

Abstract
Whether stimulant drugs like amphetamine increase or decrease choice of larger delayed reinforcers over smaller immediately available reinforcers under delay discounting procedures can depend on several factors, including the order in which delay is presented. This study examined whether the order of delay presentation impacts drug effects on discounting in rats (n=8) trained and tested under an ascending order, a descending order, as well as under a fixed delay condition. Responses on one lever delivered 1 food pellet immediately and responses on the other lever delivered 3 food pellets immediately or after a delay (4–32 s). In Experiment 1, the delay to the larger reinforcer varied within session and the order of delay presentation (ascending or descending) varied across conditions. In Experiment 2, the same delay value was presented in all blocks of the session (i.e., delay was fixed), and delay varied across phases. Under the ascending order of delay, amphetamine (0.32–1.78 mg/kg) increased choice of the larger reinforcer in some rats and decreased choice in others. In the same rats responding under the descending and fixed delay conditions, amphetamine markedly decreased choice of the larger reinforcer even in the component associated with no delay. In some subjects, the effects of amphetamine differed depending on the manner in which delay was presented, indicating that drug-induced changes in performance were due, in part, to mechanisms other than altered sensitivity to reinforcer delay. These results also suggest that a history of responding under both orders of delay presentation can modulate drug effects.
Keywords: amphetamine, delay discounting, order of delay presentation, lever press, rat
1. Introduction
Delay discounting is a process whereby the effectiveness of a consequence decreases as a function of the delay to its presentation (Mazur 1987). Delay discounting is thought to be an important behavioral process because of its apparent relevance to many socially important behavioral problems, particularly behavior that reflects greater impulsivity or a lack of self-control (Ainslie 1974; Rachlin and Green 1972; Logue 1988; Evenden 1999). For example, current drug abusers discount the value of delayed reinforcers more rapidly than former users or individuals that have never used drugs [see Bickel et al. (2012; 2013)]; enhanced discounting might predispose an individual to choose the more immediately available effects of drug taking rather than the delayed benefits of remaining abstinent such as health, income, and positive social interactions. Understanding processes that underlie such choices and knowledge of how certain experiences (e.g., drug use) further impact delay discounting will possibly aid in the development of more effective prevention and treatment strategies.
Many procedures have been developed to study how physiological, pharmacological, and behavioral factors impact delay discounting [for example, see Madden and Bickel (2010)], such as the procedure developed by Evenden and Ryan (1996) in which subjects choose between a small reinforcer (e.g., 1 food pellet) delivered immediately and a larger reinforcer (e.g., 3 food pellets) delivered immediately or following a delay. Delay to delivery of the larger reinforcer is varied systematically across blocks within the session with the most common variation of the procedure being one in which delay progressively increases across blocks (i.e., ascending delays). Delay functions obtained in this manner typically reflect a shift in preference from responding predominantly for the larger reinforcer early in the session, when the larger reinforcer is delivered immediately, to responding predominantly for the smaller reinforcer later in the session, when delivery of the larger reinforcer is delayed. The ability to rapidly assess delay discounting within a single session for individual subjects after relatively few (< 30) training sessions (e.g., Evenden and Ryan 1996) is suitable for behavioral pharmacology because it allows for determination of discounting at specific time points (e.g., acute drug effects) as well as evaluation of changes in discounting across time (e.g., during chronic drug administration or after discontinuation of drug administration) [see reviews by Perry and Carroll (2008), de Wit and Mitchell (2010), and Bari and Robbins (2013)].
The benefits of changing environmental variables such as delay within-session can be accompanied by potentially important issues (e.g., order effects) that can be addressed empirically by employing different procedural variations (Sidman 1960). For example, the effects of stimulant drugs such as amphetamine on delay discounting can differ qualitatively, either increasing or decreasing discounting, depending upon whether the delay period is paired with a unique stimulus (e.g., Cardinal et al. 2000). A recent study (Tanno et al. 2014) showed that the effects of amphetamine and methylphenidate on performance under a delay discounting procedure vary depending on the order in which delays are presented within the session. Both drugs increased choice of the larger delayed reinforcer in rats responding under an ascending order of delay, consistent with effects reported by others (e.g., Barbelivien et al. 2008; Cardinal et al. 2000; Huskinson et al. 2012; Pitts and McKinney 2005; Slezak and Anderson 2011; Slezak et al. 2013; Van Gaalen et al. 2006; Winstanley et al. 2003; 2005), but markedly decreased choice of the larger reinforcer in a separate group of rats responding under a descending order. A similar study (Slezak and Anderson 2009) examined the effects of amphetamine in rats trained and tested under both an ascending and a descending order of delay. Amphetamine decreased choice of the larger reinforcer under both orders of delay; however, the effects of amphetamine were more pronounced under the descending order, possibly reflecting an interaction with delay order within the same subject.
Taken together, the results of these studies raise the possibility that changes in performance under an ascending delay procedure are influenced by factors other than, or in addition to, changes in delay discounting. For example, stimulant drugs might increase perseveration, alter the estimation of the passage of time, or change sensitivity to reinforcer amount [see discussions by Pitts and Febbo (2004), Pitts and McKinney (2005), Richards et al. (1997), and Slezak and Anderson (2009)]. Because Tanno et al. (2014) assessed the interaction between drug effects and delay order using a between-groups design, differences might be due to factors other than, or in addition to, delay order; it might be the case that a history of responding exclusively with one order of delay enhances the apparent perseverative effects of amphetamine. One goal of the current study (Experiment 1) was to examine whether the delay-order effect reported previously could be demonstrated for an individual subject; therefore, the effects of amphetamine were assessed in rats that were trained and tested under both ascending and descending orders of delay presentation.
