

Abstract
AIM: Nitric oxide (NO) has been implicated in the pathogenesis of liver cirrhosis. We have found inducible nitric oxide synthase (iNOS) can be induced in hepatocytes of cirrhotic liver. This study further investigated the temporal expression and activity of hepatic iNOS in cirrhosis development.
METHODS: Cirrhosis was induced in rats by chronic bile duct ligation (BDL). At different time points after the operation, samples were collected to examine NO concentration, liver function, and morphological changes. Hepatocytes were isolated for determination of iNOS mRNA, protein and enzymatic activity.
RESULTS: Histological examination showed early cirrhosis 1-2 wk after BDL, with advanced cirrhosis at 3-4 wk. Bilirubin increased dramatically 3 d after BDL, but decreased by 47% on d 14. Three weeks after BDL, it elevated again. Systemic NO concentration did not increase significantly until 4 wk after BDL, when ascites developed. Hepatocyte iNOS mRNA expression was identified 3 d after BDL, and enhanced with time to 3 wk, but reduced thereafter. iNOS protein showed a similar pattern to mRNA expression. iNOS activity decreased from d 3 to d 7, but increased again thereafter till d 21.
CONCLUSION: Hepatic iNOS can be induced in the early stage, which increases with time as cirrhosis develops. Its enzymatic activity is significantly correlated with protein expression and histological alterations of the liver, but not with systemic NO levels, nor with absolute values of liver function markers.
Keywords: Liver Cirrhosis, Inducible nitric oxide synthase, Nitric Oxide, Bile duct ligation
INTRODUCTION
Nitric oxide (NO) is a pluripotent gaseous free radical that possesses a wide range of physiologic functions. It is produced from L-arginine by nitric oxide synthase (NOS), a family with three members, namely endothelial NOS (eNOS), inducible NOS (iNOS) and neuronal NOS (nNOS)[1,2]. Recently, NO has been implicated in the pathogenesis of liver cirrhosis and/or portal hypertension[3]. Many studies have demonstrated that enhanced production of NO in the systemic as well as splanchnic vasculature of both cirrhotic animal models and human patients, as indicated by elevated plasma levels of nitrites and nitrates, increased aortic NOS activity and cGMP levels[4-10]. In the liver itself, impaired sinusoidal eNOS activity has been suggested to be responsible, at least in part, for the increased intra-hepatic resistance, which is one of the characteristic dynamic changes of liver cirrhosis and/or portal hypertension[11,12]. In a previous study, we found that all the three isoforms of NOS were expressed in cirrhosis, which was induced by chronic bile duct ligation (BDL), but with different patterns. eNOS protein was redistributed in cirrhotic livers, while its activity decreased. nNOS and iNOS were induced in hepatocytes at both mRNA and protein levels with iNOS activity increased significantly[10]. These studies suggest complex functions and regulations of NO and NOS in liver cirrhosis.
However, the exact role of NO in liver cirrhosis is still not clear. This is true especially with its expression and function in the course of liver cirrhosis development, in which NO may be an important mediator. Although there were some studies showing NOS expression in acute liver injuries or at the very early stage of liver cirrhosis[13,14], the NO production, modulation and more importantly, its patho-physiological role at different stages of cirrhosis formation are still largely unknown. To shed more light on this issue, this study was designed to assess the hepatic NO/NOS activity at different stages of cirrhosis development with a cirrhosis model induced by BDL. Hepatocytes are the major cell types, comprising 80% of liver by volume and 65% of liver by cell number[15]. Moreover, iNOS has been found to be up-regulated in hepatocytes of cirrhotic livers with increased activity. Therefore, we evaluated iNOS expression and activity in hepatocytes at different time points from cirrhosis initiation to advancement. NO concentration was monitored regularly in plasma as well as the supernatant of hepatocyte culture. In addition, histological examination and liver function test were performed, and correlation analysis was done to assess their relationships with hepatic iNOS expression and activity and NO levels.
MATERIALS AND METHODS
Induction of cirrhosis in rats and grouping
Liver cirrhosis was induced in male Sprague-Dawley rats, weighing 180-250 g, by BDL as described previously[16,17]. Control rats received a sham operation, which was in the same manner, except that the bile duct was only gently manipulated, but not ligated or sectioned. At different time points, namely 3 d, 1-4 wk at weekly intervals after surgery, some BDL or sham-operated rats were randomly chosen for sacrifice by anesthesia for study. Enough subjects were included to make sure there were at least 8 BDL or 6 sham-operated rats in each group upon harvest. Half the subjects from each group were used for histological examination and chemistry analysis, while the rest were used for isolating hepatocytes and investigating NOS expression and activity. These protocols were in accordance with the institute guidelines and all rats received humane care.
General treatment
At each time point, rats (4 for BDL, 3 for sham-operated) were randomly selected for histological examination and chemistry analysis. For morphological study, livers were removed from the abdomen cavity, washed in phosphate buffered saline (PBS) and fixed in 40 g/L formaldehyde. The liver tissue blocks were dehydrated, cleared and then embedded in paraffin. Sections were cut at 4 micrometers in thickness for routine hematoxylin and eosin staining. Slides were read by two pathologists independently. A histological scoring system modified from that of Ishak et al[18] was set up to evaluate the pathological changes of the liver semi-quantitatively. For chemistry analysis, blood was taken from the inferior vena cava for liver function test as well as measurement of systemic NO concentration. Various parameters of liver function including bilirubin, aspartate aminotransferase (AST), alanine aminotransferase (ALT) were determined by routine clinical chemistry at National University Hospital, Singapore. Systemic NO level was detected by measuring its stable metabolic products, nitrite and nitrate, using a NO kit (Oxford Biomedical Research, USA) based on the colorimetric Griess reaction.
