage RAW 264.7 cell line was purchased from Shanghai Institute of Cell Biology (Chinese Academy of Sciences). RAW 264.7 cells were cultured in RPMI-1640 with 10% FBS (v/v) at 37 °C and 5% CO2 in a humidified atmosphere. GW4064 (2 μmol/L) was administered for 24 h before treatment with LPS (1 μg/mL). RNA was then extracted from cells (see below).
RNA extraction and real-time polymerase chain reaction
Total RNA was extracted from liver tissues and cells using TRIzol reagent (Invitrogen, Carlsbad, CA) and complementary DNA was synthesized according to the manufacturer’s instructions (Takara, Shiga, Japan). Real-time quantitative polymerase chain reaction (PCR) assays (SYBR Green) were carried out using a 7900HT Fast Real-Time PCR System (ABI/Life Technologies). The primer sequences of Mcp1, Il1b, Il17, Il6, Ifng and Tnf are described in Table 1. 18S rRNA, a housekeeping gene, was used as an internal standard.
Table 1.
Primer sequences
| Mcp1 | CAGCCAGATGCAGTTAACGC and GCCTACTCATTGGGATCATCTTG |
| Il6 | TCCATCCAGTTGCCTTCTTG and TTCCACGATTTCCCAGAGAAC |
| Tnf | CCACATCTCCCTCCAGAAAA and AGGGTCTGGGCCATAGAACT |
| Ifng | TCAAGTGGCATAGATGTGGAAGAA and TGGCTCTGCAGGATTTTCATG |
| Il1b | GCAACTGTTCCTGAACTCAACT and ATCTTTTGGGGTCCGTCAACT |
| Il17 | TTTAACTCCCTTGGCGCAAAA and CTTTCCCTCCGCATTGACAC |
| 18S | CGGACAGGATTGACAGATTGATAGCandTGCCAGAGTCTCGTTCGTTATCG |
Statistical analysis
All values are expressed as mean ± SD. The group means were compared by one-way analysis of variance (ANOVA). All statistical analyses were performed using GraphPad Prism 5 (GraphPad Software, San Diego, California). P values < 0.05 were considered statistically significant.
RESULTS
GW4064 alleviates LPS-induced liver injury in a murine NAFLD model
To determine whether activation of bile acid receptors (FXR) affects hepatic inflammation, we administered the FXR bile acid receptor agonist GW4064 to HF diet-fed mice for 1 wk. In mice treated with LPS only, there were many inflammatory cells, but after administration of GW4064 the number of inflammatory cells within the liver decreased (Figure 1A). No changes in bile duct morphology were observed in either group. In order to evaluate the histological changes in mice treated with GW4064, the NAFLD activity score was used. The group treated with GW4064 had a lower NAFLD score than mice treated with LPS only (Figure 1B). Treatment with the FXR agonist GW4064 also significantly decreased serum ALT and AST levels in HF diet-fed mice treated with LPS, further suggesting that FXR may have a protective role against liver injury in this NAFLD model (Figure 1C, D).
Figure 1.

GW4064 alleviates lipopolysaccharide-induced liver injury in a murine nonalcoholic fatty liver disease model. Mice fed on a high-fat (HF) diet for 8 wk received GW4064 for a week before the administration of lipopolysaccharide (LPS). Serum and liver tissues were obtained 6 h after LPS injection. A: Representative hematoxylin and eosin staining of liver sections (arrows) from normal diet, HF, HF + LPS, and HF + GW4064 + LPS mice; B: NAFLD activity scores (NAS) of normal diet, HF, HF + LPS, and HF + GW4064 + LPS mice; C: Serum levels of aspartate aminotransferase (AST) (mean ± SD) of normal diet, HF, HF + LPS, and HF + GW4064 + LPS mice (n = 5/group). aP < 0.05, bP < 0.01 vs HF + LPS group; D: Serum levels of alanine aminotransferase (ALT) (mean ± SD) of normal diet, HF, HF + LPS, and HF + GW4064 + LPS mice (n = 5/group). aP < 0.05, bP < 0.01 vs HF + LPS group.
