

Abstract
AIM: Although epidemiological and experimental studies validate influence of genetic, environmental and dietary factors in the causation of various types of cancers including colon, results from all these sources are inconclusive. Hypothesizing that high fat diet and obesity are among the major predisposing factors in the incidence of colon cancer, we evaluated the role of diet constituted with food material derived from a tropical plant, Tamarindus indica Linn (TI).
METHODS: A two part randomized double-blind study was conducted employing inbred Swiss albino mice from a single generation for the whole investigation. One day-old neonates (n = 12) were subcutaneously administered with monosodium glutamate (MSG) to induce obesity (OB). At weaning these animals were maintained on modified AIN-76 diet supplemented with 10% TI and 10% fat bolus (w/w, TIFB) for 8 wk. Subsequently, in the second part of study, four groups of animals belonging to the same generation, age and gender (n = 12 per group), were maintained on: AIN-76 control diet (CD); AIN-76 mixed with 10% TI extract (TI); and, mixed with 10% TI and 10% FB (TIFB) for 8 wk, to determine intestinal crypt cell proliferation, functionally-specific enzyme activities, fermentation profile, and energy preferences.
RESULTS: We observed a significant increase in the crypt cell production rate in distal colonic segment of experimental animals when compared with the controls. This segment also contained significantly low butyrate levels compared to control and TIFB groups. All the experimental groups showed a gross decrease in the enzyme activities viz., succinate dehydrogenase, acid-galactosidase and dipeptidyl amino peptidase IV demonstrating pathological stress caused by the test regimens, and an altered metabolic flux in the cellular environment.
CONCLUSION: We have demonstrated a cumulative response to the three dietary factors, one of which (TI) is reported, herein, for the first time to modulate kinetics of large intestinal mucosa, contributing to total risk posed by these test agents.
Keywords: Large intestinal mucosa, Kinetics, Tamarindus indica Linn, Diet
INTRODUCTION
Dietary factors have been estimated to be responsible for up to 40% of cancers including those of the gastrointestinal tract (GIT)[1-4]. Numerous life style factors have also been included such as, high fat diet consumption, inadequate physical activity and obesity[5-7]. High fat/low fiber diet has been found to have greater influence on the development of colon cancer[8]. Several studies have been conducted worldwide to relate obesity with cancer risk especially of the large intestine[9-13].
Tamarind forms an important ingredient of Indian cookery. Tamarind pulp powder (hereafter referred as TI) as used in the present study is prepared by concentrating, drying and milling the pulp into a powdered form. It is one of the convenient food products produced commercially by several manufacturers in India. Manjunath et al[14] reported that on average the total solid content of tamarind pulp powder varied between 18.6 and 25%, and acidity between 8.7 and 11.1% with an average value of 9.9% (as tartaric acid). The moisture content ranges between 3.5 and 8.8%. Digestible starch is the major ingredient in tamarind pulp powder (20-41%). In some recent reports, the presence of nicotinic acid (about 5%), and primary amines (148.8 ppm) were discussed with emphasis on precursor carcinogens that added up to the list of environmental hazards to human beings[15-17]. The average composition of the dried pulp powder of TI is given in Table 1.
Table 1.
Proximate composition of dried pulp of tamarind fruit.
| Constituents | Proximate composition of the dried pulp powder (%)1 |
| Moisture | 3.5-8.8 |
| Protein | 1.7-2.4 |
| Total sugars | 15.8-25.0 |
| Ash | 2.1-3.2 |
| Free tartaric acid | 8.7-11.1 |
| Starch | 20-41.3 |
1Source: Reference[14].
In view of the above literature, we wondered if the type of starch and high content of sugar present in the dried pulp of T. indica (hereafter referred as TI) might exert its effect on the colonic cell kinetics, either positively or negatively, when fed to the experimental animals chronically as a mix with AIN-76 control diet ad libitum. Influence of dietary sugars/starch and microbial contents on colonic cell proliferation has been studied extensively[18]. It is likely that non-digestible starch increases apoptotic cell death and inhibits cellular growth in colonic mucosa. DNA damage is considered to play a crucial role in the induction of almost every type of cancer including that of colon. Studies have been carried out until recently on the DNA damage events by the exposure to various forms of starch using animal models[19,20].
In addition, we attempted to evaluate the influence of TI- derived components with fat bolus in experimental animals post-challenged with neonatal obesity. It is neither evident from the previous literature that TI might trigger genesis of any type of cancer in humans epidemiologically, nor is it experimentally demonstrated to cause any inductive/promotive response towards carcinogenesis. However, Block[21] and Patterson[22] reported that plant-derived components had significant impact on the mucosal properties in the entire gastrointestinal tract.
Hence, a protocol was designed to evaluate whether TI could act independently on the colonic mucosal architecture of the host system, and/or facilitate the fatty diet and/or obesity - mediated cellular damage and upregulate the proliferation rate, a basic prerequisite of neoplastic transformation. For the present communication, crypt cell proliferation rate (CCPR) and tritiated thymidine incorporation assays, biochemical enzyme profile, short chain fatty acids profile (SCFAs) and energy preferences such as lactate, were chosen to address possible effects of TI, per se as well as in combination with the well-established predisposing factors, on colonic mucosa of Swiss albino mice.
MATERIALS AND METHODS
Chemicals and isotopes
All the chemicals and solvents used in this study were of analytical grade. The isotope [3H] - thymidine was purchased from Board of Radiation and Isotope Technology, Bombay, India. Monosodium glutamate (marketed as “aginomoto salt”) and the dried pulp of tamarind (marketed as “dry tam- without mess”) were purchased from the local market.