Isolation of hepatocytes
Hepatocytes were isolated by in situ collagenase perfusion as described previously[19,20] with minimal modifications. Briefly, after anesthesia, BDL or sham-operated rats were perfused with 400 mL pre-perfusion buffer (154 mmol/L NaCl, 27 mmol/L KCl, 5.5 mmol/L D-glucose, 5.5 mmol/L NaHCO3, 20 mmol/L HEPES and 1 mmol/L EDTA, pH 7.2) at a rate of 20 mL/min, followed by perfusion buffer (pre-perfusion buffer without EDTA, plus 11 mmol/L CaCl2) for 5 min. The livers of sham-operated rats were perfused with 500 mg/L collagenase (Sigma-Aldrich Fine Chemicals, USA) solution (collagenase in perfusion buffer) for 10 min. For BDL rats, the percentage of collagenase used was 1 g/L and the perfusion time ranged from 10 to 30 min depending on the stage of liver cirrhosis. The resultant digested liver was excised and minced and the dispersed cells were passed through a steel screen (100 mesh, Sigma-Aldrich Fine Chemicals, USA). The cells were then washed twice with cold perfusion buffer and centrifuged at 30 g for 2 min at 4 °C. Hepatocyte viability was determined by trypan blue exclusion and characterized by phase-contrast microscopy. Purified hepatocytes were then suspended in Williams medium E supplemented with 100 mL/L FBS, 0.5 U/L insulin, 15 mmol/L HEPES, 50 U/mL penicillin, and 50 μg/mL streptomycin and maintained in a humidified CO2 (50 mL/L) incubator at 37 °C. For further study, 5×106 hepatocytes were plated into each 75 cm2 flask. Two hours after incubation, hepatocytes were usually attached, and the monolayer was then used for total RNA extraction, Western blotting and enzymatic activity estimation.
Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA from hepatocyte monolayer was extracted using TRIzolTM Reagent (Gibco BRL, Grand Island, NY), according to the manufacturer’s protocol. Integrity of the total RNA was checked with formaldehyde agarose gel (10 g/L) and indicated by the presence of the 28 S and 18 S rRNA bands. RT was performed with oligodT using SUPERSCRIPT II RNase H- reverse transcriptase (Gibco BRL, Germany). First strand cDNA synthesis was performed at 42 °C for 1 h. The reaction was stopped by heating the mixture at 95 °C for 10 min and stored at -20 °C until further use. PCR was done using a DNA amplification reagent kit (QIAGENE, Germany) with Hybaid PCR express thermal cycler (UK). The primers of iNOS were as follows: 5’ AATAGAGGAACATCTGGCCAGG 3’ (forward) and 5’ ACTTCCTCCAGGATGTTGTA 3’ (reverse). The PCR product was predicted to be a 710 bp fragment from position 468 to position 1178 in the cDNA sequence of rat iNOS (GenBank Accession No. U03699). Each PCR reaction contained 0.2 μL of RT product, 0.4 μmoL of sense or antisense primers, 50 μmoL of each dNTPs, and 0.5 U of Taq polymerase in a final volume of 25 μL. The amplification procedure consisted of 30 cycles with the following parameters: denaturation at 94 °C for 1 min, annealing at 55 °C for 2 min, and extension at 72 °C for 1 min. Each final PCR product was loaded on agarose gel (18 g/L), eletrophoresed and visualised by ethidium bromide staining.
Competitive PCR
Competitive PCR is an established method for quantitative measurement of gene expression, which has been widely used[21,22]. To examine the levels of mRNA expression, a constant amount of the sample cDNA was co-amplified with a known concentration of the internal competitor DNA, MIMIC, which was constructed according to the manufacture’s protocol of PCR MIMICTM construction kit (CLONTECH Laboratories Inc, USA). To determine the amount of iNOS MIMIC to be used in the PCR amplification, a preliminary experiment in each case was done. A constant amount of cDNA of iNOS was first amplified together with a series of ten-fold dilution of its MIMIC. This reaction provided a fine-tuned two-fold dilution serial of the MIMIC. The same amount of cDNA was then co-amplified with the two-fold MIMIC dilution series. The amount of MIMIC, which gave an optical density ratio of 1:1 with the target, was chosen to co-amplify with the rest of RT products. After PCR, the products were eletrophoresed on agarose gel. The ratio of the optical density of the PCR product pairs (target vs MIMIC) was determined using the Kodak 1D imaging system (Eastman Kodak Company, USA).
