

Abstract
AIM: To investigate the loss of heterozygosity (LOH) and mutation of tumor suppressor gene PTEN in gastric cancer and precancerous lesions.
METHODS: Thirty cases of normal gastric mucosa, advanced and early stage gastric cancer, intestinal metaplasia, atrophic gastritis, and atypical hyperplasia were analyzed for PTEN LOH and mutations within the entire coding region of PTEN gene by PCR-SSCP denaturing PAGE gel electrophoresis, and PTEN mutation was detected by PCR-SSCP sequencing followed by silver staining.
RESULTS: LOH rate found in respectively atrophic gastritis was 10% (3/30), intestinal metaplasia 10% (3/30), atypical hyperplasia 13.3% (4/30), early stage gastric cancer 20% (6/30), and advanced stage gastric cancer 33.3% (9/30), None of the precancerous lesions and early stage gastric cancer showed PTEN mutations, but 10% (3/30) of the advanced stage gastric cancers, which were all positive for LOH, showed PTEN mutation.
CONCLUSION: LOH of PTEN gene appears in precancerous lesions, and PTEN mutations are restricted to advanced gastric cancer, LOH and mutation of PTEN gene are closely related to the infiltration and metastasis of gastric cancer.
Keywords: Gastric cancer, Precancerous lesions, PTEN gene, Loss of heterozygosity, Mutation
INTRODUCTION
A candidate tumor suppressor gene PTEN (also known as MMAC1 or TEP1) has recently been isolated from chromosome 10q23.3[1-3]. They are found mutated in several cancer types that display LOH in this region[4,5]. The PTEN gene encodes a 403 amino acid protein homologous to some protein phosphatases, and the protein has been shown to possess protein phosphatase activity in vitro[6-8]. It is thought that PTEN protein dephosphorylates the 3 positions of phosphatidylinositol 3,4,5-triphosphate (PIP3), a well-known intracellular messenger of certain cell-growth stimulators[9,10]. The molecular mechanisms of PTEN have been elucidated recently, and it is considered that PTEN belongs to a class of tumor suppressor genes together with p53, Rb, and APC. Gastric cancer is the most common digestive tract cancer diagnosed in China. In spite of its impact on human health, the molecular mechanisms involved in the pathogenesis of gastric cancer remain relatively unknown. PTEN is expressed in normal gastric mucosa, implying that loss of function may have some consequences in gastric cancer. We examined patients of advanced stage gastric cancer, especially the precancerous lesions for LOH at the loci, and observed the PTEN gene and its mutation from different visual aspects. We found the frequent presence of LOHs at 10q23.3 in gastric cancer and the precancerous lesions and mutations of PTEN gene in patients with advanced stage gastric cancer, suggesting that the inactivation of PTEN gene might be more closely related to the infiltration and metastasis of gastric cancer than previous observations.
MATERIALS AND METHODS
Tissue samples
Thirty cases of normal gastric mucosa, atrophic gastritis, intestinal metaplasia, atypical hyperplasia, and the early and advanced stage gastric cancer were obtained from the Endoscopic Center and operation rooms of China Medical University from 2000 to 2002. The tissues were extracted 5 cm away from the lesion for the control group. All the specimens were embedded in OCT rather then snap-frozen in the liquid nitrogen, and kept frozen at -70 °C. The cryostat sections from each tumor specimen were examined histologically, and only those blocks of tumor tissues composed of more than 70% neoplastic cells were selected for subsequent DNA isolation. DNA was from gastric cancers, precancerous lesions, and the corresponding normal tissues following the saturated sodium chloride method.
LOH analysis
Three microsatellite markers (D10S215, D10S541 and D10S 2491) were used to evaluate LOH on 10q23.3[11]. All the primers used in this study were obtained from AoKe Corporation, Beijing. The sequences of the primers are shown in Table 1. Each of the PCR mixtures contained 20 ng of genomic DNA, 2 μL of 10×PCR buffer, 0.4 μmol/L of each primer, 1.5 mmol/L Mg2+, 200 mmol/L of dNTPs, and 1 unit of Taq DNA polymerase (Takara, Dalian). PCR was carried out over 35 amplification cycles for 45 s at 94 °C, 45 s at 55 °C, 60 s at 72 °C. The PCR mixture was subjected to predegeneration for 5 min at 95 °C and a further extension for 10 min at 72 °C. After the amplification, PCR products were resolved on 160 g/L denaturing polyacrylamide gel and sliver staining was performed for analysis.
