

Abstract
Three technologies have emerged as therapies to restore light sensing to profoundly blind patients suffering from late-stage retinal degenerations: (1) retinal prosthetics, (2) optogenetics, and (3) chemical photoswitches. Prosthetics are the most mature and the only approach in clinical practice. Prosthetic implants require complex surgical intervention and provide only limited visual resolution but can potentially restore navigational ability to many blind patients. Optogenetics uses viral delivery of type 1 opsin genes from prokaryotes or eukaryote algae to restore light responses in survivor neurons. Targeting and expression remain major problems, but are potentially soluble. Importantly, optogenetics could provide the ultimate in high-resolution vision due to the long persistence of gene expression achieved in animal models. Nevertheless, optogenetics remains challenging to implement in human eyes with large volumes, complex disease progression, and physical barriers to viral penetration. Now, a new generation of photochromic ligands or chemical photoswitches (azobenzene-quaternary ammonium derivatives) can be injected into a degenerated mouse eye and, in minutes to hours, activate light responses in neurons. These photoswitches offer the potential for rapidly and reversibly screening the vision restoration expected in an individual patient. Chemical photoswitch variants that persist in the cell membrane could make them a simple therapy of choice, with resolution and sensitivity equivalent to optogenetics approaches. A major complexity in treating retinal degenerations is retinal remodeling: pathologic network rewiring, molecular reprogramming, and cell death that compromise signaling in the surviving retina. Remodeling forces a choice between upstream and downstream targeting, each engaging different benefits and defects. Prosthetics and optogenetics can be implemented in either mode, but the use of chemical photoswitches is currently limited to downstream implementations. Even so, given the high density of human foveal ganglion cells, the ultimate chemical photoswitch treatment could deliver cost-effective, high-resolution vision for the blind.
Keywords: Retinal degeneration, blindness, prosthetics, optogenetics, chemical photoswitches, remodeling, reprogramming
The international effort to restore vision to the profoundly blind spans diverse approaches such as computational modeling,1 electrical and bioengineering of prosthetic microelectronics,2−7 suppressive8,9 and reparative gene therapies,10 cell therapies,11 optogenetics therapies,12−16 and now photochromic ligand or chemical photoswitch therapies.17,18 A key challenge in evaluating these varied approaches is developing a common assessment platform based on animal models that mimic late-stage human retinal degenerations.19 Compared to other strategies, chemical photoswitch therapies provide fast, flexible, nonsurgical, nongenetic, and reversible tools for profiling and treating patients as well as for optimizing therapies in animal models.17,18
The Retina
The retina is a sheet of brainlike tissue embedded in complex support architecture of transport epithelia and Müller cells (glia) that both isolate the retina from its vascular beds and serve as its nutritional support system. The retina is composed of a discrete sensory layer of photoreceptor cells that synaptically drive a discrete neural layer of processing networks composed of over 60 kinds of neurons.20 Mammalian rod and cone photoreceptor cells drive different sets of glutamatergic bipolar cells that ultimately synapse on amacrine and/or ganglion cell targets in the inner plexiform layer (Figure 1A). Ganglion cells are ultimately separable into ∼15–20 classes that project to a range of central nervous system loci, providing the critical information for perceptual vision, visual world mapping to coordinate eye movements and gaze, and non-image-forming operations such as circadian phase-setting.
Figure 1.
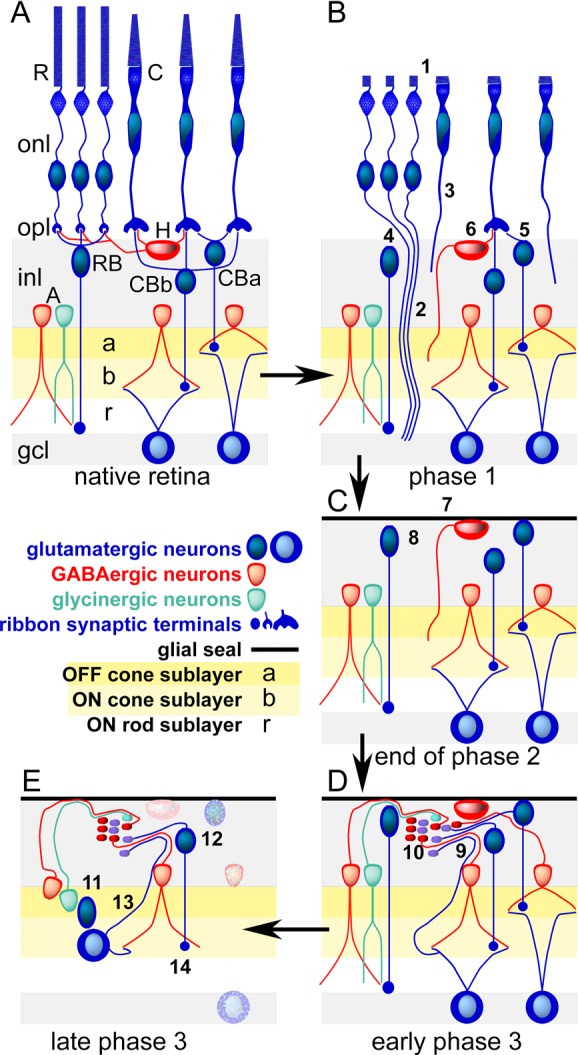
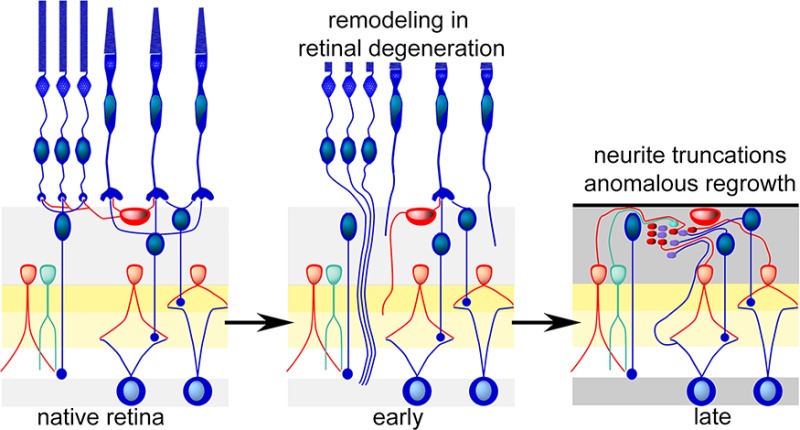