Studies using either a between-groups (Tanno et al. 2014) or within-subject (Slezak and Anderson 2009) design failed to show that amphetamine increases choice of larger delayed reinforcers (e.g., reduces delay discounting) when delay is presented in a descending order within session. If drug effects are mediated through changes in sensitivity to reinforcer delay, then such changes should be evident under various other conditions in which delay impacts behavior. Some data support the notion that amphetamine reduces sensitivity to reinforcer delay (e.g., Ta et al. 2008); however, amphetamine also impacts other behavioral processes thought to be relevant to delay discounting such as sensitivity to reinforcer amount (Maguire et al. 2009). A second goal of the current study (Experiment 2) was to determine whether amphetamine increases choice of larger, delayed reinforcers under conditions in which the impact of the order of delay presentation is reduced. Thus, the delay to the larger reinforcer was held constant within and across sessions, and delay was varied systematically across conditions.
2. Materials and methods
2.1. Subjects
Eight experimentally naïve, adult male Sprague-Dawley rats (Harlan Sprague-Dawley, Inc., Indianapolis, IN), approximately 3 months old at the beginning of the experiment, were housed individually in 45 × 24 × 20 cm high plastic cages containing rodent bedding (Sani-chips, Harlan Teklad, Madison, WI) in a colony room maintained on a 14:10 light/dark cycle with lights on at 0630 hr; experiments were conducted during the light period. Rats were fed chow (Rat Sterilizable Diet, Harlan Teklad) post-session to maintain their body weights at approximately 360 g. Water was available continuously in the home cage.
2.2. Apparatus
Sessions were conducted in sound-attenuating, ventilated enclosures (ENV-022M; Med Associates, Inc., St Albans, VT), which contained an operant conditioning chamber (ENV-008CT; Med Associates, Inc.) with an interior space measuring 31 × 24 × 21 cm high. The front door and rear panel were clear polycarbonate and both ends were aluminum panels. The right panel was equipped with two response levers horizontally aligned 11.5 cm apart, above each of which was a 2.5-cm diameter translucent circle that could be trans-illuminated white with a 100 mA light (lever lights). A 5 × 5 cm opening was centrally located between the two levers through which 45-mg food pellets (PJAI-0045; Noyes Precision Pellets, Research Diets Inc., New Brunswick, NJ) were delivered from a food hopper. The panel on the opposite side of the chamber was equipped with a 100 mA houselight centrally located near the top of the chamber. Data were collected using MED-PC IV software and a PC-compatible interface (Med Associates, Inc.).
2.3. Behavioral procedures
2.3.1. Initial training
Sessions began with illumination of the houselight and both lever lights; a response on either lever extinguished lever lights and delivered one food pellet immediately followed by re-illumination of the lever lights, signaling the next opportunity to respond. After 3 consecutive sessions in which 50 food pellets were delivered within 30 min, the number of food pellets available on the non-preferred lever, defined as the lever on which less than 50% of responses occurred for 3 consecutive sessions, increased to 3. After the first session in which at least 80% of responses occurred on the lever that delivered 3 pellets, the contingencies were reversed until at least 80% of responses occurred on the opposite lever. After at least 2 such alternations, the experimental procedure was introduced. The lever that delivered 3 food pellets was counterbalanced across rats and was maintained for an individual rat for the entire study.
2.3.2. Delay-discounting procedure
The behavioral procedure used in the current study was based on the procedure developed by Evenden and Ryan (1996) and recently described by Tanno et al. (2014). Daily sessions were divided into 5 blocks, each of which comprised 2 forced trials followed by 5 choice trials. The houselight was illuminated at the beginning of the block and remained illuminated for the duration of the block. Blocks were separated by a 30-s blackout period, during which all lights were extinguished. The beginning of a trial was signaled by illumination of one (forced trials) or both (choice trials) lever lights. A response on an active lever (i.e., located directly below an illuminated lever light) delivered either 1 food pellet immediately or 3 food pellets either immediately or after a delay of 4, 8, 16, or 32 s. When food was delivered immediately, lever lights were extinguished immediately upon the response. When food was delivered after a delay, lever lights were extinguished immediately upon the response and the light located above the lever associated with delayed food flashed at a rate of 1 Hz for the duration of the delay. If no response occurred within 20 s of the beginning of a trial (limited hold), lever lights were extinguished and the trial was recorded as an omission. Food delivery or an omission initiated an inter-trial blackout period during which the lever lights were extinguished. The duration of the inter-trial blackout period was adjusted for each trial such that trials started every 60 s.
2.4. Experiment 1: Ascending versus descending order of delay presentation
The first experiment assessed the effects of amphetamine in rats trained and tested under both an ascending and a descending order of delay presentation. Initially, rats chose between 1 and 3 food pellets delivered immediately for all 5 blocks of the session (no-delay sessions). After rats responded reliably on the lever that delivered the larger reinforcer (i.e., at least 80% choice of the larger reinforcer in each of 5 blocks) for 1 session, they were randomly assigned to either an ascending or a descending order of delay presentation with the constraint that 4 rats began under each order. Under the ascending order, both reinforcers were delivered immediately during the first block of the session and the delay to delivery of the larger reinforcer increased progressively across successive blocks. Under the descending order, the longest delay (32 s) was presented in the first block and delay decreased across successive blocks until the final block, during which both reinforcers were delivered immediately. When either an ascending or descending order of delay was in effect, responding was considered stable after 2 sessions with at least 80% choice of the larger reinforcer in the block with no delay and no more than 40% choice of the larger reinforcer in the block with the 32-s delay. After responding was stable, no-delay sessions were implemented until the rat responded predominantly for the larger reinforcer as described above, then either the same order of delay presentation was reintroduced or the alternate order of delay was introduced. Whether an ascending or a descending order was implemented was randomly determined, and the cycle of alternating delay (ascending or descending) with no-delay conditions continued for at least 8 presentations of each delay order prior to testing with amphetamine.
