

Abstract
Spatial capture-recapture (SCR) models have advanced our ability to estimate population density for wide ranging animals by explicitly incorporating individual movement. Though these models are more robust to various spatial sampling designs, few studies have empirically tested different large-scale trap configurations using SCR models. We investigated how extent of trap coverage and trap spacing affects precision and accuracy of SCR parameters, implementing models using the R package secr. We tested two trapping scenarios, one spatially extensive and one intensive, using black bear (Ursus americanus) DNA data from hair snare arrays in south-central Missouri, USA. We also examined the influence that adding a second, lower barbed-wire strand to snares had on quantity and spatial distribution of detections. We simulated trapping data to test bias in density estimates of each configuration under a range of density and detection parameter values. Field data showed that using multiple arrays with intensive snare coverage produced more detections of more individuals than extensive coverage. Consequently, density and detection parameters were more precise for the intensive design. Density was estimated as 1.7 bears per 100 km2 and was 5.5 times greater than that under extensive sampling. Abundance was 279 (95% CI = 193–406) bears in the 16,812 km2 study area. Excluding detections from the lower strand resulted in the loss of 35 detections, 14 unique bears, and the largest recorded movement between snares. All simulations showed low bias for density under both configurations. Results demonstrated that in low density populations with non-uniform distribution of population density, optimizing the tradeoff among snare spacing, coverage, and sample size is of critical importance to estimating parameters with high precision and accuracy. With limited resources, allocating available traps to multiple arrays with intensive trap spacing increased the amount of information needed to inform parameters with high precision.
Introduction
Knowledge of population size and spatial distribution is important for protection of threatened or endangered species [1]–[3], and management of harvested animal populations [4], [5]. Estimates of species’ abundance or density are useful as a baseline for developing protected areas [6], [7], prioritizing conservation actions [8], [9], and allocating harvest quotas [10]. However, large mammals often persist at low densities over large areas, are not uniformly distributed, and have large home ranges [11]–[13]. These characteristics may undermine abundance estimation and hinder subsequent conservation efforts [14]–[15].
Capture-recapture methods are often used to estimate density and abundance of rare or elusive carnivores [2], [15], [16]. Remote collection of DNA samples (e.g., hair, feces) enables researchers to sample wide geographic areas [17], [18], and has become almost universal for bear (Ursus spp.) capture-recapture studies [19], [20]. Nonetheless, trap configurations that do not adequately reflect population distributions and individual variation in space use may limit precise and accurate estimates of density and abundance [15], [21].
The spatial nature of sampling designs (e.g., trap distribution) and wildlife populations (e.g., home range distribution) are important components of estimating animal abundance [22]–[24]. Non-spatial capture-recapture models often require study designs to cover several times the area of an individual home range [25], while maintaining trap spacing narrow enough to ensure individuals have nonzero and homogenous capture probabilities [26], [27]. However, for species with large home ranges and individual movements, logistical constraints may require a tradeoff between extensive coverage of a study area with wide trap spacing or intensive coverage of a portion of the study area with close spacing [28], [29].
Spatial capture-recapture (SCR) models explicitly include animal movement and trap distribution, and therefore reduces constraints placed on sampling wide ranging species over large areas [30], [31]. Moreover, SCR defines a spatial point process model to estimate the home range (i.e., activity) centers of individuals detected, eliminating the need for ad hoc estimates of the effective sampling area [32]. Therefore, SCR models address a primary source of heterogeneity inherent in most carnivore populations by addressing unequal exposure to traps and edge effects [33], [34]. Simulations of SCR parameter estimates from black bear trapping configurations were unbiased when movement was at least half the distance between traps and when trap coverage was similar to the extent of movement [29], [35]. Although SCR models are robust to unequal trap exposure and appear flexible to various spatial trapping designs [34], few studies have empirically tested the efficacy of SCR models using different large-scale trap array configurations.
The large home ranges of bears and constraints to large-scale sampling often preclude adequate coverage of individual space use [36]. We tested a spatially extensive and intensive trapping scenario to compare how trap coverage and spacing affects precision of SCR parameter estimates using black bear (Ursus americanus) DNA encounter history data from hair snare arrays. To generalize the findings of our spatial sampling configurations and evaluate their accuracy, we also conducted simulations to measure bias under realistic densities and detection probabilities for large carnivores. We also consider effects of snare design on detections and provide insights towards implementing large scale capture-recapture sampling designs for SCR models for low density, wide ranging species.