Western blot analysis
To detect iNOS immunoreactive proteins, hepatocyte monolayer was homogenized in a lysis buffer containing 320 mmol/L sucrose, 20 mmol/L HEPES, 0.5 mmol/L EDTA, 1 mmol/L DL-dithiothreitol, 0.01 mg/mL leupeptin, 0.01 mg/mL soybean trypsin inhibitor, and 2 μg/mL aprotinin (pH 7.2), and then centrifuged at 20000 g for 30 min at 4 °C. Protein quantification of the supernatants was performed using the standard Bio-Rad protein microassay. Samples were subjected to 6% SDS-polyacrylamide gel electrophoresis and proteins were transferred to nitrocellulose membranes using a Mini Trans-Blot cell (Bio-Rad Laboratories, CA, USA). The membranes were then incubated with 1:500 dilution of anti-iNOS monoclonal antibody (Santa Cruz Biotechnology Inc, USA) for 1 h. After washing, the membranes were incubated with a 1:1000 dilution of horseradish peroxide-conjugated secondary antibody (Amersham-operated Pharmacia Biotech, NJ, USA). Blots were then visualized using the ECL kit from Amersham-operated according to the manufacturer’s protocol. Relative densities of the bands were analyzed using the Kodak 1D imaging system.
NOS activity determination
The conversion of 14C-labeled L-arginine to 14C-labeled L-citrulline was used to determine NOS activity as described previously[23,24] with minimal modifications. In brief, the hepatocyte monolayer was homogenized in the lysis buffer as used in Western blotting assay. Samples were centrifuged at 20000 g for 30 min at 4 °C. Supernatants (50 μL) were then incubated with 50 μmol/L 1-[U-14C]-arginine (0.1 μCi, Amersham-operated Pharmacia Biotech, NJ, USA), 2 mmol/L NADPH and 0.45 mmol/L CaCl2 in a final volume of 0.4 mL at 37 °C for 45 min. Immediately after incubation, the mixtures were passed through a 2 mL Dowex 50WX-8 (200-400 dry mesh, H+ form, Aldrich Chemical Company Inc. Milwaukee, USA) column. The flow-through and the eluate (2 mL of water) were collected and quantitated by liquid scintillation spectroscopy (Beckman LS 3801 counter, USA). Samples containing both 1 mmol/L L-NG-nitro-L-arginine methyl ester (L-NAME) and 1 mmol/L EGTA were used as basal controls. The activity of the calcium-dependent enzyme was determined from the difference between [14C] citrulline generated from samples with and without 1 mmol/L EGTA. Retrieved [14C] citrulline values were normalized to total protein.
Statistical analysis
Data were presented as mean±SD and analyzed using the statistical software package SPSS, v12. Probability values <0.05 were taken to indicate statistical significance.
RESULTS
General and morphological changes
Jaundice usually developed 1-3 d after BDL, and receded slowly thereafter in some rats. Body weight dropped within one week after operation, but increased with time after that. Histological examination showed bile duct proliferation, mild edema and acute inflammatory reaction in the enlarged portal areas 3 d after BDL. However, the basic lobular architectural pattern remained intact. By d 7, portal areas became obviously expanded by bile duct proliferation and fibrosis, accompanied with inflammatory reaction, predominant mononuclear cells. Portal tracts were often linked to form a fibrous network, and the parenchymal limiting plates were irregular. By wk 2, bile duct proliferation was even more apparent and often extended into the lobules with proliferated connective tissues. As a result, the normal lobular pattern was disorganized. Three to 4 wk after operation, thick and thin septas formed and bridged the remaining parenchyma into islets of different size. Well-formed nodules were readily seen in most rats. As shown in Figure 1, the histology index for BDL rats increased steadily from d 3 to d 28. In addition, ascites, a marker of uncompensated cirrhosis, was observed in some BDL rats by d 21. By d 28, all BDL rats developed ascites. For sham-operated rats, the liver structure was normal, and no significant changes with time were identified.
Figure 1.
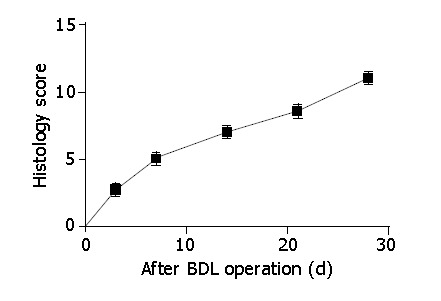
Histology index of BDL livers. The liver sections were read by two pathologists independently. The histology score was based on necrosis, inflammatory infiltration, ductular proliferation as well as fibrosis. Data were presented as mean±SD.
Changes of liver function and systemic NO concentration
As shown in Table 1, bilirubin was dramatically increased to 113 μmol/L 3 d after BDL, but dropped to 60.3 μmol/L by d 14. Thereafter, it was elevated again 3 wk after BDL. ALT was also significantly elevated 3 d after the surgery, but reduced on d 7 by 80%. It maintained that level, which was still significantly higher than that of the sham-operated rats, from wk 2 to wk 4. AST shared a similar pattern to ALT except that it increased further again from wk 3 to wk 4. Unlike AST or ALT absolute values, AST/ALT ratio increased generally with time from d 3 to wk 4, and was significantly correlated with the liver histological changes (r = 0.67, P<0.01). In contrast, no change of liver function with time was found in sham-operated rats. Systemic NO level was indicated by the concentration of its metabolic end products, nitrite and nitrate in plasma. Although it slightly increased in BDL rats on d 3 compared to that in sham-operated rats, no significant alteration was observed until wk 4, when ascites developed in all BDL rats.
Table 1.