GW4064 reduces apoptosis and macrophage infiltration in a murine NAFLD model
Apoptosis and macrophage infiltration are hallmarks of NASH, with macrophages regarded as the major drivers of NASH pathogenesis given that they are the major source of proinflammatory cytokines in both adipose tissue and the liver in NASH. Macrophages can cause hepatocyte damage and promote insulin resistance by secreting cytokines such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6)[23]. We wondered whether the FXR agonist GW4064 would affect hepatocyte apoptosis and macrophage infiltration in a murine model of NAFLD. Whereas only a few TUNEL-positive cells were observed in mice fed an ND or HF diet only, more TUNEL-positive cells were observed after treatment with LPS (about ten in each high power field, HPF). However, if we treated mice with GW4064 for 1 wk before administration of LPS, TUNEL-positive cells were decreased (Figure 2A, B). Consistent with these findings, immunohistochemical staining for the macrophage marker F4/80 in liver tissues was also increased upon LPS treatment. Moreover, GW4064 reduced apoptosis and macrophage infiltration induced by LPS in HF diet-fed mice (Figure 2C-E).
Figure 2.

GW4064 reduces apoptosis and macrophage infiltration in a murine nonalcoholic fatty liver disease model. A: Representative terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining of liver sections (magnification × 400) from normal diet, high-fat (HF), HF + lipopolysaccharide (LPS), and HF + GW4064 + LPS mice; B: Quantification of TUNEL in each high power field (HPF); C: Representative immunohistochemical staining of F4/80 in the livers (magnification × 400) of normal diet, HF, HF + LPS, and HF + GW4064 + LPS mice; D: Quantification of F4/80-positive cells in each HPF. aP < 0.05, bP < 0.01 vs HF + LPS group.
GW4064 reduces hepatic proinflammatory cytokine expression in a murine model of NAFLD and in murine macrophages
Proinflammatory cytokines such as TNF-α, MCP-1, IL-1β, and IL-6 participate in macrophage infiltration, lipid metabolism, and insulin resistance and are involved in the pathogenesis of NAFLD. Neutralization of TNF-α or MCP-1 activity has been shown to ameliorate insulin resistance and fatty liver disease in animal models[24,25]. In order to demonstrate whether GW4064 affects proinflammatory cytokines in a murine model of NAFLD, RT-PCR was used to detect the mRNA levels of Tnfa, Mcp1, Il1b, and Il6. We found that expression levels of Tnfa, Mcp1, Il1b, and Il6 mRNAs were significantly increased in the livers of HF diet-fed mice treated with LPS, whereas GW4064 treatment attenuated LPS-induced expression of Tnfa, Mcp1, Il1b and Il6 in HF-diet fed mice (Figure 3).
Figure 3.