Rats were randomly assigned to be tested first under either an ascending or a descending order of delay with the constraint that 4 rats were first tested under each order. After being assigned to the first test condition, rats began receiving daily injections of saline 15-min prior to the start of each session. Tests with amphetamine began after at least 3 sessions under a particular delay order and a determination that the pattern of responding was stable for each individual rat as described above. Thereafter, other doses of amphetamine were tested so long as at least 3 baseline sessions occurred since the previous test and responding was stable; injections of amphetamine occurred no more frequently than once every 4 sessions. Doses were tested in irregular order across subjects and across conditions. After the effects of amphetamine were determined with one order of delay presentation, no-delay sessions were implemented until rats responded predominantly for the larger reinforcer for one session, followed by implementation of the alternate order of delay and then tests with amphetamine.
2.5. Experiment 2: Fixed delays
Following completion of the first experiment, rats chose between 1 and 3 food pellets delivered immediately. After responding was stable, rats either continued to respond with no delay or a delay to the larger reinforcer was introduced and the same delay was presented in all 5 blocks of the session. The delay was held constant (fixed) within and across sessions until responding was stable, at which point the effects of amphetamine were determined. Responding was considered stable when the total number of choices of the larger reinforce across all blocks of the session (ranging from 0, indicating exclusive responding for the smaller reinforcer, to 25, indicating exclusive responding for the larger reinforcer) did not vary by more than ± 5 for 3 consecutive sessions. The effects of amphetamine were determined in a manner similar to Experiment 1; that is, amphetamine was administered when at least 3 baseline sessions were conducted since the last test and responding was stable. After the effects of amphetamine were determined under a delay condition, no-delay sessions were implemented until responding was stable, followed by introduction of the next delay in the sequence of 0, 4, 8, 16, and 32 s. Four rats began with no delay and the delay to the larger reinforcer increased across conditions; the remaining rats began with a 32-s delay and the delay to the larger reinforcer decreased across conditions.
2.6. Data Analyses
For each session, the total number of trials completed, the time to complete each trial (latency), and the percentage of responses for the larger reinforcer during choice trials (lever responses for the larger reinforcer/ total responses * 100) were recorded for each block. For Experiment 1, delay functions were determined each session by plotting the percentage of responses for the larger reinforcer as a function of the delay, and area under the curve (AUC) was calculated using a trapezoidal rule (e.g., Myerson et al. 2001). At least half of the trials during a block (3 trials/block) had to be completed in order for data from that block to be included in the delay function, and data from all 5 blocks were required to calculate AUC. For each session in Experiment 2, percent choice of the larger reinforcer was averaged across each of the 5 blocks to yield an overall session mean. Under each delay condition, the session means across the last three sessions immediately prior the first test with amphetamine were averaged and plotted as a function of delay; thus, the delay function comprised the overall means from each of the 5 delay conditions and AUC was calculated as described above. For all comparisons, data from the 3 sessions immediately preceding the first test with amphetamine under that condition were used as control data.
Comparisons were analyzed using either one- or two-way repeated measures ANOVA with all independent variables (delay, dose, and delay condition) entered as within-subject factors. Post-hoc comparisons were conducted using Dunnett’s test. Analyses were conducted using NCSS 9 (Kaysville, UT) which allows for ANOVAs to be calculated when AUC values are missing for an individual subject (e.g., due to decreases in the number of trials completed). The significance level for all tests was set at p < .05.
2.7. Drugs
Amphetamine was provided by the National Institute on Drug Abuse (Research Technology Branch, Rockville, MD, USA) and dissolved in 0.9% sodium chloride (saline). Saline or amphetamine was administrated i.p. in a volume of 1.0 ml/kg of body weight 15-min prior to the rat being placed in the chamber and the start of the first block. Each dose of amphetamine was tested once per subject per condition, totaling 7 injections of each dose across the entire study.
3. Results
3.1. Training
After lever pressing was established, rats required an average of 2.2 (median=2, range=1–7), 5.1 (median=5, range=2–11), and 5.2 (median=5, range=2–10) sessions to satisfy training criteria under the no-delay, ascending, and descending training conditions, respectively, prior to the first test with amphetamine. During control sessions, rats responded predominantly for the larger reinforcer when both reinforcers were delivered immediately (Figure 1A, points above “0”), with responding for the larger reinforcer decreasing and responding for the smaller reinforcer increasing as a function of delay. Two way ANOVA revealed a significant main effect of delay [F(4,120) = 124.14, p < .001] but no main effect of delay order (p=.38) and no delay by order interaction (p=.27). Likewise, area under the curve (mean ± SEM) for the ascending (0.36 ± 0.05), descending (0.49 ± 0.04), and fixed (0.46 ± 0.04) conditions were not significantly different [F(2,24) = 1.88, p = .18]. Control AUC values for individual subjects (Figure 1B) were generally similar among conditions within individual rats with the exceptions of Rat 1 which showed a lower AUC under the ascending order as compared to the other two conditions, Rat 3 which showed a higher AUC under the descending condition as compared to the other two conditions, and Rat 4 which showed a higher AUC under the fixed-delay condition.
Fig 1.