Preparation of T. indica fruit pulp extract
The dried fruit pulp of T. indica weighing 100 g was taken in conical flask and about 500 mL of boiled water was added and allowed to soak for 1 h. The cooled, aqueous extract[16] was filtered through a sieve (100 mesh) and used as an ingredient in the synthetic diet. The particular preparation method logically simulated the conditions applied by the Indian cookery system.
Experimental design
Swiss albino mice were purchased from Tamil Nadu Veterinary and Animal Science University (TNVASU), Chennai, India, and bred in the department’s animal house. Inbred animals belonging to a single generation were selected for all the experimental conditions. In the first part of investigation, 75 animals from the third day of their birth were subcutaneously administered with 100 μL of 20 mg/mL MSG at two spots, regularly for eleven days[23]. A mortality rate of about 60% was observed during the induction of obesity and remaining 40% live animals developed obesity successfully, eighteen animals of which were chosen for the present study plan (OB), and therefore, caged separately. They were fed the AIN-76 modified diet[24], constituted with 10% (w/w) TI, 10% (w/w) fat bolus and 2% cholesterol, as designated for the TIFB-treated group.
For the second set of study, animals were divided into four groups (twenty four animals in each group). Control animals received controlled AIN-76 synthetic diet (CD). Animals belonging to TI - treated group (TI) were maintained on the synthetic diet mixed with 10% TI. Further, for TIFB - treated group (TIFB) of animals, the diet was mixed with 10% TI, 2% cholesterol, and 10% fat bolus.
Table 2 describes the diet composition of control and experimental groups of animals. Adjustment on caloric value of the diet was done with respect to the chemical composition of TI fruit extract, saturated animal fat and cholesterol. Choline and corn oil were excluded from the dietary regimens so as to analyze the effects of pure animal fat and cholesterol on the cellular membrane integrity without the interference of choline and other lipids. Treatment of animals with the present study setup was within the permissible criteria proposed by Government of India’s Ministry of Social Justice and Empowerment, and Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), (as published recently in Volume 19, Issue 23, November 9-22, 2002 Frontline). The rules and laws were followed from Poynter Center for the Study of Ethics and American Institutions, Indiana University, 410 N. Park Ave., Bloomington, IN, USA.
Table 2.
Composition of the diet administered to control and experimental animals.
| Constituents | Control diet | Experimental diet |
| Caesin (%) | 23.8 | 23.8 |
| DL- methionine (%) | 0.5 | 0.5 |
| Corn starch (%) | 30 | 22.7 |
| Sucrose (%) | 30 | 27.5 |
| Cellulose (%) | 6.5 | 6.5 |
| Corn oil (%) | 5 | 5 |
| Salt mix (%) | 3 | 3 |
| Vitamin mix (%) | 1 | 1 |
| Choline bitartrate (%) | 0.2 | 0.2 |
| TI (10%)1- mixed | ||
| Sucrose (%) | - | 2.5 |
| Tartaric/Malic/Citric acid (%) | - | 3.3 |
| Starch (%) | - | 4 |
| Animal fat (%)2 - mixed | - | 10 |
| Cholesterol (%) | - | 2 |
1TI was mixed (10%) in the total diet and compensated with corn starch and sucrose of the control diet for the nutrients composition for preparing experimental diet 2Choline bitartrate and corn oil were excluded from the dietary regimens of groups3 and 4.
Crypt cell proliferation rate assay
At intervals of 2, 4 and 8 wk, 6 animals from the experimental and control groups were given 1 mg/kg of colchicine intraperitoneally. The tissue samples were excised from different parts of the large intestine. The number of arrested metaphases increased with time after injection. To prevent variation in the kinetic results due to diurnal changes[25], in each group, the first animal was killed 30 min after injection with colchicine and thereafter one animal was killed every 15 min up to 90 min. Three segments were taken viz., cecum, ascending colon, and descending colon. Fixing of the segments was carried out with glacial acetic acid for 24 h and stored in 750 mL/L ethanol. For measurement of the crypt cell proliferation rate, the tissue specimens were rehydrated on an alcohol gradient and hydrolyzed in 1 N HCl for 10 min and stained using periodic acid Schiff’s reagent. The focal areas of crypts exhibiting abnormal characteristics were microscopically dissected and squashed under cover slips. The number of arrested metaphases was counted in ten crypts per sample. The mean metaphase count per crypt was plotted against the time that the samples were taken (five points, 30 to 90 min). A linear regression curve was drawn for each site in each group, and from the slope of the graph mean CCPR/crypt/h was calculated and plotted.
3H-Thymidine incorporation assay
Control and experimental animals (six in each) after 2, 4 and 8 wk of experimental period were taken for this assay. For the determination of [3H]-dThd into DNA, 100 μci/kg body weight of [3H]-dThd (3H-(methyl) thymidine; specific activity 18 μci/mmole) was injected intraperitoneally to the animals of experimental and control groups. Animals were sacrificed by cervical dislocation after 3 h of injection of [3H]-dThd. The three colonic segments were excised and placed in saline at 4 °C. During the analysis, the mucosa was scraped and DNA content was extracted and counted as described previously[26-28]. The results are represented as counts per minute (cpm) per μg DNA.
Measurement of SCFAs and lactate
Colonic contents were collected at respective intervals from 6 animals in each group. The contents were acidified with 500 mL/L sulphuric acid solution. The solution was then extracted with 2 mL ether and 1 μL of the extract was injected into a capillary column containing a column stationary phase of 10% fatty acids, of 30 m long (inner diameter: 0.32 mm, thickness: 0.5 μm, split, 1:80). A Hewlett Packard gas chromatograph (Model 6890) equipped with a flame ionization detector was used. A column temperature of 70 °C and a detector temperature of 230 °C were used. Peaks of SCFAs were confirmed by running known standards under similar conditions. Integration of the peaks was carried out using the HP 3389A software; values are represented as percentage distribution of each SCFA in the profile. Lactate levels in the control and experimental samples were measured by UV- enzymatic method as described in the commercial kit (Boehringer, Mannheim, Germany) and calculated as μmol/g wet content by establishing the standard curve.
