

Abstract
Patterns of plant distribution by palaeoenvironment were examined across the Pennsylvanian–Permian transition in North–Central Texas. Stratigraphically recurrent packages of distinct lithofacies, representing different habitats, contain qualitatively and quantitatively different macrofloras and microfloras. The species pools demonstrate niche conservatism, remaining closely tied to specific habitats, during both short-term cyclic environmental change and a long-term trend of increasing aridity. The deposits examined principally comprise the terrestrial Markley and its approximate marine equivalent, the Harpersville Formation and parts of lower Archer City Formation. Fossiliferous deposits are lens-like, likely representing fill sequences of channels formed during abandonment phases. Palaeosols, represented by blocky mudstones, comprise a large fraction of the deposits. They suggest progressive climate change from minimally seasonal humid to seasonal subhumid to seasonal dry subhumid. Five lithofacies yielded plants: kaolinite-dominated siltstone, organic shale, mudstone beds within organic shale, coarsening upward mudstone–sandstone interbeds and channel sandstone. Both macro- and microflora were examined. Lithofacies proved compositionally distinct, with different patterns of dominance diversity. Organic shales (swamp deposits), mudstone partings (swamp drainages) and coarsening upward mudstone–sandstone interbeds (floodplains) typically contain Pennsylvanian wetland vegetation. Kaolinite-dominated siltstones and (to the extent known) sandstones contain taxa indicative of seasonally dry substrates. Some kaolinite-dominated siltstones and organic shales/coals yielded palynomorphs. Microfloras are more diverse, with greater wetland–dryland overlap than macrofloras. It appears that these two floras were coexistent at times on the regional landscape.
The fossil record offers a unique opportunity to track species pools through time and to examine their responses to long-term, directional changes in physical conditions. The ability to estimate environmental variables, such as temperature, rainfall amount (and its seasonal distribution) and atmospheric CO2, is improving continuously. Links between proxy measures of such physical factors and quantitative data on changes in ecosystem composition and dynamics, formerly the purview of Pleistocene palaeoecology, are now available, even in the deep past, for both terrestrial (e.g. Wing et al. 2005; Falcon-Lang et al. 2006; Montañez et al. 2007; Secord et al. 2008; Currano et al. 2008; Tabor et al. 2011) and marine (Palaeozoic examples: Olszewski & Erwin 2009; Ivany et al. 2009; Fall & Olszewski 2010; Bonelli & Patzkowsky 2011; Holland & Zaffos 2011) systems. Plant fossils themselves can be especially useful as palaeoclimate proxies, even in the distant world of the Palaeozoic (e.g. Falcon-Lang et al. 2009), because of the strong sensitivities of plants to habitat and regional climate. As sessile organisms, plants are directly and inextricably linked to climate at the level of individual, population, local community and biome.
Quantitative floristic studies of vegetation from the Late Palaeozoic are numerous (e.g. Scott 1978; Pfefferkorn & Thomson 1982; Bartram 1987; Eble et al. 1994, 2006; Pryor & Gastaldo 2000; DiMichele et al. 2007; Opluštil et al. 2007; Šimůnek & Martínek 2009; Libertín et al. 2009; Bashforth et al. 2010), although few have had a framework with which to link plants and their associated environments to changing global climate (e.g. Phillips & Peppers 1984; Falcon-Lang 2004). It is a particular challenge to find a long, relatively uninterrupted stratigraphic section that contains both a common species pool or pools and a clear signal of an overall, directional and interpretable environmental change. The latest Pennsylvanian–earliest Permian Markley Formation of north–central Texas provides such an opportunity.
The Markley Fm was positioned in the western Pangean tropics during the Pennsylvanian–Permian transition, a time of global climate change that accompanied the onset of Permian glaciation (Fielding et al. 2008). Within the Markley Fm, its partial equivalent – the Harpersville Fm – and into the base of its successor – the Archer City Fm – there are distinct packets of lithofacies that repeat in the stratigraphic section. Each lithofacies contains a distinctive fossil-plant assemblage, characterized and described by Romanchock et al. (in press), which can be followed through the Markley Fm. The goal of this paper is to go beyond such a qualitative flora-to-facies correlation and to examine quantitatively the floristic similarity within and among these facies in space and through time in order to investigate, and in this instance document, the coherence of species assemblages in the face of short-term and long-term environmental changes, and to evaluate spatio-temporal scales on which niche versus neutral biological community assembly patterns may operate (e.g. Alonso et al. 2006; Weiher et al. 2011). In addition, we wish to examine the conformance of the Markley Fm data with interpretive models that call for climate change as the controlling variable on repeating patterns of lithofacies and plant distribution through time, in the short and long term (allocyclicity), v. models that call for changes in sedimentary environments as the driver of temporally recurrent floristic change under a relatively constant background climate (autocyclicity).
Geological setting
Geographic location
Collections from the Markley Fm, on which this study is based, were made across north–central Texas from 20 localities (Fig. 1). Eighteen of these localities are spread in a NE–SW transect across four counties: Montague, Jack, Young and Archer. Single collections come from the Harpersville Fm in Stephens County and the Archer City Fm in Clay County. Localities are lettered consecutively from A to T in Figure 1.
Fig. 1.

Locality map of Markley Fm collecting sites, showing general collecting area and outcrop extent of Markley, Archer City and Harpersville Fm. Collecting sites are labelled by letter (A–T). The USNM or USGS locality numbers for individual collections from each site are listed after the informally given name of the general collecting site. T – Kola Switch USNM 40069, 40070, 40071; USGS 9998; S – Malone Ranch USNM 42135; R – Turnbow USNM 39995, 39996, 39997, 39998; Q – 281 Road Cut USNM 40078, 40079, 40080, 40596, 40633; P – Voyles USNM 39989, 39994; O – Maxey USNM 40602, 40603; N – North of Markley USNM 40006, 40064; USGS 10164; M – Walker USNM 40007, 40086, 40087, 40088, 40595, 43529; L – Scott USNM 40641, 40687, 40688; K – Lycopod B USNM (East) 43546, 40081, 40682; (West) 40085; J – Gillespie Ranch (two localities) USNM (Ironstones) 40694; (not ironstones) 40607, 40608, 40609; (North) 43534; I – Gas Well USNM 40610, 40611; H – Squaw Mountain Coal Mine USNM 40632; G – Squaw Mountain Road Cut USNM 39990, 40002; F – Bloodworth USNM (Ironstones) 40003; (not ironstones) 40004, 40005, 40692, 40693, 43547; USGS 10166; E – Williamson Drive USNM 40012, 40013; D – Cooper USNM 39991, 39992, 39999, 40000, 40001, 40068, 43528, 43530, 43531, 43532, 43533; USGS 10162; C – Brannon Mine USNM 40084; B – Loving USNM (East) 40082; (West) 40083; A – Coal Mountain USNM 40612, 40613.
During the latest Pennsylvanian, the study area was located in the western Pangean tropics between 0° and 5°N (Scotese 1999).
Stratigraphic context of study
Stratigraphic correlation with the marine-equivalent Harpersville Fm indicates that the Markley Fm is composed of uppermost Pennsylvanian and lower-most Permian strata (Hentz 1988). Because of the paucity of fossiliferous marine beds in the Markley Fm, direct biostratigraphic correlation to a global standard is poor. However, Wardlaw (2005) identified the Pennsylvanian–Permian conodont boundary below the Stockwether Limestone of the Pueblo Fm, which has been lithostratigraphically correlated very close to locality Q (Fig. 1, Table 1), an exposure within the upper Markley Fm on US Highway 281 to the north of Jacksboro, Texas, USA.
Table 1.
Relative stratigraphic positions of localities sampled in this study, located relative to sandstones as mapped and numbered by Hentz (1988)
| Archer City Fm |
| Sandstone 8 |
| T-Kola Switch (Clay County) |
| Markley Fm |
| Sandstone 14 |
| S-Malone |
| Sandstone 12 |
| Q-US 281 Roadcut (Jack County) |
| Sandstone 11 |
| N- North of Markley (Young County) |
| O-Maxey Ranch (Jack County) |
| P-Voyles (Jack County) |
| Sandstone 10 |
| B-Loving (Young County) |
| D-Cooper (Jack County) |
| E-Williamson Drive (Jack County) |
| F-Bloodworth (Jack County) |
| H-Squaw Mountain Coal Mine (Jack County) |
| I-Gas Well (Jack County) |
| J-Gillespie Ranch (Jack County) |
| K-Lycopod B (Young County) |
| M-Walker (Jack County) |
| Sandstone 9 |
| L-Scott (Montague County) |
| Sandstone 8 |
| R-Turnbow (Montague County) |
| Sandstone 6 |
| C-Brannon Mine (Jack County) |
| G-Squaw Mountain Road Cut (Jack County) |
| Harpersville Fm |
| Sandstone 5 |
| A-Coal Mountain |
The Markley Fm was deposited in a fluvial-dominated coastal plain setting on the Eastern Shelf of the Midland Basin. Source areas for Markley Fm sediments include the Ouachita fold belt to the east and the Wichita–Amarillo and Arbuckle highlands to the north. Fluvial systems emptied into open marine depositional systems to the west and south, including the proximal Midland and distal Delaware basins (Hentz 1988). In the study area, Markley Fm strata have a regional NE–SW strike and dip c. 0.5°NW (Hentz 1988). The fluvio-deltaic strata of the Markley Fm transition towards the SW to the more marine strata of the Harpersville Fm (Fig. 1). Immediately above the Markley Fm lie the fluvial-dominated strata of the Archer City Fm (Fig. 1).
Collections of plant fossils were made from a range of stratigraphic positions throughout the entire Markley Fm, from portions of the laterally equivalent, marine-dominated Harpersville Fm, and from the lower Archer City Fm that overlies the Markley Fm (Fig. 1, Table 1).
Sedimentology
The Markley Fm is poorly exposed in the majority of its outcrop belt. Based on the characteristics of modern soils developed upon Markley Fm strata, it is likely that the formation is predominantly composed of fine claystones and mudstones (Hentz 1988; Tabor & Montanez 2004).
The best outcrop exposures are usually located on the flanks of erosional cuestas that are capped by fluvial sandstones of the Markley Fm (Hentz 1988). Outcrops are typically limited in lateral extent, ranging from a few tens of metres to c. 200 m (Figs 2 & 3). The majority of Markley Fm exposures consist of mudstones and sandstones, which were affected by pedogenesis in Permo-Carboniferous floodplain environments (Tabor & Montanez 2004) and typified by the Markley Fm stratotype section (Hentz 1988). Fossiliferous outcrops, however, differ from the stratotype section in that such fossiliferous strata appear to have been deposited in shallow abandoned channels, or as part of backfill sequences resulting from rising groundwater table or regional base-level, and were not subject to extensive pedogenic modification. The pattern of rock units in these channel-fill sequences is repeated throughout the Markley Fm and into the basal Archer City Fm, and is similar in its lithostratigraphic content to stacking patterns described in facies models for meandering streams (e.g. Nichols 2009). A schematic diagram of a typical fossiliferous sedimentary sequence and its facies associations in the Markley Fm is shown in Figure 4. Descriptions of lithologies and sedimentary structures, as well as interpretations of depositional environments of the facies, are provided in Table 2. Figure 4 is an idealized representation, of course, and between any two localities, minor differences in lithology and relative bedding thicknesses may occur or some lithologies/facies may be entirely absent. Despite these differences, the overall bedding organization at any locality follows the sequence illustrated in Figure 4.
Fig. 2.

Markley and Harpersville formation outcrops. Key: P, basal palaeosol; K, kaolinite-dominated siltstone; O, organic shale; F, coarsening upward interbedded siltstone–sandstone; S, channel sandstone. (a) Maxey Ranch, locality ‘O’; note thickness of basal palaeosol, prominence of quartz-kaolinitic siltstone, and weathered channel sandstone boulders. (b) Maxey Ranch, locality ‘O’, detail of erosionally resistant quartz-kaolinitic siltstone and overlying organic shale. (c) Maxey Ranch, locality ‘O’; note lateral continuity of beds. (d) Lycopod B, locality ‘K’; note both upper and lower coal intervals. (e) Squaw Mountain Coal Mine, locality ‘H’; note interbeds in transition between organic shale and floodplain mudstone. (f) Coal Mountain, locality ‘A’; note size of channel sandstone-dominated ridge in background.
Fig. 3.

Markley and Harpersville formation outcrops. Key: P, basal palaeosol; K, kaolinite-dominated siltstone; O, organic shale; F, coarsening upward interbedded siltstone–sandstone. (a) Cooper, locality ‘D’; note thick palaeosol and transition between organic shale facies and coarsening upward interbedded siltstone–sandstone facies, and blocks of channel sandstone at top of exposure. (b) Voyles, locality ‘P’; note dipping beds in coarsening upward interbedded siltstone–sandstone facies. (c) Loving, locality ‘B’; note both upper and lower coal intervals and channel sandstone in background. (d) Walker, locality ‘M’; note concave shape of beds in outcrop. Kaolinite-dominated siltstone bed is present directly below organic shale but not visible in photograph. (e) Bloodworth, locality ‘F’; note truncation of floodplain mudstone beds directly above the heads of the people in the photograph and large Fe-oxide concretions near their feet; dull orange beds at top of outcrop are Quaternary alluvium.
Fig. 4.

Idealized vertical profile of a single Markley Fm sedimentary sequence illustrating lithology, sedimentary structures and bed geometry. Vertical scale not listed due to variability in bed thickness (see Figs 2 & 3 for comparison of bed thickness variations).
Table 2.
Lithological descriptions and interpretations for the six facies represented at the majority of plant bearing outcrops in the Markley, Harpersville and Archer City formations
| Lithology | Interpretation | |
|---|---|---|
| Channel sandstone | Lenticular bodies of tan-reddish tan, fine to very coarse sand. Can be locally conglomeratic. Cross-beds present. Individual beds often pinch and swell in thickness at outcrop scale. Overall, this unit is virtually non-fossiliferous, with only a small number of identifiable plant fossils and unidentifiable stem fragments observed at a few localities. Sharp, erosional basal contact. |
Fluvial channel deposit |
|
Floodplain
mudstone |
Medium grey mud/silt. Weathers to tan-rusty red. Well to fair bedded, ripple laminated, often fissile. Charcoal fragments widespread, but sparse. Plant fossils may be abundant (sometimes covering entire cleaved surfaces of a hand sample) in some intervals and absent in others, with no observed lithological differences between fossiliferous and non-fossiliferous intervals. Basal contact typically indistinct, with interbedded organic shales gradually thinning and being replaced by this unit. |
Fluvial floodplain deposit |
|
Mudstone parting
beds |
Medium grey, poorly bedded clay. May contain charcoal fragments. Beds are thin (<10 cm) and are deposited within organic shale beds. Plant fossils are usually present, often in abundance. |
Fluvial channel deposit |
| Organic shale | Chocolate brown, organic-rich, fissile mud/silt. Weathers to rusty brown. Charcoal fragments (usually < 2 cm in diameter and <5 mm thick) widespread, sometimes in abundance. Bedding is very thin and planar. Coal deposits locally present, ranging from 1 cm to nearly 1 m thick. Coal is sub-bituminous and impure. Thicker coal beds often contain several interbedded shaly, clastic-rich intervals. Organic beds may be rich in fossil plant foliage, but may also be dominated by stem fragments, almost to the exclusion of foliage. Basal contact gradational for c. 2–5 cm from underlying grey clay to more organic-rich mud. |
Swamp deposit |
|
Quartzitic-kaolinitic
siltstone |
Light to medium grey, quartzitic silt/fine sand clasts in a kaolinite-rich matrix. At outcrop scale, bedding is clearly present; hand samples usually show poorly developed bedding. Plant fossils locally abundant or absent. Sharp basal contact. At outcrop scale, this unit is typically lens-shaped, indicating an erosional base or deposition in a topographic depression. |
Fluvial channel deposit |
| Basal palaeosol | Grey to reddish grey, mottled claystone, enriched in kaolinite and iron oxides. Has blocky structure. No bedding present. Colour differences are present in basal palaeosol outcrops, probably due to soil horizonation. This unit contains root traces and root casts, but no identifiable plant material was observed. This unit is identical to the ‘Type B paleosols’ described by Tabor & Montañez (2004). |
Palaeoultisol |
Material
Macrofossil collections and palynological slide preparations from the Markley Fm are housed in the Paleobotanical Collections of the United States National Museum of Natural History, Smithsonian Institution. At the request of property owners, locality information is limited to the general collecting areas rather than exact sites. The majority of the material surveyed has been collected over the past 20 years, although additional collections date back as far as the early 1900s. Selections of this older material were examined to gain a better understanding of the flora at a particular site, although this material was not analysed quantitatively.