Methods
Ethics Statement
All sampling methods complied with guidelines established by the American Society of Mammalogists and this study was approved by the Institutional Animal Care and Use Committee protocol (approval 10-037) at Mississippi State University. Sampling locations and procedures were approved by the Missouri Department of Conservation and sampling procedures did not involve endangered species.
Study area
We collected data from a recolonizing black bear population in the south-central Ozark Highlands ecological region of Missouri (36°30′–37°13′N, 91°16′–93°52′W), USA (Figure 1). The Ozark Highlands comprise 52% of the state’s total area and contains 57% forest, 32% crop and pasture, 2% grassland, and 7% developed areas [37]. About 80% of forest in Missouri occurs in the Ozark Highlands and is primarily upland oak-hickory (Quercus spp., Carya spp.) and oak-pine (Pinus spp.; [38]). Landownership in the Ozark Highlands includes private homesteads, farms, and public lands (e.g., Mark Twain National Forest, Ozark National Scenic Riverways). Elevation in Missouri ranges from 70 to 540 m with greatest elevations in the Ozark Highlands [39], [40].
Figure 1. Trap array configurations.

Location of the extensive and intensive configurations to estimate black bear density in south-central Missouri, USA. State space boundary for extensive (solid line) and intensive (dotted line) configurations represents the area used to estimate population size. For the extensive design, snares (black circles) were allocated proportionate to density of historical bear sightings. For the intensive design, five arrays were distributed in areas of expected bear occurrence and one snare was placed in each cell; specific locations omitted for clarity. The five arrays were designated alphabetically (A–E) from west to east.
Data collection
We collected black bear hair samples using barbed-wire hair snares [41]. Snares were constructed using 4-barbed, 15.5 gauge wire to create an enclosure (∼1.5 m radius) around three or more trees. For the extensive design, we constructed snares using a single strand of barbed-wire set 50 cm above ground. For the intensive design, we constructed snares using two strands of barbed-wire with strands 20 and 50 cm above ground. We applied raspberry oil (intensive design only; Mother Murphy’s Laboratories, Inc., Greensboro, NC), anise oil (Minnesota Snareline Products, Pennock, MN), and Ultimate Bear Lure (Wildlife Research Center, Ramsey, MN) on perimeter trees forming the enclosure, about 2 m above ground. We placed decaying logs in the center of the enclosure and saturated them with 0.5 L of fish oil as an attractant [19]. Hair snare stations were re-lured every 10 days at the beginning of each consecutive sampling session. We collected hair samples at the end of each session and considered all hair found on a barb or single tree one sample. We placed hair in separate coin envelopes, and air dried samples before processing.
Extensive sampling design
We derived the sampling boundary from a 70% fixed kernel isopleth applied to historical bear sightings (1989–2010) reported to the Missouri Department of Conservation by citizens throughout the state [42]. This area comprised 13,508 km2 in south-central Missouri. We used the 70% isopleth because this was the maximum logistically feasible extent of sampling and contained the core area of interest by state biologists. We used an array with 10×10 km cells to generate a distribution of bear sightings per array cell, excluding cells with zero bear sightings. We allocated 378 snares proportionate to the number of sightings per cell (Figure 1) following methods similar to Dreher et al. [18]. Cells containing 1–3 bear sightings received one snare, cells containing 4–5 sightings received four snares, cells with 6–7 sightings received five snares, and cells with >8 sightings received six snares. Cells with suitable habitat (i.e., forest) containing zero sightings adjacent to cells with similar habitat containing bear sightings were allocated snares similar to adjacent cells. We selected hair snare locations based on habitat characteristics and availability of forested private and public land. We used ArcMap 9.3.1 (Environmental Systems Research Institute, Redlands, California, USA) to select approximate locations for hair snares using forest cover data (30 m resolution, [43]) as initial criteria to maximize bear detection; excluding open water, agricultural, and developed areas.
We selected final snare locations within about 300 m of initial locations and out of sight from human trails or dwellings. Additionally, we used previous bear sightings, recent bear activity, and habitat and topographic features to select hair snare locations to maximize black bear capture [18]. We attempted to maintain a minimum distance of 3 km between snare sites, and conducted oversampling of snare locations in the event existing land use or ownership precluded snare placement. We monitored snares over six, 10-day sessions during June–August 2011.