Liver function tests and NO levels of rats at different time points after operation (mean±SD).
| Sham-operated | BDL-3d | BDL-1 wk | BDL-2 wk | BDL-3 wk | BDL-4 wk | |
| Bilirubin (mmol/L) | 2.0±0.0 | 113.0±7.1 | 80.0±30.5 | 60.3±5.9 | 96.3±14.8 | 119.4±35.7 |
| ALT (U/L) | 61.9±10.1 | 539.5±150.6 | 102.7±23.8 | 109.3±33.2 | 95.0±11.8 | 134.8±23.3 |
| AST (U/L) | 74.0±26.1 | 691.5±116.7 | 222.3±84.9 | 296.0±127.0 | 233.3±44.1 | 431.7±91.8 |
| NO (mmol/L) | 7.3±0.2 | 8.6±0.3 | 6.9±0.4 | 8.0±0.7 | 7.4±0.9 | 10.5±0.4 |
iNOS gene expression in hepatocytes
iNOS gene expression at mRNA level was determined by competitive RT-PCR, and was identified 3 d after surgery in hepatocytes of BDL rats. By d 7, the signal became intense and clear, indicating a higher level of iNOS mRNA expression. From wk 1 to wk 3, iNOS expression increased with time. For sham-operated rats, a faint iNOS mRNA signal was seen in hepatocytes on d 3. However, the signal reduced significantly with time thereafter, and vanished at the end of wk 2. These results suggested that unlike as in BDL rats, iNOS expression in sham-operated livers was temporal, which could be due to the stress caused by the operation itself (Figure 2A). Further quantitative analysis showed that iNOS mRNA in hepatocytes of BDL rats increased by 0.5-fold from d 3 (0.80±0.13×10-2 attomole/μg RNA) to d 7 (1.17±0.08×10-2 attomole/μg RNA), but by 3.5-fold from wk 1 to wk 2 (4.07±0.33×10-2 attomole/μg RNA, P<0.01 versus wk 1), and by 1.33-fold (5.43±0.01×10-2 attomole/μg RNA, P<0.01 versus week 2) from wk 2 to wk 3. However, it decreased by almost 60% from wk 3 to wk 4 (2.32±0.01×10-2 attomole/μg RNA, P<0.01 versus wk 3), which was still significantly higher than that on d 7 of BDL rats and sham-operated rats at any time point (Figure 2B).
Figure 2.
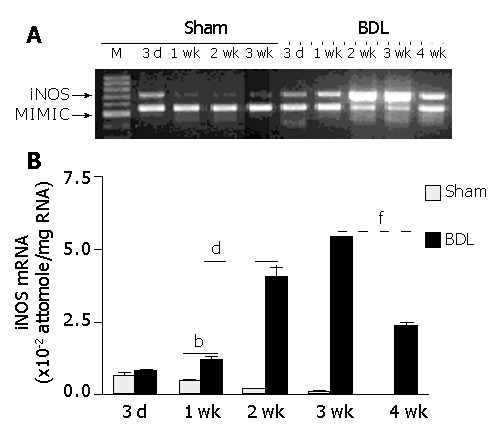
Temporal iNOS mRNA expression in hepatocytes. At different time points after BDL operation, hepatocytes were isolated and iNOS mRNA expression was determined by competitive RT-PCR. A: Typical agarose gel of hepatic iNOS expression (representative of at least three separate experiments). B: Quantitative analysis of iNOS mRNA expression in hepatocytes in the process of cirrhosis development. 3d-4 wk, three days to four weeks after surgery. bP<0.01 for BDL versus sham-operated, 1-3 wk after surgery; dP<0.01 for 2 wk versus all other time points after BDL; fP<0.01 for 4 wk versus 3 wk after BDL.
Expression of iNOS protein in hepatocytes
iNOS protein in hepatocytes was determined by immunoblotting analysis. As shown in Figure 3, iNOS protein was not detected in heptoctytes of sham-operated rats at any time. However, it was observed in hepatocytes of BDL rats, and the expressional levels increased with cirrhosis development. Semi-quantitative analysis showed that iNOS expression increased by 1-fold from d 3 to d 7, and about 40% from wk 1 to wk 2. From wk 2 to wk 3, iNOS protein expression level increased by about 60%. However, from wk 3 to wk 4, iNOS protein expression only increased slightly but not significantly.
Figure 3.
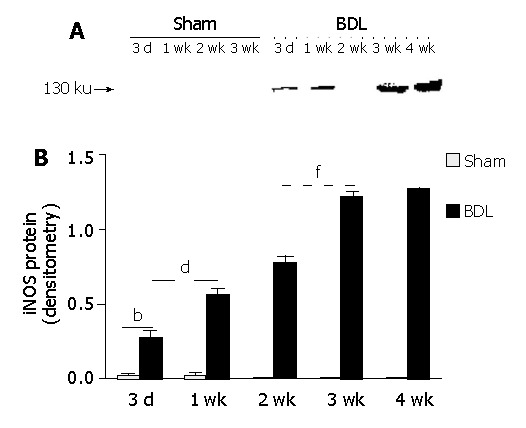
Temporal expression of iNOS protein in hepatocytes. At different time points after BDL, hepatocytes were isolated and iNOS protein was examined by immunoblot analysis. A: Typical result of immunoblotting for iNOS. B: Semi-quantitative analysis of iNOS protein expression. 3d-4 wk, three days to four weeks after surgery. bP<0.01 for sham-operated vs BDL at each time point; dP<0.01 for 2 wk vs all other time points after BDL; fP<0.01 for 3 wk vs all other time points after BDL except wk 4.