Eighty-six separate macrofossil collections were analysed from the 20 localities shown in Figure 1, containing a total of 3544 hand samples. Collections outnumber localities because multiple collections were often made at a single locality, either from a number of stratigraphic intervals at that locality or as replicate collections in one particular bed.
Pieces of unoxidized matrix were removed from hand samples of macrofossils for palynological processing and analysis. Ten samples either did not contain palynomorphs or were too sparse for detailed floristic evaluation (Table 3). Samples from three localities (localities A, B and K; Tables 1 & 3) in the Harpersville and Markley formations between Sandstone 5 and Sandstone 10 of Hentz (1988) produced palynomorphs. Locality T in the Archer City Fm was the only other site included in this study to yield palynomorphs. Additionally, two suites of samples were collected directly from Markley Fm outcrop specifically for palynological processing, from localities A and K (Fig. 1, Table 3). Lithologies throughout the Markley Fm display a high degree of oxidation, so it is not surprising that many samples were unproductive. In addition, most of the outcrops are deeply weathered, causing post-depositional oxidation. Fresher matrix, for example, from drill cores, might prove to be more productive.
Table 3.
Locality numbers and codes for palynological samples processed
| Locality number | Lithofacies | DCA graph label | Locality name | Productive |
|---|---|---|---|---|
| USNM 42135 | F | Malone Ranch | − | |
| USNM 40069-A | K | T-13 | Kola Switch 9013 Lower Bed | + |
| USNM 40633-A | K | 281 Road Cut 9215 | − | |
| LycB90-1 | K | K-1 | Lycopod B East (Conifer layer) | + |
| LycB90-2 | K/O | K-2 | Lycopod B East (Cuticle layer) | + |
| USNM 40081 | K/O | K-2.5 | Lycopod B East (Bottom bed) | + |
| LycB90-3 | O | K-3 | Lycopod B East (Lower coal) | + |
| LycB90-7 | O | K-7 | Lycopod B East (Blocky mudstone) | + |
| LycB90-10 | O | K-10 | Lycopod B East (Blocky mudstone) | + |
| LycB90-12 | O | K-12 | Lycopod B East (Brown organic shale) | + |
| LycB90-13 | O | K-13 | Lycopod B East (Upper coal) | + |
| LycB90-14 | O | K-14 | Lycopod B East (Organic shale) | + |
| USNM 40083-A | O | B-1 | Loving West (Brown organic shale) | + |
| USNM 40612 (1991-15-3) | O | A-3 | Coal Mt. 9115A(Coal B) | + |
| USNM 40612 (1991-15A) | O | A-4 | Coal Mt. 9115A (Organic shale) | + |
| USNM 39989 | O | Voyles 8911 | − | |
| USNM 39994-5A | O | Voyles 9029 | − | |
| USNM 39991-A | O | Cooper 9010 (Brown organic shale) | − | |
| USGS 5665 | O | Newcastle Coal | + | |
| USNM 40602 | P | Maxey Ranch A | − | |
| USNM 40082 (Coal) | O | Loving East Coal | − | |
| USNM 40083 (Coal) | O | Loving West Coal | − | |
| MTS | U | Markley type section (Grey mudstone) | − |
+ , productive sample; −, unproductive sample. Key to lithofacies classification: F, coarsening upward interbedded siltstone and sandstone; K, quartz–kaolinite siltstone; O, organic shale; K/O, transitional between kaolinite-dominated siltstone and organic shale; P, mudstone parting beds; U, unclassified.
Palaeobotanical methodology
Collecting
The following is a detailed account of the collecting methods used during this study, together with the challenges, recognized biases and limitations encountered during collection of Markley Fm plant fossils. Some palynological samples were prepared from matrix removed from macrofossil samples, so the collecting methods used were the same. At localities A and K, samples were collected from distinct beds exposed in freshly dug trenches in the course of measuring outcrop sections. These details are necessary for the meaningful interpretation of our data and permit critical comparison of this study with other palaeoecological floristic analyses (Bennington et al. 2009).
No attempt was made to study a single outcrop exhaustively, as a major collecting goal of this study was to sample as many stratigraphic positions and lithofacies, and from as large a geographical area of Markley Fm exposures as possible. Although we recognize that such an exhaustive study could provide a more accurate assessment of relative abundances and diversity of taxa at that single site, practical limitations and the scope of the project prevented such an approach.
Quarry test pits were excavated into beds considered likely to contain plant fossils, followed by more extensive quarrying if warranted. Standard dimensions for a quarry were 1 m (width) × 0.5 m (depth) × 0.3 m (thickness), although these dimensions may be highly variable for a number of reasons: (i) fossiliferous deposits thinner than 0.3 m were excavated through the entire thickness of the deposit; (ii) fossiliferous deposits in some instances were highly localized and were excavated until plant fossils were exhausted; (iii) some outcrops were simply too difficult to quarry at desired dimensions and, in this case, the excavation was made as large as time and energy permitted; (iv) if several fossiliferous lithologies were distinct from each other, collections were then made on a bed-by-bed basis.
A collecting objective was to obtain a sample of plant macrofossils that accurately represented the proportional taxonomic composition of the deposit. Collections from different lithofacies were not mixed. All rocks on which plant fossils could be visually identified were examined, including those with only highly fragmentary debris. The collections housed at the NMNH usually number at least 50 hand samples, if possible, and may reach nearly 200 hand samples. This number varies, mainly due to logistical limitations encountered in the field. If fossils were scarce, then all specimens found were collected. If fossils were abundant, then a subset of what was found was collected. A minor bias was (or could have been) introduced at this stage by favouring rare taxa. A subobjective was to capture the broadest possible range of species diversity at a site. As a consequence, the rarest of taxa at a given locality may be over-represented (but given the generally low numbers of such specimens, this ‘over-representation’ will have little effect on the assessment of the quantitative floristic composition).
Different methods of field preparation were used, as dictated by the nature of the rocks. Using a rock hammer, fissile shales were cleaved and non-fissile rocks were fractured. Fissile shales were sometimes forcibly fractured to expose fossils not preserved on planes of weakness in the rock (see discussion of sampling issues below).
For a given lithofacies at a given locality, excavation of one or more replicate quarries was attempted. This was done to assess the spatial and temporal variability in fossiliferous deposits in an outcrop. Replicate quarries may have been excavated simultaneously, or during successive field seasons. Replicate quarries were either laterally equivalent to the original quarries (usually 1–2 m from the original quarry) or vertically displaced to some degree, depending on what aspect of variability was of interest. There were no replicates in instances where the contents of a fossiliferous deposit were exhausted during initial excavation and no equivalent deposits could be located. If, after examination, the fossil contents from the replicate quarry were determined to be virtually identical in diversity and type to the original quarry, a small voucher collection was made accompanied by a field count. In some cases, no voucher collection was made but a field count accompanied by photographs served as documentation.
Macrofossil quantification method and biases
Our sampling and collecting methods were chosen for their simplicity, efficiency and applicability to the wide range of fossiliferous deposits to be analysed. Some of these deposits, either by their general lithological nature or taphonomic biases, present challenges for any attempt to study their fossil contents quantitatively. The following paragraphs list a few notes on these sampling issues with the aim of clarifying the sampling method used in this analysis.
Determination of the quantitative abundances of various macrofossil taxa in a given collection was carried out using the hand sample quadrat method of Pfefferkorn et al. (1975). In this method, each hand sample is considered to be a sampling quadrat; size differences among these quadrats are minor and so do not affect the overall results. Counts of taxa were made by scanning the entire surface of a hand sample for identifiable plant fossil material. Whenever two samples were found to be a part/counterpart couple, only one of the duplicate surfaces was counted. All portions of the non-mirror-image surface of the counterpart were counted as a separate quadrat. When a given taxon was identified as present on a hand sample, it was counted only once for that quadrat. The total abundance of a taxon in a given collection, therefore, is expressed first as its occurrence on a specific number of quadrats, and then normalized to a proportion or percentage of the total number of quadrats in the collection. This method produces a frequency distribution because it is theoretically possible for all taxa to be present on every quadrat. In practice, this is not the case, and in tests of the hand-sample-quadrat method against point-count methods, rank-order abundance is preserved (Lamboy & Lesnikowska 1988; DiMichele et al. 1991). The quadrat method tends to raise the apparent abundance of rare taxa and suppress the abundance of abundant taxa, relative to a point-count analysis (Wing & DiMichele 1995).
Fragmentary preservation, fragmentation during collection, and obscured plant characters (due to rock colour, grain size, poor fossil/matrix contrast, weathering, burial in rock matrix, etc.) rendered much of the fossil material unidentifiable. Furthermore, although some species could be identified from small fragments due to highly diagnostic morphology, others required larger surfaces for positive identification. For example, to positively identify Cordaites leaves (which can be easily confused with finely striated axes – stems and rachises – belonging to many groups), a sufficiently large and/or finely detailed specimen was necessary. In contrast, Pseudomariopteris cordato-ovata or Nemejcopteris feminaeformis generally could be identified from a fragment of a pinna or pinnule. In addition, some species have a natural tendency to fragment into small bits, which are inherently difficult to identify, whereas others are naturally robust and tend to be well preserved. When a positive identification to species level was not possible, identification was made to the lowest taxonomic level possible. Beyond this (usually for the most poorly preserved specimens), the fossil was ignored. In either of the latter two cases, the lack of positive identification was grounds for not including the specimen in the final floral count. It should be noted that a positive identification, either to genus or species level, was possible for the vast majority of specimens.
Because of the taphonomic factors described above, certain groups (e.g. Cordaites) may be unavoidably under-represented in the floral counts of some collections. However, this sampling method minimizes the amount of bias that enters into a floral count of a sufficiently large sample because some identifiable specimens are usually encountered, which provides a ‘tip off’ that the group is present and thus sharpens the eye of the observer. This is most likely to be the case if the group is common to abundant in the collection. If a taxon was fragmentary in its preservation, difficult to identify from fragmentary specimens and present in very low abundance, the likelihood of ultimate identification was greatly reduced. In this kind of analysis, the basic dominance patterns are likely to be robust as reconstructed. However, total species richness is likely to be underestimated due to low identifiability of the rarest elements.
The degree of ease with which fossiliferous shales could be split to plant-bearing bedding surfaces introduced bias into the analysis. Most of the rocks collected were fissile shale, which naturally cleaves along planes of weakness. These planes of weakness are most common on surfaces with large plant remains. Therefore, broad, thin plant material (e.g. various kinds of axes, large pteridosperm foliage, etc.) is likely to be the most commonly exposed during excavation or subsequent splitting of larger excavated shale blocks, if such large material is present. Conversely, small or sparse plant compressions (e.g. isolated pecopterid pinnules, sphenopsid foliage, seeds, etc.) usually will not create a significant plane of weakness and may be under-represented on the rock surfaces analysed.
Palynological preparation and biases
Samples were subjected to 10% hydrochloric and 70% hydrofluoric acid maceration, followed by heavy liquid separation in zinc bromide (Specific Gravity = 2), carried out by Global Geolab Inc (Medicine Hat, Canada). Residues were sieved through a 10 mm filter, strew-mounted in glycerine jelly on glass slides, and examined using a Nikon 80i differential interference compound microscope with transmitted light.
Fourteen samples proved productive, including the Newcastle Coal, which, due to poor preservation, was not included in this analysis. A 200-grain count was performed on each sample (Table 3). More complete analyses will be presented elsewhere.
Palynology complements data from the plant macrofossil record, but presents its own potential biases. Although more susceptible to oxidative destruction than plant macrofossils, which can yield taxonomic information even if oxidized, palynomorphs when preserved are much more abundant than plant macrofossils and capture less common or more fragile taxa not preserved in the macrofossil record. Palynomorphs, being small and readily transported by water and wind, sample a broader source area than plant macrofossils, thus capturing a larger proportion of the total regional flora, including taxa from sites undergoing active erosion that otherwise would not be preserved in the fossil record. This quality can potentially introduce noise into facies/habitat distinctions, but, in practice, in fluvial terrestrial settings, the most common palynomorphs tend to be locally derived, that is from the vegetation surrounding the site of deposition. Palynological samples can be swamped by a single or few hyperproductive plant taxa (e.g. Marattiales); conversely, taxa with low or sporadic production may be under-represented or even absent, so relative or absolute abundance data are often a poor indicator of actual abundance of parent plants on the landscape. Most palynological samples (except those processed from a single depositional event, such as a lake varve) represent some degree of time averaging, so do not offer the temporal precision of single macrofossil assemblages. Palynological samples can and often do include reworked specimens, as rivers erode their banks and release previously deposited grains back into the system for redeposition. Reworked specimens often display a higher degree of physical damage or chemical weathering, and may display a higher (or lower) degree of thermal alteration, but can be physically indistinguishable from non-reworked (or only locally reworked) grains. Finally, taxonomic resolution for palynomorphs is generally less precise compared to plant macrofossils, because many palynomorph taxa lack any known macrofossil counterpart, although almost every pollen type can be ascribed to some biologically meaningful rank, for example seed plant or fern. Fortunately, a relatively high percentage of Pennsylvanian dispersed pollen and spore morphotaxa have been linked to a parent plant. It should also be noted that palynomorph morphospecies tend to be over-split, based on minor morphological, developmental and taphonomic variation. Consequently, overall diversity based on palynomorph morphospecies probably overestimates actual species richness.
Data analysis
Based on the original quadrat counts, abundance lists of dispersed macrofossil organ types in each collection were compiled. The original count data were condensed to create, as far as possible, ‘whole plants’ by combining different dispersed organs or by using only those organs that are most commonly encountered and most reliable for taxonomic purposes. So, for example, where a plant typically is represented by multiple dispersed organs, such as Sigillaria brardii, which, in addition to the characteristic stems, is represented by distinctive leaves, reproductive organs and deeper bark adpressions, all dispersed organs were combined under that one species. Pteridosperms and ferns, on the other hand, were overwhelmingly represented by foliage, with large amounts of stem and rachial axes, and minor (almost always quantitatively insignificant) occurrences of seeds or spore-bearing organs. The unidentifiable vegetative parts often could be combined with foliage when diversity consisted of a single species, or were divided proportionately based on the proportions of identifiable foliage. Disassociated reproductive organs generally could not be combined confidently with foliage, and in most cases were kept separate and not used in the quantitative analyses of taxonomic abundance. This approach is essentially the same as that used by Gastaldo et al. (2004), Cleal (2007) and Cleal et al. (2012).