Table 1.
Primers of PTEN microsatellite loci and exons.
| Loci | Position | Primer sequence |
| D10S215 | 10q22-23 | P1 TGGCATCATTCTGGGGA |
| P2 TTACGTTTCTTCACATGGT | ||
| D10S541 | 10q22-23 | P1 AAGCAAGTGAAGTCTTAGAACCACC |
| P2 CCACAAGTAACAGAAAGCCTGTCTC | ||
| D10S2491 | 10q23-23 | P1 TTATAAGGACTGAGTGAGGGA |
| P2 GTTAGATAGAGTACCTGCACTC | ||
| Exon 5 | P1 CTTATTCTGAGGTTATCTTTTTTACC | |
| P2 CTCAGAATCCAGGAAGAGGA | ||
| Exon 8 | P1 ACACATCACATACATACAAGTC | |
| P2 GTGCAGATAATGACAAGGAATA |
PTEN mutations
Exons 5 and 8 of PTEN were amplified separately using the primer sets described by Risinger et al[12]. The primer sequences of exons 5 and 8 are shown in Table 1. Genomic DNA was subjected to PCR amplification in 20 μL of the reaction mixture, which contained 20 ng of genomic DNA, 2.5 mol/L of each primer, 2 μL of 10×PCR buffer, 2.0 mmol/L Mg2+, 200 mmol/L dNTP, and 1 unit of TaqDNA polymerase (Takara, Dalian). PCR was carried out over 35 amplification cycles for 45 s at 94 °C, 30 s at 58 °C, 60 s at 72 °C. The PCR mixture was subjected to predenaturation at 95 °C for 5 min and a final extension at 72 °C for 10 min. After completion of PCR, the ratio of 1:5 volume of sequencing-stop-solution was added, heated at 95 °C and rapidly cooled on ice, and 1-5 μL of resulting mixtures was loaded on 120 g/L non-denaturing polyacrylamide gel and silver stained, then the shifted bands were sequenced.
DNA sequencing
After the electrophoresis, shifted SSCP bands were excised from the gel. We extracted the DNA from the gel with distilled water, then reamplified it using the original PCR primers. The reamplified PCR products were sequenced with an ABI PRISM310 dye terminator cycle sequencing ready reaction kit.
RESULTS
LOH analysis
We examined the genotypes of gastric cancer and precancerous lesions at three highly polymorphic loci distributed at 10q23.3 (D10S215, D10S541 and D10S2491). In our study, 10% of atrophic gastritis and intestinal metaplasia and 13.3% of atypical hyperplasia demonstrated LOH in the 10q23.3 region (Figure 1 and Table 1). To examine whether the LOH found frequently in this region was also present in gastric cancer, we further studied 30 early stage and 30 advanced stage cancers (Table 2 and Figure 1). We found that 20% of the early stage gastric cancers and 30% of the advanced stage gastric cancers demonstrated LOH at 10q23.3, while none of the normal gastric mucosa showed LOH at D10S215, D10S54, or D10S 2491.
Figure 1.
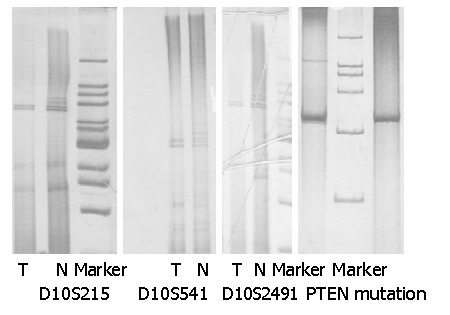
Representative silver staining showing allelic losses at markers near or within PTEN and mutation of PTEN (T: tumor; N: normal;).
Table 2.
PTEN LOH in gastric cancer and precancerous lesion.
| Lesions | D10S215 % (n) | D10S541 % (n) | D10S2491 % (n) | Total % |
| AG | 3.3 (1/30) | 3.3 (1/30) | 6.7 (2/30) | 10 |
| IM | 3.3 (1/30) | 3.3 (1/30) | 3.3 (1/30) | 10 |
| AH | 6.7 (2/30) | 3.3 (1/30) | 6.7 (2/30) | 13.3 |
| Gastric cancer | ||||
| Early stage | 6.7 (2/30) | 10 (3/30) | 6.7 (2/30) | 20 |
| Advanced stage | 13.3 (4/30) | 6.7 (2/30) | 13.3 (4/30) | 30 |
AG: atrophic gastritis; IM: intestinal metaplasia; AH: atypical hyperplasia.