Cellular composition of the retina and the three phases of remodeling associated with retinal degenerations. (Panel A) The laminated retina with rod (R) and cone (C) photoreceptor cells in the outer nuclear layer (onl) and synaptic endings in the outer plexiform layer (opl) driving rod bipolar cells (RB), ON (CBb) or OFF (CBa) cone bipolar cells, and/or horizontal cells (H). Rod and cone bipolar cells then drive separate sets of GABAergic (red) and glycinergic (green) amacrine cells (A) in the OFF (a), ON (b), and rod (r) layers of the inner plexiform layer. Only cone bipolar cells drive ganglion cells that project to the brain through the optic nerve. The ganglion cell layer (gcl) contains the cell bodies of ganglion cells positioned next to the vitreous of the eye. It is the first layer accessible through surgical approaches or intraocular injections. (Panel B) Phase 1. Stressed photoreceptor cells truncate their outer segments (1) and extend anomalous axons deep into the retina (2,3). Bipolar cells retract dendrites (4,5) and horizontal cells generate ectopic axons. (Panel C) End of phase 2. Death of photoreceptor cells and ablation of the outer nuclear layer. The remnant retina is capped by a seal of glial processes (7) and all bipolar cells have truncated their dendrites. (Panel D) Early phase 3. All surviving cells can initiate neuritogenesis, forming ectopic fascicles of processes (9) and new synaptic microneuromas (10). (Panel E) Late phase 3. Cells revise their connection repertoires (11) and some migrate to ectopic sites (12), ganglion cells generate new intraretinal axons (13), and rod bipolar cells decrease their synaptic ribbon size (14). Neuronal cell death progresses (stippled cells).
The Scope of Retinal Degenerations
Retinal degenerations are inherited disorders that directly or indirectly cause photoreceptor cell death. The majority are associated with rod photoreceptor cells. Many of these disorders are phenomenologically called retinitis pigmentosa (RP), a name that reflects the common disease phenotype of exposed pigmentation (when viewed by ophthalmoscopy) from invading retinal pigmented epithelium (RPE) cells. RP and over 9 other classes of retinal dystrophies are collections of orphan diseases20 arising from over 200 source gene defects.21 In most instances, dominant or recessive defects in various rod photoreceptor genes lead to the progressive degeneration and death of rods. In the process, the critical cone cells of high-acuity day vision are decimated by bystander killing (e.g., microglial killing) or substantially damaged and remodeled. In humans, foveal cone vision may persist for many years as the surrounding retina degenerates, but eventually many RP patients suffer total vision loss (abbreviated clinically as NLP: no light perception) or are reduced to impaired bare light perception (BLP). The incidence is variable (1 in 2000–7000), and ∼1 to 1.5 million people are afflicted globally. However, there are but a few hundred to several thousand cases worldwide of any given genotype. Disease from different genotypes can progress very differently, with varying severity in speed of vision loss,22 and many forms have delayed adult onset. Thus, developing a homogeneous patient cohort for testing therapeutic interventions is virtually impossible and most patients are never genotyped. Most genetic therapies proposed for these diseases require early, even perinatal interventions. Indeed, the most publicized gene therapies treat not degenerations sensu stricto, but rather metabolic disorders. For example, some forms of autosomal recessive Leber Congenital Amaurosis (LCA) arise from defects in the RPE65 gene expressed in RPE cells.23 RPE65 is essential for the synthesis of the retinoid 11-cis-retinaldehyde, required by all photoreceptor cells for light sensing. Photoreceptor cells lacking 11-cis-retinaldehyde can survive long periods without degenerating, allowing late onset rescue of function by adeno-associated virus (AAV) mediated gene therapy.24,25 But what of those already afflicted and in advanced stages of visual impairment? Gene therapy approaches are, so far, not options for patients who have such advanced photoreceptor cell degenerations. Before we address possible solutions, we must deal with one more complexity of retinal degenerations: remodeling.