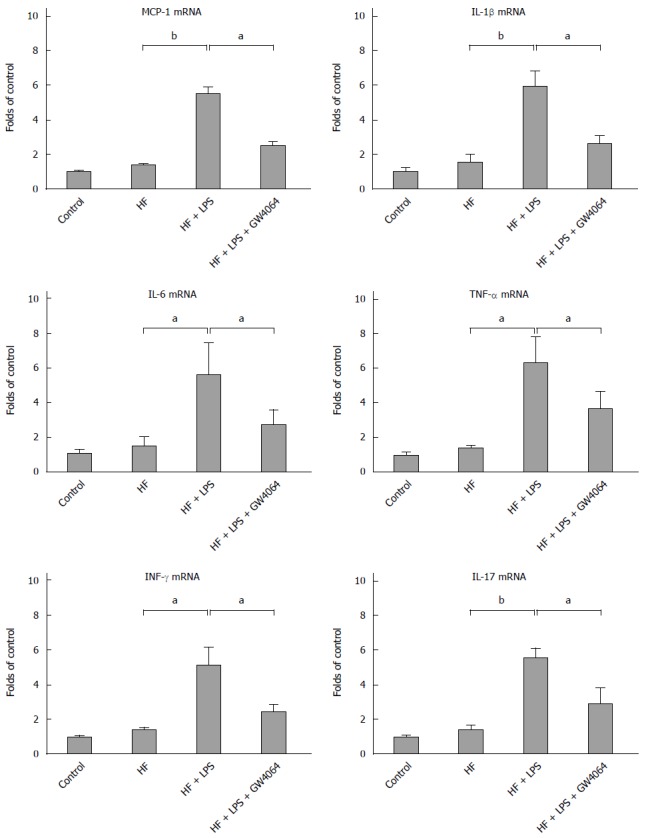
GW4064 reduces hepatic proinflammatory cytokine expression in a murine nonalcoholic fatty liver disease model. RNA was extracted from the liver tissues of normal diet, high-fat (HF), HF + lipopolysaccharide (LPS), and HF + GW4064 + LPS mice. Real-time polymerase chain reaction was used to determine the mRNA levels of Tnf, Mcp1, Il1b, Il6, Il17, and Ifng. aP < 0.05, bP < 0.01 vs HF + LPS group.
In order to determine whether GW4064 can directly regulate proinflammatory cytokine expression in macrophages, we treated the murine macrophage cell line RAW 264.7 with GW4064 for 24 h before administration of LPS. RT-PCR was then used to determine the levels of proinflammatory cytokine expression. LPS treatment dramatically increased the levels of Il1b, Il6, and Ifng, but pretreatment with GW4064 prevented the induction of cytokine expression induced by LPS (Figure 4).
Figure 4.
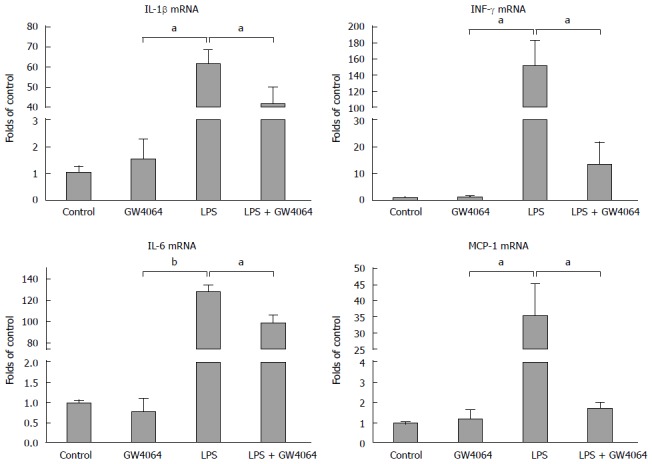
GW4064 represses lipopolysaccharide-induced proinflammatory cytokine expression in RAW 264.7 cells. RAW 264.7 cells were cultured in Dulbecco's Modified Eagle Media with 10% FBS, treated with GW4064 for 24 h, then treated with lipopolysaccharide (LPS) for 6 h. Expression of Mcp1, Il1b, Il6, and Ifng in RAW 264.7 cells was evaluated by real-time polymerase chain reaction. aP < 0.05, bP < 0.01 vs LPS group.
FXR is decreased and reversely correlated with NAS in patients
FXR expression in the livers of NASH patients was examined by immunohistochemical staining. FXR is highly expressed in hepatocytes in healthy control livers (Table 2), but less so in the livers of patients with NASH, a condition associated with high-grade steatosis, lobular inflammation, and ballooning (Figure 5A). We counted the number of FXR-positive cells in healthy controls and patients with NASH and found that there are about 25 FXR-positive cells per HPF in healthy controls, but only 10 in patients with NASH (Figure 5B). NAS, a scoring system for NAFLD, is used to assess the degree of NAFLD in clinical and experimental studies. We found that FXR expression was inversely correlated with NAS in patients with NASH (Figure 5C).