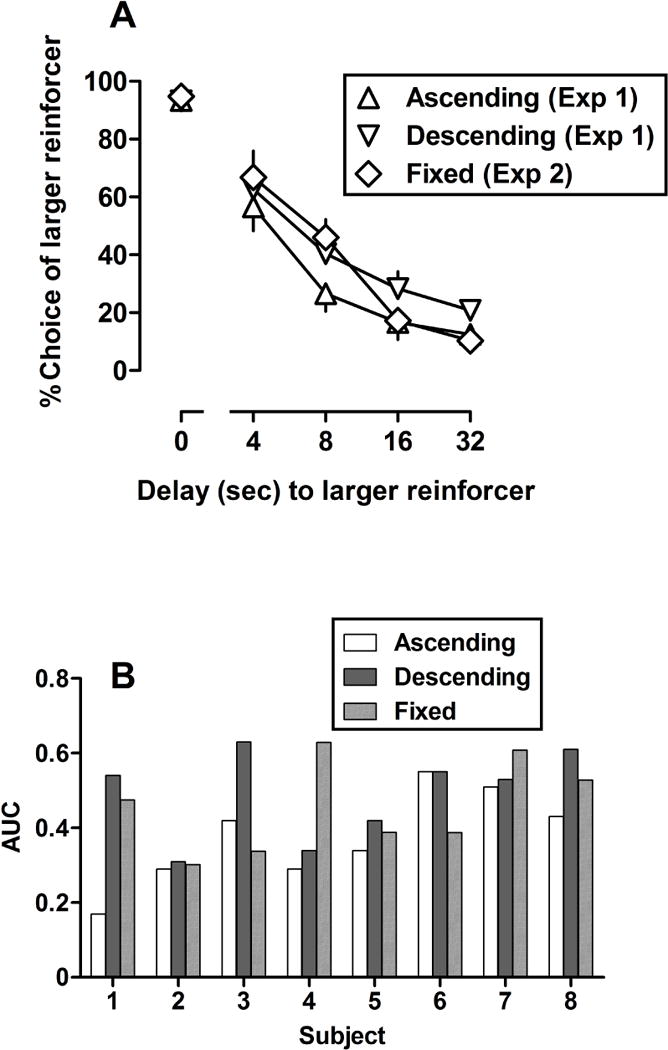
(A) Percent choice of the larger reinforcer plotted as a function of delay (in seconds) to the delivery of the larger reinforcer in the same group of rats (n=8) trained under the ascending (triangles), descending (inverted triangles), and fixed (diamonds) delay conditions. Data points indicate the group means and error bars indicate the standard error of the mean. (B) Area under the curve (AUC) for individual subjects responding under the ascending (white bars), descending (dark bars), and fixed (lightly shaded bars) delay conditions (see Data Analyses for further details).
3.2. Ascending order of delay
Under the ascending order of delay presentation (Figure 2, left panel), doses of 1.0 and 1.78 mg/kg of amphetamine significantly decreased choice of the larger reinforcer in the block with no delay (i.e., the first block of the session; points above “0”); however, amphetamine did not appear to markedly affect choice when delivery of the larger reinforcer was delayed (but see individual subject analysis below). Two-way ANOVA revealed a significant main effect of delay [F(4,152) = 37.2, p < .001] on percent choice of the larger reinforcer but no main effect of dose and no dose by delay interaction.
Fig 2.
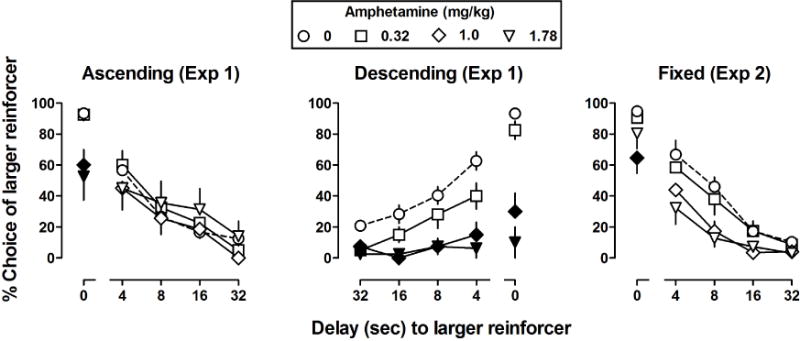
The effects of amphetamine on delay functions under the ascending (left), descending (center), and fixed (right) delay conditions. Filled symbols indicate that percent choice of the larger reinforcer was significantly different (p<.05) as compared to control. Other details are as in Fig 1.
3.3. Descending order of delay
Under the descending order of delay (Figure 2, middle panel), doses of 1.0 and 1.78 mg/kg of amphetamine decreased choice of the larger reinforcer across the entire range of delays, as well as during the block with no delay (i.e., the last block of the session; points above “0”). Two-way ANOVA revealed a significant main effect of delay [F(4,160) = 40.9, p < .001] and dose [F(3,160) = 61.7, p < .001] on percent choice of the larger reinforcer as well as a significant delay by dose interaction [F(12,160) = 5.9, p < .001].
3.4. Fixed delay
Under the fixed-delay condition (Figure 2, right panel), amphetamine significantly decreased choice of the larger reinforcer in the block with no delay. A two-way ANOVA on percent choice revealed a main effect of delay [F(4,158) = 102.1, p < .001] and a main effect of dose [F(3,158) = 14.2, p < .001] but no dose by delay interaction.
Figure 3A shows dose-effect functions for AUC under each condition. In correspondence with data shown in Figure 2, amphetamine did not have a significant effect on AUC under the ascending order of delay presentation (upright triangles). However, amphetamine dose-dependently decreased AUC under the descending order of delay (inverted triangles) with AUC following administration of each dose being significantly different from saline. Moreover, doses of 1.0 and 1.78 mg/kg of amphetamine significantly decreased AUC under the fixed-delay condition. Two way ANOVA for AUC revealed significant main effects for dose [F(3,93) = 14.5, p < .001] and order [F(3,93) = 7.3, p < .01] and a significant dose by delay interaction [F(6,93) = 3.6, p < .01].