Enzymatic analysis
In this study, activities of mitochondrial succinate dehydrogenase (SDH), brush border dipeptidyl amino peptidase IV (DAP IV) and lysosomal acid β-galactosidase (acid β-gal) were measured by histochemical quantitation (Table 3). Colon tissues were removed and cut opened longitudinally and washed thoroughly with physiological saline. Serial 10 μm cryostat sections of the colon tissues were mounted on the cover slips. Histochemical quantitation of the enzymes was carried out using specific substrate solutions and stains according to the established procedures[29-31].
Table 3.
Activities of succinate dehydrogenase, acid-B- galactosidase and dipeptidyl amino peptidase IV (values are expressed as absorbance units, mean±SE; n = 6).
| Enzymes | Crypt segments | CD | TI | TIFB | OB |
| SDH | Luminal epithelium | 0.596±0.053 | 0.533±0.038 | 0.512±0.053a | 0.524±0.067a |
| DAP IV | Luminal epithelium | 0.178±0.027 | 0.163±0.008 | 0.156±0.019 | 0.125±0.014a |
| Basal crypt epithelium | 0.196±0.012 | 0.179±0.003 | 0.162±0.005 | 0.151±0.030 | |
| Acid-b-Gal | Luminal epithelium | 0.526±0.046 | 0.443±0.035 | 0.381±0.028a | 0.314±0.029a |
| Basal crypt epithelium | 0.592±0.061 | 0.527±0.036 | 0.493±0.058 | 0.397±0.011b | |
| Apical crypt epithelium | 0.863±0.067 | 0.639±0.057c | 0.587±0.009c | 0.452±0.065c |
aP<0.05 vs CD in the crypt segment of luminal epithelium; bP<0.01 vs CD in the crypt segment of basal crypt epithelium; cP<0.05 vs CD in the crypt segment of apical crypt epithelium.
Statistical analysis
In the calculation of CCPR, linear regression analysis was carried out to determine the correlation coefficient. The standard deviations of the slopes were also calculated but, as is normal practice in this analysis, they were not quoted[32]. Cell kinetics data between each experimental group and the control group for each segment were compared for statistical significance using two tailed Student’s t-test. Chi-square test was applied to determine the significance of counts of tritiated thymidine incorporation into DNA. ANOVA was employed for calculating the statistical significance of the means of SCFA and lactate levels obtained for the experimental and control groups. Histochemical analysis was performed on rank data as low (1), moderate (2), and high (3) levels of staining. ANOVA was used to get the statistical significance with respect to the colonic segments and treatment groups. Statistical analyses were also aided by MicroCal Origin software (version 2.9, USA).
RESULTS
CCPR assay
Linear regression curve (Figure 1) was drawn by taking the mean number of cells produced at five different time intervals, 30 min after the administration of colchicine. It might be noted that each CCPR/crypt/h measurement at different sites at their specific exposure levels was carried out in a similar manner. Figures 2A, B, C show the crypt cell production rate in cecum, ascending colon and descending colon tissues isolated from the experimental and normal animals at wk 2, 4, and 8. It was observed that the rate of cell proliferation in the TI-supplemented group was approximately equal to its normal counterpart, up to 4 wk in all the three segments of GIT. Combination of TI and saturated fat (TIFB) did not influence the production of cells significantly at wk 2 in all the tissue sections. However, at four weeks of study, this group had a significantly greater rate of cell production compared to the controls. While for the OB treated animals, influence of obese condition on the production rate of crypt cells in the presence of TIFB-supplemented diet was evidently significant in all the three segments four weeks onwards.
Figure 1.
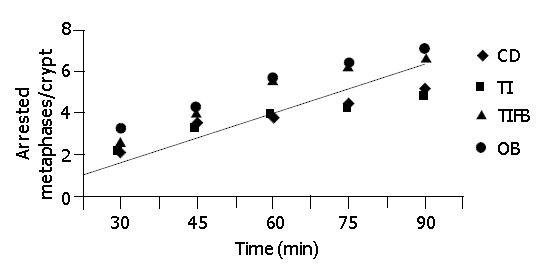
Linear relationship of metaphase arrest to time used to calculate CCPR.
Figure 2.
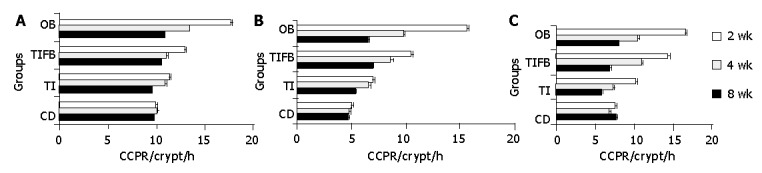
Crypt cell production rate/Crypt/h in caecum at (A), ascending colon (B) and descending colon (C) tissues isolated from experimental and control groups of animals (mean±SE) at 2, 4 and 8 wk treatment. Significance was seen between CD and the experimental groups TI, TIFB and OB.
[3H]-dThd incorporation assay
Figures 3A, B, C demonstrate the amount of [3H] - dThd incorporation into the colonocyte DNA of cecum, ascending and descending colon tissues at three different exposure levels. At wk 8, the OB animals exhibited a significantly increased incorporation in the mucosa obtained from all three segments followed by TIFB-treated group in comparison to their normal counterparts. However, on a par with the analysis of CCPR, TI- fed animals presented reduced levels of incorporation in all three tissue sections up to 4 wk. Nevertheless this group was indicated to incorporate significant numbers of radiolabeled precursor in the colonocyte DNA of ascending and descending segments at eight weeks as compared to controls.
Figure 3.