Statistical analyses in this study were based on the condensed data matrix, after normalization. Exploratory data analysis was carried out by first examining the macrofloral composition of each lithofacies, with consideration given to both diversity and quantitative abundance of different species. Composition is shown graphically with species arranged in the same sequence on each plot (Fig. 5a, c, e, g), so that changes in composition can be readily identified. The order of the taxa in these plots is based on their scores on the first axis of a Correspondence Analysis (CA), which, although not shown separately, can be seen in the species order on the x-axis of the plots in question. CA is a multivariate ordination method designed for categorical variables; the analysis was carried out in CANOCO for Windows 4.5 (Ter Braak & Šmilauer 2002).
Fig. 5.


Histograms illustrating the pattern of abundance and dominance in Markley Fm macrofossils by facies type. Species order on the x-axis is the same in (a), (c), (e) and (g). In (b), (d), (f) and (h), species are ordered by dominance along the x-axis. Letters to the side indicate the facies type: K, kaolinite-dominat ed siltstone (18 taxa, sample size 356) ; F, coarsening upward, interbedded siltstones and sandstones (25 taxa, sample size 1210);P, mudstoneparting beds (16 taxa, sample size 696); O, organic shales (15 taxa, sample size 915).
In order to examine patterns in the distribution of dominants, dominance–diversity curves were prepared. These are simple bivariate plots of taxa ordered along the x-axis by relative abundance. Curves were created for each lithofacies type (Fig. 5b, d, f, h). ‘Importance’ is measured by relative percentage abundance, calculated from the original quadrat counts (number of quadrat occurrences in a sample/total number of quadrats in a sample × 100). These curves reveal with simplicity both the dominant groups in an assemblage, based on importance value, and the proportional distribution of abundance among the other taxa in a given assemblage.
Macrofossil absolute abundance data was also normalized to relative abundance data (as above). The converted data matrix was imported into R, a computer program designed for statistical computing and graphics (R Development Core Team 2010). Non-metric multidimensional scaling (NMDS) ordination was then performed on this data matrix to examine patterns of sample similarity. NMDS is a data analysis technique that uses quantitative models to describe qualitative data and display their structure as a geometrical picture (Young 1985). The technique is robust to non-normality of distribution, discontinuous data, non-linearity of relationships among variables, and questionable scales, and as such is a very robust method with which to explore data structure (McCune & Grace 2002). NMDS converts the relative abundance matrix to a dissimilarity matrix with n objects (in this case, n = 86 collections) on each axis. The dissimilarity between any two objects is based on their shared descriptors (in this case, species abundance). The objects are plotted as points in a multidimensional Euclidian space, the number of dimensions of which is prescribed. To simplify the display of data structure, two dimensions were used for this analysis. This was accomplished within the R program by projecting the points from a three-dimensional Euclidian space onto a two-dimensional plane. Arrangement of these points is based on their dissimilarity to each other so that proximity reflects degree of dissimilarity (i.e. highly dissimilar points will plot far from one another, and highly similar points will plot near one another). Table 4 lists the R command string used to compute the NMDS.
Table 4.
R command strings. Commands are in standard R notation; explanations follow # symbol
| File Preparation |
| > library(vegan) # Runs vegan package |
| >dd=read.csv(file=file.choose(), header=T) # Choose file to analyse |
| >dds<- dd[,6:33] # Choose columns to analyze |
| >row.names(dds)<- dd[,1] # Assign cells in column "1" as headings for their respective rows |
| NMDS |
| >w.mds<- metaMDS(dds) # Runs ordination |
| >w.mds # Performs NMDS using isoMDS (from MASS package) |
| >plot(scores(w.mds)) # Creates a plot of NMDS scores |
| ANOSIM |
| >markley.jacc=vegdist(dds, method= ‘jaccard’, binary=F,diag=F,upper=F,na.rm=F) # Calculates jaccard index |
| >gg<- dd$LITHOLOGY # Defines a grouping variable, in this case the grouping variable used is lithologic type |
| >aa<- anosim(markley.jacc, gg) # Performs ANOSIM |
| >plot(aa) # Creates a box-and-whisker diagram of ANOSIM results |
Confirmatory analyses were carried out on macrofossil data using the technique of analysis of similarities (ANOSIM) in R. Table 4 lists and explains the R command string used in this study to compute the ANOSIM. Further confirmatory statistical testing was not performed on palynological data because of small number of productive samples, limiting intersample comparisons. ANOSIM tests for significant dissimilarities between two or more predetermined groups of sampling units. Forthis analysis, fossil plant observations were divided into five groups based upon the lithology/facies in which they were found: upward coarsening mudstone–sandstone interbeds, Fe-oxide nodules (ironstones), kaolinite-dominated siltstone, organic shale and mudstone parting beds within organic shale. ANOSIM is based on a distance matrix whereby pairwise distances are first converted to rank value and then separated into within-group and between-group distances. The distance values define the level of dissimilarity between any number of objects: larger distances equate to higher dissimilarity, and vice versa. The dissimilarity values are displayed on a box-and-whisker diagram.
Palynological count data was the basis for a detrended correspondence analysis, carried out using PAST 2.12 software (Hammer et al. 2001). Correspondence analysis finds the chi-squared distances among all data points, yielding axes that account for maximum variability between points, where axis 1 accounts for the highest variance, and so on. Detrending is a form of normalization applied to data exhibiting a gradient by iteratively rescaling ordination axes to suppress ‘edge’ and ‘arch’ data artefacts generated by standard correspondence analysis (Hill & Gauch 1980; Oxanen & Minchin 1997).
Lithofacies characterization
Blocky to massive mudstone
Blocky to massive mudstones within plant-bearing successions are usually found at or near the base of outcrops and are composed of red-grey mottled claystones and siltstones (Figs 2 & 3, units labelled ‘P’; Fig. 4). Thickness is often indeterminate, depending on exposure of the profile base, and the maximum exposed thickness was 6.5 m. Sedimentary bedding structure is characteristically absent. These deposits weather rapidly, producing small, low gradient slopes (<30°). This lithology is friable, very weakly indurated, and typically organized into angular blocky structures in the range c.1–5 cm throughout the unit. The lithofacies typically has a red coloration, but may also have grey and yellow mottling that varies from rare (<2% of volume) to abundant (>20% of volume) and often includes nodular and vertically to horizontally oriented tubular Fe-oxide concretions that range from a few millimetres to several tens of centimetres across. The lithofacies can be subdivided into thinner layers, or horizons, based on changes in organization of the mudstone, in the amount of claysize material, in mineral composition with depth, and in abundance of concretions. This lithofacies has been interpreted as soil profiles that formed upon the coastal plains during Late Pennsylvanian time (Tabor & Montañez 2004; pedotype B). No identifiable plant megafossils were observed in this facies (with the important exception of rooting structures). This particular sort of palaeosol morphology is restricted to the lower Markley, that is, below Sandstone 10 of Hentz (1988; Tabor & Montañez 2004).
Tabor & Montañez (2004) note an abundance of Fe-oxide nodules in the lower portions of the profiles (BCg and Cgv horizons in their fig. 4b), together with evidence for translocation of kaolinite clay and Fe- and Mn-oxide into their inferred Bt horizon. They interpreted these findings to represent a palaeoultisol that was waterlogged for prolonged periods of time near the bottom of the soil profiles (based on the Fe-oxide nodules) yet experienced relatively mild seasonal changes in soil moisture (i.e. free drainage, based on the predominance of kaolinite in the clay fraction) in the upper parts of the profiles. Taken as a whole, these observations strongly support the interpretation of these deposits as palaeosols and suggest that they likely developed on low-relief interfluves during a period of low sedimentation rates or landscape degradation. Taken a step further, it would be reasonable to assume that the climate during development of these palaeosols was predominantly humid, with periods of seasonal drying.
Whereas the blocky to massive mudstone lithofacies is typical of the base of fossiliferous stratigraphic successions and provides an appropriate representation for some non-fossiliferous successions (e.g. Markley Fm stratotype section; Hentz 1988), other palaeosol morphologies occur within different lithostratigraphic facies associations in the Markley and lower Archer City formations (Figs 2 & 4; Tabor & Montañez 2004; DiMichele et al. 2006). We describe several different palaeosol morphologies in the subsequent discussion and note their occurrence(s) among the lithofacies associations used in this study (Figs 2 & 4).
A frequently encountered morphology is blocky greenish-yellow mudstone up to 2 m thick with abundant yellow to reddish-brown nodules that may include a layer of brown to dusky red Fe-oxide nodules and concretions in the upper half of the profile (Tabor & Montañez 2004; Pedotype C). Below the zone of Fe-oxide cementation the mudstone is organized into wedge-shape aggregate structures and has large, low-angle, slickenplanes. Tabor & Montañez (2004) interpreted these profiles to be similar to modern inceptisols, and considered these profiles to have formed under very poorly drained conditions based on redoximorphic features such as gley coloration and Fe-oxide nodules and concretions. However, the presence of slickensides and Fe-oxide nodules does suggest that profiles saw at least a small amount of seasonal drying and aeration. This particular morphology is limited to the upward coarsening mudstone–sandstone interbed facies. Such palaeosols likely formed within floodplain deposits proximal to fluvial channels, as evidenced by their close lateral and stratigraphic proximity to channel-filling and crevasse-splay sandstones (Tabor & Montañez 2004).
Another common palaeosol that occurs in abundance through the Markley and lower Archer City formations (Pedotype E; Tabor & Montañez 2004) ranges from 0.2 to 2 m thick and is composed of variable lithologies ranging from claystone, to muddy fine-grained sandstone, to sandstone, with massive to very weakly developed blocky structure. Fine-grained profiles commonly preserve carbonized fossil root compressions, as well as abundant and prominent colour mottling. In contrast, sand-rich lithologies have no carbonized root compressions and typically preserve only faint mottling and rare Fe-oxide nodules. Tabor & Montañez (2004) considered this morphologically variable group of palaeosol profiles to be similar to modern entisols, and noted that they are abundant throughout the Permo-Pennsylvanian continental strata of the eastern shelf of the Midland Basin. This particular palaeosol morphology has not been observed in the stratigraphic position of the blocky to massive mudstone lithofacies association. It does, however, occur commonly in the organic shale and mudstone parting beds, coarsening upward mudstone–sandstone interbeds, at the tops of the channel sandstone lithofacies and at the tops of kaolinite-dominated siltstone associations.
Pedotype D of Tabor & Montañez (2004) is common in the upper Markley (above Sandstone 11 of Hentz 1988) and lower Archer City formations. Such profiles are composed primarily of red to purple mudstone up to 1.3 m thick with common to abundant, coarse and irregular grey and yellowish mottles. The mudstone is organized into wedge-shaped aggregates ranging from 5 cm to several tens of centimetres across. Fe-oxide nodules and concretions commonly occur in the upper half of the palaeosol profiles. These palaeosols preserve slickenplanes in the lower half of the profile, and often preserve V-shaped dykes that extend from the top, down to the middle, of the profile. These dykes are typically filled with material that is similar in composition to the material in the stratum that overlies the profile. Tabor & Montanez (2004) considered these profiles to be morphologically and mineralogically similar to modern vertisols. The wedge-shaped aggregate structures, slickenplanes and clastic dykes are all indications of seasonal shrink–swell processes in soil profiles. This kind of morphology in modern tropical soil systems occurs in seasonally moist climates with four to eight dry months annually (Dudal & Eswaran 1988). Furthermore, Nordt et al. (2006) found that modern vertisols are non-calcareous (similar to the palaeosols considered here) only when mean annual precipitation exceeds 1400 mm a−1. Such high mean annual precipitation seems consistent with the abundant mottling and redoximorphic features in these profiles and suggests extended periods of soil saturation and relatively short dry intervals. These palaeovertisols are interpreted to have formed upon interfluvial surfaces of the coastal plain, and can occur either within the blocky to massive mudstone facies association (Fig. 2) or the coarsening upward mudstone–sandstone interbed facies association.
A final palaeosol morphology that we have not observed in the Markley or lower Archer City formations includes all of the morphological and mineralogical characteristics of a palaeovertisol, but also includes horizons with calcium carbonate nodules. The presence of calcite in this palaeosol type indicates lesser amounts of rainfall (Nordt et al. 2006) and longer and more severe intervals of soil moisture deficit than non-calcareous palaeosols, so the absence of these profiles is important from a palaeoclimate and palaeoecological perspective. Although Hentz (1988) noted calcareous palaeovertisols above Sandstone 12 of the Markley Fm, in 20 years of fieldwork we have not identified calcareous palaeovertisols below Sandstone 3 in the Archer City Fm, nor anywhere in the Markley Fm. This suggests that significant drying and greater seasonality across landscapes existed in the Eastern Midland Basin after deposition of Archer City Fm Sandstone 3, and importantly, subsequent to the interval in which the fossiliferous stratigraphic sequences discussed herein were deposited.
Kaolinite-dominated siltstone lithofacies
One or more beds of siltstone composed mainly of kaolinite and quartz, with variable amounts of other clays, mainly chlorite and illite, are present in nearly all of the fossiliferous outcrops studied, most often positioned immediately above the basal palaeosol (Figs 2 & 3, labelled ‘K’; Fig. 4). Kaolinite-dominated siltstone beds also occurred at the base of scours within the stacked mudstone–sandstone lithofacies at a few sites. Such beds are laterally extensive on outcrop, variable in thickness from 10 to 80 cm, and have trough-shaped erosional basal contacts with the underlying facies. In lithological composition the kaolinite-dominated siltstone beds are very similar to the basal palaeosol facies. The most striking difference between these two facies is that the kaolinite-dominated siltstone deposits lack the pedogenic features found in the underlying palaeosols and instead show evidence of fluvial deposition, such as an erosional and scoured basal contact, poor, yet distinct sedimentary bedding, and localized deposits of identifiable plant fossils that often crosscut bedding surfaces.
The plant fossils in this facies are xeromorphic and belong to evolutionary lineages broadly interpreted as centred in subhumid to semi-arid habitats. The only evidence for soil formation in this facies is found in some instances near the top of the unit, where bedding disruption or bioturbation by rooting rarely occurs.
Overall, the kaolinite-dominated siltstone facies appears to have been formed by rapid deposition of fine-grained, highly weathered sediments, possibly as a slurry of mud and fossil plant fragments. Because this facies is so lithologically and mineralogically similar to the underlying basal palaeosol facies, it is reasonable to speculate that its sediments were derived from local erosion of the surface layers of the underlying palaeosols from an adjacent, slightly elevated source, although this cannot be confirmed. Romanchock et al. (in press) compared these deposits to the so-called ‘ball clays’ of Eocene age from the southeastern USA (Elsik & Dilcher 1974; Potter 1976; Potter & Dilcher 1980), which are typically confined to abandoned floodplain channels, and are thought to form in hot, humid climates.
Organic shale lithofacies
Organic-rich lithofacies range from finely laminated shales with a high organic content to locally developed coal beds with high ash content (Figs 2 & 3, labelled ‘O’; Fig. 4). The shale beds may contain extremely dense deposits of fossil plant material and coaly/charcoal fragments. Shales are fissile, low in density and extremely friable, yet are erosionally resistant, often forming low escarpments on outcrop. Overall, the organic shale deposits typically range between 2 and 3 m thick, with individual organic-rich beds ranging from 10 cm to nearly 2 m in thickness. At a few localities an impure coal up to 0.8 m thick was observed within the organic shale deposit. Thinner (5–20 cm), laterally discontinuous coals are present at many other localities. Fe-oxide nodules of various sizes occur within this lithofacies at a few localities, interspersed throughout the organic shale deposits in varying concentrations. These organic-rich deposits probably formed in a low-energy environment with low clastic input, such as a swamp.
Thin (<10 cm), medium-grey, clay-rich beds are locally present in the organic shales. These beds, which are referred to in this paper as mudstone parting beds, are lens-shaped on outcrop. The mudstone parting beds, based on their small size, limited extent and fine-grained composition, are interpreted as channels that periodically carried clastic sediments into the swamp environment.