Remodeling
Loss of photoreceptor cells represents deafferentation. All known deafferentation events trigger downstream changes in target neural structures.26 Deafferented neurons remodel in a variety of ways, including revision of cell shape and processes, as well as reprogramming of gene expression. The surviving neural retina remodels in response to the overall loss of photoreceptor cells and cone photoreceptors in particular19,27−31 via a spectrum of revisions independent of the genetic type of retinal degeneration. Figure 1B–E traces the progression of remodeling through its characteristic phases, spanning 90–600 days in rodents with different gene defects.27 In phase 1 (Figure 1B), stressed photoreceptor cells begin to lose their outer segments and transiently begin disconnecting from their target bipolar cells, extending anomalous neurites into the neural retina. Bipolar cells begin to retract their dendrites, and horizontal cells also extend anomalous axons into the inner plexiform layer. Phase 2 (Figure 1C) is characterized by comprehensive death of the photoreceptor cell layer and ends with the formation of a glial seal atop the remnant neural retina and truncation of all bipolar cell dendrites. Phase 3 (Figure 1D, E) is a period of complex revision that progresses throughout life, with the generation of new neurites and evolution of anomalous synaptic microneuromas, cell migration, and cell death, including loss of ganglion cells in humans and animal models.
There are three critical features of remodeling. First, anomalous neuritogenesis that corrupts signaling is initiated by retinoic acid receptor pathways.32 Thus, restoring light signaling by any means (electrical, optogenetics, chemical photoswitches) is not likely to alter this aspect of remodeling. Second, downstream glutamate receptor reprogramming events are triggered30,33 and it is not clear that these are reversible. Rescue schemas targeting upstream neurons (cones or bipolar cells) with payloads such as channelrhodopsins or other photoproteins still must cope with the progressive loss of downstream retinal neurons and rewiring. Finally, the retina uses multiple neurotransmitter cycles34 and there is now evidence that the heterocellular glutamate-glutamine cycle essential for neural signaling is altered in both animal models of retinal degeneration and human RP.31,35,36
Restoring Signaling
Three strategies for restoring light-driven signaling in the surviving neural retina currently have clinical potential: retinal prosthetics, optogenetics, and chemical photoswitches. The outcomes of each depend on which survivor cells are chosen as targets, the sensitivity of transduction, signaling speed, and the perdurance of the intervention. Based on our understanding of the division of signal outflow of normal retina into a variety of ON, OFF, and ON–OFF ganglion cells that encode a range of fundamental operations for vision (Figure 2), we can estimate the best-case scenario outcomes for each schema. Globally driving upstream cells potentially exploits extant networks, amplification, and encoding, especially generation of ON and OFF polarity signals in their appropriate ganglion cell classes (Figure 3A). Driving downstream cells avoids network remodeling and reprogramming likely to corrupt upstream signaling, but faces the challenge of driving different classes of ganglion cells with appropriate polarities, sensitivities, kinetics, and trigger features (Figure 3B). So far, no reliable means of targeting different ganglion cell classes has been found.
Figure 2.

Signal flow in the retina and the consequences of upstream and downstream stimulation. The retina has four layers of processing. Layer 1 is the photoreceptor cell layer and contains rods (r), long wave sensitive L-cones, and short wave sensitive S-cones. They drive layer 2 bipolar cells with sign-conserving (filled arrows) or sign-inverting (open arrows) synapses. Bipolar cell classes include rod, S-cone and a variety of L-cone types that are temporally fast or slow, spatially narrow or wide, and either ON or OFF (bar supra). Bipolar cells drive ganglion cells via excitatory glutamate synapses (implied, not shown). Layer 3 amacrine cell networks collect and shape bipolar cell signals (dotted lines) into a collection of layer 4 ganglion cell types that signal a range of structural and motion events in the visual system. DS, directionally selective; LED, local edge detector. The waveforms indicate the response to a nominal 1 Hz square wave ON–OFF light input. Layer 2.5 is a specialized amacrine cell (AII) that collects rod (r) bipolar cell signals and distributes them to ON cone bipolar cells via gap junctions (crossbar) or sign-inverting inhibitory glycinergic synapses (double open arrows).
Figure 3.

Retinal prosthetic networks. (Panel A) Upstream stimulation by subretinal or suprechoroidal electrodes. Electrodes drive a patch of bipolar cells of different classes with the same polarity. After layer 4 processing, many functions are normal but some are inverted. (Panel B) Downstream stimulation by epiretinal electrodes. All ganglion cell classes are driven with the same polarity and waveform of voltage.