NOS activity in hepatocytes
As shown in Figure 4A, the total NOS activity in hepatocytes of BDL rats significantly increased on d 3 compared to that of sham-operated rats. It continued to elevate till 3 wk after BDL surgery. iNOS activity, however was different from total NOS activity in that it decreased from d 3 (57.92±0.88 pmole/mg/min) to d 7 (45.20±3.39 pmole/mg/min, P<0.05 vs d 3). Thereafter, iNOS activity was significantly increased till 3 wk after BDL surgery. No more change of iNOS activity was found from d 21 (100.60±3.90 pmole/mg/min) to d 28 (96.23±1.92 pmole/mg/min, P>0.05 vs d 21). To determine the relationship of iNOS activity with its mRNA and protein expression in hepatocytes during cirrhosis development, we performed a correlation analysis. We found that iNOS activity in hepatocytes of BDL rats was significantly correlated with total NOS activity (r = 0.87, P<0.01), iNOS protein expression (r = 0.87, P<0.01), but not with its mRNA expression (r = 0.47, P>0.05). Further study should be performed to explain the discrepancy between iNOS expression and its enzymatic activity in this model.
Figure 4.
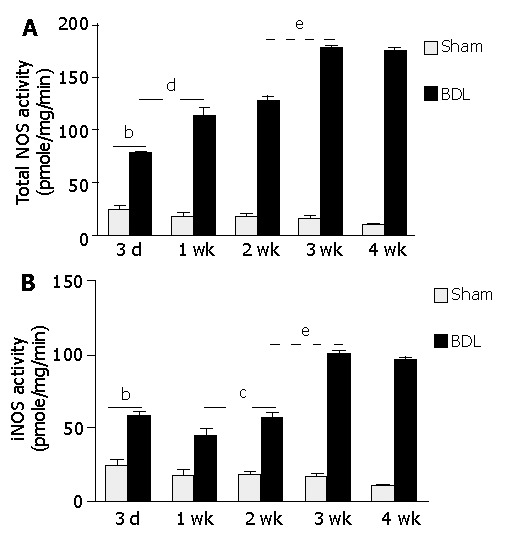
Hepatic NOS activity in the course of cirrhosis development. At different time points after operation, hepatocytes were isolated and NOS activity was determined by L-citrulline generation assay as described in Materials and Methods. 3d-4 wk, three days to four weeks after BDL or sham operation. bP<0.01 for sham-operated versus BDL at each time points; eP<0.05 for 3 wk vs all other time points after BDL except 4 wk. A: Total NOS activity, dP<0.01 for 1 wk vs all other points except 2 wk after BDL; B: iNOS activity, cP<0.05 for 1 wk vs 3 d and 2 wk after BDL.
Besides NOS activity, NO concentration was also determined in the supernatants of hepatocyte culture. However, the absolute values were unsatisfactorily low and no difference was found between different time points.
Correlation among NOS activity, NO levels, liver function, and histology index
In order to define the role of hepatic NO in liver cirrhosis, correlation study was done for NOS activity, liver function, histology index and systemic NO levels. As shown in Table 2, total hepatic NOS activity was correlated significantly with histology score (r = 0.90, P<0.01), iNOS activity (r = 0.86, P<0.01) and AST/ALT ratio (r = 0.56, P<0.05), but not with systemic NO levels, bilirubin, or AST, ALT absolute values. In addition to histology score, iNOS hepatic NOS activity, however had a correlation with bilirubin (r = 0.53, P<0.05), but not with AST/ALT ratio or their absolute values. Moreover, there was no association between iNOS activity and systemic NO levels.
Table 2.
Correlation analysis of NOS activity, NO levels, liver function and histology index.
| NO | AST/ALT | Bilirubin | Total NOS activity | iNOS activity | Histology score | |
| NO | 0.4 | 0.61a | 0.27 | 0.42 | 0.47 | |
| AST/ALT | 0.14 | 0.19 | 0.56a | 0.38 | 0.67b | |
| Bilirubin | 0.017 | 0.51 | 0.23 | 0.53a | 0.2 | |
| Total NOS activity | 0.34 | 0.03 | 0.41 | 0.86b | 0.90b | |
| iNOS activity | 0.12 | 0.17 | 0.04 | 0 | 0.77b | |
| Histology score | 0.08 | 0.006 | 0.47 | 0 | 0.001 |
Above diagonal, Pearson correlation; Below diagonal, P value.
P<0.05;
P<0.01.
DISCUSSION
This time, course study showed that hepatic iNOS was induced 3 d after bile duct ligation, and its expression increased with time towards the late stage of liver cirrhosis. iNOS activity followed a similar pattern to its protein expression, and was correlated significantly with morphological changes of the liver, but not with systemic NO levels, nor with the absolute values of AST, ALT and bilirubin.