Fig 3.
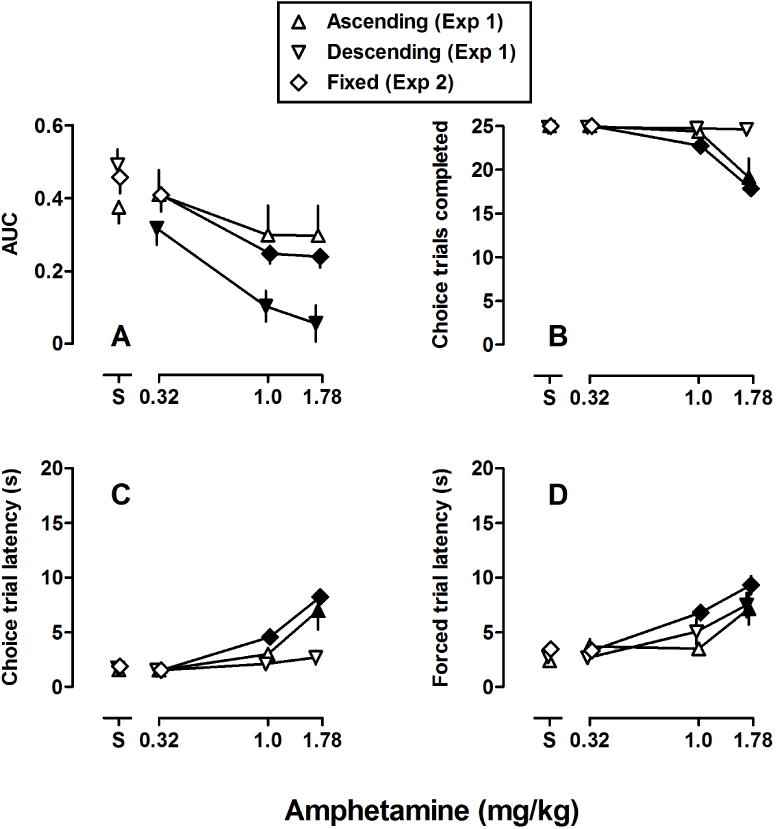
(A) AUC; (B) mean number of choice trials completed; (C) mean choice trial latency in s; and (D) mean forced trial latency in s. Data for rats (n=8) responding under the ascending (triangles), descending (inverted triangles), and fixed (diamonds) delay conditions are plotted as a function of dose of amphetamine in mg/kg body weight. Filled symbols indicate when the effects of amphetamine was significantly different (p<.05) as compared to saline (data above “S”).
Amphetamine significantly decreased the number of choice trials completed (Fig 3B) and increased the latency to complete choice and forced trials (Fig 3C and 3D, respectively). Two way ANOVA for choice trials completed revealed significant main effects for dose [F(3,96) = 27.8, p < .001] and order [F(2,96) = 10.7, p < .001] and a significant dose by delay interaction [F(6,96) = 3.6, p < .001]. For the latency to complete choice trials there were significant main effects for dose [F(3,96) = 36.2, p < .001] and order [F(2,96) = 12.3, p < .001] and a significant dose by delay interaction [F(6,96) = 5.7, p < .001]. For the latency to complete forced trials there were significant main effects for dose [F(3,96) = 28.0, p < .001] and order [F(2,96) = 4.5, p < .05] but no significant dose by delay interaction.
Figure 4 summarizes effects of amphetamine obtained in individual rats across each of three delay conditions. For each rat, the AUC following amphetamine administration is plotted as a percentage of the control AUC for that condition. Amphetamine increased AUC relative to control in some rats under the ascending order of delay and decreased AUC in others. Of the 24 total observations (8 rats × 3 doses), AUC was increased relative to control in 12 cases and decreased in 12 cases. In contrast, amphetamine predominantly decreased AUC under the descending order (middle panel) and during the fixed-delay condition (right panel). Of the 48 total observations across the descending and fixed delay conditions, AUC was decreased in 45 cases and increased slightly (the largest increase was 110% of control) in only 3 cases.
Fig 4.
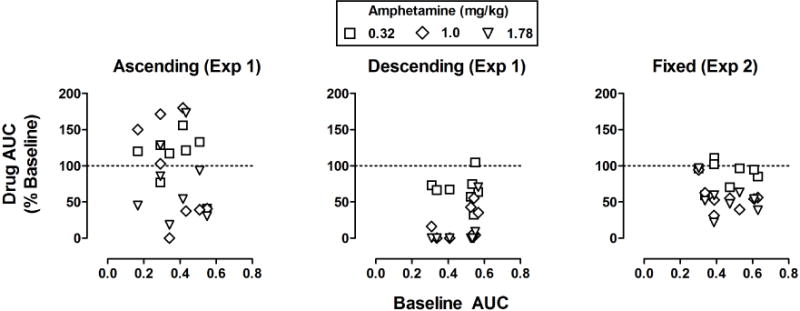
The percent change in AUC plotted as a function of baseline AUC for individual rats (n=8) under the ascending (left), descending (center), and fixed (right) delay conditions.