All three strategies have clinical potential. By far, the leading technology is the Argus II retinal prosthetic,37−39 a stimulation array with 60 platinum electrodes (60 × 10) surgically positioned over the inner surface of retina (the epiretinal configuration), coupled to an embedded scleral electronics package via a transscleral ribbon cable and driven by a head-mounted camera. The Argus II system permits significant recovery of positional, motion, and structural percepts in many but not all patients. The driving concept is that the array will drive patches of retinal ganglion cells and that induced percepts, despite being coarse, will allow recovery of mobility and independent living.38 The Argus II specifically targets retinal ganglion cells in an array spanning roughly 20° of visual angle, corresponding to each electrode addressing an area roughly 0.1 mm in diameter, covering ∼20–50 retinal ganglion cells (Figure 3) depending on retinal eccentricity.40,41 This generates the same pattern of signaling in all classes of ganglion cells, which is not the norm (Figure 3B). Further electrode and stimulus refinement could improve epiretinal array resolution42 and lower thresholds.43 Alternate schemas include subretinal implants positioned in the remnant space between the RPE and the surviving retina6,44 and suprachoroidal implants where the stimulating array is completely positioned in the choroid, outside the retinal space altogether.45,46 These upstream technologies drive full remnant networks, including rewired and reprogrammed elements (Figure 3A). At present, there are perhaps 100 patient implants in varied epiretinal and subretinal device trials.
Optogenetic approaches have not yet reached clinical testing, but should soon. Optogenetics therapies are nonsurgical; their spatial resolution should match surviving networks and they can be targeted to specific cells. There have been numerous reviews of optogenetics, and we only gloss the concept. Genes for various type 1 opsins are packaged with promoter or enhancer elements into varied serotypes of AAV capsids and delivered by subretinal or intravitreal injection as nonreplicating, long-expression episome systems in mammalian neurons. By hijacking existing trafficking mechanisms, the translated proteins are sometimes successfully delivered to the cell membrane, acquire residual retinoids, and become photosensitive. Some versions require additional trafficking motifs to enhance expression. Activated type 1 opsins can trigger sufficient current flow to modulate cell voltage, transmitter release and/or spiking. Cation-selective channelrhodopsin-2 variants (ChR2) from Chlamydomonas reinhardtii(47) and anion-selective halorhodopsin variants (NpHR) from Natronomonas phoraonis(48,49) are the major opsins in use. Implementation problems include targeting appropriate cells via AAV capsid variants,50 efficient regional expression, perduring expression during remodeling, and low sensitivity. Some studies have tested sensitive type 2 mammalian opsins such as melanopsin, and, while effective, these opsins signal too slowly to achieve functional vision when expressed in ganglion cells.51 Different transduction mechanisms in ON bipolar cells may allow direct control of melanopsin transduction turnoff, yielding faster kinetics, however. But the quandary remains: is it best to target upstream or downstream elements (Figure 4)? For example, expression of NpHR in remnant mouse cones in early stages of retinal degeneration appears to properly trigger different classes of responses in ganglion cells.14 But such cones eventually die, and no one has yet shown that optogenetics interventions can retard remodeling. Bipolar cells are longer-lived, and ON bipolar cells have been targeted with ChR2 (Figure 3B) via their mGluR6 pathways.12,15 However, mGluR6 trafficking in this pathway also seems corrupted over time.52,53 Upstream remodeling might be avoided by targeting ganglion cells with ChR2 (Figure 4A), which leads to behavioral light detection in mice.54 This leads us to the use of chemical photoswitches.
Figure 4.

Chemical signaling schemas. (Panel A) Upstream stimulation by targeting ChR2 to either ON bipolar cells or the AII amacrine cell. In principle, this provides the most faithful pathway encoding, barring corruption by remodeling. (Panel B) Downstream stimulation by targeting ChR2 or DENAQ to ganglion cells. The entire ganglion cell stimulus-response domain is homogeneous, as we lack tools to differentially target and drive individual ganglion cells.
Fast Chemical Photoswitches
A completely different strategy involves the use of photochromic ligands or chemical photoswitches: small molecules that modulate neuronal electrical activity.55 Polosukhina et al.18 injected AAQ (acrylamide-azobenzene-quaternary ammonium) intravitreally in rd1 mice (an aggressive retinal degeneration) and readily generated light-driven activity in otherwise silent ganglion cells. trans-AAQ is an effective K-channel blocker, which in turn induces neuronal excitation; photoisomerization driven by short wave light (∼380 nm) to cis-AAQ unblocks K-channels, theoretically enabling outward currents to silence the cell.56 AAQ can slowly thermally isomerize back to trans or be reisomerized with long wave light (∼540 nm). While this was an exciting advance, retinal injections of AAQ elicited rather poor photosensitivity, reversed quickly, and appeared to target all cells indiscriminately. And, paradoxically, trans → cis AAQ photoconversions excited rather than silenced ganglion cells in the retina. It turned out that most of the light-driven activity triggered by AAQ originated in inhibitory amacrine cells rather than in ganglion cells. Since amacrine cells make up over 98% of the neuronal membrane density in the remnant inner plexiform layer57,58 and are inhibitory, this explains the sign-inverting switch.