Organic accumulations of this nature, especially trending to impure coals, require a humid climate and persistent standing water conditions for most of the year to retard decay and permit the accumulation and preservation of peat deposits (Cecil & Dulong 2003; DiMichele et al. 2006; Greb et al. 2006). Furthermore, a humid climate promotes the thick growth of vegetation necessary to stabilize the landscape and severely limit the erosion and transport of sediments into the fluvial system. Vegetationally controlled inhibition of sediment flux, as a secondary consequence of humid climatic conditions, helps explain the relatively low quantitites of clastic sediment in the organic shale facies, permitting the concentration of organic matter. This is especially relevant considering the lack of evidence for change in landscape elevation or topographic slope, and the repetitive appearance of this facies at different stratigraphic levels, but in the same position relative to the other facies.
Coarsening upward mudstone–sandstone interbed lithofacies
The organic shale facies is directly overlain by a coarsening upward sequence of interbedded mudstone and sandstone (Figs 2 & 3, labelled ‘F’; Fig. 4). The contact between these two facies may be sharp and erosional or gradational. In most of the instances we observed, this facies association can be artificially separated into three sections: a ‘bottom’ transitional section consisting of interbedded thin layers of organic-rich shale and mudstone, a ‘middle’ section consisting of mudstone and interspersed weakly developed palaeosols (types C, D and E described above), and an ‘upper’ section that is composed of interbedded mudstones and sandstones that collectively define an upward coarsening sequence of sedimentary strata beneath the channel sandstone lithofacies. The predominant lithology is grey-to-tan coloured, clay-and-silt rich, mudstone and fissile shale with low-to-moderate organic content, sometimes containing charcoal fragments. Beds are thin (0.5–10 cm) and discontinuous, lenticular in shape, and often ripple-laminated, giving them an undulating surface. At outcrop scale, packages of beds frequently truncate one another.
The various bed geometries and sedimentary features mentioned above suggest that the lower part of the upward coarsening mudstone–sandstone interbed facies was deposited in shallow channels within broad, low-profile channel belts.
This depositional pattern continues upward into the middle section where the organic shales disappear, the mudstones take on a more uniform lithological character, and palaeosols begin to appear within the unit. These palaeosols are angular and blocky in texture and exhibit various levels of development (Pedotypes C, D and E, discussed above). Most are thin and only slightly obscure the original sedimentary structure of the mudstones, resembling immature palaeoentisols (Pedotype E discussed above). Others are thicker and more mature, sometimes including blocky or wedge-shaped mudstone clasts and containing a horizon of densely packed Fe-oxide nodules, resembling palaeoinceptisols (Pedotype E discussed above) and palaeovertisols (Pedotype D discussed above). The upper surface of any of these immature palaeosols represents a hiatus in deposition and implies the presence of significant diastems. This middle section also contains plant fossils, which are found in small, localized lenses of sediment that are often trough-shaped and appear to have erosional basal contacts, possibly originating as scour surfaces. The enclosed plant fossils are sometimes preserved along bedding planes but often crosscut bedding planes suggesting rapid rates of sediment deposition.
Within the upper section, the floodplain mudstone facies is replaced by channel sandstone facies. This transition may happen at a single lithological contact or as a series of interbeds of mudstone and sandstone of up to 3 m in thickness that is similar in appearance to crevasse-splay deposits emplaced on floodplains by overbank flooding of meandering streams (e.g. Nichols 2009).
Based on the fine-grained nature of the sediments, frequent depositional hiatuses and overall coarsening-upward profile, deposition of the upward coarsening mudstone–sandstone interbed facies likely occurred in a low-energy, aggrading (i.e. regressive), coastal plain setting. Furthermore, the overall coarsening upward pattern in this lithofacies is consistent with a temporal transition from an initially distal floodplain site to one that was proximal to meandering river channels (e.g. Walker & James 1992).
Channel sandstone lithofacies
Sandstones are present at the uppermost portion of nearly all outcrops studied, either as intact beds or weathered blocks of talus material that are now scattered across the outcrops (Fig. 2a, labelled ‘S’; Fig. 4). The sandstones cap outcrops because they are more resistant to erosion than the fine-grained mudstones. The channel sandstone lithofacies is characterized by erosional basal contacts with the coarsening upward mudstone–sandstone interbeds, which, where visible, are broadly U-shaped in cross-section. Overall unit thickness varies from a few metres to a few tens of metres, although accurate thickness measurements are complicated by modern weathering that destroyed or buried the uppermost portions of sandstones. Their internal geometries exhibit extensive pinching and swelling of individual beds, with bed thickness ranging from 0.5 m to less than 5 cm, and trough cross-bedding. Grain size ranges from fine to very coarse sand, with chert, quartz and Fe-oxide pebble conglomerates locally present at the base of trough-cross bedded units.
Plant fossils were very sparse or absent from this facies at most localities examined. In the few instances where plant fossils were present, Cordaites sp., unidentifiable axes and rare Pecopteris sp. were most common. Perhaps most interestingly within the scope of this study, the conifer Walchia was also observed at one locality (locality K); the only other occurrences of this taxon within Pennsylvanian strata are in the kaolinite-dominated siltstone facies (Walchia becomes common in younger Permian strata). Because of the scarcity of fossils, a meaningful quantitative analysis of the channel sandstone facies flora is not possible.
It is likely that these deposits fill channels that were once part of larger stream networks. Channel cutting and sand deposition were possibly contemporaneous with the development of soils on interfluve surfaces and may account for the floristic similarity between the channel sandstone and kaolinite-dominated siltstone lithofacies, if the kaolinite-dominated siltstones were derived from erosion of soil surfaces.
Temporal trends in depositional patterns
At the temporal scale of the entire Markley Fm deposition, facies recur in distinct packages, as discussed above. Yet there is a long-term trend of differential facies abundance through the Markley Fm most marked by the diminishment, and ultimately the disappearance, of organic-rich beds. In the lower Markley Fm, for example, virtually every locality from which plant fossils were collected contains deposits of organic shale, often of considerable thickness (up to 1.5 m). Progressing stratigraphically upward through the Markley Fm, these organic shale deposits decrease in number, extent and thickness until they become virtually absent from the uppermost Markley Fm and the overlying Archer City Fm. In the basal Archer City Fm, most plant-bearing shales occur in channel-fill facies on outcrops of less lithological complexity than typical of the Markley Fm. In fact, the characteristic sedimentary cycles of the Markley Fm disappear almost entirely and are replaced by a new sedimentary pattern consisting mostly of well-developed palaeosols, floodplain or channel-fill mudstones, and fluvial sandstones.
The disappearance of the organic shale–coaly facies, while at the same time the other typical Markley facies persist in sequence, strongly suggests a climatic trend toward long-term drying, or at least increased seasonality of precipitation whereby productivity and preservation of plant organic matter was increasingly limited. While one may argue that this lithological shift could also have resulted from a steepening of the topographic gradient that would have prohibited the formation of organic-rich swamps, it is inconsistent with a long period of tectonic quiescence in this region during and after deposition of the Markley Fm (Hentz 1988). Although basin subsidence did continue during this time, the sediment influx was suffcient to maintain a nearly level coastal plain with a shoreline well to the west. Further support of the inferred long-term drying trend comes from other sedimentological indicators of dry climate in formations younger than the Markley. These include widespread development of soil carbonate and gypsum nodules in the upper Bowie and Wichita groups (Tabor & Montañez 2004; Tabor et al. 2008), a complete lack of both organic-rich palaeosol profiles (Histosols; coal) and deeply weathered palaeoultisols indicative of humid climates, as well as marine beds of gypsum in the Wichita Group ranging from 3 to 10 m thick (Jones 1971; Jones & Hentz 1988).
Results of macrofossil palaeobotanical analysis
Dominance–diversity curves
Patterns of composition and dominance and diversity for each of the four lithofacies are illustrated in Figure 5. Each of these curves shows the full list of 28 taxa identified in the Markley Fm. However, the number of taxa identified in each facies differs considerably. The mudstone–sandstone interbed lithofacies includes 25 taxa, whereas the other facies encompass between 15 and 18. In addition, the mudstone–sandstone interbed lithofacies includes many relatively abundant taxa that are rare to absent in the other facies, such as Pseudomariopteris cordato-ovata, Neuropteris ovata, Asterophyllites equisetiformis, Neurodontopteris auriculata and Sphenophyllum oblongifolium.
Species dominance patterns in the coarsening upward mudstone–sandstone interbed facies are different from those of the other three facies. In the mudstones of the coarsening upward mudstone–sandstone interbed facies, no species occurs on more than 20% of the sampling quadrats, giving this facies the most even distribution of dominance among the four fossiliferous lithofacies types. Even so, eight or nine species are present at a frequency of 5% or more of the sampling quadrats in each of the four fossiliferous facies except for the organic shales, at 6 species. However, the organic shale, parting bed and kaolinite-dominated siltstone facies were more uniform from site to site than the coarsening upward mudstone–sandstone interbed facies. Furthermore, the dominance–diversity curves for each facies, which are composites of all sample sites from a given facies, are generally similar from site to site in all but the coarsening upward mudstone–sandstone interbed facies, where very high dominance by one species may occur within any local assemblage despite an overall average uniformity of species abundances among the sampling sites. These observations, together with field observations of the rocks, suggest that the coarsening upward mudstone–sandstone interbed facies was environmentally heterogeneous. Much like today, Late Pennsylvanian floodplains, which we presume were the basic depositional settings of the coarsening upward mudstone–sandstone interbed facies, were subject to high levels of disturbance and supported spatially heterogeneous, riparian plant communities.
The three remaining lithofacies are all clearly dominated by a single species. The organic shale and mudstone parting bed facies are dominated by Macroneuropteris scheuchzeri (present on 47% and 46% of quadrats, respectively), whereas the kaolinite-dominated siltstone facies is dominated by Sphenopteridium manzanitanum (present on 57% of quadrats). In each of these three facies, the second ranked species was identified on 14–23% of quadrats. Although the shapes of the dominance–diversity curves are similar for these three facies, the quartz kaolinitic siltstones have a very different taxonomic composition from the organic shale and mudstone parting bed facies. High dominance by a single species is consistent with a standing-water swamp interpretation for the physical habitat of the organic shales and mudstone parting beds. Relatively low species richness and high dominance by one or a few species are well documented for clastic and peat-swamp settings in the Pennsylvanian (e.g. Gastaldo et al. 2004; DiMichele et al. 2007; Opluštil et al. 2007, 2009a, b). Conversely, the limited floral diversity in the kaolinite-dominated siltstone facies may reflect the fact that this flora is transported to some extent. Low species richness may reflect a low diversity of trees on the landscape immediately surrounding the environment of deposition from which the flora is most likely drawn, a conclusion supported by modern actualistic studies of plant taphonomic patterns (e.g. Scott 1978; Scheihing & Pfefferkorn 1984; Burnham & Spicer 1986; Burnham et al. 1992). Significant taphonomic selection against plants farther from the edge of the deposit and against ground cover plants has been documented. In addition, low abundances and diversity of small, ground cover plants in fossiliferous assemblages is expected in most allochthonous depositional settings, even when the environment of deposition is immediately adjacent to the source vegetation (Scheihing 1980).
Non-metric multidimensional scaling (NMDS)
NMDS ordination of all 86 macrofossil collections (Fig. 6) reveals patterns consistent with expectations based on observations of species occurrence and dominance within the different lithofacies. Points in Figure 6 are labelled by their respective lithofacies. Recall that each point is a single excavation characterized by its quantitative taxonomic composition, determined to the lowest taxonomic rank possible.
Fig. 6.

Non-metric multidimensional scaling (NMDS) ordination of all 86 macrofossil samples. Points are labelled by facies type. Each facies type is outlined by a continuous line. Axis orientation is arbitrary and axis units represent NMDS scores. Key: F, coarsening upward interbedded siltstone–sandstone; I, Fe-oxide nodules (ironstones); K, kaolinite-dominated siltstone; O, organic shale; P, mudstone parting beds; S, channel sandstone.
The most obvious and significant pattern is the clear emergence of clusters, based on species composition, that conform to lithofacies groups. The pattern indicates that species composition has the strongest effect on the distribution of points (samples) within this ordination, and that floral composition is tied strongly to lithology/habitat of growth. Conversely, in this two-dimensional plot there is no clearly interpretable signal derived from the stratigraphic position or geographic location of a given point. In fact, there is no stratigraphically detectable pattern in the data. The ordination pattern is, then, in agreement with field and laboratory observations that the major pattern in the data is one of species by lithology, and supports niche-based models of species distribution.
The kaolinite-dominated siltstone facies sample suite does not overlap with samples of the organic shale facies (Fig. 6), consistent with the strong differences in species content of these two facies. In contrast, the suite of organic shale samples and that of the mudstone parting bed facies overlap one another nearly entirely (Fig. 6). This indicates a common species pool and is consistent with field observations of the position of the mudstone parting beds within organic shale deposits.
The area of the ordination encompassed by any given facies cluster reflects the degree of similarity among the samples of that facies. For example, the cluster of organic shale samples (n = 24) covers a relatively small area, consistent with Macroneuropteris scheuchzeri and/or Pecopteris abundance in all of these samples. In contrast, the coarsening upward mudstone–sandstone interbed facies samples (n = 25) are nearly twice as dispersed as the organic shale sample suite. Qualitative observations made during sampling, and dominance–diversity curves based on quantitative analysis, indicate that the coarsening upward mudstone–sandstone interbed facies flora is often diverse at any one site and spatiotemporally variable among sites. In contrast, organic shale samples have low richness at any given site and vary little from site to site.
A sample suite of Fe-oxide nodules (ironstones) is included in Figure 6 (red squares (grey in print version), not encompassed by a circumferential line) to show the floral variability of this collection. These samples do not form a coherent cluster but instead overlap with all other facies clouds. These nodules formed in – and were collected from – various facies, indicating that they are a taphonomic feature without vegetational or floristic significance. Locally, the ironstones often preserve plant impressions in exquisite detail and so are of great value in characterizing the flora of those deposits in which they occur.
A single sample representing the only quantifiable channel sandstone facies collection is also included in Figure 6. Whereas a single collection from this facies is perhaps of limited significance, it is interesting to note that this point is located among peripheral samples (filled orange dot (grey in print version) on the right side) of the coarsening upward mudstone–sandstone interbed facies cloud (marked by blue (grey in print version) diamonds), which are interpreted to have been deposited upon floodplain sites.
The influence exerted by five of the most abundant species on the NMDS cluster pattern was assessed by plotting the abundances of these taxa on five separate NMDS-based ordinations (Fig. 7). These subsets reveal those collecting localities (points) that contain an abundance of that particular taxon. The cutoff for abundance varied for each taxon, and was chosen to enhance the visual aspect of the pattern by maximizing the number of points while simultaneously maximizing their concentration on the NMDS plot.
Fig. 7.

NMDS-based plots for selected dominant macrofossil taxa, indicated by taxon label. Percentage values indicate relative abundance cutoff values used for that particular plot. Orientation of axes is arbitrary and axis units represent NMDS scores. Key: F, coarsening upward interbedded siltstone–sandstone; I, Fe-oxide nodules (ironstones); K, kaolinite-dominated siltstone; O, organic shale; P, mudstone parting beds.