4. Discussion
Delay discounting is thought to be relevant to many socially important behavioral problems such as drug abuse (e.g., Bickel et al. 2013) and several procedures have been developed to study delay discounting in human and non-human subjects (Madden and Bickel 2010). Among the most commonly used is a procedure developed by Evenden and Ryan (1996) in which subjects choose between a smaller and a larger reinforcer with the delay to delivery of the larger reinforcer increasing progressively during the session. A recently published delay discounting study (Tanno et al. 2014) showed that the effects of amphetamine and methylphenidate varied between rats responding under an ascending order of delay and those responding under a descending order of delay; the same doses of amphetamine increased responding for a larger reinforcer in the former group and decreased responding for a larger reinforcer in the latter group. The current study determined whether this effect, possibly related to the order of delay presentation, also occurs within the same subject and whether amphetamine-induced changes in delay discounting are evident when delay is varied across rather than within sessions.
In the current study, responding maintained under both orders of delay presentation as well as under the fixed delay condition was sensitive to reinforcer amount and delay. When given a choice between 1 and 3 food pellets, both delivered immediately, rats responded predominantly for the larger reinforcer, and responding for the larger reinforcer decreased as a function of delay to its delivery. Delay functions generated under each delay condition were similar at the group level, yielding group-mean AUC values that were not significantly different. Given that control (no drug) delay functions were comparable among conditions, it is unlikely that performance under baseline contributed to the differences observed among groups in the effects of amphetamine.
Amphetamine increased responding for the larger, delayed reinforcer under the ascending order of delay presentation in some, but not all, rats. Increased choice of larger, delayed reinforcers is consistent with previous reports (e.g., Cardinal et al. 2000; Van Gaalen et al. 2006; Winstanley et al. 2003; 2005) and supports the view that amphetamine decreases delay discounting. However, that effect of amphetamine was observed in only half of the subjects and overall was less robust as compared with results from studies in which rats were trained under only an ascending order of delay (e.g., Tanno et al. 2014). That is, under the ascending order of delay presentation, amphetamine increased choice of the larger reinforcer in some rats and decreased choice of the larger reinforcer in others. In contrast, amphetamine either had no effect on or decreased responding for the larger reinforcer under the descending and fixed delay conditions, including when there was no delay. Notably, amphetamine failed to increase responding for the larger delayed reinforcer under the descending and fixed delay conditions, even in subjects that showed an increase in responding for the larger reinforcer under the ascending order.
Responding under the delay discounting procedures appears to be under the control of multiple behavioral processes (e.g., sensitivity to reinforcer delay and amount) and drug effects can result from perturbation of one or more of these processes. For example, amphetamine-induced increases in choice of larger delayed reinforcers can result from a reduction in sensitivity to reinforcer delay; that is, delays become functionally shorter resulting in less discounting of the delayed reinforcer (e.g., Pitts and Febbo 2004). If the effects of amphetamine on delay discounting were due simply to decreased sensitivity to delay, then amphetamine should increase responding for the larger reinforcer under a range of delay discounting procedures and conditions. Many, but not all (e.g., Evenden and Ryan 1996; Koffarnus et al. 2011), studies report that amphetamine increases responding for the larger reinforcer with increasing delay (e.g. attenuates delay discounting); however, in most studies, subjects were trained with one, usually ascending, order of the delay presentation. Only in one other published study (Slezak and Anderson 2009) were the same rats trained and tested with amphetamine under an ascending and a descending order of delay. That study also failed to show amphetamine-induced increases in choice of the larger delayed reinforcer under an ascending order of delay presentation. In fact, amphetamine decreased choice of the larger reinforcer at all delays as well as when both reinforcers were delivered immediately (Slezak and Anderson 2009, Fig 5). Studying the effects of amphetamine on responding under a descending order of delay presentation (Slezak and Anderson 2009; Tanno et al. 2014, Fig 2a; Experiment 1 of the current study) has consistently shown that amphetamine can decrease, rather than increase, choice of the larger reinforcer. Taken together, data from two prior studies (Slezak and Anderson 2009; Tanno et al. 2014) and from the current study strongly suggest that factors other than, possibly in addition to, sensitivity to delay contribute to the effects of amphetamine and other stimulant drugs on delay discounting.
Decreased choice of a larger delayed reinforcer suggests enhanced rather than reduced delay discounting. However, in many cases amphetamine also decreased responding for the larger reinforcer when it was delivered immediately. The effect of amphetamine on responding under conditions without delay would not be expected if drug effects reflected exclusively changes in delay discounting. One alternative possibility is that amphetamine reduced sensitivity to reinforcer amount, an effect that has been reported previously with amphetamine and methamphetamine (Pitts and Febbo 2004; Maguire et al. 2009). However, the effects observed in the component without delay under the descending order exceeded those observed under the ascending order and fixed delay conditions; choice of the larger reinforcer was decreased from greater than 90% under control conditions to 10% under the descending order, 53% under the ascending order, and 65% under the fixed delay condition. Moreover, a similar effect was reported in a previous study (Slezak and Anderson 2009, Table 3) using a similar procedure in which choice of the larger reinforcer was decreased in the component with no delay, under both orders of delay, with more marked effects occurring under the descending order. Thus, although amphetamine appeared to attenuate sensitivity to reinforcer amount, the magnitude of effect was modulated by the context in which it was studied (i.e., order of delay presentation). These data support the view that the effects of amphetamine were mediated through other processes (e.g., increased behavioral perseveration), insofar as rats responding under the descending order continued to respond on the lever delivering the smaller reinforcer despite the immediate delivery of the larger reinforcer (Tanno et al. 2014). Thus, under otherwise identical conditions, responding appeared to be insensitive to reinforcer amount under the descending order whereas under the ascending order this effect was less apparent, and under the fixed delay condition, the effect was negligible. It is possible that a perseverative effect of amphetamine interacts with other factors (delay and amount), resulting in qualitatively different patterns of responding. Taken together, these results demonstrate show that across a range of delay discounting conditions amphetamine decreases the slope of the delay function.