SSCP and sequencing
SSCP analysis of the PTEN gene was performed on gastric cancer and precancerous lesions. We screened the exons 5 and 8 of PTEN coding region, and no mutation was detected in early stage gastric cancer or precancerous lesions. However, one mutation in exon 5, and two mutations in exon 8 of PTEN coding region were detected in advanced gastric cancer (Table 3 and Figure 1)
Table 3.
Mutation of PTEN in gastric cancer.
| Case | Exon | LOH | Coden | Base change | Stage | Pathology |
| 11 | 5 | + | 91 | GAA to CAA | IIIa | Signet-ring cell cancer |
| 19 | 8 | + | 335 | CGA to TGA | II | Hypodifferentiated adenocarcinoma |
| 22 | 8 | + | 329 | Del 4 bp | IIIb | Undifferentiate gastric cancer |
DISCUSSION
PTEN/MMAC1 was originally isolated from a region homozygously deleted in several cancer cell lines, including glioma and cancers of the breast and prostate. Mutations in this gene have been reported as glioma, endometrial carcinoma, breast tumors, and malignant melanoma[13-16]. Germ-line mutation of PTEN/MMAC1 is also associated with two autosomal dominant disorders belonging to the family of hamartomatous polyposis syndrome[17,18]. Thus some cancers seem devoid of PTEN/MMAC1 alterations (e.g., serous carcinoma of endometrium and cervical cancer)[19]. Genetic changes of PTEN/MMAC1 occur in multiple types of cancer, suggesting that inactivation of PTEN/MMAC1 may play an important but perhaps somewhat general role in the pathogenesis of a variety of human malignancies.
There is strong circumstantial evidence that allelic loss of PTEN is found in a large variety of human cancers. The inactivation of PTEN gene may be due to mutation and LOH or decreasing expression of PTEN mRNA or protein[20-22]. The earliest research done on PTEN gene was for experimentation of glioma. The result showed that LOH rate was 70-80%, while the mutation rate was 27-30% for the PTEN gene in glioma[23]. In prostate cancer, PTEN LOH was reported to be 32-63% while PTEN mutation rate was 12-25%[24,25]. In endometrial carcinoma, PTEN mutation rate was 19%[26], and ovary cancer showed 27.3-42.1% of PTEN LOH with a mutation rate of 8.3-21%[27]. In the benign lesion of ovary cyst, the PTEN LOH rate was 56.3%, and PTEN mutation rate was 20.6%[28]. It has been speculated that PTEN mutation and its relation to gastrointestinal tumor often occur in the colorectal tumor with hereditary tendency such as familial polyposis[29]. The mutation rate is low in sporadic colorectal cancers[30]. Taniyama et al[31] studied 32 cases of sporadic colon cancer, but observed a bi-allelic genomic alteration that caused loss of function of the gene only in one case. Wang et al[32] found only one somatic mutation of PTEN gene out of 72 colon cancer cases. Regarding the relationship between PTEN mutation and gastric cancer, Zheng et al[33] found PTEN was closely related to the infiltration and metastasis of gastric cancer. Our previous study showed PTEN inactivation was also related to the precancerous lesion of gastric cancer. In this study, the rates of PTEN LOH in atrophic gastritis, intestinal metaplasia, and atypical hyperplasia were 10, 10, and 13.3%, respectively. In early stage gastric cancer, PTEN LOH rate was 20%, while it was 30% in the advanced gastric cancer. There were no PTEN mutations in precancerous lesions or in any early stage gastric cancer. The PTEN mutation rate was observed in 10% of advanced gastric cancers, showing that the loss or inactivation of PTEN gene is related to the tumor genesis of gastric cancer, especially to the invasion and metastasis of gastric cancer. For the three cases of PTEN showing mutations, one tumor located in the anus prolonged and spread to the lower part of esophagus, and the pathology showed signet cancer. One case showing the tumor located in the corpus without penetrating the seroma, with the lymph node metastasis around the artery, pathology showed Undifferentiated cancer. Another case showing the tumor located in the sinus was able to infiltrate to muscular layer, with the metastasis of lymph node within 5 cm of the tumor margin. The pathology for this particular case was regarded as poorly-differentiated adenocarcinoma.