These plots (Fig. 7) demonstrate the strong effect of dominant taxa on the NMDS pattern. For example, the Macroneuropteris scheuchzeri cluster (n = 24) is the main influence on the overlap between floodplain mudstone, organic shale and mudstone parting bed facies. Similarly, Sphenopteridium manzanitanum (n = 11) and Walchia sp. (n = 7) account for the distinctiveness of the kaolinite-dominated siltstones, but their rare occurrences in deposits of the coarsening upward mudstone–sandstone interbed facies are major factors in creating a zone of overlap between these two facies clusters. Pseudomariopteris cordato-ovata (n = 4), in contrast, is conspicuously abundant only in a subset of samples from the coarsening upward mudstone–sandstone interbed facies. High abundance in each of these four taxonomic groups is relatively tightly clustered in the NMDS. The opposite pattern is found in pecopterid ferns (n = 17), which do not form a discrete cluster and have a high abundance in a variety of facies types, occurring in many collections of the coarsening upward mudstone–sandstone interbed, organic shale and mudstone parting bed facies. These findings are consistent with a high degree of habitat specificity for many of these taxa. Species such as M. scheuchzeri, S. manzanitanum and P. cordato-ovata appear to have high habitat specificity towards wetland swamps, well-drained soils and wet flood plains, respectively. Pecopterid ferns, on the other hand, are common in species pools of both floodplain mudstones of the coarsening upward mudstone–sandstone interbed facies and organic shale–mudstone parting bed facies. Pecopterid ferns may be a very complex group in terms of both species diversity and ecological tolerances, but this is difficult to clarify because the taxonomy of pecopterids is refractory (e.g. Knight 1985; Pšenička et al. 2009), particularly where specimens are preserved mainly as small fragments, as in the Markley Fm samples (Romanchock et al. in press). As a group, pecopterid marattialean ferns appear to have been able to grow in a wide range of landscapes as long as they were able to locate even small areas of sufficiently wet substrates on those landscapes (DiMichele & Phillips 2002; DiMichele et al. 2006).
Analysis of similarities
ANOSIM (Fig. 8) compares the degree of dissimilarity within prespecified groups to the amount of variability between those groups. It calculates the statistic ‘R’ to determine if these dissimilarities are significantly different. In the case of the Markley Fm data, it is a test of the hypothesis that there are distinct lithofacies-based assemblages, not an exploratory analysis.
Fig. 8.

Analysis of similarities (ANOSIM) plot showing dissimilarity between and within facies. Bold horizontal bar in box indicates median; bottom of box indicates 25th percentile; top of box indicates 75th percentile; whiskers extend to the most extreme data point, which is no more than the range (i.e. 1.5) times the interquartile range from the box; width of bar directly proportional to sample size; grey-shaded boxes highlight notch width; if the notches of any two plots do not overlap this is ‘strong evidence’ that the two medians differ (Chambers et al. 1983, p. 62). The ANOSIM statistic R is based on the difference of mean ranks between groups (r_B) and within groups (r_W), such that: R = (r_B − r_W)/(N*(N − 1)/4). Key: Between, between-group dissimilarity; F, coarsening upward interbedded. siltstone–sandstone; I, Fe-oxide nodules (ironstones); K, kaolinite-dominated siltstone; O, organic shale; P, mudstone parting beds.
In this analysis, the lithofacies-specific floras formed the predefined groups. The most significant aspect of this test is the between-group dissimilarity. The test was carried out with and without the Ironstone samples and a similar result was obtained in each analysis; only the with-Ironstones results are reported here.
The median between-group dissimilarity is higher than the dissimilarity within any individual lithofacies group, except for ironstones (which are not facies-specific and represent floras from several different lithofacies). This indicates that the dissimilarity within any facies is significantly less than that among facies. Thus, the ANOSIM is consistent with the patterns seen in the NMDS and other qualitative observations. The kaolinite-dominated siltstone, organic shale and coarsening upward mudstone–sandstone interbed facies have relatively low within-group dissimilarities. In contrast, the coarsening upward mudstone–sandstone interbed facies has a much higher level of dissimilarity, again consistent with NMDS and visual inspection of collections, indicating high site-to-site variability.
Results of palynological analyses
Of the five lithofacies, only two – organic shale and kaolinite-dominated siltstone – were palynologically productive (Table 3). These two lithofacies contrast in palynomorph composition, largely but not completely matching the pattern observed in the plant macrofossil record.
Composition and dominance patterns
The organic shales, broadly speaking, are dominated by spores of Marattiales, Sigillaria and other lycopsids (e.g. Chaloneria), as well as sphenophylls and calamites (Fig. 9). Microspores of Sigillaria appear more abundant in the organic-rich clastic units, whereas the coals appear to be dominated by spores of Marattiales and sphenophylls; however, sample size was insufficient to support this impression statistically. The palynological assemblage of the organic shale facies displays a greater degree of evenness of species in quantitative composition when compared to the dominance of Macroneuropteris in the macrofossil records. The spores of non-marattialian ferns display the greatest species richness, although not necessarily abundance, in this facies. Significantly, pollen of walchian conifers and seed ferns is rare but consistent in samples observed from the organic shale lithofacies (abundance, 0.5%).
Fig. 9.

Taxonomic composition and relative abundance of major taxonomic groups in palynological samples. Two lithofacies are represented: kaolinite-dominated siltstones (K) and organic shales (O), with two transitional samples (K/O). Labels of palynological samples as in Table 3.
Certain macrofossil taxa are poorly represented in the microfossil record and, conversely, many microfossil taxa lack obvious counterparts in the macrofossil record. The most notable incongruence is the very low abundance of Schopfipollenites, the pre-pollen of Macroneuropteris and other medullosan seed ferns (Alethopteris, Neurodontopteris, Neuropteris). Reasons for this are unclear, but may include size sorting in the depositional process, because of the large size of Schopfipollenites pre-pollen (up to 500 μm in length). Breakage during deposition or sample processing may be another factor, because Schopfipollenites sometimes occurs as fragments in strew mounts, as well as lower pollen output. In contrast to the scarcity of medullosan pre-pollen, cordaitalean pollen is abundant in the organic shale lithofacies, especially in the clastic phases, compared to the rarity of Cordaites leaves. This discrepancy is likely due to the difficulty of recognizing Cordaites leaf fragments in the macrofossil record, as noted above.
The palynological assemblage composition of the kaolinite-dominated siltstone lithofacies is also largely congruent with the macrofossil data. The most abundant pollen type in this facies is one or more species of non-taeniate bisaccate, or pseudobisaccate forms such as Falcisporites, Alisporites or Pseudovesicaspora. It is tempting to attribute one or more of these forms to the seed fern Sphenopteridium manzanitum, which is dominant in this lithofacies; however, there is no direct evidence for this. Pollen of walchian conifers (Potonieisporites and Nuskoisporites) is subdominant, along with spores of sphenophylls and marattialian ferns. Dominance–diversity curves for palynomorphs largely follow those of the macrofossils, with the exception of the virtual absence of medullosan pre-pollen (Fig. 5). Lycopsid spores and cordaitalean pollen are rare in this lithofacies, as are spores of Sigillaria. Marattialian fern spores display low to moderate abundance, consistent with the presence of macrofossils of this clade in many environments. A few non-marattialian ferns are sometimes abundant in this lithofacies, but their overall species richness is much lower than observed in the organic shale lithofacies.
The palynofloras from both facies are notably more species-rich than the respective macrofossil assemblages. Roughly 110 palynomorph morphospecies were recognized in the 13 productive samples, compared to 28 in the macrofossil flora. Per sample diversity is also much higher in palynological assemblages, with a mean of 30 (ranging from 19 to 42). Some of this apparent diversity may be due to taxonomic oversplitting as noted above, but the greater part of this signal is probably real. The greatest diversity occurs within the non-marattialian ferns, with at least 20 distinct morphotypes represented, many of unknown affinity. These ferns, including members of the Tedeleaceae, Zygopteridaceae, Gleicheniaceae, among others, never comprise more than 15% of the total flora, more usually ranging from 2% to 5%, but are always present at some level. The palynological assemblages may be more diverse, in part because of time averaging, reflecting a temporal succession of floral elements represented in the macrofossil record by a single time slice, or because they are drawing from a broader source area, or probably both.
Ordination analysis
Detrended correspondence analysis of palynological abundance data (Fig. 10) reveals more overlap between the organic shale and kaolinite-dominated siltstone lithofacies than is apparent in the macrofossil samples. Axis 1 of the graph is interpreted as a wet-dry gradient. Axis 2, on the other hand, manifests primarily variation within the organic-shale facies, and appears to reflect abundance of Sigillaria microspores; this may indicate a subtle wet–dry gradient within that facies. Samples from the organic shale lithofacies (palynological samples B-2, A-3, A-4, K-3, K-7, K-10, K-12, K-13 and K-14) group to the left of the graph, presumably on the wet end of the spectrum, whereas the two macrofossil-bearing kaolinite-dominated siltstone beds (palynological samples K-1 and T-13) group to the right, showing a clear disjunction in palynoflora composition between these two lithofacies types. However, two samples between the kaolinite-dominated siltstone bed and the basal coal at locality K (palynological samples K-2 and K-2.5) are transitional in palynofloristic composition between the wetland and dryland floras (Fig. 10). The lithology of samples K-2 and K-2.5 becomes increasingly rich in organics and plant debris upsection. Both units contain abundant conifer and non-taeniate bisaccate pollen, together with an almost equal abundance of marattialian and non-marattialian fern, sphenopsid and lycopsid spores (Figs 9 & 10). Significantly, walchian twigs occur just below the basal coal in one of these layers. There is no evidence of a sedimentological break or hiatus between kaolinite-dominated siltstone lithofacies and the organic shale beds at locality K.
Fig. 10.
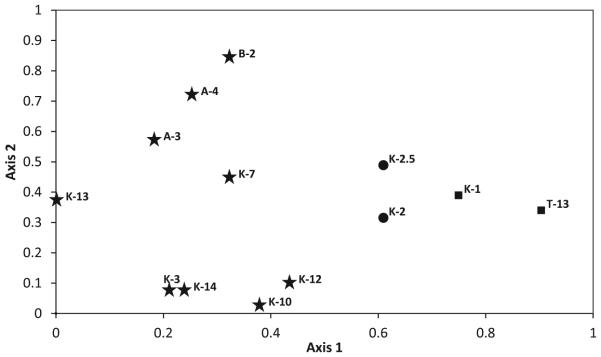
Palynological site detrended correspondence analysis (DCA) based on count data of 109 taxa from 13 localities. Lithofacies are either organic shales (stars, n = 9), kaolinite-dominated siltstones (squares, n = 2) or transitional between these facies (circles, n = 2) (see also Fig. 4). Locality letter designations as in Table 3. Eigenvalues for Axis 1 and Axis 2 are 0.7599 and 0.3258, respectively.
Discussion
Evidence of niche-based assembly dynamics
Perhaps the most significant specific finding of this study is that species content and relative abundance in a given facies are remarkably similar regardless of geographic location or stratigraphic position within the Markley or basal Archer City formations. These patterns of niche conservatism are superimposed on a stratigraphic interval of uncertain duration, but likely encompassing several hundred thousand to perhaps as many as a million years. This duration is estimated based on the several distinct intervals of peat formation separated by periods of palaeosol development and marine incursion. This cyclic repetition of lithofacies packages is evidence of recurrent base level and likely climate change, in addition to the documented ecological changes. This means that a high degree of ‘niche assembly’ is called for in the face of changing landscape conditions within which certain very specific habitats (represented by the lithofacies types) periodically reappear.
Thus the species that comprise these different habitats do not appear to be randomly distributed but instead have strong preferences for physical conditions that reflect amount and duration of soil wetness, degree of disturbance, and possibly amount of light availability. Although a few species are present in more than one habitat, each habitat contains a unique suite of plants and dominance distribution, whereas species richness is roughly comparable. So strong are these habitat relationships that they permit statistical differentiation among the facies solely based on taxonomic composition and quantitative abundance. The strength of these conclusions about flora/facies relationships can certainly be tested in other parts of the Late Pennsylvanian (Stephanian) tropics, given the widespread distribution of these basic species pools (e.g. Wagner 1983; Doubinger et al. 1995; Castro 2005; Wagner & Álvarez-Vázquez 2010; Bashforth et al. 2010).
Vegetational patterns
Plant assemblages of the Markley Fm can be grouped into two basic vegetational types: those of periodically flooded and generally wet substrates and those of rarely or never flooded, periodically dry substrates. At the wet end of this environmental spectrum are the organic shale beds, interpreted to have formed in swamps, and containing a typical Late Pennsylvanian wetland flora (Crookall 1959, 1966; Darrah 1969; Wagner & Álvarez-Vázquez 2010; Wagner & Castro 2010; Bashforth et al. 2010), most commonly dominated by Sigillaria brardii, Macroneuropteris scheuchzeri and marattialean ferns. Floodplain assemblages, part of the coarsening upward mudstone–sandstone interbed facies, also fall broadly into this category, and are dominated by typically Pennsylvanian wetland plants. At the drier end of the spectrum are floras from the kaolinite-dominated siltstone facies, dominated by Sphenopteridium manzanitanum and Walchia spp., with various types of Pecopteris as accessory taxa. The kaolinite-dominated siltstone beds are interpreted to be composed of clay-rich sediment formed under a hot, humid climate, and later eroded from palaeosol surfaces and transported into the deposystem, perhaps in the early phases of landscape degradation and filling of abandoned channels. The flora is the key to the environmental interpretation: Sphenopteridium is found most commonly in association with other indicators of seasonally dry climates, both in the Mississippian and in the later Pennsylvanian and earlier Permian (Mamay 1992). Similarly, conifers are typically found in strata that include evidence for seasonality of moisture availability at the time of deposition, probably in more dry to wet subhumid climates than in semi-arid conditions (e.g. Rothwell et al. 1997; Feldman et al. 2005; Looy 2007; DiMichele et al. 2008; Hernandez-Castillo et al. 2009).
A more complex picture of the vegetated landscape appears with the addition of less abundant accessory taxa. Usually common to rare within any particular collection, these taxa may occur in abundance locally and may be the dominant species within a narrow stratigraphic interval or at a single collecting site. Some of these accessory taxa are widespread and appear in a broad range of lithologies (e.g. Equisetalean sphenopsids, Cordaites and some types of Pecopteris) whereas others are lithologically specific (e.g. Pseudomariopteris cordato-ovata and Odontopteris brardii in floodplain mudstones of the coarsening upward mudstone–sandstone interbed facies or Alethopteris zeilleri in organic shales). Among many possibilities, one of the most likely explanations for this pattern is habitat heterogeneity combined with niche specificity, some taxa being tolerant of a greater range of physical conditions than others. Dispersal limitation could also play a major role within a pool of species with otherwise very similar physical tolerances, particularly within the context of a heavily vegetated wetland landscape in which incumbency and standing population size would strongly affect the likelihood of colonizing available sites (e.g. Belyea & Lancaster 1999). Such a process may account for taxa that exhibit a very restricted distribution in the Markley outcrop belt, geographically and stratigraphically, such as (?)Lepidodendron, Neurodontopteris, Alethopteris and Odontopteris.
Space v. time and lithofacies/habitat relationships: differentiating allocyclicity from autocyclicity
The clear differentiation of vegetation by lithofacies, and the ecological implications thereof pose a question that arises in any temporal study of Pennsylvanian-age vegetation, in which the ability to assess the spatial scope of a landscape is limited (e.g. compare Dimitrova et al. 2011 with Dolby et al. 2011, examining and interpreting differently the same dataset, and published back to back in the same journal), namely, ‘Did these different vegetation types coexist in relatively close proximity side by side on a landscape, shifting location through time under a fluctuating, but generally consistent climate (‘autocyclic coexistence model’)?’ Or did they succeed one another in time, expanding and contracting within the region as physical, mainly climatic, conditions changed significantly over the course of time (‘allocyclic succession model’)? Or some combination of these? Clearly, this is a question of spatial scale because the different vegetation types did coexist on Earth, and in the case of the Pennsylvanian equatorial coal belts, perhaps even in the same basic latitudinal zones. We use the terms autocyclic and allocyclic from Beerbower (1964) as modifiers for reasons that will become clear below.