The results of the current study underscore the importance of considering how longer-term experimental history (e.g., training under different orders of delay presentation) might interact with the more recent history (e.g., current order of delay presentation) to impact the effects of amphetamine on delay discounting. However, the mechanisms underlying these effects remain unclear. Training with only one order can lead to the development of a persistent pattern of responding. Typical performance is characterized by a within-session shift in responding from the lever associated with the larger reinforcer to the lever associated with the smaller reinforcer (e.g., Evenden and Ryan 1996). Early in training, this shift in responding is likely under the control of variables such as reinforcer delay and amount. However, following extended exposure to the same experimental conditions, responding might come under the control of other variables such as the passage of time within the session [see Pitts and McKinney (2005) and Slezak and Anderson (2009) for discussions]. One method for assessing the contribution of training history might be to conduct systematic comparisons of drug effects early versus later in training. However, in a previous study (Tanno et al. 2014) neither the length of training under one particular order nor intervening tests with other drugs appeared to modulate the effects of amphetamine.
If perseveration contributes to the effects of amphetamine in delay discounting procedures, then those effects should be evident when variables other than delay are manipulated. Indeed, in a recent study (St. Onge et al. 2010) different groups of rats could choose between a smaller certain reinforcer and a larger uncertain reinforcer with the likelihood of receiving the larger reinforcer either increasing or decreasing within session. Amphetamine shifted the probability-discounting function upward in the group responding under the descending order of probability, increasing choice of the larger less certain reinforcer, but shifted the function downward with the ascending order, decreasing choice of the larger less certain reinforcer and demonstrating an interaction between order and the effects of amphetamine. Results of that study support the view that the effects of amphetamine under a probability discounting procedure involve mechanisms other than, or in addition to, sensitivity to delay, amount, or probability.
5. Conclusions
In conclusion, various delay discounting procedures have been developed and have proven to be very useful for studying behavioral, pharmacological, and neurobiological aspects of impulsive decision making; however, a better understanding of factors that impact baseline performance and those that modulate drug effects is necessary to interpret the effects of drugs and other manipulations on behavior that is as complex as delay discounting. This study, taken together with previous research (e.g., Tanno et al. 2014), demonstrates the complex interaction between pharmacological (drugs) and environmental variables and underscores the importance of the environment and history in drug effects.
Highlights.
Order of delay presentation can alter the effects of drugs on delay discounting
Amphetamine appears to attenuate discounting under an ascending order of delay
Amphetamine does not attenuate discounting under a descending order
Training with varied orders modifies the effects of amphetamine on discounting
Across a range of conditions, amphetamine decreases the slope of delay functions
Acknowledgments
Animals were maintained and experiments were conducted in accordance with the Institutional Animal Care and Use Committee, The University of Texas Health Science Center at San Antonio, and with the 2011 Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources on Life Sciences, National Research Council, and the National Academy of Sciences). This work was supported, in part, by United States Public Health Service Grants T32DA031115, F32DA035605 (DRM), R25NS080684 (CH), and K05DA17918 (CPF). Research reported in this publication was supported by the National Institute on Drug Abuse and the National Institute of Neurological Disorders And Stroke of the National Institutes of Health. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
The authors have no conflict of interest.
References
- Ainslie GW. Impulse control in pigeons. J Exp Anal Behav. 1974;21:485–489. doi: 10.1901/jeab.1974.21-485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbelivien A, Billy E, Lazarus C, Kelche C, Majchrzak M. Rats with different profiles of impulsive choice behavior exhibit differences in responses to caffeine and d-amphetamine and in medial prefrontal cortex 5-HT utilization. Behav Brain Res. 2008;187:273–283. doi: 10.1016/j.bbr.2007.09.020. [DOI] [PubMed] [Google Scholar]
- Bari A, Robbins TW. Inhibition and impulsivity: behavioral and neural basis of response control. Prog Neurobiol. 2013;108:44–79. doi: 10.1016/j.pneurobio.2013.06.005. [DOI] [PubMed] [Google Scholar]
- Bickel WK, Jarmolowicz DP, Mueller ET, Koffarnus MN, Gatchalian KM. Excessive discounting of delayed reinforcers as a trans-disease process contributing to addiction and other disease-related vulnerabilities: emerging evidence. Pharmacol Ther. 2012;134:287–297. doi: 10.1016/j.pharmthera.2012.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bickel WK, Koffarnus MN, Moody L, Wilson AG. The behavioral- and neuroeconomic process of temporal discounting: a candidate behavioral marker of addiction. Neuropharmacology. 2013 doi: 10.1016/j.neuropharm.2013.06.013. published ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardinal RN, Robbins TW, Everitt BJ. The effects of amphetamine,chlordiazepoxide, α-flupenthixol and behavioural manipulations on choice of signaled and unsignaled delayed reinforcement in rats. Psychopharmacology (Berl) 2000;152:362–375. doi: 10.1007/s002130000536. [DOI] [PubMed] [Google Scholar]