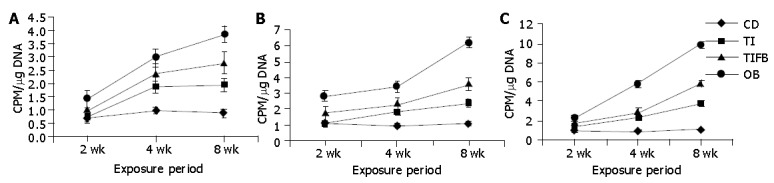
Incorporation of [3H] – dThd into the DNA of cecum (A), ascending colon (B) and descending colon (C) tissues at different exposure periods in experimental and control groups (mean±SE). Significance was seen between CD and the experimental groups TI, TIFB and OB.
Fermentation profile
TI - per se fed animals showed a significant decrease in the levels of butyrate and lactate at 8 wk, in the ascending and descending colon segments, while there was no significant change at 2 and 4 wk of exposure (Figure 4). Acetate and propionate were as equal as that of control samples. However, insignificant changes were observed in the cecum tissue of TI -fed animals compared to control group with regard to all of these SCAFs. The TIFB-and OB-induced groups produced significantly increased amount of lactate in the two colon segments, whereas, the cecum tissues of these groups producd more propionate and butyrate. Acetate concentrations in all the three segments from these two treated groups were not altered. The changes in these two groups were significant at 4 wk and increases continued thereafter.
Figure 4.
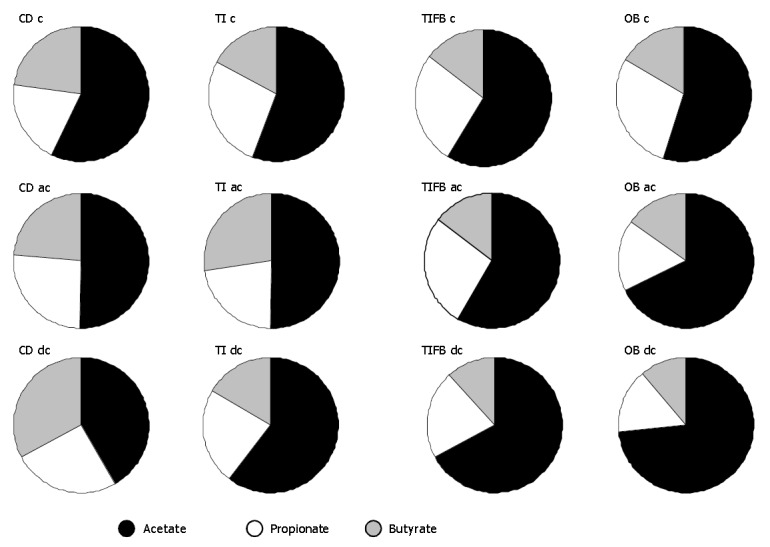
Mean (%) of acetic, propionic and butyric acids in the mucosal contents of cecum (c), ascending colon (ac) and descending colon (dc) tissue segments isolated from mice of CD, TI, TIFB and OB groups. Results were drawn from the total mean(%) distribution of SCFAs throughout the experimental period (n = 6).
In the cecum, lactate concentrations in the animals of control and experimental groups showed a positive correlation with other SCFAs, indicating the fate of microbial content of the mucosa at different dietary exposures (Figure 5). TI fed group showed an insignificant change in its lactate content compared to that of controls at 2 wk. While this group showed a significant elevation in the levels of lactate at wk 4 and 8 (P<0.004 and P<0.016, respectively). Ascending and descending colon tissues showed elevated lactate concentrations at 2 wk (P<0.047, and P<0.002, respectively). However, decreased lactate production was evident in these two segments later on (P = NS). The results were attributed to the amount of starch that escaped digestion producing high lactate at about 2 wk. The animals were seemed to get acclimatized to the dietary conditions thereafter. Upon analysis of the same samples isolated from the other two groups, viz., TIFB and OB, there seemed to be a significant production of lactate from 4 wk onwards in all the three segments indicating a close association with the high acetate levels (about 73%) and propionate levels (around 45% in the cecum specifically) found in these experimental animals.
Figure 5.
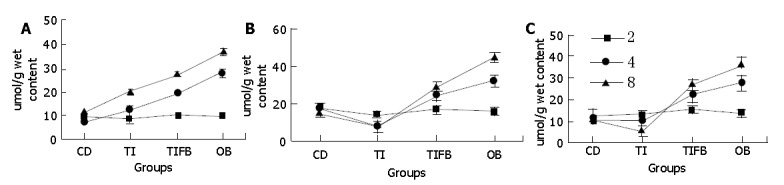
Levels of lactate in mice fed with various dietary factors in CD, TI, TIFB and OB groups for 2, 4 and 8 wk of test periods in the isolated cecum (A), ascending colon (B) and descending colon (C). Values are represented as μmol/g wet mucosal content, mean±SE.
Enzyme activities
The selection of the three enzymes was dependent upon the occurrence of these enzymes in colonic crypts. The distribution of these enzymes in the crypt follows a function-specific pattern. The activity of succinate dehydrogenase is pronounced in the luminal epithelium of the colonic crypt, whereas the basal epithelium contains DAP IV activity and the activity of acid β-gal occupies the apical crypt at a high level. Distribution of reactive DAP IV is also seen in the luminal crypt. Similarly, a moderate level distribution of acid β-gal is seen in the luminal and basal portions of the colonic crypt[33]. Histochemical analysis of the enzymes indicated a gross decrease in the activities of all the three enzymes along the entire large intestine suggesting impaired metabolic activities of the colonic mucosa, due in part to the dietary exposures. The effect was more prominent when observed critically in the OB - group, as there was an additive influence of TIFB in association with obese condition. Distribution patterns of enzyme activities in the tissue sections of cecum region of TI per se - treated group did not reveal much variation in their activities at all the three time points. However, in combination with other two factors, viz., FB and OB, it was shown to have an impact as a positive stimulator. There was of course, a reduction of enzyme activities observed in the other two segments after 4 wk of treatment with TI.