Table 2.
Clinical and biochemical features of healthy control subjects and patients with nonalcoholic steatohepatitis
| Healthy control (n = 4) | NASH (n = 18) | P value | |
| Female (n) | 2 | 7 | NS |
| Age (yr) | 47.20 ± 6.04 | 51.29 ± 12.33 | NS |
| ALT (U/L) | 32.40 ± 17.09 | 128.27 ± 98.07 | < 0.05 |
| AST (U/L) | 16.80 ± 4.92 | 71.71 ± 53.47 | < 0.05 |
| Alkaline phosphatase (U/L) | 96.20 ± 17.83 | 101.74 ± 24.09 | NS |
| Gamma-glutamyl transferase (U/L) | 14.00 ± 15.68 | 95.40 ± 56.94 | < 0.05 |
| BMI (kg/m2) | 19.78 ± 1.19 | 26.63 ± 1.55 | < 0.05 |
| Fasting insulin (uIU/mL) | 7.75 ± 4.15 | 14.05 ± 2.94 | < 0.05 |
| Triglycerides (mmol/L) | 1.59 ± 0.10 | 2.85 ± 0.92 | < 0.05 |
All values are means ± SD. NASH: Nonalcoholic steatohepatitis; ALT: Alanine aminotransferase; AST: Aspartate aminotransferase; NS: Not significant.
Figure 5.
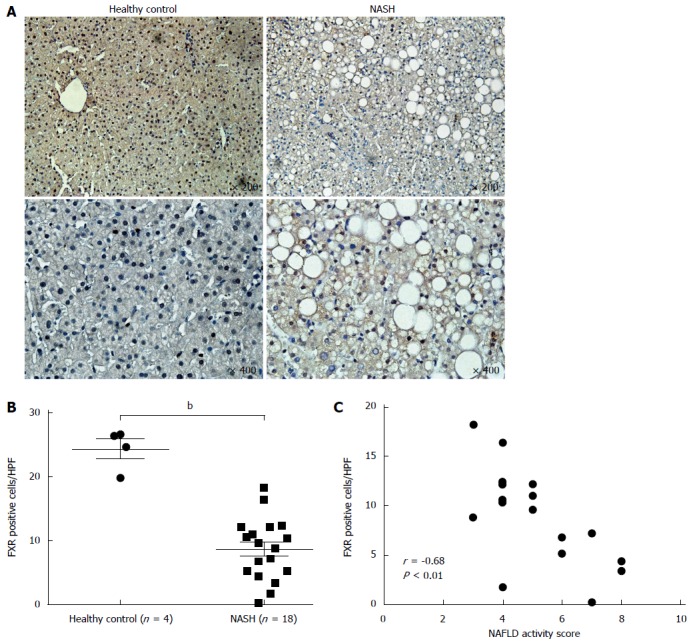
Farnesoid X receptor expression is decreased and inversely correlated with nonalcoholic fatty liver disease activity score in patients. Expression of farnesoid X receptor (FXR) in patients was detected by immunohistochemical staining. A: Representative FXR staining of liver sections from healthy control subjects and patients with nonalcoholic steatohepatitis (NASH); B: Quantification of FXR-positive cells in each high power field (HPF); C: The correlation between FXR expression and nonalcoholic fatty liver disease activity score (NAS) in NASH patients. bP < 0.01 vs health control group.
DISCUSSION
The current study demonstrates that activation of FXR by GW4064 alleviates hepatic inflammation and macrophage infiltration in a murine model of NAFLD. GW4064 treatment significantly decreased the NAS, serum levels of ALT and AST, and proinflammatory cytokines, all of which are characteristics of hepatic injury. LPS-induced hepatocyte apoptosis and macrophage infiltration were dramatically reduced in mice treated with GW4064, and GW4064 repressed LPS-induced proinflammatory cytokine expression in macrophages in vitro. In patients with NASH, FXR expression was decreased compared with healthy controls, and the level of FXR expression was negatively correlated with NAS. These results provide direct evidence that FXR activation is an effective method to block hepatic inflammation induced by an HF diet and LPS that operates by repressing proinflammatory cytokine expression in macrophages.