NO has been implicated in the pathogenesis of liver cirrhosis. Outside the liver, numerous studies have shown that NO production in the splanchnic as well as systemic circulation increased[5,25-27], induction and expression of iNOS were responsible, at least in part, for it[7,25]. Inside the liver, we found previously that iNOS was induced in hepatocytes of cirrhotic rats. Both its expression and activity significantly increased as compared to its sham-operated controls[10]. Enhanced iNOS expression was also observed in human hepatocytes from patients with alcoholic cirrhosis[28], primary biliary cirrhosis as well as autoimmune hepatitis[29]. These data indicate that iNOS could be upregulated in hepatocytes of at least some, if not all kinds of liver cirrhosis that occurred in patients as well as in experimental animal models.
As cirrhosis is usually a chronic process, we hypothesized that the hepatic iNOS might play an important role in this course ranging from initiation to perpetuation, in addition to its expression and functions in late or advanced stage of cirrhosis as mentioned above. Thus, we designed this time course study to investigate the temporal expression of iNOS in hepatocytes and its contribution to the pathogenesis of cirrhosis. We found that iNOS was induced in hepatocytes at both mRNA and protein levels just 3 d after BDL, when morphologically there were only bile duct proliferation and acute inflammatory reaction in the portal tract. Thereafter, its expression and activity were increased with time till 3 wk after BDL, when advanced cirrhosis started to form, as indicated by the appearance of ascites. No further change of NOS activity was observed from d 21 to d 28 in hepatocytes of BDL rats. This pattern was correlated significantly with histological alterations of the liver in that fibrosis, inflammatory infiltration and ductular proliferation also increased with time to advanced stage. For sham-operated rats, although a faint signal of iNOS mRNA was observed in hepatocytes 3 d after operation, the signal vanished thereafter. Moreover, iNOS protein was not detected at any time point, indicating that the iNOS mRNA expression in sham-operated rats was transient and could be caused by the surgery itself. These results showed that hepatic NO was an important mediator in cirrhosis development. Further study to examine whether it contributes to fibrosis directly or not is warranted.
In order to elucidate the role of hepatic iNOS in cirrhosis development, liver function was determined at different time points after BDL. It was found that bilirubin and AST, ALT were elevated dramatically 3 d after BDL, but decreased thereafter until the late stage of cirrhosis. These results were in agreement with previous reports[30,31] suggesting the liver function is impaired very early in BDL rats, but can be largely compensated by remaining healthy or regenerated hepatocytes after acute phase. The impairment of liver function was aggravated by wk 3 or 4, when advanced cirrhosis developed. Correlation analysis indicated that AST/ALT ratio but not their absolute values was associated with the degree of histological changes of the liver suggesting that AST/ALT ratio may be used as a clinical marker to evaluate biliary cirrhosis. The AST/ALT ratio was also correlated with total hepatic NOS activity implying that hepatic NO may affect liver function in certain mechanisms.
NO levels were also measured in the supernatant of hepatocyte culture as well as plasma. We did not find any difference in NO concentration of the supernatants. In the systemic circulation, NO levels fluctuated from d 3 to d 21, but significant increase was observed only by 4 wk after BDL surgery, when ascites developed in all BDL rats. This was largely in agreement with previous reports, where elevated systemic NO was often found in uncompensated cirrhosis[5,25,27]. Although the mechanisms underlying the systemic NO overproduction are not fully understood yet, enhanced shear stress in response to chronically increased blood flow has been suggested as a major contributor[32-34]. In contrast, no correlation was found between systemic NO levels and hepatic NOS activity, suggesting that NO produced from hepatic iNOS may not be responsible for the systemic NO levels. In addition to hepatocytes, Kupffer cells were the other source of iNOS in cirrhosis[10,29]. However, it constituted only 15% of liver by cell numbers[15], and iNOS expression was even lower than that in hepatocytes[10]. Thus, the study of iNOS function in liver has been focused largely on its induction in hepatocytes[35].
In summary, we found that iNOS was induced in hepatocytes soon after BDL; its expression and activity increased with time in cirrhosis development ranging from initiation to perpetuation. This was correlated with the morphological changes of liver, as well as AST/ALT ratio, but not with systemic NO levels, which significantly increased only in the late stage of cirrhosis. Further study is necessary to define the function of hepatic NO during cirrhosis development.