- de Wit H, Mitchell SH. Drug effects on delay discounting. In: Madden GJ, Bickel WK, editors. Impulsivity: the behavioral and neurological science of discounting. American Psychological Association; Washington D.C.: 2010. pp. 213–241. [Google Scholar]
- Evenden JL. Varieties of impulsivity. Psychopharmacology (Berl) 1999;146:348–361. doi: 10.1007/pl00005481. [DOI] [PubMed] [Google Scholar]
- Evenden JL, Ryan CN. The pharmacology of impulsive behaviour in rats: the effects of drugs on response choice with varying delays of reinforcement. Psychopharmacology (Berl) 1996;128:161–170. doi: 10.1007/s002130050121. [DOI] [PubMed] [Google Scholar]
- Huskinson SL, Krebs CA, Anderson KG. Strain differences in delay discounting between Lewis and Fischer 344 rats at baseline and following acute and chronic administration of d-amphetamine. Pharmacol Biochem Behav. 2012;101:403–416. doi: 10.1016/j.pbb.2012.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koffarnus MN, Newman AH, Grundt P, Rice KC, Woods JH. Effects of selective dopaminergic compounds on a delay-discounting task. Behav Pharmacol. 2011;22:300–311. doi: 10.1097/FBP.0b013e3283473bcb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logue AW. Research on self-control: an integrating framework. Behav Brain Sci. 1988;11:665–679. [Google Scholar]
- Madden GJ, Bickel WK. Impulsivity: the behavioral and neurological science of discounting. American Psychological Association; Washington D.C.: 2010. [Google Scholar]
- Maguire DR, Rodewald AM, Hughes CE, Pitts RC. Rapid acquisition of preference in concurrent schedules: effects of d-amphetamine on sensitivity to reinforcement amount. Behav Processes. 2009;81:238–243. doi: 10.1016/j.beproc.2009.01.001. [DOI] [PubMed] [Google Scholar]
- Mazur JE. An adjusting procedure for studying delayed reinforcement. In: Commons ML, Mazur JE, Nevin JA, Rachlin H, editors. Quantitative analyses of behavior. Vol. 5. The effect of delay and of intervening events on reinforcement value; Erlbaum, New Jersey: 1987. pp. 55–73. [Google Scholar]
- Myerson J, Green L, Warusawitharana M. Area under the curve as a measure of discounting. J Exp Anal Behav. 2001;76:235–243. doi: 10.1901/jeab.2001.76-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perry JL, Carroll ME. The role of impulsive behavior in drug abuse. Psychopharmacology (Berl) 2008;200:1–26. doi: 10.1007/s00213-008-1173-0. [DOI] [PubMed] [Google Scholar]
- Pitts RC, Febbo SM. Quantitative analyses of methamphetamine’s effects on self-control choices: implications for elucidating behavioral mechanisms of drug action. Behav Processes. 2004;66:213–233. doi: 10.1016/j.beproc.2004.03.006. [DOI] [PubMed] [Google Scholar]
- Pitts RC, McKinney AP. Effects of methylphenidate and morphine on delay-discount functions obtained within sessions. J Exp Anal Behav. 2005;83:297–314. doi: 10.1901/jeab.2005.47-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rachlin H, Green L. Commitment, choice and self-control. J Exp Anal Behav. 1972;17:15–22. doi: 10.1901/jeab.1972.17-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards JB, Mitchell SH, de Wit H, Seiden LS. Determination of discount functions in rats with an adjusting-amount procedure. J Exp Anal Behav. 1997;67:353–366. doi: 10.1901/jeab.1997.67-353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidman M. Tactics of scientific research. Basic Books; New York: 1960. [Google Scholar]
- Slezak JM, Anderson KG. Effects of variable training, signaled and unsignaled delays, and d-amphetamine on delay-discounting functions. Behav Pharmacol. 2009;20:424–436. doi: 10.1097/FBP.0b013e3283305ef9. [DOI] [PubMed] [Google Scholar]
- Slezak JM, Anderson KG. Effects of acute and chronic methylphenidate on delay discounting. Pharmacol Biochem Behav. 2011;99:545–551. doi: 10.1016/j.pbb.2011.05.027. [DOI] [PubMed] [Google Scholar]
- Slezak JM, Ricaurte GA, Tallarida RJ, Katz JL. Methylphenidate and impulsivity: a comparison of effects of methylphenidate enantiomers on delay discounting in rats. Psychopharmacology (Berl) 2013 doi: 10.1007/s00213-013-3220-8. published electronically ahead of print. [DOI] [PubMed] [Google Scholar]
- St Onge JR, Chiu YC, Floresco SB. Differential effects of dopaminergic manipulations on risky choice. Psychopharmacology (Berl) 2010;211:209–221. doi: 10.1007/s00213-010-1883-y. [DOI] [PubMed] [Google Scholar]
- Ta WM, Pitts RC, Hughes CE, McLean AP, Grace RC. Rapid acquisition of preference in concurrent chains: effects of d-amphetamine on sensitivity to reinforcement delay. J Exp Anal Behav. 2008;89:71–91. doi: 10.1901/jeab.2008.89-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanno T, Maguire DR, Henson C, France CP. Effects of amphetamine and methylphenidate on delay discounting in rats: interactions with order of delay presentation. Psychopharmacology (Berl) 2014;231:85–95. doi: 10.1007/s00213-013-3209-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Gaalen MM, van Koten R, Schoffelmeer ANM, Vanderschuren LJMJ. Critical involvement of dopaminergic neurotransmission in impulsive decision making. Biol Psychiatry. 2006;60:66–73. doi: 10.1016/j.biopsych.2005.06.005. [DOI] [PubMed] [Google Scholar]
- Winstanley CA, Dalley JW, Theobald DEH, Robbins TW. Global 5-HT depletion attenuates the ability of amphetamine to decrease impulsive choice on a delay-discounting task in rats. Psychopharmacology (Berl) 2003;170:320–331. doi: 10.1007/s00213-003-1546-3. [DOI] [PubMed] [Google Scholar]
- Winstanley CA, Theobald DEH, Dalley JW, Robbins TW. Interactions between serotonin and dopamine in the control of impulsive choice in rats: therapeutic implications for impulse control disorders. Neuropsychopharmacology. 2005;30:669–682. doi: 10.1038/sj.npp.1300610. [DOI] [PubMed] [Google Scholar]