Kupffer cells, also known as stellate macrophages, are the largest component of the reticuloendothelial system and represent about 10% of the resting total liver cell population[26,27]. Kupffer cells are important to innate immunity and are responsible for the rapid containment and clearance of exogenous particulate and immunoreactive material. In NAFLD, Kupffer cells primarily recognize endotoxin. After activation by LPS in NAFLD, Kupffer cells are enabled to initiate a biochemical attack and interactions with hepatocytes by releasing proinflammatory cytokines, chemokines, reactive oxygen species, and nitric oxide[28]. We found that GW4064 treatment decreased the level of LPS-induced macrophage infiltration in the livers of mice fed an HF diet in association with a reduction in hepatic inflammation.
There are two macrophage phenotypes in the livers of patients with NAFLD: the classical M1 (proinflammatory phenotype) and M2 (alternatively activated macrophages)[29]. The alternative activation of macrophages is primarily induced by IL-4 and IL-13[30] and characterized by the expression of M2 signature genes such as those encoding arginase-1 and the mannose receptor. Recently, McMahan et al[31] reported that INT-767 could alter the phenotype of monocytes. We have shown that FXR activation by GW4064 inhibits the production of proinflammatory cytokines by macrophages and decreases intrahepatic expression of the genes encoding TNF-α, MCP-1, IL-1β, and IL-6.
FXR, a metabolic nuclear receptor abundantly expressed in the liver, intestine, and kidney, is a major transcriptional factor involved in glucose and lipid metabolism in the liver[32]. Previous studies showed that mice with FXR deficiency displayed hepatic steatosis as well as glucose and insulin intolerance, the main hallmarks of NAFLD in humans. At the molecular level, FXR controls lipid and glucose homeostasis through the regulation of PGC-1α, FOXO1 and SIRT1[33]. Recent findings have shown that FXR also participates in lipid metabolism by regulating CD36, a mediator of long-chain fatty acid transport, which reduces hepatic steatosis in mice fed an HF diet[34]. Repression of FXR in obese mice promotes hepatic steatosis[35], and hepatic FXR was reduced in Zucker fatty rats[36], which are consistent with our findings in patients with NASH.
Collectively, our data indicate that treatment with the FXR agonist GW4064 decreases intrahepatic inflammation in a murine model of NAFLD by inhibiting proinflammatory cytokine expression in macrophages and reducing hepatocyte apoptosis and serum levels of ALT and AST. These findings suggest that FXR activation using synthetic agonists may represent a promising therapeutic option for fatty liver diseases.
COMMENTS
Background
Nonalcoholic fatty liver disease (NAFLD) affects a large population worldwide. NAFLD is frequently associated with metabolic syndrome including insulin resistance, diabetes, obesity, and hyperlipidemia. The spectrum of NAFLD ranges from simple steatosis without inflammation to nonalcoholic steatohepatitis (NASH) and cirrhosis. Currently, liver transplantation is the only therapeutic option for patients with end-stage NASH.
Research frontiers
The current study demonstrates that activation of farnesoid X receptor (FXR) by GW4064 alleviates hepatic inflammation and macrophage infiltration in a murine model of NAFLD. In patients with NASH, FXR expression was decreased compared with healthy controls, and the level of FXR expression was negatively correlated with NAS.
Innovations and breakthroughs
Authors found that the synthetic FXR agonist GW4064 attenuated lipopolysaccharide-induced hepatic injury in high-fat mice by reducing proinflammatory cytokine expression in macrophages. Moreover, they found that FXR expression was decreased in patients with nonalcoholic steatohepatitis, the most extreme form of nonalcoholic fatty liver disease (NAFLD), and was inversely correlated with the NAFLD activity score. Together, these findings provide insights into the etiology of NAFLD and suggest that FXR activation with synthetic agonists may represent a potential therapeutic option for patients with NAFLD, which they believe will be of great interest to the readers of the journal.