A second-generation chemical photoswitch, DENAQ (diethylamino-azobenzene-quaternary ammonium), solves many of these problems.17,55 Its action spectrum is red-shifted relative to AAQ, and it is much more photosensitive, rendering it competitive with ChR2 and NpHR opsins. It is fast acting but with a much longer physiological half-life of 2.1 days. Importantly, it rapidly isomerizes thermally to the inactive state. But the key ingredient here is serendipity. Phase 3 retinal degeneration involves significant reprogramming of signaling pathways,30,31 and we only know a small part of this repertoire. Corruptive spontaneous spiking in ganglion cells from retinal degeneration models has both network-driven and cell-autonomous components.59−62 It now appears that ganglion cells in the rd1 mouse either upregulate expression of hyperpolarization-activated cyclic-nucleotide gated (HCN) cation channels that generate rebound spiking in neurons63 and/or some other mechanism facilitates selective loading with DENAQ. For reasons unknown, DENAQ appears to target HCN channels as well as K channels. Thus, the application of DENAQ to normal retinas evokes little responsivity, but elicits large responses from rd1 and rd4 mice.17
DENAQ represents a fast tool to target retinal ganglion cells but is limited to downstream targeting (Figure 4B), with all the defects appertaining thereto. Chemical photoswitches act rapidly after a single intravitreal injection and are apparently nontoxic. They could be used to screen patients, establishing baseline visual performance against which other therapies could be calibrated. Targeting high-density human foveal ganglion cells with chemical photoswitches may rapidly generate high resolution vision, and developing either photoswitches with longer physiological half-lives or delivery strategies providing long-term depots could make chemical photoswitches an option of choice. But, in the end, would not it be marvelous if all these technologies worked well?
Author Contributions
R.M. performed the literature review and wrote the manuscript. B.J. reviewed the manuscript, focusing on remodeling concepts. R.P. reviewed the manuscript, focusing on optogenetics and metabolism in retinal degenerations.
NIH R01 EY02576 (R.M.), NIH EY R01 EY015128 (R.M.), NIH P30 EY14800 (R.M.), Research to Prevent Blindness.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
References
- Nirenberg S.; Pandarinath C. (2012) Retinal prosthetic strategy with the capacity to restore normal vision. Proc. Natl. Acad. Sci. U.S.A. 109, 15012–15017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margalit E.; Maia M.; Weiland J. D.; Greenberg R. J.; Fujii G. Y.; Torres G.; Piyathaisere D. V.; O’Hearn T. M.; Liu W.; Lazzi G.; Dagnelie G.; Scribner D. A.; de Juan E. J.; Humayun M. S. (2002) Retinal prosthesis for the blind. Surv. Ophthalmol. 47, 335–356. [DOI] [PubMed] [Google Scholar]
- Zrenner E. (2002) Will Retinal Implants Restore Vision?. Science 295, 1022–1025. [DOI] [PubMed] [Google Scholar]
- Eberdt M.; Brown P. K.; Lazzi G. (2003) Two-dimensional SPICE-linked multiresolution impedance method for low-frequency electromagnetic interactions. IEEE Trans. Biomed. Eng. 50, 881–889. [DOI] [PubMed] [Google Scholar]
- Xie J.; G.J W.; Yow L.; Humayun M. S.; Weiland J. D.; Cela C. J.; Jadvar H.; Lazzi G.; Dhrami-Gavazi E.; Tsang S. H. (2012) Preservation of retinotopic map in retinal degeneration. Exp. Eye Res. 98, 88–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stingl K.; Bartz-Schmidt K. U.; Besch D.; Braun A.; Bruckmann A.; Gekeler F.; Greppmaier U.; Hipp S.; Hörtdörfer G.; Kernstock C.; Koitschev A.; Kusnyerik A.; Sachs H.; Schatz A.; Stingl K. T.; Peters T.; Wilhelm B.; Zrenner E. (2013) Artificial vision with wirelessly powered subretinal electronic implant alpha-IMS. Proc. Biol. Sci. 280, 20130077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zrenner E. (2013) Fighting blindness with microelectronics. Sci. Transl. Med. 5, 210–216. [DOI] [PubMed] [Google Scholar]