DISCUSSION
Our investigation has shown that dietary factors had a significant impact on large intestine of experimental animals. It was critically noted that all the parameters included in this study were dependent on each other. Generally, epithelial restitution is a basic physiologic response elicited against any kind of mucosal damage causing an insult to the epithelial layer. Rate of cell proliferation was increased in such instances and normal differentiated cells migrated towards and replaced the damaged epithelia. Crypt fission occurred under these conditions leading to increased numbers of crypts thereby stabilizing mode of spread of mutated clones in the mucosa[34]. Our findings go in parallel with the literature cited with respect to colonic epithelial cell proliferation. Crypt cell proliferation rate was increased in all the three segments of OB- and TIFB- treated animals with a positive correlation with decreased levels of butyrate and enzyme activities. It was reported that butyrate acted as an inducer of apoptosis under in vivo conditions and in contrast it was implied as an inducer of cellular differentiation under in vitro conditions[35-37]. A biopsy obtained from human cecum showed a response to SCFA supplementation with an increase in cell proliferation when cultured in vitro[38]. In our study butyrate levels went down in the groups of TIFB and OB at an early stage compared to the TI- per se treated group and controls, leading to uncontrolled proliferation of mucosal epithelium. Earlier reports also suggested similar hypotheses[6,12]. It was also noted that distribution of mitotic cells (in OB and TIFB-treated groups) was seen in the upper 40% of the crypt, which was considered to be a sensitive biomarker of cancer risk[39].
Interestingly, in the present study, alterations in the colonic mucosa were evident due in part to the high digestible starch and acidic content of TI which might have affected the microfloral content of the colon as shown by altered short chain fatty acids profile and impaired enzyme profile, thereby aggravating the adverse effects of fatty diet and obese condition. Large bowel microfloral contents were found to be very low when the cecum and distal colon tissues were weighed in comparison to that of control tissues, as it is evident[40,41] that microbial digestion of the partially digested food that escape from the small intestine are ultimately digested by the microflora of the cecum (wherein the digested material stays for a maximum time period, approximately 4 h in the case of mouse). In addition, our suggested hypothesis is in agreement with the findings of McIntyre et al[42], who suggested the beneficial effects of increased butyrate production in the large bowel due to consumption of fiber-enriched diet which helps protect the rat from developing colon cancer. In our study, lactate production showed an indifferent pattern of results with a close similarity to other SCFAs profiles. The profile indicated that the amount of lactate produced in the TIFB and OB - groups was a reflection to some extent of high bacterial glycolysis in combination with a high proliferative index of mucosal cells as noted with an increased CCP rate and radioactive labeling. Other reports of Fleming et al[43] and Caderni et al[44] also suggested these two phenomena in the mucosa under carcinogenic/pathological conditions induced by high fat diet.
Further, the rate of proliferation of the crypt epithelium and radiolabeled precursor incorporated DNA content in the cells of colon tissues isolated from OB and TIFB- fed groups, were increased in a linear fashion from 4 to 8 wk . It is speculated that both physiological and biochemical parameters seem to be affected significantly after 4 wk attributing to the fact that acclimatization of animals to their respective diets ad libitum might take nearly 2 wk. Previous reports also supported our findings that ad libitum feeding of high-fat diet enhanced formation of aberrant crypt/ foci in experimental animals after four weeks[45-47]. Patients with familial polyposis coli, clinically manifested in association with obesity, have been indicated to have increased thymidine - labeled cells in the upper third of the morphologically normal crypt[45]. It was demonstrated by another scientific group that specified pathogen-free gut conditions might have a positive impact on the development of polyps[48]. Association of fatty diet consumption and obesity has been studied extensively[5]. Ogus et al[49] have described that high fat diet - induced obesity in normal and transgenic mice results in an increase in body weight and decrease in energy expenditure thus establishing a leptin - resistant state. High fat intake rich in saturated fatty acids promotes colon cancer risk in azoxy methane (AOM) - induced rats.
On the other hand, TI - treated group showed an insignificant correlation between CCPR and thymidine labeling at four weeks, supporting the hypothesis that DNA synthesizing cells remain microscopically unaltered[50, 51]. A further continuation of DNA synthesis in those cells in which it normally ceased was the earliest and most reliable pathological sign of the commencement of neoplastic alterations[52], as we have observed in the OB and TIFB - treated groups, suggesting that obesity and fatty diet influenced the intensity of response to TI. It was likely that the colonocytes of TI- per se treated group stayed in phase I, according to Lipkin[53] wherein, cells remained microscopically normal and moved towards medial two thirds of the crypt, and in TIFB- and OB- treated groups, the cells stayed in phase II stage, wherein proliferating cells were not shed into the lumen and, due to inadequate suppression of the DNA synthesis, accumulated in the mucosa, leading to neoplastic transformation.
Observations of a highly significant reduction of SDH levels in OB and TIFB - treated animals could be attributed to the fact that the exposures to TI and FB of normal and obese animals might have caused damage to the mitochondrial membrane and impaired the aerobic metabolism and electron transport chain in the mitochondria[54]. Further, attenuation and dispersal of cell organelles were affected due to fusion of cells during proliferation. This may also lead to decreased enzyme activities[55].