Li et al has previously studied the expression of PTEN encoding products in gastric cancer and precancerous lesions by immunohistochemistry. The result showed PTEN was down-regulated in the tumorigenesis of gastric cancer.
Based on the findings from this study, we conclude that PTEN LOH and the mutation rates are lower in gastric cancer compared with that in the other tumors (10-33.3% for PTEN LOH and 0-10% for the mutation rate). This further supports that the pathogenesis of gastric cancer is a complicated molecular mechanism closely associated with genes, such as oncogene, tumor suppressor gene, mismatch repair gene, telomere and telomerase, cellular adhesive factors, etc.
Footnotes
Supported by the National Natural Science Foundation of China, No. 30070845
Edited by Kumar M and Ma JY
References
- 1.Kim S, Domon-Dell C, Wang Q, Chung DH, Di Cristofano A, Pandolfi PP, Freund JN, Evers BM. PTEN and TNF-alpha regulation of the intestinal-specific Cdx-2 homeobox gene through a PI3K, PKB/Akt, and NF-kappaB-dependent pathway. Gastroenterology. 2002;123:1163–1178. doi: 10.1053/gast.2002.36043. [DOI] [PubMed] [Google Scholar]
- 2.McConnachie G, Pass I, Walker SM, Downes CP. Interfacial kinetic analysis of the tumour suppressor phosphatase, PTEN: evidence for activation by anionic phospholipids. Biochem J. 2003;371:947–955. doi: 10.1042/BJ20021848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Li J, Yen C, Liaw D, Podsypanina K, Bose S, Wang SI, Puc J, Miliaresis C, Rodgers L, McCombie R, et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science. 1997;275:1943–1947. doi: 10.1126/science.275.5308.1943. [DOI] [PubMed] [Google Scholar]
- 4.Zhao H, Dupont J, Yakar S, Karas M, LeRoith D. PTEN inhibits cell proliferation and induces apoptosis by downregulating cell surface IGF-IR expression in prostate cancer cells. Oncogene. 2004;23:786–794. doi: 10.1038/sj.onc.1207162. [DOI] [PubMed] [Google Scholar]
- 5.Konopka B, Janiec-Jankowska A, Paszko Z, Goluda M. The coexistence of ERBB2, INT2, and CMYC oncogene amplifications and PTEN gene mutations in endometrial carcinoma. J Cancer Res Clin Oncol. 2004;130:114–121. doi: 10.1007/s00432-003-0518-7. [DOI] [PubMed] [Google Scholar]
- 6.Brown KS, Blair D, Reid SD, Nicholson EK, Harnett MM. FcgammaRIIb-mediated negative regulation of BCR signalling is associated with the recruitment of the MAPkinase-phosphatase, Pac-1, and the 3'-inositol phosphatase, PTEN. Cell Signal. 2004;16:71–80. doi: 10.1016/s0898-6568(03)00113-x. [DOI] [PubMed] [Google Scholar]
- 7.Cho SH, Lee CH, Ahn Y, Kim H, Kim H, Ahn CY, Yang KS, Lee SR. Redox regulation of PTEN and protein tyrosine phosphatases in H(2)O(2) mediated cell signaling. FEBS Lett. 2004;560:7–13. doi: 10.1016/s0014-5793(04)00112-7. [DOI] [PubMed] [Google Scholar]
- 8.Wishart MJ, Dixon JE. PTEN and myotubularin phosphatases: from 3-phosphoinositide dephosphorylation to disease. Trends Cell Biol. 2002;12:579–585. doi: 10.1016/s0962-8924(02)02412-1. [DOI] [PubMed] [Google Scholar]
- 9.Mills GB, Kohn E, Lu Y, Eder A, Fang X, Wang H, Bast RC, Gray J, Jaffe R, Hortobagyi G. Linking molecular diagnostics to molecular therapeutics: targeting the PI3K pathway in breast cancer. Semin Oncol. 2003;30:93–104. doi: 10.1053/j.seminoncol.2003.08.011. [DOI] [PubMed] [Google Scholar]