Footnotes
Edited by Wang XL
References
- 1.Lowenstein CJ, Dinerman JL, Snyder SH. Nitric oxide: a physiologic messenger. Ann Intern Med. 1994;120:227–237. doi: 10.7326/0003-4819-120-3-199402010-00009. [DOI] [PubMed] [Google Scholar]
- 2.Moncada S, Palmer RM, Higgs EA. Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev. 1991;43:109–142. [PubMed] [Google Scholar]
- 3.Wiest R, Groszmann RJ. The paradox of nitric oxide in cirrhosis and portal hypertension: too much, not enough. Hepatology. 2002;35:478–491. doi: 10.1053/jhep.2002.31432. [DOI] [PubMed] [Google Scholar]
- 4.Cahill PA, Redmond EM, Hodges R, Zhang S, Sitzmann JV. Increased endothelial nitric oxide synthase activity in the hyperemic vessels of portal hypertensive rats. J Hepatol. 1996;25:370–378. doi: 10.1016/s0168-8278(96)80124-3. [DOI] [PubMed] [Google Scholar]
- 5.Guarner C, Soriano G, Tomas A, Bulbena O, Novella MT, Balanzo J, Vilardell F, Mourelle M, Moncada S. Increased serum nitrite and nitrate levels in patients with cirrhosis: relationship to endotoxemia. Hepatology. 1993;18:1139–1143. [PubMed] [Google Scholar]
- 6.Hori N, Wiest R, Groszmann RJ. Enhanced release of nitric oxide in response to changes in flow and shear stress in the superior mesenteric arteries of portal hypertensive rats. Hepatology. 1998;28:1467–1473. doi: 10.1002/hep.510280604. [DOI] [PubMed] [Google Scholar]
- 7.Martin PY, Xu DL, Niederberger M, Weigert A, Tsai P, St John J, Gines P, Schrier RW. Upregulation of endothelial constitutive NOS: a major role in the increased NO production in cirrhotic rats. Am J Physiol. 1996;270:F494–F499. doi: 10.1152/ajprenal.1996.270.3.F494. [DOI] [PubMed] [Google Scholar]
- 8.Niederberger M, Martin PY, Ginès P, Morris K, Tsai P, Xu DL, McMurtry I, Schrier RW. Normalization of nitric oxide production corrects arterial vasodilation and hyperdynamic circulation in cirrhotic rats. Gastroenterology. 1995;109:1624–1630. doi: 10.1016/0016-5085(95)90652-5. [DOI] [PubMed] [Google Scholar]
- 9.Ros J, Jiménez W, Lamas S, Clària J, Arroyo V, Rivera F, Rodés J. Nitric oxide production in arterial vessels of cirrhotic rats. Hepatology. 1995;21:554–560. [PubMed] [Google Scholar]
- 10.Wei CL, Khoo HE, Lee KH, Hon WM. Differential expression and localization of nitric oxide synthases in cirrhotic livers of bile duct-ligated rats. Nitric Oxide. 2002;7:91–102. doi: 10.1016/s1089-8603(02)00103-9. [DOI] [PubMed] [Google Scholar]
- 11.Rockey DC, Chung JJ. Reduced nitric oxide production by endothelial cells in cirrhotic rat liver: endothelial dysfunction in portal hypertension. Gastroenterology. 1998;114:344–351. doi: 10.1016/s0016-5085(98)70487-1. [DOI] [PubMed] [Google Scholar]
- 12.Shah V, Toruner M, Haddad F, Cadelina G, Papapetropoulos A, Choo K, Sessa WC, Groszmann RJ. Impaired endothelial nitric oxide synthase activity associated with enhanced caveolin binding in experimental cirrhosis in the rat. Gastroenterology. 1999;117:1222–1228. doi: 10.1016/s0016-5085(99)70408-7. [DOI] [PubMed] [Google Scholar]
- 13.Rockey DC, Chung JJ. Regulation of inducible nitric oxide synthase and nitric oxide during hepatic injury and fibrogenesis. Am J Physiol. 1997;273:G124–G130. doi: 10.1152/ajpgi.1997.273.1.G124. [DOI] [PubMed] [Google Scholar]
- 14.Tu W, Kitade H, Satoi S, Zhang ZT, Kaibori M, Kwon AH, Kamiyama Y, Okumura T. Increased nitric oxide production in hepatocytes is involved in liver dysfunction following obstructive jaundice. J Surg Res. 2002;106:31–36. doi: 10.1006/jsre.2002.6436. [DOI] [PubMed] [Google Scholar]
- 15.Wanless IR. Physioanatomic considerations. In: Schiff ER, Sorrell MF, Maddrey WC, eds. Schiff’s Diseases of the Liver. Lippincott Williams Wilkins; 2003. pp. 18–50. [Google Scholar]
- 16.Kountouras J, Billing BH, Scheuer PJ. Prolonged bile duct obstruction: a new experimental model for cirrhosis in the rat. Br J Exp Pathol. 1984;65:305–311. [PMC free article] [PubMed] [Google Scholar]
- 17.Lee SS, Sharkey KA. Capsaicin treatment blocks development of hyperkinetic circulation in portal hypertensive and cirrhotic rats. Am J Physiol. 1993;264:G868–G873. doi: 10.1152/ajpgi.1993.264.5.G868. [DOI] [PubMed] [Google Scholar]
- 18.Ishak K, Baptista A, Bianchi L, Callea F, De Groote J, Gudat F, Denk H, Desmet V, Korb G, MacSween RN. Histological grading and staging of chronic hepatitis. J Hepatol. 1995;22:696–699. doi: 10.1016/0168-8278(95)80226-6. [DOI] [PubMed] [Google Scholar]