Autocyclic coexistence models
In the strictest construction of the autocyclic coexistence model, widespread regional climate is assumed to be relatively constant, lithological changes reflecting only shifting loci of sedimentary dynamics. During most of the Pennsylvanian Period, in those areas where coal is abundant and well developed, this calls for wet conditions with high rainfall for most of the year, exceeding evapotranspiration (e.g. Dimitrova et al. 2010). On such a landscape, vegetational zones are thought to be controlled by topography and consequent differences in soil drainage (for another example, see Cazzulo-Klepzig et al. 2007). Under this model, changes in the distribution of vegetation through time reflect changes in loci of sedimentation and erosion intrinsic to the local setting and region. In terms of a regional sedimentary system applicable to the Pennsylvanian tropics, the deposystem might look much like the deltaic settings proposed for Pennsylvanian coal-bearing sequences (e.g. Ferm & Cavaroc 1968; Esterle & Ferm 1986; Ferm & Weisenfluh 1989), in which the driver of peat accumulation is envisioned to have been the simple cutoff of wetland areas from clastic input, thus preventing dilution of accumulating organic matter, with no consideration given to climate. In this model, significant siliciclastic transport and deposition continues within the delta elsewhere, contemporaneously with peat accumulation in such areas as abandoned, sinking delta lobes. In the Markley Fm, this model would need to be reformulated such that the prevailing climate was seasonally dry, given the record of floras (Romanchock et al. in press) and palaeosols (Tabor & Montañez 2004), but still with high rainfall for much of the year. As discussed above, the climate also seems to have been becoming more seasonally dry through time, culminating in the restriction of the wetland flora to limited areas in the Archer City Fm, deposited during the Early Permian.
A tectonic variant of the autocyclic coexistence model has recently been proposed to explain major changes in Pennsylvanian-age equatorial vegetation. In this model, mountain building events affect such things as drainage patterns, elevation and other factors that control plant biogeographic distribution (see Cleal et al. 2011 for a summary). In the Pennsylvanian, for example, the growth of the European Variscan mountains has been suggested to be the cause of widespread and rapid changes in vegetation during the Middle–Late Pennsylvanian transition, in contrast to widespread climatic changes. This model strongly downplays, though does not rule out, climatic effects on vegetational changes in time and space. This is not the place to critique in detail this particular application of such a model. Its most significant weaknesses are the great disparity in the rates at which vegetation is known to change its spatial distribution v. the rates of tectonically induced changes in elevation and drainage patterns, and the somewhat geographically local, and temporally irregular effects a tectonic model can offer when examining sequences in which there are repeated patterns of facies and vegetation through time over vast areas, such as equatorial Euramerica. Tectonic effects on the landscape of any region are important long-term modifiers of many physical variables, including climate, and are perhaps best considered as an explanatory variable for biological changes that take place on such long-term timescales.
Another version of the coexistence model is found in those sequence stratigraphic interpretations of Pennsylvanian peat-forming systems in which climate is not integral to the model (e.g. Flint et al. 1995; Heckel 1995, 2008; Bisnett & Heckel 1996; Bohacs & Suter 1997). Such models generally refer explicitly to eustatically driven sea-level fluctuations as the proximate driver of sedimentary sequences on the shelf-edge and cratonic surface and, as such, may be referred to as ‘allocyclic coexistence’ models. Peat formation is envisioned to have been driven by landscape paludification caused by rising sea-level, and is part of a system of coexistent, terrestrial and marine, sedimentary belts, pushed across the craton during sea-level rise and fall, thus making each environment time transgressive. A ‘default’ climate is generally not specified, but must be somewhere in the range of wet subhumid to humid, in the terminology of Cecil & Dulong (2003), meaning rainfall exceeding evapotranspiration for 8–10 months of the year if there is to be peat (coal) accumulation as part of the system. We can find no examples where this particular aclimatic sequence stratigraphic model has been used as a framework for detailed interpretation of terrestrial palaeoecological patterns on Pennsylvanian landscapes, although such interpretations are implicit in the model and have been alluded to broadly, particularly as they apply to peat formation, palaeosols and non-marine limestones (e.g. Jerrett et al. 2011; Belt et al. 2011). We do not consider this model further.
Allocyclic succession model
The allocyclic succession model requires the imposition of climatic changes on the local and regional sedimentary systems and biota, thus strongly affecting both (e.g. Cecil 1990; Gastaldo et al. 1993; Falcon-Lang 2004; DiMichele et al. 2008, 2010; Falcon-Lang & DiMichele 2010). When placed in a sequence stratigraphic framework (e.g. Ziegler et al. 1987; Gastaldo et al. 1993; Cecil et al. 2003; Falcon-Lang 2004; Peyser & Poulsen 2008; Gastaldo et al. 2009), this model calls for strong linkages between eustacy, climate and sediment transport dynamics in order to account for lithofacies patterns. Such models suppose a significant element of climate control on the formation of peat (deposited under humid to perhumid climates), clastics (transport being highest under seasonal climate regimes) and carbonates (marine and non-marine, forming mainly under the more strongly seasonal to arid climates). Because plants are so sensitive to climate (see discussion and citations in DiMichele et al. 2008, 2009), these climatic changes are expected to have had spatially extensive effects on vegetation (Falcon-Lang & DiMichele 2010; Falcon-Lang et al. 2009; Dolby et al. 2011), effectively centring different species pools, or biomes, in different parts of the palaeogeographic landscape at any one time, with minimal zones of overlap. Thus, lithofacies–biofacies patterns, in the context of the Pennsylvanian ice house world, would be expected to be a reflection of changing climate and depositional environment, driven by glacial–interglacial fluctuations. Different vegetation types through time are interpreted to reflect climatic shifts to which vegetation responded, similar to that seen during the Pleistocene and Holocene in response to glacial–interglacial climatic cycles (e.g. Falcon-Lang 2004).
Autocyclicity v. allocyclicity in the Markley Fm
Markley Fm vegetational and sedimentary analyses provide clear evidence of differences in physical conditions on the landscape. At question, however, is whether these differences reflect widespread, cyclical patterns of fluctuation in climate, particularly precipitation, or whether the Markley was deposited under a fairly stable climatic background within which autocyclicity dominated. The physical data, that is, the facies sequences and their spatial expression, to the degree they are known, do not lead to a clear resolution of this problem. Outcrop exposures are limited in areal extent and are widely spaced geographically. At a local, spatial scale, at all sampled sites through time and space, the environmental sequence is the same; habitats shift from palaeosols (evidencing well-drained conditions with seasonally high rainfall) and associated kaolinite-rich fluvial deposits dominated by a relatively homogeneous association of seed ferns and walchian conifers, to monotonous medullosan pteridosperm and sigillarian lycopsid-rich swamp settings, to floodplains with a diverse, spatially heterogeneous wetland flora.
Macrofossil analyses of the kaolinite-dominated siltstone and organic shale lithofacies indicate little overlap of the two species pools: (i) the ‘dry’ flora, dominated by the xeromorphic pteridosperm Sphenopteridium manzanitanum and conifers and (ii) the ‘wet’ flora, dominated by a diversity of pteridosperms, lycopsids and ferns. However, floodplain lithofacies include rare occurrences of dryland elements, within an overall wetland floristic framework. The compositional differences indicate, at a minimum, strong taphonomic segregration of the floras. The processes by which the organic remains are shed, transported and buried, and the depositional conditions under which these processes operate, are at least segregated spatially or differ in terms of the major operative sedimentary processes.
The palynological data also support the existence of two distinct floras, one a typical Late Pennsylvanian wetland flora dominated by lycopsids, sphenopsids, tree and herbaceous ferns and cordaitaleans, and the other a dryland flora dominated by seed ferns, walchian conifers and more tolerant pteridophyte taxa. However, palynology introduces nuances that are not apparent in the macrofossil record, and suggests that the allocyclic succession and autocyclic coexistence models are not necessarily mutually exclusive.
That all of the palynologically productive samples from the Markley occur within a restricted stratigraphic interval characterized by regionally developed coals suggests that these beds were deposited during an overall more humid phase. Absence of pollen (due presumably to syndepositional oxidative destruction) from stratigraphically higher intervals within the Markley is consistent with a pattern of increasing aridity in younger strata; this agrees with the disappearance of Histosols and Ultisols above Sandstone 10 (Tabor & Montañez 2004). Nevertheless, palynological data indicate a spatial overlap between dryland and wetland floras, for example, the presence of rare Sigillaria microspores in kaolinite-dominated siltstones, and the presence of rare pollen grains of walchian conifers and seed ferns in organic shales. The transitional palynofloras preserved at locality K contain a mixture of wetland and dryland elements, in addition to Sigillaria macrofossils in the same beds from which the palynologically mixed floras are derived, suggesting that they may represent an ecotonal boundary. This boundary between wetland and dryland was probably quite narrow spatially, evidenced by walchian conifer twigs occurring just below a transitional kaolinite-dominated siltstone bed with Sigillaria in its upper portions and several centimetres below an organic shale also containing Sigillaria. This pattern suggests that, at least during all or perhaps the earlier parts of this wetter phase on this particular landscape, the latest Pennsylvanian wetland and dryland floras existed in relatively close proximity.
Most or all of the organic shale and coal deposits examined in this study of the Markley and lower Archer City formations are underlain by kaolinite-dominated siltstones in abandoned channel fills, suggesting the existence of a genetic relationship between the two types of lithofacies. We propose that kaolin-rich sediments were deposited during the abandonment phases of shallow channels active during the late stages of soil formation, lining channel bottoms. These clays were stripped from surrounding interfluves, sometimes carrying plant debris derived from the well-drained interfluve floras, dominated by walchian conifers and seed ferns. There, the clays formed impediments to drainage (aquicludes), creating poorly drained depressions and the formation of local wetlands that were dominated by typical Pennsylvanian wetland species. A modern analogue might be that observed today in Borneo (Gastaldo 2010). Clay-rich deposits there form under a hot, humid climate through intensive chemical leaching, suggesting that the emplaced flora within the kaolinite-dominated siltstone lithofacies may similarly have existed under hot, humid conditions. If well-drained interfluves were dominated by ‘dryland’ floras, particularly during more intensely seasonal periods of time, wetland elements would not be expected in the kaolinite-dominated siltstone lithofacies; thus, the apparent separation of these two types of floras could be due largely to taphonomic biases operative during a transitional period from a more seasonally dry to a more seasonally wet portion of any given glacial–interglacial climatic and sedimentary cycle. This model proposes that at times, all observed floristic elements within the Markley coexisted temporally, and were spatially rather than temporally separated.
Integration of allocyclic and autocyclic succession models still allows for high-frequency climate fluctuations between humid and more seasonal precipitation regimes as the underlying driver of the habitat changes that resulted in vegetation shifts. These shifts would most likely have been driven by glacial–interglacial cycles and the attendant changes in tropical climate and sea-level accompanying them, which occurred during the Late Pennsylvanian (Olszewski & Patzkowsky 2003). At a coarser level of resolution, entire habitats linked to humid climates disappear and are replaced through time by those of drier and more seasonal climates. Moving upward into younger portions of the stratigraphic section, this long-term drying trend becomes more pronounced, with the development of calcrete-bearing palaeosols and evaporite deposits in the Archer City Fm (Jones 1971; Tabor & Montañez 2004).
Still, a question remains: ‘What was responsible for these changes in climate at any and all scales?’ To answer this, the patterns of floral and sedimentary dynamics described by this study must be viewed in the context of Late Palaeozoic climate in the western Pangean tropics. It is now well established that this region experienced a long-term drying trend accompanied by increased seasonality in precipitation beginning in latest Pennsylvanian time (Cecil 1990; Tabor & Montañez 2004; Tabor & Poulsen 2008 and references therein). This climate shift occurred at different temporal scales and was influenced by multiple drivers operating at each of these scales. For example, tectonic drivers active during the formation of Pangea (e.g. orographic uplift and erosion, change in continental position through time) likely operated at scales of 106–107 years (Tabor & Poulsen 2008), whereas glacial–interglacial cycles operated at scales of c.105 year (Heckel 1986; Davydov et al. 2010; Eros et al. 2011). Although tectonic drivers cannot account for the repetitive, high-frequency sedimentary facies and climate shifts present in the Markley Fm, ice-volume fluctuations and coincident changes in atmospheric CO2 are viable primary drivers (Poulsen et al. 2007; Peyser & Poulsen 2008). Evidence for massive expansion of multiple glacial-spreading centres during the latest Pennsylvanian has been reported from various Gondwanan basins (Isbell et al. 2003; Fielding et al. 2008). Depending on the number of spreading centres and amount of ice ablation during interglacial periods, estimates of attendant eustatic changes range from 50 m to 115 m (Isbell et al. 2003). Eustatic changes of this magnitude would have been sufficient to account for the observed orderly repetition of facies in the Markley Fm, given the lowland coastal plain on which deposition took place. Furthermore, conceptual (Cecil et al. 2003) and numerical (Peyser & Poulsen, 2008) climate models suggest that ice-volume fluctuations would have shifted the position of the Intertropical Convergence Zone (ITCZ; via the conceptual model) or altered Hadley Cell strength and, consequently, amounts of precipitation near the equator (via the numerical model). Although the mechanism is different in each model, the end result is that mean annual precipitation increases and seasonality decreases at low latitudes during glacial periods and, in contrast, mean annual precipitation decreases during interglacial periods and the seasonality of its distribution increases. When this high-frequency cycle is superimposed on the tectonically driven long-term drying trend, the average trend within the high-frequency cycles is for the wetter intervals to become less wet and the drier intervals to become more dry over the longer spans of time. This overall trend matches the precipitation signal inferred from outcrop- and regional-scale floral, pedogenic and sedimentological patterns in the Markley Fm.
Suppose, in contrast to the allocyclic succession interpretation, that the Markley Fm lithofacies–biofacies patterns reflect strictly small-scale spatial variation in the coastal plain landscape, in accordance with an autocyclic coexistence interpretation. In this case, the background climate would most likely have fluctuated no further than between humid and wet–subhumid, at least during the early and middle intervals of Markley Fm deposition, such that peat accumulation could continue on the landscape at all times, anywhere clastic input to persistently wet areas was diminished. These swamp deposits, and the buried remains of the constituent floras, are among the most likely to be preserved on a landscape with slow, tectonic creation of accommodation space, because they are low-lying, sediment-accumulating locations. The vast majority of the lowlands, in contrast, would have consisted of moderate to well drained surfaces supporting xeromorphic vegetation. This vegetation would have grown on the soils that became the palaeosol profiles that are the predominant lithology of the Markley Fm, and, as such, even though relatively poorly represented in the Markley fossil record, would have been the persistently dominant vegetation type on the coastal plain at all times. Preservation of the dryland flora occurs mainly in reworked kaolinite-dominated siltstones that seem to have been deposited in shallow channels during the final phases of abandonment. The sedimentary matrix in which the fossils are buried would have been stripped from the soil surface, over an extended time, by repeated events of erosional runoff, accounting for the variable and generally fragmentary preservation of the plant fossils. Floodplain interbedded siltstones and sandstones likely represent the encroachment phases of channels, although we cannot differentiate a fluvial system dominated by meandering processes from one in which aggradation and avulsion are the predominant processes. Such landscapes would have had relatively high soilmoisture, been subjected to high levels of disturbance, and would have had complex drainage networks, consistent with the scour and fill nature of these deposits on outcrop, the weakly developed palaeosols, and the patchiness and variability of the vegetation.