- 10.Orchiston EA, Bennett D, Leslie NR, Clarke RG, Winward L, Downes CP, Safrany ST. PTEN M-CBR3, a versatile and selective regulator of inositol 1,3,4,5,6-pentakisphosphate (Ins(1,3,4,5,6)P5). Evidence for Ins(1,3,4,5,6)P5 as a proliferative signal. J Biol Chem. 2004;279:1116–1122. doi: 10.1074/jbc.M310933200. [DOI] [PubMed] [Google Scholar]
- 11.Whang YE, Wu X, Suzuki H, Reiter RE, Tran C, Vessella RL, Said JW, Isaacs WB, Sawyers CL. Inactivation of the tumor suppressor PTEN/MMAC1 in advanced human prostate cancer through loss of expression. Proc Natl Acad Sci USA. 1998;95:5246–5250. doi: 10.1073/pnas.95.9.5246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Risinger JI, Hayes AK, Berchuck A, Barrett JC. PTEN/MMAC1 mutations in endometrial cancers. Cancer Res. 1997;57:4736–4738. [PubMed] [Google Scholar]
- 13.Ding X, Endo S, Zhang SJ, Saito T, Kouno M, Kuroiwa T, Washiyama K, Kumanishi T. Primary malignant lymphoma of the brain: analysis of MMAC1 (PTEN) tumor suppressor gene. Brain Tumor Pathol. 2001;18:139–143. doi: 10.1007/BF02479427. [DOI] [PubMed] [Google Scholar]
- 14.Kanamori Y, Kigawa J, Itamochi H, Sultana H, Suzuki M, Ohwada M, Kamura T, Sugiyama T, Kikuchi Y, Kita T, et al. PTEN expression is associated with prognosis for patients with advanced endometrial carcinoma undergoing postoperative chemotherapy. Int J Cancer. 2002;100:686–689. doi: 10.1002/ijc.10542. [DOI] [PubMed] [Google Scholar]
- 15.Chung MJ, Jung SH, Lee BJ, Kang MJ, Lee DG. Inactivation of the PTEN gene protein product is associated with the invasiveness and metastasis, but not angiogenesis, of breast cancer. Pathol Int. 2004;54:10–15. doi: 10.1111/j.1440-1827.2004.01576.x. [DOI] [PubMed] [Google Scholar]
- 16.Tsao H, Goel V, Wu H, Yang G, Haluska FG. Genetic interaction between NRAS and BRAF mutations and PTEN/MMAC1 inactivation in melanoma. J Invest Dermatol. 2004;122:337–341. doi: 10.1046/j.0022-202X.2004.22243.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vega A, Torres J, Torres M, Cameselle-Teijeiro J, Macia M, Carracedo A, Pulido R. A novel loss-of-function mutation (N48K) in the PTEN gene in a Spanish patient with Cowden disease. J Invest Dermatol. 2003;121:1356–1359. doi: 10.1111/j.1523-1747.2003.12638.x. [DOI] [PubMed] [Google Scholar]
- 18.Huang SC, Chen CR, Lavine JE, Taylor SF, Newbury RO, Pham TT, Ricciardiello L, Carethers JM. Genetic heterogeneity in familial juvenile polyposis. Cancer Res. 2000;60:6882–6885. [PubMed] [Google Scholar]
- 19.Martini M, Ciccarone M, Garganese G, Maggiore C, Evangelista A, Rahimi S, Zannoni G, Vittori G, Larocca LM. Possible involvement of hMLH1, p16(INK4a) and PTEN in the malignant transformation of endometriosis. Int J Cancer. 2002;102:398–406. doi: 10.1002/ijc.10715. [DOI] [PubMed] [Google Scholar]
- 20.Ning K, Pei L, Liao M, Liu B, Zhang Y, Jiang W, Mielke JG, Li L, Chen Y, El-Hayek YH, et al. Dual neuroprotective signaling mediated by downregulating two distinct phosphatase activities of PTEN. J Neurosci. 2004;24:4052–4060. doi: 10.1523/JNEUROSCI.5449-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hlobilková A, Knillová J, Bártek J, Lukás J, Kolár Z. The mechanism of action of the tumour suppressor gene PTEN. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2003;147:19–25. [PubMed] [Google Scholar]