Dipeptidyl amino peptidase IV is a brush border enzyme. It plays an important role perhaps, together with other amino peptidases, in the final digestion of nutritional constituents[56]. Dipeptidyl aminopeptidase liberates dipeptides from the amino terminal end of the polypeptides, derived from other digestive enzymes such as trypsin, chymotrypsin and pepsin. Transport of these amino acids as well as dipeptides is done by the brush border hydrolases, therefore known as carriers[57,58]. A decrease in such an enzyme activity as observed in this study might lead to an increase in the intracellular amino acids pool. In addition, obesity induction has been found to be associated with alterations in the cholesterol and membrane bound- lipid components[59]. We, therefore, suggested that altered membrane fluidity due in part to the effects of TI and FB, might have affected the glycoprotein constituents of the cell membrane resulting in decreased peptidase activity. A similar pattern of reduction in the activity of acid β-galactosidase has been observed in the entire crypt segment, the reduction of this enzyme was found to be more pronounced in the apical crypt epithelium, with respect to TIFB treatment and OB induction. Literature evidenced[60] that cellular membrane damage caused release of lysosomal enzymes into the blood stream, leading to an insult to the cellular integrity.
Finally, our design involved a multifactorial exposure to dietary components of either sex of animals. It excluded typical chemical inducers to cause genetic susceptibility. In addition, we employed a method of inducing obesity with MSG, so as to relate with the realistic regular human diets constituted with this typical flavoring agent, classified as a non-nutritional factor by World Health Organization[61]. Apart from the probable mechanisms so far discussed, there might be some other parameters, such as intestinal enzyme activities and gross metabolic alterations, detailed studies on these aspects have been carried out and results are yet to be communicated.
ACKNOWLEDGEMENTS
Thanks are due to Professor S. Govindasamy, our former Head, Department of Biochemistry and Molecular Biology, University of Madras, Guindy Campus, Chennai - 600025 India.
Footnotes
Supported by Senior Research fellowship from Council of Scientific and Industrial Research (CSIR), New Delhi, India, SRF No. [9/115 (462) / 98 EMR-1- BKR]
Edited by Wang XL and Zhu LH
References
- 1.Deng DJ. Progress of gastric cancer etiology: N-nitrosamides 1999s. World J Gastroenterol. 2000;6:613–618. doi: 10.3748/wjg.v6.i4.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.O'Brien H, Matthew JA, Gee JM, Watson M, Rhodes M, Speakman CT, Stebbings WS, Kennedy HJ, Johnson IT. K-ras mutations, rectal crypt cells proliferation, and meat consumption in patients with left-sided colorectal carcinoma. Eur J Cancer Prev. 2000;9:41–47. doi: 10.1097/00008469-200002000-00006. [DOI] [PubMed] [Google Scholar]
- 3.Giovannucci E. Obesity, gender, and colon cancer. Gut. 2002;51:147. doi: 10.1136/gut.51.2.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kaaks R, Lukanova A. Effects of weight control and physical activity in cancer prevention: role of endogenous hormone metabolism. Ann N Y Acad Sci. 2002;963:268–281. doi: 10.1111/j.1749-6632.2002.tb04118.x. [DOI] [PubMed] [Google Scholar]
- 5.Woods SC, Seeley RJ, Rushing PA, D'Alessio D, Tso P. A controlled high-fat diet induces an obese syndrome in rats. J Nutr. 2003;133:1081–1087. doi: 10.1093/jn/133.4.1081. [DOI] [PubMed] [Google Scholar]
- 6.Saglam K, Aydur E, Yilmaz M, Göktaş S. Leptin influences cellular differentiation and progression in prostate cancer. J Urol. 2003;169:1308–1311. doi: 10.1097/01.ju.0000055903.18400.25. [DOI] [PubMed] [Google Scholar]
- 7.Rock CL, Demark-Wahnefried W. Can lifestyle modification increase survival in women diagnosed with breast cancer? J Nutr. 2002;132:3504S–3507S. doi: 10.1093/jn/132.11.3504S. [DOI] [PubMed] [Google Scholar]
- 8.Reddy BS, Hedges AR, Laakso K, Wynder EL. Metabolic epidemiology of large bowel cancer: fecal bulk and constituents of high-risk North American and low-risk Finnish population. Cancer. 1978;42:2832–2838. doi: 10.1002/1097-0142(197812)42:6<2832::aid-cncr2820420644>3.0.co;2-l. [DOI] [PubMed] [Google Scholar]
- 9.Terry PD, Miller AB, Rohan TE. Obesity and colorectal cancer risk in women. Gut. 2002;51:191–194. doi: 10.1136/gut.51.2.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nelson DE, Bland S, Powell-Griner E, Klein R, Wells HE, Hogelin G, Marks JS. State trends in health risk factors and receipt of clinical preventive services among US adults during the 1990s. JAMA. 2002;287:2659–2667. doi: 10.1001/jama.287.20.2659. [DOI] [PubMed] [Google Scholar]
- 11.Deslypere JP. Obesity and cancer. Metabolism. 1995;44:24–27. doi: 10.1016/0026-0495(95)90316-x. [DOI] [PubMed] [Google Scholar]
- 12.Weber RV, Stein DE, Scholes J, Kral JG. Obesity potentiates AOM-induced colon cancer. Dig Dis Sci. 2000;45:890–895. doi: 10.1023/a:1005560621722. [DOI] [PubMed] [Google Scholar]