- Jiang L.; Li T. Z.; Boye S. E.; Hauswirth W. W.; Frederick J. M.; Baehr W. (2013) RNAi-mediated gene suppression in a GCAP1(L151F) cone-rod dystrophy mouse model. PLoS One 8, e57676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang L.; Zhang H.; Dizhoor A. M.; Boye S. E.; Hauswirth W. W.; Frederick J. M.; Baehr W. (2011) Long-term RNA interference Gene Therapy in a dominant retinitis pigmentosa mouse model. Proc. Natl. Acad. Sci. U.S.A. 108, 18476–18481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stieger K.; Cronin T.; Bennett J.; Rolling F. (2011) Adeno-associated virus mediated Gene Therapy for retinal degenerative diseases. Methods Mol. Biol. 807, 179–218. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Cordero A.; West E. L.; Pearson R. A.; Duran Y.; Carvalho L. S.; Chu C. J.; Naeem A.; Blackford S. J.; Georgiadis A.; Lakowski J.; Hubank M.; Smith A. J.; Bainbridge J. W.; Sowden J. C.; Ali R. R. (2013) Photoreceptor precursors derived from three-dimensional embryonic stem cell cultures integrate and mature within adult degenerate retina. Nat. Biotechnol. 31, 741–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lagali P. S.; Balya D.; Awatramani G. B.; Münch T. A.; Kim D. S.; Busskamp V.; Cepko C. L.; Roska B. (2008) Light-activated channels targeted to ON bipolar cells restore visual function in retinal degeneration. Nat. Neurosci. 11, 667–675. [DOI] [PubMed] [Google Scholar]
- Busskamp V.; Picaud S.; Sahel J. A.; Roska B. (2012) Optogenetic therapy for retinitis pigmentosa. Gene Ther. 19, 169–175. [DOI] [PubMed] [Google Scholar]
- Busskamp V.; Duebel J.; Baly D.; Fradot M.; Viney T. J.; Siegert S.; Groner A. C.; Cabuy E.; Forster V.; Seeliger M.; Biel M.; Humphries P.; Paques M.; Mohand-Said S.; Trono D.; Deisseroth K.; Sahel J. A.; Picaud S.; Roska B. (2010) Genetic reactivation of cone photoreceptors restores visual responses in retinitis pigmentosa. Science 329, 413–417. [DOI] [PubMed] [Google Scholar]
- Doroudchi M. M.; Greenberg K. P.; Liu J.; Silka K. A.; Boyden E. S.; Lockridge J. A.; Arman A. C.; Janani R.; Boye S. E.; Boye S. L.; Gordon G. M.; Matteo B. C.; Sampath A. P.; Hauswirth W. W.; Horsager A. (2011) Virally delivered channelrhodopsin-2 safely and effectively restores visual function in multiple mouse models of blindness. Mol. Ther 19, 1220–1229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutter M.; Swietek N.; Münch T. A. (2014) Salvaging ruins: reverting blind retinas into functional visual sensors. Methods Mol. Biol. 1148, 149–160. [DOI] [PubMed] [Google Scholar]
- Tochitsky I.; Polosukhina A.; Degtyar V. E.; Gallerani N.; Smith C. M.; Friedman A.; Van Gelder R. N.; Trauner D.; Kaufer D.; Kramer R. H. (2014) Restoring visual function to blind mice with a photoswitch that exploits electrophysiological remodeling of retinal ganglion cells. Neuron 81, 800–813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polosukhina A.; Litt J.; Tochitsky I.; Nemargut J.; Sychev Y.; De Kouchkovsky I.; Huang T.; Borges K.; Trauner D.; Van Gelder R. N.; Kramer R. H. (2012) Photochemical restoration of visual responses in blind mice. Neuron 75, 271–282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones B. W.; Kondo M.; Terasaki H.; Lin Y.; McCall M.; Marc R. E. (2012) Retinal remodeling. Jpn. J. Ophthalmol. 56, 289–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marc R. E. (2010) Synaptic Organization of the Retina. In Adler’s Physiology of the Eye (Levin L. A., Nilsson S. F. E., Ver Hoeve J., Wu S. M., Kaufman P. L., and Alm A., Eds.), pp 443–458, Elsevier, Amsterdam. [Google Scholar]
- Daiger S. P., Sullivan L. S., and Bowne S. (2014) http://www.sph.uth.tmc.edu/Retnet/ (accessed 23 July 2014).
- Schuster A.; Weisschuh N.; Jägle H.; Besch D.; Janecke A. R.; Zierler H.; Tippmann S.; Zrenner E.; Wissinger B. (2005) Novel rhodopsin mutations and genotype-phenotype correlation in patients with autosomal dominant retinitis pigmentosa. Br. J. Ophthalmol. 89, 1258–1264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redmond T. M.; Yu S.; Lee E.; Bok D.; Hamasaki D.; Chen N.; Goletz P.; Ma J. X.; Crouch R. K.; Pfeifer K. (1998) Rpe65 is necessary for production of 11-cis-vitamin A in the retinal visual cycle. Nat. Genet. 20, 344–351. [DOI] [PubMed] [Google Scholar]
- Acland G. M.; Aguirre G. D.; Ray J.; Zhang Q.; Aleman T. S.; Cideciyan A. V.; Pearce-Kelling S. E.; Anand V.; Zeng Y.; Maguire A. M.; Jacobson S. G.; Hauswirth W. W.; Bennett J. (2001) Gene Therapy restores vision in a canine model of childhood blindness. Nat. Genet. 28, 92–95. [DOI] [PubMed] [Google Scholar]
- den Hollander A. I.; Black A.; Bennett J.; Cremers F. P. (2010) Lighting a candle in the dark: advances in genetics and gene therapy of recessive retinal dystrophies. J. Clin. Invest. 120, 3042–3053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perederiy J. V.; Westbrook G. L. (2013) Structural plasticity in the dentate gyrus- revisiting a classic injury model. Front. Neural Circuits 7, 17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones B. W.; Watt C. B.; Frederick J. M.; Baehr W.; Chen C. K.; Levine E. M.; Milam A. H.; LaVail M. M.; Marc R. E. (2003) Retinal remodeling triggered by photoreceptor degenerations. J. Comp. Neurol. 464, 1–16. [DOI] [PubMed] [Google Scholar]