- 19.Guo FH, De Raeve HR, Rice TW, Stuehr DJ, Thunnissen FB, Erzurum SC. Continuous nitric oxide synthesis by inducible nitric oxide synthase in normal human airway epithelium in vivo. Proc Natl Acad Sci USA. 1995;92:7809–7813. doi: 10.1073/pnas.92.17.7809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Titos E, Clària J, Bataller R, Bosch-Marcé M, Ginès P, Jiménez W, Arroyo V, Rivera F, Rodés J. Hepatocyte-derived cysteinyl leukotrienes modulate vascular tone in experimental cirrhosis. Gastroenterology. 2000;119:794–805. doi: 10.1053/gast.2000.17831. [DOI] [PubMed] [Google Scholar]
- 21.Gilliland G, Perrin S, Blanchard K, Bunn HF. Analysis of cytokine mRNA and DNA: detection and quantitation by competitive polymerase chain reaction. Proc Natl Acad Sci USA. 1990;87:2725–2729. doi: 10.1073/pnas.87.7.2725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hon WM, Moochhala S, Khoo HE. Adenosine and its receptor agonists potentiate nitric oxide synthase expression induced by lipopolysaccharide in RAW 264.7 murine macrophages. Life Sci. 1997;60:1327–1335. doi: 10.1016/s0024-3205(97)00078-7. [DOI] [PubMed] [Google Scholar]
- 23.Brown JF, Tepperman BL, Hanson PJ, Whittle BJ, Moncada S. Differential distribution of nitric oxide synthase between cell fractions isolated from the rat gastric mucosa. Biochem Biophys Res Commun. 1992;184:680–685. doi: 10.1016/0006-291x(92)90643-y. [DOI] [PubMed] [Google Scholar]
- 24.Salter M, Knowles RG, Moncada S. Widespread tissue distribution, species distribution and changes in activity of Ca(2+)-dependent and Ca(2+)-independent nitric oxide synthases. FEBS Lett. 1991;291:145–149. doi: 10.1016/0014-5793(91)81123-p. [DOI] [PubMed] [Google Scholar]
- 25.Morales-Ruiz M, Jiménez W, Pérez-Sala D, Ros J, Leivas A, Lamas S, Rivera F, Arroyo V. Increased nitric oxide synthase expression in arterial vessels of cirrhotic rats with ascites. Hepatology. 1996;24:1481–1486. doi: 10.1053/jhep.1996.v24.pm0008938184. [DOI] [PubMed] [Google Scholar]
- 26.Niederberger M, Ginès P, Tsai P, Martin PY, Morris K, Weigert A, McMurtry I, Schrier RW. Increased aortic cyclic guanosine monophosphate concentration in experimental cirrhosis in rats: evidence for a role of nitric oxide in the pathogenesis of arterial vasodilation in cirrhosis. Hepatology. 1995;21:1625–1631. [PubMed] [Google Scholar]
- 27.Sieber CC, Lopez-Talavera JC, Groszmann RJ. Role of nitric oxide in the in vitro splanchnic vascular hyporeactivity in ascitic cirrhotic rats. Gastroenterology. 1993;104:1750–1754. doi: 10.1016/0016-5085(93)90655-v. [DOI] [PubMed] [Google Scholar]
- 28.McNaughton L, Puttagunta L, Martinez-Cuesta MA, Kneteman N, Mayers I, Moqbel R, Hamid Q, Radomski MW. Distribution of nitric oxide synthase in normal and cirrhotic human liver. Proc Natl Acad Sci USA. 2002;99:17161–17166. doi: 10.1073/pnas.0134112100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sanz-Cameno P, Medina J, García-Buey L, García-Sánchez A, Borque MJ, Martín-Vílchez S, Gamallo C, Jones EA, Moreno-Otero R. Enhanced intrahepatic inducible nitric oxide synthase expression and nitrotyrosine accumulation in primary biliary cirrhosis and autoimmune hepatitis. J Hepatol. 2002;37:723–729. doi: 10.1016/s0168-8278(02)00266-0. [DOI] [PubMed] [Google Scholar]
- 30.Ni Y, Lukito G, Marchal G, Cresens E, Yu J, Petré C, Baert AL, Fevery J. Potential role of bile duct collaterals in the recovery of the biliary obstruction: experimental study in rats using microcholangiography, histology, serology and magnetic resonance imaging. Hepatology. 1994;20:1557–1566. doi: 10.1002/hep.1840200627. [DOI] [PubMed] [Google Scholar]
- 31.Parola M, Leonarduzzi G, Robino G, Albano E, Poli G, Dianzani MU. On the role of lipid peroxidation in the pathogenesis of liver damage induced by long-standing cholestasis. Free Radic Biol Med. 1996;20:351–359. doi: 10.1016/0891-5849(96)02055-2. [DOI] [PubMed] [Google Scholar]
- 32.García-Cardeña G, Fan R, Shah V, Sorrentino R, Cirino G, Papapetropoulos A, Sessa WC. Dynamic activation of endothelial nitric oxide synthase by Hsp90. Nature. 1998;392:821–824. doi: 10.1038/33934. [DOI] [PubMed] [Google Scholar]
- 33.Pateron D, Tazi KA, Sogni P, Heller J, Chagneau C, Poirel O, Philippe M, Moreau R, Lebrec D. Role of aortic nitric oxide synthase 3 (eNOS) in the systemic vasodilation of portal hypertension. Gastroenterology. 2000;119:196–200. doi: 10.1053/gast.2000.8554. [DOI] [PubMed] [Google Scholar]
- 34.Tazi KA, Barrière E, Moreau R, Heller J, Sogni P, Pateron D, Poirel O, Lebrec D. Role of shear stress in aortic eNOS up-regulation in rats with biliary cirrhosis. Gastroenterology. 2002;122:1869–1877. doi: 10.1053/gast.2002.33586. [DOI] [PubMed] [Google Scholar]
- 35.Clemens MG. Nitric oxide in liver injury. Hepatology. 1999;30:1–5. doi: 10.1002/hep.510300148. [DOI] [PubMed] [Google Scholar]