- 13.Alfieri M, Pomerleau J, Grace DM. A comparison of fat intake of normal weight, moderately obese and severely obese subjects. Obes Surg. 1997;7:9–15. doi: 10.1381/096089297765556150. [DOI] [PubMed] [Google Scholar]
- 14.Manjunath MN, Sattigeri VD, Rama Rao SN, Udah Rani M, Nagaraja KV. Physico-chemical composition of commercial tamarind powder. Indian Food Packer. 1991;45:39–42. [Google Scholar]
- 15.Shankaracharya NB. Tamarind–chemistry, technology and uses–A critical appraisal. J Food Sci Tech Mysore. 1988;35:193–203. [Google Scholar]
- 16.Mustapha A, Yakasai IA, Aguye IA. Effect of Tamarindus indica L. on the bioavailability of aspirin in healthy human volunteers. Eur J Drug Metab Pharmacokinet. 1996;21:223–226. doi: 10.1007/BF03189717. [DOI] [PubMed] [Google Scholar]
- 17.Kobayashi A, Adenan MI, Kajiyama S, Kanzaki H, Kawazu K. A cytotoxic principle of Tamarindus indica, di-n-butyl malate and the structure-activity relationship of its analogues. Z Naturforsch C. 1996;51:233–242. doi: 10.1515/znc-1996-3-415. [DOI] [PubMed] [Google Scholar]
- 18.Poulsen M, Mølck AM, Thorup I, Breinholt V, Meyer O. The influence of simple sugars and starch given during pre- or post-initiation on aberrant crypt foci in rat colon. Cancer Lett. 2001;167:135–143. doi: 10.1016/s0304-3835(01)00487-6. [DOI] [PubMed] [Google Scholar]
- 19.Le Leu RK, Brown IL, Hu Y, Young GP. Effect of resistant starch on genotoxin-induced apoptosis, colonic epithelium, and lumenal contents in rats. Carcinogenesis. 2003;24:1347–1352. doi: 10.1093/carcin/bgg098. [DOI] [PubMed] [Google Scholar]
- 20.Le Leu RK, Hu Y, Young GP. Effects of resistant starch and nonstarch polysaccharides on colonic luminal environment and genotoxin-induced apoptosis in the rat. Carcinogenesis. 2002;23:713–719. doi: 10.1093/carcin/23.5.713. [DOI] [PubMed] [Google Scholar]
- 21.Block G, Patterson B, Subar A. Fruit, vegetables, and cancer prevention: a review of the epidemiological evidence. Nutr Cancer. 1992;18:1–29. doi: 10.1080/01635589209514201. [DOI] [PubMed] [Google Scholar]
- 22.Patterson BH, Block G. Food choices and the cancer guidelines. Am J Public Health. 1988;78:282–286. doi: 10.2105/ajph.78.3.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bunyan J, Murrell EA, Shah PP. The induction of obesity in rodents by means of monosodium glutamate. Br J Nutr. 1976;35:25–39. doi: 10.1079/bjn19760005. [DOI] [PubMed] [Google Scholar]
- 24.Report of the American Institute of Nurtition ad hoc Committee on Standards for Nutritional Studies. J Nutr. 1977;107:1340–1348. doi: 10.1093/jn/107.7.1340. [DOI] [PubMed] [Google Scholar]
- 25.Deschner EE, Lipkin M. In: Gastrointestinal tract cancer. Ed: Lipkin M, Good RA. Sloane kettering Institute, Cancer Series, New York; 1978. p. 14. [Google Scholar]
- 26.DeRubertis FR, Craven PA, Saito R. Role of prostaglandins in bile salt-induced changes in rat colonic structure and function. J Clin Invest. 1984;74:1614–1624. doi: 10.1172/JCI111577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Burton K. A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem J. 1956;62:315–323. doi: 10.1042/bj0620315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kornblum HI, Loughlin SE, Leslie FM. Effects of morphine on DNA synthesis in neonatal rat brain. Brain Res. 1987;428:45–52. doi: 10.1016/0165-3806(87)90081-2. [DOI] [PubMed] [Google Scholar]
- 29.Pearse AGE. Histochemistry, Theoritical and applied. Third edition. Churchill Livingstone London. 1972;2:1342–1343. [Google Scholar]
- 30.Gossrau R. Splitting of naphthol AS-BI beta-galactopyranoside by acid beta-galactosidase. Histochemie. 1973;37:89–91. doi: 10.1007/BF00306863. [DOI] [PubMed] [Google Scholar]
- 31.McDonald JK, Schwabe C. In: Proteinases in mammalian cells and tissues, North Holland, Amsterdam 1977: 311 [Google Scholar]
- 32.Goeting N, Trotter GA, Cooke T, Kirkham N, Taylor I. Effect of warfarin on cell kinetics, epithelial morphology and tumour incidence in induced colorectal cancer in the rat. Gut. 1985;26:807–815. doi: 10.1136/gut.26.8.807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sandforth F, Heimpel S, Balzer T, Gutschmidt S, Riecken EO. Characterization of stereomicroscopically identified preneoplastic lesions during dimethylhydrazine-induced colonic carcinogenesis. Eur J Clin Invest. 1988;18:655–662. doi: 10.1111/j.1365-2362.1988.tb01283.x. [DOI] [PubMed] [Google Scholar]
- 34.Wright NA. Epithelial stem cell repertoire in the gut: clues to the origin of cell lineages, proliferative units and cancer. Int J Exp Pathol. 2000;81:117–143. doi: 10.1046/j.1365-2613.2000.00146.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sakata T. Stimulatory effect of short-chain fatty acids on epithelial cell proliferation in the rat intestine: a possible explanation for trophic effects of fermentable fibre, gut microbes and luminal trophic factors. Br J Nutr. 1987;58:95–103. doi: 10.1079/bjn19870073. [DOI] [PubMed] [Google Scholar]
- 36.Whitehead RH, Young GP, Bhathal PS. Effects of short chain fatty acids on a new human colon carcinoma cell line (LIM1215) Gut. 1986;27:1457–1463. doi: 10.1136/gut.27.12.1457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cummings JH. Colonic absorption: the importance of SCFAs in man. Scand J Gastroenterol Suppl. 1984;93:117–131. [PubMed] [Google Scholar]