Under the autocyclic coexistence scenario, the longer-term pattern of diminishment, and ultimately disappearance, of the organic-rich facies and the shifts in palaeosol characteristics from ultisols to vertisols can be accounted for by the same processes as described above for the allocyclic succession model.
Whereas it may seem somewhat unsatisfying to leave the matter of the environmental framework unresolved, we are, as a group of authors, unresolved on which of these scenarios is most likely. The increasing refinement of sequence stratigraphic models for Pennsylvanian tropical, coal-bearing sequences points ever more strongly to the need for a significant climatic element in these models. Sea-level alone is an insufficient explanation to account for the diversity of lithologies and their repeated, stereotypical successions with a sequence of Pennsylvanian cyclothems. Furthermore, an aclimatic sequence model cannot account for the widely varied climatic signals represented by, say, coal beds and vertic, sometimes calcic palaeosols, which often lie in immediate successional juxtaposition.
Palaeobotanical patterns create still further dilemmas. Repeated temporal fluctuations occur within basins, thus in the same biogeographical space, between typical wetland assemblages and so-called ‘extrabasinal’ floras (if they are extrabasinal, what are they doing in the basins?). Both vegetation types occur in autochthonous to parautochthonous preservation, which indicates growth in areas close to the sites of deposition. Yet there is only minimal compositional overlap between these different floras, suggesting that they represent different species pools, likely reflecting different climatic backgrounds. Extension of such findings, made elsewhere, to the Markley Fm is stymied by its limited marine influence, indicating a relatively inland position on the coastal plain, its position in the western parts of the Pangean equatorial zone, placing it under a relatively drier climate than the eastern North American coal basins, and the limitations of surface exposure. Thus, at present, we believe that while many data favour an allocyclic successional interpretation, the autocyclic, coexistence interpretation cannot be ruled out, and it is most likely that the two play important complementary roles, although perhaps in different spatial and temporal frames of reference.
Acknowledgments
We thank the numerous property owners on whose land these exposures are found. Without their permission and material assistance this study would not have been possible. We are indebted to R. W. Hook and K. Craddock for their advice and participation in various aspects with this project, to the late S. Mamay and N. Hotton III for their mentorship and collaboration, and to J. Nelson for helpful discussion. The fieldwork on which this project is based was carried out over a 20-year period with various grants from the Smithsonian Institution, including the Scholarly Studies Program, Endowed Funds, and NMNH Small Grants to W.D. and to the late N. Hotton III. The research of C.L.H. was supported in part by the Intramural Research Program of the National Institutes of Health, National Library of Medicine. The paper was improved by the thoughtful comments of two anonymous reviewers, whom we thank.
References
- Alonso D, Etienne RS, McKane AJ. The merits of neutral theory. Trends in Ecology and Evolution. 2006;21:451–457. doi: 10.1016/j.tree.2006.03.019. [DOI] [PubMed] [Google Scholar]
- Bartram KM. Lycopod succession in coals: an example from the Low Barnsley Seam (Westphalian B), Yorkshire, England. In: Scott AC, editor. Coal and Coal-bearing Strata: Recent Advances. Vol. 32. Geological Society; London: 1987. pp. 187–199. Special Publications. [Google Scholar]
- Bashforth AR, Falcon-Lang HJ, Gibling MR. Vegetation heterogeneity on a Late Pennsylvanian braided-river plain draining the Variscan Mountains, La Magdalena Coalfield, northwestern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology. 2010;292:367–390. [Google Scholar]
- Beerbower JR. Cyclothems and cyclic deposition mechanisms in alluvial plain sedimentation. Kansas Geological Survey Bulletin. 1964;169:31–42. [Google Scholar]
- Belt ES, Heckel PH, Lentz LL, Bragonier WA, Lyons TW. Record of glacial–eustatic sea-level fluctuations in complex middle to late Pennsylvanian facies in the Northern Appalachian Basin and relation to similar events in the Midcontinent basin. Sedimentary Geology. 2011;238:79–100. [Google Scholar]
- Belyea LR, Lancaster J. Assembly rules within a contingent ecology. Oikos. 1999;86:402–416. [Google Scholar]
- Bennington JB, DiMichele WA, et al. Critical issues of scale in paleoecology. Palaios. 2009;24:1–4. [Google Scholar]
- Bisnett AJ, Heckel PH. Sequence stratigraphy helps to distinguish offshore from nearshore black shales in the Midcontinent Pennsylvanian succession. In: Witzke BJ, Ludvigson GA, Day J, editors. Paleozoic Sequence Stratigraphy: Views from the North American Craton. Vol. 306. Geological Society of America; 1996. pp. 341–350. Special Papers. [Google Scholar]
- Bohacs K, Suter J. Sequence stratigraphic distribution of coaly rocks: fundamental controls and paralic examples. American Association of Petroleum Geologists Bulletin. 1997;81:1612–1639. [Google Scholar]
- Bonelli JR, Jr., Patzkowsky ME. Taxonomic and ecologic persistence across the onset of the Late Paleozoic Ice Age: evidence from the Upper Mississippian (Chesterian Series), Illinois Basin, United States. Palaios. 2011;26:5–17. [Google Scholar]
- Burnham RJ, Spicer RA. Forest litter preserved by volcanic activity at El Chichon, Mexico; a potentially accurate record of the pre-eruption vegetation. Palaios. 1986;1:158–161. [Google Scholar]
- Burnham RJ, Wing SL, Parker GG. The reflection of deciduous forest communities in leaf litter: implications for autochthonous litter assemblages from the fossil record. Paleobiology. 1992;18:30–49. [Google Scholar]
- Castro Martínez MP. La flora estefaniense B de La Magdalena (Léon, España), un referente europeo. 1 & 2. Instituto Geológico y Minero de España; Madrid: 2005. [Google Scholar]
- Cazzulo-Klepzig M, Guerra-Sommer M, Menegar R. Peat-forming environment of Permian coal seams from the Faxinal coalfield (Paraná Basin) in southern Brazil, based on palynology and palaeobotany. Revista brasileira de Paleontologia. 2007;10:117–127. [Google Scholar]
- Cecil CB. Paleoclimate controls on stratigraphic repetition of chemical and siliciclastic rocks. Geology. 1990;18:533–536. [Google Scholar]
- Cecil CB, Dulong FT. Precipitation models for sediment supply in warm climates. In: Cecil CB, Edgar NT, editors. Climate Controls on Stratigraphy. Vol. 77. SEPM Special Publications; Tulsa: 2003. pp. 21–27. [Google Scholar]
- Cecil CB, Dulong FT, West RR, Stamm R, Wardlaw B, Edgar NT. Climate controls on the stratigraphy of a middle Pennsylvanian cyclothem in North America. In: Cecil CB, Edgar NT, editors. Climate Controls on Stratigraphy. Vol. 77. SEPM Special Publications; Tulsa: 2003. pp. 151–180. [Google Scholar]
- Chambers JM, Cleveland WS, Kleiner B, Tukey PA. Graphical Methods for Data Analysis. Wadsworth; Boston: 1983. [Google Scholar]
- Cleal CJ. The Westphalian–Stephanian macrofloral record from the South Wales Coalfield. Geological Magazine. 2007;144:465–486. [Google Scholar]
- Cleal CJ, Opluštil S, Thomas BA, Tenchov Y. Pennsylvanian vegetation and climate in tropical Variscan Euramerica. Episodes. 2011;34:3–12. [Google Scholar]
- Cleal CJ, Uhl D, Cascales-Miñana B, Thomas BA, Bashforth AR, King SC, Zodrow EL. Plant biodiversity changes in Carboniferous tropical wetlands. Earth-Science Reviews. 2012;114:124–155. [Google Scholar]
- Crookall R. Fossil plants of the Carboniferous rocks of Great Britain, 2nd section, Neuropteridae. Memoirs of the Geological Survey of Great Britain, London, Palaeontology. 1959;4:85–216. [Google Scholar]
- Crookall R. Fossil plants of the Carboniferous rocks of Great Britain, 2nd section. Memoirs of the Geological Survey of Great Britain, Palaeontology. 1966;4:355–571. [Google Scholar]
- Currano ED, Wilf P, Wing SL, Labandeira CC, Lovelock EC, Royer DL. Sharply increased insect herbivory during the Paleocene-Eocene thermal maximum. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:1960–1964. doi: 10.1073/pnas.0708646105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darrah WC. A critical review of the Upper Pennsylvanian floras of eastern United States with notes on the Mazon Creek flora of Illinois. Gettysburg, Pennsylvania: 1969. Privately printed. [Google Scholar]
- Davydov VI, Crowley JL, Schmitz MD, Poletaev VI. High-precision U–Pb zircon age calibration of the global Carboniferous time scale and Milankovitch band cyclicity in the Dontz Basin, eastern Ukraine. Geochemistry, Geophysics, Geosystems. 2010;11 http://dx.doi.org/10.1029/2009GC002736. [Google Scholar]
- DiMichele WA, Phillips TL. The ecology of Paleozoic ferns. Review of Palaeobotany and Palynology. 2002;119:143–159. [Google Scholar]
- DiMichele WA, Phillips TL, McBrinn GE. Quantitative analysis and paleoecology of the Secor coal and roof-shale floras (Middle Pennsylvanian, Oklahoma) Palaios. 1991;6:390–409. [Google Scholar]
- DiMichele WA, Tabor NJ, Chaney DS, Nelson WJ. From wetlands to wetspots: environmental tracking and the fate of Carboniferous elements in Early Permian tropical floras. In: Greb SF, DiMichele WA, editors. Wetlands Through Time. Vol. 399. Geological Society of America Special Papers; 2006. pp. 223–248. [Google Scholar]
- DiMichele WA, Falcon-Lang HJ, Nelson WJ, Elrick SD, Ames PR. Ecological gradients within a Pennsylvanian forest mire. Geology. 2007;35:415–418. [Google Scholar]
- DiMichele WA, Kerp H, Tabor NJ, Looy CV. The so-called ‘Paleophytic–Mesophytic’ transition in equatorial Pangea: multiple biomes and vegetational tracking of climate through geological time. Palaeogeography, Palaeoclimatology, Palaeoecology. 2008;268:152–163. [Google Scholar]
- DiMichele WA, Montañez IP, Poulsen CJ, Tabor NJ. Climate and vegetational regime shifts in the late Paleozoic ice age earth. Geobiology. 2009;7:220–226. doi: 10.1111/j.1472-4669.2009.00192.x. [DOI] [PubMed] [Google Scholar]
- DiMichele WA, Cecil CB, Montañez IP, Falcon-Lang HJ. Cyclic changes in Pennsylvanian paleoclimate and effects on floristic dynamics in tropical Pangaea. International Journal of Coal Geology. 2010;83:329–344. [Google Scholar]
- Dimitrova TK, Zodrow EL, Cleal CJ, Thomas BA. Palynological evidence for Pennsylvanian (Late Carboniferous) vegetation change in the Sydney Coalfield, eastern Canada. Geological Journal. 2010;45:388–396. [Google Scholar]
- Dimitrova TK, Cleal CJ, Thomas BA. Palynological evidence for Pennsylvanian extra-basinal vegetation in Atlantic Canada. Journal of the Geological Society, London. 2011;168:559–569. [Google Scholar]
- Dolby G, Falcon-Lang HJ, Gibling MR. A conifer-dominated palynological assemblage from Pennsylvanian (late Moscovian) alluvial drylands in Atlantic Canada: implications for the vegetation of tropical lowlands during glacial phases. Journal of the Geological Society, London. 2011;168:571–584. [Google Scholar]
- Doubinger J, Vetter P, Langiaux J, Galtier J, Broutin J. The flore fossile du basin houiller de Saint-Étienne. Mémoires du Muséum National D’Histoire Naturelle. 1995;164:1–355. [Google Scholar]
- Dudal R, Eswaran H. Distribution, Properties and Classification of Vertisols. Publication Soil Management Support Services, US Department of Agriculture, Natural Resources Conservation Service; Washington DC: 1988. pp. 1–22. [Google Scholar]
- Eble CF, Gastaldo RA, Demko TM, Liu Y. Coal compositional changes along a mire interior to mire margin transect in the Mary Lee coal bed, Warrior Basin, Alabama, USA. International Journal of Coal Geology. 1994;26:43–62. [Google Scholar]
- Eble CF, Grady WC, Pierce BS. Compositional characteristics and inferred origln of three Late Pennsylvanian coal beds from the northern Appalachian Basin. In: Greb SF, DiMichele WA, editors. Wetlands Through Time. Vol. 399. Geological Society of America Special Papers; 2006. pp. 191–222. [Google Scholar]
- Elsik WC, Dilcher DL. Palynology and age of clays exposed in Lawrence Clay Pit, Henry County, Tennessee. Palaeontographica. 1974;146B:65–87. [Google Scholar]
- Eros JM, Montañez IP, Osleger D, Davydov V, Nemyrovska T, Poletaev V, Zhykalyak M. Upper Carboniferous sequence stratigraphy and relative sea level history, Donets Basin, Ukraine. Palaeogeography, Palaeoclimatology, Palaeoecology. 2011;313–314:1–25. [Google Scholar]
- Esterle JS, Ferm JC. Relationship between petrographic and chemical properties and coal seam geometry, Hance seam, Breathitt Fm, southeastern Kentucky. International Journal of Coal Geology. 1986;6:199–214. [Google Scholar]
- Falcon-Lang HJ. Pennsylvanian tropical rain forests responded to glacial–interglacial rhythms. Geology. 2004;32:689–692. [Google Scholar]
- Falcon-Lang HJ, DiMichele WA. What happened to the coal forests during Pennsylvanian glacial phases? Palaios. 2010;25:611–617. [Google Scholar]
- Falcon-Lang HJ, Benton MJ, Braddy SJ, Davies SJ. The Pennsylvanian tropical biome reconstructed from the Joggins Fm of Nova Scotia, Canada. Journal of the Geological Society, London. 2006;163:561–576. [Google Scholar]
- Falcon-Lang HJ, Nelson WJ, Elrick SD, Looy CV, Ames PR, DiMichele WA. Incised channel-fills containing conifers imply that seasonally-dry vegetation dominated Pennsylvanian tropical lowlands. Geology. 2009;37:923–926. [Google Scholar]
- Fall LM, Olszewski TD. Environmental disruptions influence taxonomic composition of brachiopod paleocommunities in the Middle Permian Bell Canyon Fm (Delaware Basin, West Texas) Palaios. 2010;25:247–259. [Google Scholar]
- Feldman HR, Franseen EK, Joeckel RM, Heckel PH. Impact of longer-term modest climate shifts on architecture of high-frequency sequences (cyclothems), Pennsylvanian of midcontinent U.S.A. Journal of Sedimentary Research. 2005;75:350–368. [Google Scholar]
- Ferm JC, Cavaroc VV., Jr. A nonmarine sedimentary model for the Allegheny rocks of West Virginia. In: Klein GD, editor. Late Paleozoic and Mesozoic Continental Sedimentation, Northeastern North America. Vol. 106. Geological Society of America Special Papers; 1968. pp. 1–19. [Google Scholar]
- Ferm JC, Weisenfluh GA. Evolution of some depositional models in late Carboniferous rocks of the Appalachian coal fields. International Journal of Coal Geology. 1989;12:259–292. [Google Scholar]