Peer review
In the present manuscript, authors investigated the role of FXR in a model of NAFLD. The authors used the FXR agonist GW4064 for their study. C57BL/6 mice were fed either a normal diet or a high fat diet. In addition animals were additionally injected with lipopolysaccharide. In vitro the study was performed with the murine macrophage cell line RAW 264.7.
Footnotes
P- Reviewer: Plentz RR S- Editor: Gou SX L- Editor: Wang TQ E- Editor: Ma S
References
- 1.Bechmann LP, Kocabayoglu P, Sowa JP, Sydor S, Best J, Schlattjan M, Beilfuss A, Schmitt J, Hannivoort RA, Kilicarslan A, et al. Free fatty acids repress small heterodimer partner (SHP) activation and adiponectin counteracts bile acid-induced liver injury in superobese patients with nonalcoholic steatohepatitis. Hepatology. 2013;57:1394–1406. doi: 10.1002/hep.26225. [DOI] [PubMed] [Google Scholar]
- 2.Puri P, Wiest MM, Cheung O, Mirshahi F, Sargeant C, Min HK, Contos MJ, Sterling RK, Fuchs M, Zhou H, et al. The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology. 2009;50:1827–1838. doi: 10.1002/hep.23229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sheth SG, Gordon FD, Chopra S. Nonalcoholic steatohepatitis. Ann Intern Med. 1997;126:137–145. doi: 10.7326/0003-4819-126-2-199701150-00008. [DOI] [PubMed] [Google Scholar]
- 4.Cohen JC, Horton JD, Hobbs HH. Human fatty liver disease: old questions and new insights. Science. 2011;332:1519–1523. doi: 10.1126/science.1204265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Li Y, Jadhav K, Zhang Y. Bile acid receptors in non-alcoholic fatty liver disease. Biochem Pharmacol. 2013;86:1517–1524. doi: 10.1016/j.bcp.2013.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Neuschwander-Tetri BA. Hepatic lipotoxicity and the pathogenesis of nonalcoholic steatohepatitis: the central role of nontriglyceride fatty acid metabolites. Hepatology. 2010;52:774–788. doi: 10.1002/hep.23719. [DOI] [PubMed] [Google Scholar]
- 7.Marchesini G, Brizi M, Morselli-Labate AM, Bianchi G, Bugianesi E, McCullough AJ, Forlani G, Melchionda N. Association of nonalcoholic fatty liver disease with insulin resistance. Am J Med. 1999;107:450–455. doi: 10.1016/s0002-9343(99)00271-5. [DOI] [PubMed] [Google Scholar]
- 8.Hebbard L, George J. Animal models of nonalcoholic fatty liver disease. Nat Rev Gastroenterol Hepatol. 2011;8:35–44. doi: 10.1038/nrgastro.2010.191. [DOI] [PubMed] [Google Scholar]
- 9.Harrison SA, Oliver D, Arnold HL, Gogia S, Neuschwander-Tetri BA. Development and validation of a simple NAFLD clinical scoring system for identifying patients without advanced disease. Gut. 2008;57:1441–1447. doi: 10.1136/gut.2007.146019. [DOI] [PubMed] [Google Scholar]
- 10.Bhargava P, Li C, Stanya KJ, Jacobi D, Dai L, Liu S, Gangl MR, Harn DA, Lee CH. Immunomodulatory glycan LNFPIII alleviates hepatosteatosis and insulin resistance through direct and indirect control of metabolic pathways. Nat Med. 2012;18:1665–1672. doi: 10.1038/nm.2962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chen Y, Song X, Valanejad L, Vasilenko A, More V, Qiu X, Chen W, Lai Y, Slitt A, Stoner M, et al. Bile salt export pump is dysregulated with altered farnesoid X receptor isoform expression in patients with hepatocellular carcinoma. Hepatology. 2013;57:1530–1541. doi: 10.1002/hep.26187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Modica S, Petruzzelli M, Bellafante E, Murzilli S, Salvatore L, Celli N, Di Tullio G, Palasciano G, Moustafa T, Halilbasic E, et al. Selective activation of nuclear bile acid receptor FXR in the intestine protects mice against cholestasis. Gastroenterology. 2012;142:355–365.e1-e4. doi: 10.1053/j.gastro.2011.10.028. [DOI] [PubMed] [Google Scholar]