- 38.Samelson SL, Nelson RL, Nyhus LM. Protective role of faecal pH in experimental colon carcinogenesis. J R Soc Med. 1985;78:230–233. doi: 10.1177/014107688507800311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Barthold SW. Relationship of colonic mucosal background to neoplastic proliferative activity in dimethylhydrazine-treated mice. Cancer Res. 1981;41:2616–2620. [PubMed] [Google Scholar]
- 40.Goodlad RA, Ratcliffe B, Fordham JP, Wright NA. Does dietary fibre stimulate intestinal epithelial cell proliferation in germ free rats? Gut. 1989;30:820–825. doi: 10.1136/gut.30.6.820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Folino M, McIntyre A, Young GP. Dietary fibers differ in their effects on large bowel epithelial proliferation and fecal fermentation-dependent events in rats. J Nutr. 1995;125:1521–1528. doi: 10.1093/jn/125.6.1521. [DOI] [PubMed] [Google Scholar]
- 42.McIntyre A, Gibson PR, Young GP. Butyrate production from dietary fibre and protection against large bowel cancer in a rat model. Gut. 1993;34:386–391. doi: 10.1136/gut.34.3.386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fleming SE, Fitch MD, De Vries S. The influence of dietary fiber on proliferation of intestinal mucosal cells in miniature swine may not be mediated primarily by fermentation. J Nutr. 1992;122:906–916. doi: 10.1093/jn/122.4.906. [DOI] [PubMed] [Google Scholar]
- 44.Caderni G, Stuart EW, Bruce WR. Dietary factors affecting the proliferation of epithelial cells in the mouse colon. Nutr Cancer. 1988;11:147–153. doi: 10.1080/01635588809513982. [DOI] [PubMed] [Google Scholar]
- 45.Lupton JR, Coder DM, Jacobs LR. Influence of luminal pH on rat large bowel epithelial cell cycle. Am J Physiol. 1985;249:G382–G388. doi: 10.1152/ajpgi.1985.249.3.G382. [DOI] [PubMed] [Google Scholar]
- 46.Freeman HJ, San RH. Use of unscheduled DNA synthesis in freshly isolated human intestinal mucosal cells for carcinogen detection. Cancer Res. 1980;40:3155–3157. [PubMed] [Google Scholar]
- 47.Reddy BS, Wang CX, Maruyama H. Effects of restricted caloric intake on azoxymethane-induced colon tumor incidence in wale f344 rats. Cancer Res. 1987;74:1226–1228. [PubMed] [Google Scholar]
- 48.Rasmussen RE, Boyd CH, Dansie DR, Kouri RE, Henry CJ. DNA replication and unscheduled DNA synthesis in lungs of mice exposed to cigarette smoke. Cancer Res. 1981;41:2583–2588. [PubMed] [Google Scholar]
- 49.Ogus S, Ke Y, Qiu J, Wang B, Chehab FF. Hyperleptinemia precipitates diet-induced obesity in transgenic mice overexpressing leptin. Endocrinology. 2003;144:2865–2869. doi: 10.1210/en.2002-0178. [DOI] [PubMed] [Google Scholar]
- 50.Deschner EE, Lipkin M. Study of human rectal epithelial cells in vitro. III. RNA, protein, and DNA synthesis in polyps and adjacent mucosa. J Natl Cancer Inst. 1970;44:175–185. [PubMed] [Google Scholar]
- 51.Lieb LM, Lisco H. In vitro uptake of tritiated thymidine by carcinoma of the human colon. Cancer Res. 1966;26:733–740. [PubMed] [Google Scholar]
- 52.Zhuang XQ, Yuan SZ, Wang XH, Lai RQ, Luo ZQ. Oncoprotein expression and inhibition of apoptosis during colorectal tumorigenesis. China Nati J New Gastroenterol. 1996;2:3–5. [Google Scholar]
- 53.Lipkin M. Proliferative changes in the colon. Cancer Suppl. 1974;34:878–888. [Google Scholar]
- 54.Borthwick GM, Johnson MA, Ince PG, Shaw PJ, Turnbull DM. Mitochondrial enzyme activity in amyotrophic lateral sclerosis: implications for the role of mitochondria in neuronal cell death. Ann Neurol. 1999;46:787–790. doi: 10.1002/1531-8249(199911)46:5<787::aid-ana17>3.0.co;2-8. [DOI] [PubMed] [Google Scholar]
- 55.Maskens AP, Dujardin-Loits RM. Experimental adenomas and carcinomas of the large intestine behave as distinct entities: most carcinomas arise de novo in flat mucosa. Cancer. 1981;47:81–89. doi: 10.1002/1097-0142(19810101)47:1<81::aid-cncr2820470115>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 56.Gutschmidt S, Gossrau R. A quantitative histochemical study of dipeptidylpeptidase IV (DPP IV) Histochemistry. 1981;73:285–304. doi: 10.1007/BF00493027. [DOI] [PubMed] [Google Scholar]
- 57.Ward PE, Sheridan MA, Hammon KJ, Erdös EG. Angiotensin I converting enzyme (kininase II) of the brush border of human and swine intestine. Biochem Pharmacol. 1980;29:1525–1529. doi: 10.1016/0006-2952(80)90603-6. [DOI] [PubMed] [Google Scholar]
- 58.Matthews DM, Adibi SA. Peptide Absorption. Gastroenterology. 1976;71:151–161. [PubMed] [Google Scholar]
- 59.Reynier M, Sari H, d'Anglebermes M, Kye EA, Pasero L. Differences in lipid characteristics of undifferentiated and enterocytic-differentiated HT29 human colonic cells. Cancer Res. 1991;51:1270–1277. [PubMed] [Google Scholar]
- 60.Kyriacou K, Garrett JR. Histochemistry of hydrolytic enzymes in resting submandibular glands of rabbits. Histochem J. 1985;17:683–698. doi: 10.1007/BF01003520. [DOI] [PubMed] [Google Scholar]
- 61.Robert HR. In: Food Safety. Ed: Roberts Hr, New York Wiley; 1999. p. 239. [Google Scholar]