- Fielding CR, Frank TD, Isbell JL. The late Paleozoic ice age–a review of current understanding and synthesis of global climate patterns. In: Fielding CR, Frank TD, Isbell JL, editors. Resolving the Late Paleozoic Ice Age in Time and Space. Vol. 441. Geological Society of America Special Papers; 2008. pp. 343–354. [Google Scholar]
- Flint S, Aitken J, Hampson G. Application of sequence stratigraphy to coal-bearing coastal plain successions: implications for the UK Coal Measures. In: Whateley MKG, Spears DA, editors. European Coal Geology. Vol. 82. Geological Society; London: 1995. pp. 1–16. Special Publications. [Google Scholar]
- Gastaldo RA. Peat or no peat: why do the Rajang and Mahakam deltas differ? International Journal of Coal Geology. 2010;83:162–172. [Google Scholar]
- Gastaldo RA, Demko TM, Liu Y. Application of sequence and genetic stratigraphic concepts to Carboniferous coal-bearing strata: an example from the Black Warrior basin, USA. Geologische Rundschau. 1993;82:212–226. [Google Scholar]
- Gastaldo RA, Stevanovic-Walls I, Ware WN, Greb SF. Community heterogeneity of Early Pennsylvanian peat mires. Geology. 2004;32:693–696. [Google Scholar]
- Gastaldo RA, Purkyňov́ E, Šimůnek Z. Megafloral perturbation across the Enna Marine Zone in the Upper Silesian Basin attests to Late Mississippian (Serpukhovian) deglaciation and climate change. Palaios. 2009;24:351–366. [Google Scholar]
- Greb SF, DiMichele WA, Gastaldo RA. Evolution and importance of wetlands in Earth history. In: Greb SF, DiMichele WA, editors. Wetlands Through Time. Vol. 399. Geological Society of America Special Papers; 2006. pp. 1–40. [Google Scholar]
- Hammer Ø, Harper DAT, Ryan PD. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica. 2001;4:1–9. [Google Scholar]
- Heckel PH. Sea-level curve for Pennsylvanian eustatic marine transgressive-regressive depositional cycles along mid-continent outcrop belt, North America. Geology. 1986;14:330–334. [Google Scholar]
- Heckel PH. Glacial-eustatic base-level – climatic model for late Middle to Late Pennsylvanian coal-bed formation in the Appalachian Basin. Journal of Sedimentary Research. 1995;B65:348–356. [Google Scholar]
- Heckel PH. Pennsylvanian cyclothems in Mid-continent North America as far-field effects of waxing and waning of Gondwana ice sheets. In: Fielding CR, Frank TD, Isbell JL, editors. Resolving the Late Paleozoic Ice Age in Time and Space. Vol. 441. Geological Society of America Special Papers; 2008. pp. 275–289. [Google Scholar]
- Hentz TF. Lithostratigraphy and paleoenvironments of Upper Paleozoic Continental Red Beds, North-Central Texas–Bowie (new) and Wichita (revised) Groups. Bureau of Economic Geology Report of Investigations; Austin, TX: 1988. Report 170. [Google Scholar]
- Hernandez-Castillo GR, Stockey RA, Rothwell GW, Mapes G. Reconstructing Emporia lockardii (Voltziales: Emporiaceae) and initial thoughts on Paleozoic conifer ecology. International Journal of Plant Sciences. 2009;170:1056–1074. [Google Scholar]
- Hill MO, Gauch HG., Jr. Detrended correspondence analysis: an improved ordination technique. Vegetatio. 1980;42:47–58. [Google Scholar]
- Holland SM, Zaffos A. Niche conservatism along an onshore–offshore gradient. Paleobiology. 2011;37:270–286. [Google Scholar]
- Isbell JL, Miller MF, Wolfe KL, Lenaker PA. Timing of late Paleozoic glaciation in Gondwana: was glaciation responsible for the development of northern hemisphere cyclothems? In: Chan MA, Archer AW, editors. Extreme Depositional Environments: Mega End Members in Geologic Time. Vol. 370. Geological Society of America Special Papers; 2003. pp. 5–24. [Google Scholar]
- Ivany LC, Brett CE, Wall HLB, Wall PD, Handley JC. Relative taxonomic and ecologic stability in Devonian marine faunas of New York State: a test of coordinated stasis. Paleobiology. 2009;35:499–524. [Google Scholar]
- Jerrett RM, Hodgson DM, Flint SS, Davies RC. Control of relative sea level and climate on coal character in the Westphalian C (Atokan) Four Corners Fm, Central Appalachian Basin, U.S.A. Journal of Sedimentary Research. 2011;81:420–445. [Google Scholar]
- Jones JO. The Blaine Fm of North Texas. University of Iowa; Iowa City: 1971. Unpublished PhD dissertation. [Google Scholar]
- Jones JO, Hentz TR. Geological Society of America Centennial Field Guide – South-Central Section. Geological Society of America; Boulder, Colorado: 1988. Permian strata of North-Central Texas; pp. 309–316. [Google Scholar]
- Knight J. The stratigraphy of the Stephanian rocks of the Sabero coalfield, León (NW Spain) and an investigation of the fossil flora. Palaeontographica. 1985;197B:1–80. [Google Scholar]
- Lamboy W, Lesnikowska A. Some statistical methods useful in the analysis of plant paleoecological data. Palaios. 1988;3:86–94. [Google Scholar]
- Libertín M, Dašková J, Opluštil S, Bek J, Edress N. A palaeoecological model for a vegetated early Westphalian intramontane valley (Intra-Sudetic Basin, Czech Republic) Review of Palaeobotany and Palynology. 2009;155:175–203. [Google Scholar]
- Looy CV. Extending the range of derived late Paleozoic conifers: Lebowskia gen. nov. (Majonicaceae) International Journal of Plant Sciences. 2007;168:957–972. [Google Scholar]
- Mamay SH. Sphenopteridium and Telangiopsis in a Diplopteridium-like association from the Virgilian (Upper Pennsylvanian) of New Mexico. American Journal of Botany. 1992;79:1092–1101. doi: 10.1002/j.1537-2197.1992.tb13703.x. [DOI] [PubMed] [Google Scholar]
- McCune B, Grace JB. Analysis of Ecological Communities. MJM Software Design; Gleneden Beach, OR: 2002. [Google Scholar]
- Montañez IP, Tabor NJ, et al. CO2-forced climate and vegetation instability during late Paleozoic deglaciation. Science. 2007;315:87–91. doi: 10.1126/science.1134207. [DOI] [PubMed] [Google Scholar]
- Nichols G. Sedimentology and Stratigraphy. 2nd Wiley-Blackwell; London: 2009. [Google Scholar]
- Nordt L, Orosz M, Driese S, Tubbs J. Vertisol carbonate properties in relation to mean annual precipitation: Implications for paleoprecipitation estimates. The Journal of Geology. 2006;114:501–510. [Google Scholar]
- Olszewski TD, Erwin DH. Change and stability in the Permian brachiopod communities of western Texas. Palaios. 2009;24:27–40. [Google Scholar]
- Olszewski TD, Patzkowsky ME. From cyclothems to sequences: the record of eustasy and climate on an icehouse epeiric platform (Pennsylvanian–Permian, North American Midcontinent) Journal of Sedimentary Research. 2003;73:15–30. [Google Scholar]
- Opluštil S, Pšenička J, Libertín M, Šimůnek Z. Vegetation patterns of Westphalian and lower Stephanian mire assemblages preserved in tuff bed of the continental basins of Czech Republic. Review of Palaeobotany and Palynology. 2007;143:107–154. [Google Scholar]
- Opluštil S, Pšenička J, Libertín M, Bashforth AR, Šimůnek Z, Daškov́ J. A Middle Pennsylvanian (Bolsovian) peat-forming forest preserved in situ in volcanic ash of the Whetstone Horizon in the Radnice Basin, Czech Republic. Review of Palaeobotany and Palynology. 2009a;155:234–274. [Google Scholar]
- Opluštil S, Pšenička J, Libertín M, Bek J, Daškov́ J, Šimůnek Z. Composition and structure of an in situ Middle Pennsylvanian peat-forming plant assemblage buried in volcanic ash, Radnice Basin (Czech Republic) Palaios. 2009b;24:726–746. [Google Scholar]
- Oxanen J, Minchin PR. Instability of ordination results under changes in input data order: explanations and remedies. Journal of Vegetation Science. 1997;8:447–454. [Google Scholar]
- Peyser CE, Poulsen CJ. Soreghan GS, Montañez IP, editors. Controls on Permo-Carboniferous precipitation over tropical Pangaea: a GCM sensitivity study. Special Issue on the Late Paleozoic Earth System. Palaeogeography, Palaeoclimatology, Palaeoecology. 2008;268:181–192. [Google Scholar]
- Pfefferkorn HW, Thomson M. Changes in dominance patterns in Upper Carboniferous plant-fossil assemblages. Geology. 1982;10:641–644. [Google Scholar]
- Pfefferkorn HW, Mustafa H, Hass H. Quantitative Charakterisierung ober–karboner Abdruckfloren (quantitative characterization of compression–impression floras of Upper Carboniferous age) Neues Jahrbuch Geologie Palaeontologie, Abhandlungen. 1975;150:253–269. [Google Scholar]
- Phillips TL, Peppers RA. Changing patterns of Pennsylvanian coal-swamp vegetation and implications of climatic control on coal occurrence. International Journal of Coal Geology. 1984;3:205–255. [Google Scholar]
- Potter FW., Jr. Investigations of angiosperms from the Eocene of southeastern North America: pollen assemblages from Miller Pit, Henry County, Tennessee. Palaeontographica. 1976;157B:44–96. [Google Scholar]
- Potter FW, Jr., Dilcher DL. Biostratigraphic analysis of Eocene clay deposits in Henry County, Tennessee. In: Dilcher DL, Taylor TN, editors. Biostratigraphy of Fossil Plants. Dowden, Hutchinson, and Ross; Stroudsburg, PA: 1980. pp. 211–225. [Google Scholar]
- Poulsen CJ, Pollard D, Montañez IP, Rowley D. Late Paleozoic tropical climate response to Gondwanan deglaciation. Geology. 2007;35:771–774. [Google Scholar]
- Pryor JS, Gastaldo RA. Paleoecological analysis of two Early Pennsylvanian mineral-substrate wetlands. Palaios. 2000;15:3–13. [Google Scholar]
- Pšenička J, Bek J, Cleal CJ, Wittry J, Zodrow EL. Description of synangia and spores of the holotype of the Carboniferous fern Lobatopteris miltoni, with taxonomic comments. Review of Palaeobotany and Palynology. 2009;155:133–144. [Google Scholar]
- R DEVELOPMENT CORE TEAM . R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; Vienna, Austria: 2010. http://www.R-project.org. [Google Scholar]
- Romanchock CM, DiMichele WA, Chaney D, Hook RW. A latest Pennsylvanian fossil flora from the Markley Fm, north-central Texas. Review of Paleobotany and Palynology. in press.
- Rothwell GW, Mapes G, Mapes RH. Late Paleozoic conifers of North America: structure, diversity and occurrences. Review of Palaeobotany and Palynology. 1997;95:95–113. [Google Scholar]
- Scheihing MH. Reduction of wind velocity by the forest canopy and the rarity of non-arborescent plants in the Upper Carboniferous fossil record. Argumenta Palaeobotanica. 1980;6:133–138. [Google Scholar]
- Scheihing MH, Pfefferkorn HW. The taphonomy of land plants in the Orinoco Delta: a model for the incorporation of plant parts in clastic sediments of Late Carboniferous age of Euramerica. Review of Palaeobotany and Palynology. 1984;41:205–240. [Google Scholar]
- Scotese CR. PALEOMAP Animations ‘Paleogeography’. PALEOMAP Project, Department of Geology, University of Texas; Arlington, TX: 1999. [Google Scholar]
- Scott AC. Sedimentological and ecological control of Westphalian B plant assemblages from West Yorkshire. Proceedings of the Yorkshire Geological Society. 1978;41:461–508. [Google Scholar]
- Secord R, Wing SL, Chew A. Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology. 2008;34:282–300. [Google Scholar]
- Šimůnek Z, Martínek K. A study of Late Carboniferous and Early Permian plant assemblages from the Boskovice Basin, Czech Republic. Review of Palaeobotany and Palynology. 2009;155:275–307. [Google Scholar]
- Tabor NJ, Montañez IP. Morphology and distribution of fossil soils in the Permo-Pennsylvanian Wichita and Bowie Groups, north-central Texas, USA: implications for western equatorial Pangean palaeoclimate during icehouse–greenhouse transition. Sedimentology. 2004;51:851–884. [Google Scholar]
- Tabor NJ, Poulsen CJ. Soreghan GS, Montañez IP, editors. Palaeoclimate across the Late Pennsylvanian–Early Permian tropical palaeolatitudes: a review of climate indicators, their distribution, and relation to palaeophysiographic climate factors. Special Issue on the late Paleozoic Earth System. Palaeogeography, Palaeoclimatology, Palaeoecology. 2008;268:293–310. [Google Scholar]
- Tabor NJ, Montañez IP, Scotese CR, Poulsen CJ, Mack GH. Paleosol archives of environmental and climatic history in paleotropical western Pangea during the latest Pennsylvanian through Early Permian. In: Fielding CR, Frank TD, Isbell JL, editors. Resolving the Late Paleozoic Ice Age in Time and Space. Vol. 441. Geological Society of America Special Papers; 2008. pp. 291–303. [Google Scholar]
- Tabor NJ, Smith RMH, Steyer JS, Sidor CA, Poulsen CJ. The Permian Moradi Fm of northern Niger: paleosol morphology, petrography and mineralogy. Palaeogeography, Palaeoclimatology, Palaeoecology. 2011;299:200–213. [Google Scholar]
- Ter Braak CJF, Šmilauer P. CANOCO Reference manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5) Microcomputer Power; Ithaca: 2002. [Google Scholar]
- Wagner RH. Upper Stephanian stratigraphy and palaeontology of the Puertollano Basin, Ciudad Real, Spain. Anais da Faculdade de Ciências do Universidade Porto. 1983;64:171–231. [Google Scholar]
- Wagner RH, Álvarez-Vázquez C. The Carboniferous floras of the Iberian Peninsula: a synthesis with geological connotations. Review of Palaeobotany and Palynology. 2010;162:239–324. [Google Scholar]
- Wagner RH, Castro MP. Compositional changes in a mid-Stephanian (Kasimovian) flora in relation to alluvial plain deposits derived from westward-receding mountains and bordered by the paleotethys: La Magdalena Coalfield, northwestern Spain. Palaios. 2010;26:33–54. [Google Scholar]
- Walker RG, James NP. Facies Models: Response to Sea Level Changes. Geological Association of Canada; St. John’s: 1992. [Google Scholar]
- Wardlaw BR. Age assignment of the Pennsylvanian-Early Permian succession of North Central Texas. Permophiles. 2005;46:21–22. [Google Scholar]
- Weiher E, Freund D, Bunton T, Stefanski A, Lee T, Bentivenga S. Advances, challenges and a developing synthesis of ecology community assembly theory. Philosophical Transactions of the Royal Society of London B. 2011;366:2403–2413. doi: 10.1098/rstb.2011.0056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wing SL, DiMichele WA. Conflict between local and global changes in plant diversity through geological time. Palaios. 1995;10:551–564. [Google Scholar]
- Wing SL, Harrington GJ, Smith FA, Bloch JI, Boyer DM, Freeman KH. Transient floral change and rapid global warming at the Paleocene–Eocene boundary. Science. 2005;310:993–996. doi: 10.1126/science.1116913. [DOI] [PubMed] [Google Scholar]
- Young FW. Multidimensional Scaling. In: Kotz S, Johnson NL, editors. Encyclopedia of Statistical Sciences. Vol. 5. Wiley; New York: 1985. pp. 649–659. [Google Scholar]
- Ziegler AM, Raymond A, Gierlowski TC, Horrell MA, Rowley DB, Lottes AL. Coal, climate and terrestrial productivity: the present and Early Cretaceous compared. In: Scott AC, editor. Coal and Coal-Bearing Strata: Recent Advances. Vol. 32. Geological Society; London: 1987. pp. 25–49. Special Publications. [Google Scholar]