- 13.Chen F, Ma L, Dawson PA, Sinal CJ, Sehayek E, Gonzalez FJ, Breslow J, Ananthanarayanan M, Shneider BL. Liver receptor homologue-1 mediates species- and cell line-specific bile acid-dependent negative feedback regulation of the apical sodium-dependent bile acid transporter. J Biol Chem. 2003;278:19909–19916. doi: 10.1074/jbc.M207903200. [DOI] [PubMed] [Google Scholar]
- 14.Lefebvre P, Cariou B, Lien F, Kuipers F, Staels B. Role of bile acids and bile acid receptors in metabolic regulation. Physiol Rev. 2009;89:147–191. doi: 10.1152/physrev.00010.2008. [DOI] [PubMed] [Google Scholar]
- 15.Claudel T, Zollner G, Wagner M, Trauner M. Role of nuclear receptors for bile acid metabolism, bile secretion, cholestasis, and gallstone disease. Biochim Biophys Acta. 2011;1812:867–878. doi: 10.1016/j.bbadis.2010.12.021. [DOI] [PubMed] [Google Scholar]
- 16.Zollner G, Marschall HU, Wagner M, Trauner M. Role of nuclear receptors in the adaptive response to bile acids and cholestasis: pathogenetic and therapeutic considerations. Mol Pharm. 2006;3:231–251. doi: 10.1021/mp060010s. [DOI] [PubMed] [Google Scholar]
- 17.Watanabe M, Houten SM, Wang L, Moschetta A, Mangelsdorf DJ, Heyman RA, Moore DD, Auwerx J. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J Clin Invest. 2004;113:1408–1418. doi: 10.1172/JCI21025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ma K, Saha PK, Chan L, Moore DD. Farnesoid X receptor is essential for normal glucose homeostasis. J Clin Invest. 2006;116:1102–1109. doi: 10.1172/JCI25604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang Y, Lee FY, Barrera G, Lee H, Vales C, Gonzalez FJ, Willson TM, Edwards PA. Activation of the nuclear receptor FXR improves hyperglycemia and hyperlipidemia in diabetic mice. Proc Natl Acad Sci USA. 2006;103:1006–1011. doi: 10.1073/pnas.0506982103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stayrook KR, Bramlett KS, Savkur RS, Ficorilli J, Cook T, Christe ME, Michael LF, Burris TP. Regulation of carbohydrate metabolism by the farnesoid X receptor. Endocrinology. 2005;146:984–991. doi: 10.1210/en.2004-0965. [DOI] [PubMed] [Google Scholar]
- 21.Prawitt J, Abdelkarim M, Stroeve JH, Popescu I, Duez H, Velagapudi VR, Dumont J, Bouchaert E, van Dijk TH, Lucas A, et al. Farnesoid X receptor deficiency improves glucose homeostasis in mouse models of obesity. Diabetes. 2011;60:1861–1871. doi: 10.2337/db11-0030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang Y, Ge X, Heemstra LA, Chen WD, Xu J, Smith JL, Ma H, Kasim N, Edwards PA, Novak CM. Loss of FXR protects against diet-induced obesity and accelerates liver carcinogenesis in ob/ob mice. Mol Endocrinol. 2012;26:272–280. doi: 10.1210/me.2011-1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Baffy G. Kupffer cells in non-alcoholic fatty liver disease: the emerging view. J Hepatol. 2009;51:212–223. doi: 10.1016/j.jhep.2009.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li Z, Yang S, Lin H, Huang J, Watkins PA, Moser AB, Desimone C, Song XY, Diehl AM. Probiotics and antibodies to TNF inhibit inflammatory activity and improve nonalcoholic fatty liver disease. Hepatology. 2003;37:343–350. doi: 10.1053/jhep.2003.50048. [DOI] [PubMed] [Google Scholar]