- 22.Mahimainathan L, Choudhury GG. Inactivation of platelet-derived growth factor receptor by the tumor suppressor PTEN provides a novel mechanism of action of the phosphatase. J Biol Chem. 2004;279:15258–15268. doi: 10.1074/jbc.M314328200. [DOI] [PubMed] [Google Scholar]
- 23.Sano T, Lin H, Chen X, Langford LA, Koul D, Bondy ML, Hess KR, Myers JN, Hong YK, Yung WK, et al. Differential expression of MMAC/PTEN in glioblastoma multiforme: relationship to localization and prognosis. Cancer Res. 1999;59:1820–1824. [PubMed] [Google Scholar]
- 24.Whang YE, Wu X, Suzuki H, Reiter RE, Tran C, Vessella RL, Said JW, Isaacs WB, Sawyers CL. Inactivation of the tumor suppressor PTEN/MMAC1 in advanced human prostate cancer through loss of expression. Proc Natl Acad Sci USA. 1998;95:5246–5250. doi: 10.1073/pnas.95.9.5246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhao H, Dupont J, Yakar S, Karas M, LeRoith D. PTEN inhibits cell proliferation and induces apoptosis by downregulating cell surface IGF-IR expression in prostate cancer cells. Oncogene. 2004;23:786–794. doi: 10.1038/sj.onc.1207162. [DOI] [PubMed] [Google Scholar]
- 26.Salvesen HB, Stefansson I, Kalvenes MB, Das S, Akslen LA. Loss of PTEN expression is associated with metastatic disease in patients with endometrial carcinoma. Cancer. 2002;94:2185–2191. doi: 10.1002/cncr.10434. [DOI] [PubMed] [Google Scholar]
- 27.Obata K, Hoshiai H. Common genetic changes between endometriosis and ovarian cancer. Gynecol Obstet Invest. 2000;50 Suppl 1:39–43. doi: 10.1159/000052877. [DOI] [PubMed] [Google Scholar]
- 28.Sato N, Tsunoda H, Nishida M, Morishita Y, Takimoto Y, Kubo T, Noguchi M. Loss of heterozygosity on 10q23.3 and mutation of the tumor suppressor gene PTEN in benign endometrial cyst of the ovary: possible sequence progression from benign endometrial cyst to endometrioid carcinoma and clear cell carcinoma of the ovary. Cancer Res. 2000;60:7052–7056. [PubMed] [Google Scholar]
- 29.Negoro K, Takahashi S, Kinouchi Y, Takagi S, Hiwatashi N, Ichinohasama R, Shimosegawa T, Toyota T. Analysis of the PTEN gene mutation in polyposis syndromes and sporadic gastrointestinal tumors in Japanese patients. Dis Colon Rectum. 2000;43:S29–S33. doi: 10.1007/BF02237223. [DOI] [PubMed] [Google Scholar]
- 30.Guanti G, Resta N, Simone C, Cariola F, Demma I, Fiorente P, Gentile M. Involvement of PTEN mutations in the genetic pathways of colorectal cancerogenesis. Hum Mol Genet. 2000;9:283–287. doi: 10.1093/hmg/9.2.283. [DOI] [PubMed] [Google Scholar]
- 31.Taniyama K, Goodison S, Ito R, Bookstein R, Miyoshi N, Tahara E, Tarin D, Urquidi V. PTEN expression is maintained in sporadic colorectal tumours. J Pathol. 2001;194:341–348. doi: 10.1002/path.908. [DOI] [PubMed] [Google Scholar]
- 32.Wang Q, Wang X, Hernandez A, Hellmich MR, Gatalica Z, Evers BM. Regulation of TRAIL expression by the phosphatidylinositol 3-kinase/Akt/GSK-3 pathway in human colon cancer cells. J Biol Chem. 2002;277:36602–36610. doi: 10.1074/jbc.M206306200. [DOI] [PubMed] [Google Scholar]
- 33.Zheng HC, Chen Y, Kuang LG, Yang L, Li JY, Wu DY, Zhang SM, Xin Y. Expression of PTEN-encoding product in different stages of carcinogenesis and progression of gastric carcinoma. Zhonghua ZhongLiu ZaZhi. 2003;25:13–16. [PubMed] [Google Scholar]