- Marc R. E.; Jones B. W.; Watt C. B.; Strettoi E. (2003) Neural Remodeling in Retinal Degeneration. Prog. Retinal Eye Res. 22, 607–655. [DOI] [PubMed] [Google Scholar]
- Marc R. (2010) Injury and Repair: Retinal Remodeling. In Encyclopedia of the Eye (Dana R., Ed.), pp 414–420, Elsevier, Amsterdam. [Google Scholar]
- Marc R. E.; Jones B. W.; Anderson J. R.; Kinard K.; Marshak D. W.; Wilson J. H.; Wensel T.; Lucas R. J. (2007) Neural reprogramming in retinal degeneration. Invest. Ophthalmol. Visual Sci. 48, 3364–3371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones B. W.; Kondo M.; Terasaki H.; Watt C. B.; Rapp K.; Anderson J. R.; Lin Y.; Shaw M. V.; Yang J. H.; Marc R. E. (2011) Retinal remodeling in the Tg P347L rabbit, a large-eye model of retinal degeneration. J. Comp. Neurol. 519, 2713–2733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y.; Jones B. W.; Liu A.; Tucker J. F.; Rapp K.; Luo L.; Baehr W.; Bernstein P. S.; Watt C. B.; Yang J. H.; Shaw M. V.; Marc R. E. (2012) Retinoid receptors trigger neuritogenesis in retinal degenerations. FASEB J. 26, 81–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y.; Jones B. W.; Liu A.; Vazquez-Chona F. R.; Lauritzen J. S.; Ferrell W.; Marc R. E. (2012) Rapid glutamate receptor 2 trafficking during retinal degeneration. Mol. Neurodegener. 7, 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marc R. E. (2004) Retinal Neurotransmitters. In The Visual Neurosciences (Chalupa L. M., and Werner J., Eds.), pp 315–330, MIT Press, Cambridge, MA. [Google Scholar]
- Marc R.; Jones B.; Watt C.; Vazquez-Chona F.; Vaughan D.; Organisciak D. (2008) Extreme retinal remodeling triggered by light damage: Implications for AMD. Mol. Vision 14, 782–806. [PMC free article] [PubMed] [Google Scholar]
- Pfeiffer R. L.; Jones B. W.; Marc R. E. (2014) Metabolic changes associated with Muller cells in a transgenic rabbit model of retinal degeneration. Invest. Ophthalmol. Visual Sci. 55, 829. [Google Scholar]
- Ahuja A. K.; Behrend M. R. (2013) The Argus II retinal prosthesis: factors affecting patient selection for implantation. Prog. Retinal Eye Res. 36, 1–23. [DOI] [PubMed] [Google Scholar]
- Dorn J. D.; Ahuja A. K.; Caspi A.; da Cruz L.; Dagnelie G.; Sahel J. A.; Greenberg R. J.; McMahon M. J.; Group A. I. S. (2013) The Detection of Motion by Blind Subjects With the Epiretinal 60-Electrode (Argus II) Retinal Prosthesis. JAMA Ophthalmol. 131, 183–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rizzo S.; Beltin C.; Cinelli L.; Allegrini L.; Genovesi-Ebert F.; Barca F.; di Bartolo E. (2014) The Argus II Retinal Prosthesis: Twelve-Month Outcomes from a Single-Study Center. Am. J. Ophthalmol. 157, 1282–1290. [DOI] [PubMed] [Google Scholar]
- Perry V. H.; Cowey A. (1985) The ganglion cell and cone distributions in the monkey’s retina: Implications for central magnification factors. Vision Res. 25, 1795–1810. [DOI] [PubMed] [Google Scholar]
- Wässle H.; Grünert U.; Röhrenbeck J.; Boycott B. B. (1989) Cortical magnification factor and the ganglion cell density of the primate retina. Nature 341, 643–646. [DOI] [PubMed] [Google Scholar]
- Jepson L. H.; Hottowy P.; Mathieson K.; Gunning D. E.; Dąbrowski W.; Litke A. M.; Chichilnisky E. J. (2014) Spatially patterned electrical stimulation to enhance resolution of retinal prostheses. J. Neurosci. 34, 4871–4881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weitz A. C.; Behrend M. R.; Ahuja A. K.; Christopher P.; Wei J.; Wuyyuru V.; Patel U.; Greenberg R. J.; Humayun M. S.; Chow R. H.; Weiland J. D. (2014) Interphase gap as a means to reduce electrical stimulation thresholds for epiretinal prostheses. J. Neural Eng. 11, 016007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stingl K.; Bartz-Schmidt K. U.; Gekeler F.; Kusnyerik A.; Sachs H.; Zrenner E. (2013) Functional outcome in subretinal electronic implants depends on foveal eccentricity. Invest. Ophthalmol. Visual Sci. 54, 7658–7665. [DOI] [PubMed] [Google Scholar]
- Nishida K.; Kamei M.; Kondo M.; Sakaguchi H.; Suzuki M.; Fujikado T.; Tano Y. (2010) Efficacy of suprachoroidal-transretinal stimulation in a rabbit model of retinal degeneration. Invest. Ophthalmol. Visual Sci. 51, 2263–2268. [DOI] [PubMed] [Google Scholar]