- 25.Si Y, Tsou CL, Croft K, Charo IF. CCR2 mediates hematopoietic stem and progenitor cell trafficking to sites of inflammation in mice. J Clin Invest. 2010;120:1192–1203. doi: 10.1172/JCI40310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bouwens L, Baekeland M, De Zanger R, Wisse E. Quantitation, tissue distribution and proliferation kinetics of Kupffer cells in normal rat liver. Hepatology. 1986;6:718–722. doi: 10.1002/hep.1840060430. [DOI] [PubMed] [Google Scholar]
- 27.Bogdanos DP, Gao B, Gershwin ME. Liver immunology. Compr Physiol. 2013;3:567–598. doi: 10.1002/cphy.c120011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ioannou GN. Implications of elevated serum alanine aminotransferase levels: think outside the liver. Gastroenterology. 2008;135:1851–1854. doi: 10.1053/j.gastro.2008.11.005. [DOI] [PubMed] [Google Scholar]
- 29.Mylonas KJ, Hoeve MA, MacDonald AS, Allen JE. Alternative activation of macrophages by filarial nematodes is MyD88-independent. Immunobiology. 2013;218:570–578. doi: 10.1016/j.imbio.2012.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Biswas SK, Mantovani A. Macrophage plasticity and interaction with lymphocyte subsets: cancer as a paradigm. Nat Immunol. 2010;11:889–896. doi: 10.1038/ni.1937. [DOI] [PubMed] [Google Scholar]
- 31.McMahan RH, Wang XX, Cheng LL, Krisko T, Smith M, El Kasmi K, Pruzanski M, Adorini L, Golden-Mason L, Levi M, et al. Bile acid receptor activation modulates hepatic monocyte activity and improves nonalcoholic fatty liver disease. J Biol Chem. 2013;288:11761–11770. doi: 10.1074/jbc.M112.446575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Thomas C, Pellicciari R, Pruzanski M, Auwerx J, Schoonjans K. Targeting bile-acid signalling for metabolic diseases. Nat Rev Drug Discov. 2008;7:678–693. doi: 10.1038/nrd2619. [DOI] [PubMed] [Google Scholar]
- 33.Lee J, Seok S, Yu P, Kim K, Smith Z, Rivas-Astroza M, Zhong S, Kemper JK. Genomic analysis of hepatic farnesoid X receptor binding sites reveals altered binding in obesity and direct gene repression by farnesoid X receptor in mice. Hepatology. 2012;56:108–117. doi: 10.1002/hep.25609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ma Y, Huang Y, Yan L, Gao M, Liu D. Synthetic FXR agonist GW4064 prevents diet-induced hepatic steatosis and insulin resistance. Pharm Res. 2013;30:1447–1457. doi: 10.1007/s11095-013-0986-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lu Y, Ma Z, Zhang Z, Xiong X, Wang X, Zhang H, Shi G, Xia X, Ning G, Li X. Yin Yang 1 promotes hepatic steatosis through repression of farnesoid X receptor in obese mice. Gut. 2014;63:170–178. doi: 10.1136/gutjnl-2012-303150. [DOI] [PubMed] [Google Scholar]
- 36.Fiorucci S, Rizzo G, Donini A, Distrutti E, Santucci L. Targeting farnesoid X receptor for liver and metabolic disorders. Trends Mol Med. 2007;13:298–309. doi: 10.1016/j.molmed.2007.06.001. [DOI] [PubMed] [Google Scholar]