- Nayagam D. A.; Williams R. A.; Allen P. J.; Shivdasani M. N.; Luu C. D.; Salinas-LaRosa C. M.; Finch S.; Ayton L. N.; Saunders A. L.; McPhedran M.; McGowan C.; Villalobos J.; Fallon J. B.; Wise A. K.; Yeoh J.; Xu J.; Feng H.; Millard R.; McWade M.; Thien P. C.; Williams C. E.; Shepherd R. K. (2014) Chronic electrical stimulation with a suprachoroidal retinal prosthesis: a preclinical safety and efficacy study. PLoS One 9, e97182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagel G.; Szellas T.; Huhn W.; Kateriya S.; Adeishvili N.; Berthold P.; Ollig D.; Hegemann P.; Bamberg E. (2003) Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. U.S.A. 100, 13940–13945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolbe M.; Besir H.; Essen L. O.; Oesterhelt D. (2000) Structure of the light-driven chloride pump halorhodopsin at 1.8 Å resolution. Science 288, 1390–1396. [DOI] [PubMed] [Google Scholar]
- Zhang F.; Wang L. P.; Brauner M.; Liewald J. F.; Kay K.; Watzke N.; Wood P. G.; Bamberg E.; Nagel G.; Gottschalk A.; Deisseroth K. (2007) Multimodal fast optical interrogation of neural circuitry. Nature 446, 633–639. [DOI] [PubMed] [Google Scholar]
- Dalkara D.; Byrne L. C.; Lee T.; Hoffmann N. V.; Schaffer D. V.; Flannery J. G. (2012) Enhanced gene delivery to the neonatal retina through systemic administration of tyrosine-mutated AAV9. Gene Ther. 19, 176–181. [DOI] [PubMed] [Google Scholar]
- Lin B.; Koizumi A.; Tanaka N.; Panda S.; Masland R. H. (2008) Restoration of visual function in retinal degeneration mice by ectopic expression of melanopsin. Proc. Natl. Acad. Sci. U.S.A. 105, 16009–16014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strettoi E.; Pignatelli V.; Rossi C.; Porciatti V.; Falsini B. (2003) Remodeling of second-order neurons in the retina of rd/rd mutant mice. Vision Res. 43, 867–877. [DOI] [PubMed] [Google Scholar]
- Strettoi E.; Porciatti V.; Falsini B.; Pignatelli V.; Rossi C. (2002) Morphological and functional abnormalities in the inner retina of the rd/rd mouse. J. Neurosci. 22, 5492–5504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- G N.; Tan A.; Farhatnia Y.; Rajadas J.; Hamblin M. R.; Khaw P. T.; Seifalian A. M. (2013) Channelrhodopsins: visual regeneration and neural activation by a light switch. Nat. Biotechnol. 30, 461–474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mourot A.; Kienzler M. A.; Banghart M. R.; Fehrentz T.; Huber F. M.; Stein M.; Kramer R. H.; Trauner D. (2011) Tuning photochromic ion channel blockers. ACS Chem. Neurosci. 2, 536–543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fortin D. L.; Banghart M. R.; Dunn T. W.; Borges K.; Wagenaar D. A.; Gaudry Q.; Karakossian M. H.; Otis T. S.; Kristan W. B.; Trauner D.; Kramer R. H. (2008) Photochemical control of endogenous ion channels and cellular excitability. Nat. Methods 5, 331–338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marc R. E.; Jones B. W. (2002) Molecular phenotyping of retinal ganglion cells. J. Neurosci. 22, 412–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marc R. E.; Liu W. (2000) Fundamental GABAergic amacrine cell circuitries in the retina: nested feedback, concatenated inhibition, and axosomatic synapses. J. Comp. Neurol. 425, 560–582. [DOI] [PubMed] [Google Scholar]
- Margolis D. J.; Gartland A. J.; Singer J. H.; Detwiler P. B. (2014) Network oscillations drive correlated spiking of ON and OFF ganglion cells in the rd1 mouse model of retinal degeneration. PLoS One 9, e86253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margolis D. J.; Newkirk G.; Euler T.; Detwiler P. B. (2008) Functional stability of retinal ganglion cells after degeneration-induced changes in synaptic input. J. Neurosci. 28, 6526–6536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stasheff S. F. (2008) Emergence of sustained spontaneous hyperactivity and temporary preservation of OFF responses in ganglion cells of the retinal degeneration (rd1) mouse. J. Neurophysiol. 99, 1408–1421. [DOI] [PubMed] [Google Scholar]
- Yee C. W.; Toychiev A. H.; Sagdullaev B. T. (2012) Network deficiency exacerbates impairment in a mouse model of retinal degeneration. Front Syst. Neurosci. 6, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He C.; Chen F.; Li B.; Hu Z. (2014) Neurophysiology of HCN channels: from cellular functions to multiple regulations. Prog. Neurobiol. 112, 1–23. [DOI] [PubMed] [Google Scholar]