Intensive sampling design
We established 403 hair snares in five, 9×9 sampling arrays (A–E) with 2.6 km2 cells (210 km2/array) in south-central Missouri ( = 81 snares/array, SD = 1; Figure 1). We selected array locations to maximize coverage of the largest available forested regions known or expected to contain bears based on information from our extensive sampling effort and prior radio-collaring efforts [28]. Four arrays (A–D) were contained within the previous extensive survey area and array E was about 15 km east of the extensive survey boundary (Figure 1). We allocated one hair snare to each cell and monitored all snares over six, 10-day sessions during June–August 2012. Snares were set in forests on both private and public lands. We selected preliminary and final snare sites following the same criteria as the extensive sampling protocol.
= 81 snares/array, SD = 1; Figure 1). We selected array locations to maximize coverage of the largest available forested regions known or expected to contain bears based on information from our extensive sampling effort and prior radio-collaring efforts [28]. Four arrays (A–D) were contained within the previous extensive survey area and array E was about 15 km east of the extensive survey boundary (Figure 1). We allocated one hair snare to each cell and monitored all snares over six, 10-day sessions during June–August 2012. Snares were set in forests on both private and public lands. We selected preliminary and final snare sites following the same criteria as the extensive sampling protocol.
DNA extraction and microsatellite genotyping
We cut follicles from the shafts of hair to minimize the amount of melanin, a known PCR inhibitor [44], within each DNA extract. Number of follicles per sample ranged from one to twelve depending on amount of hair removed from each barb. We placed follicles in 1.5 mL Eppendorf tubes before adding 250 µL InstaGene matrix (BioRad, Hercules, CA). Samples incubated overnight at 56°C, then at 100°C for 15 min [45]. Following centrifugation at 13 krpm for 3 min, we transferred supernatant to a clean 1.5 mL tube before use in downstream reactions.
We genotyped samples at 15 microsatellite loci (G1A, G10B, G10C, G1D, G10J, G10L, G10M, G10O, G10P, G10U, UarMU05, UarMU10, UarMU23, UarMU59, and P2H03; [46]–[48]) following the protocol of Puckett et al. [49], where UarMU05 and UarMU10 were added to the multiplex panel in 2012 (Dataset S1). We randomly selected 25 samples genotyped at all loci to calculate the probability of identity between siblings (PIDsib; α = 0.001) in Gimlet [50]. When determining recaptures we allowed two mismatches between samples. We determined the sex of unique individuals by amplification of the Amelogenin gene followed by BslI digestion [51].
Population analysis
We used DNA-based encounter history data from hair snares and SCR models to estimate black bear density in south-central Missouri. We analyzed data using package secr (version 2.7.0; [52]) in program R [53]. We fit each model using a binomial observation model with the half-normal detection function, where the parameter g0 is the probability of detection at the activity center of an individual and σ is the spatial scale parameter of the detection function [54]. The spatial scale parameter describes the rate of decrease in capture probability as a function of increasing distance from a trap and an individual’s activity center [32]. We compared 12 a priori models for each array configuration (Table 1). We fit a null model with no covariates and 11 models with varying effects on the detection parameters (g0, σ). We created models based on expected sources of variation in black bear detection probability within our study area. Models with effects on g0 included one model with time as a factor (t), and three models with behavioral responses following initial detection (global learned response (b), snare-specific learned response (bk), and a snare-specific Markovian response (Bk)). We also fit seven models using sex as a categorical individual covariate to specify sex-specific effects on g0 and σ and in combination with the behavioral response models.
Table 1. Model selection results for fitted models ranked by AICc with number of parameters (K), log likelihood (LL), and AICc weights (wi) to estimate black bear density in south-central Missouri, USA, for extensive and intensive sampling designs.
| Design | Model | K | LL | AICc | ΔAICc | wi |
| Extensive | g0(bk), σ(.) | 4 | −254.5 | 519.0 | 0.0 | 0.5 |
| g0(bk), σ(sex) | 5 | −253.6 | 520.3 | 1.4 | 0.3 | |
| g0(Bk), σ(.) | 4 | −255.9 | 521.8 | 2.8 | 0.1 | |
| g0(Bk), σ(sex) | 5 | −254.9 | 523.0 | 4.0 | 0.1 | |
| g0(sex), σ(sex) | 5 | −266.6 | 546.4 | 27.4 | 0.0 | |
| g0(.), σ(.) | 3 | −274.0 | 555.1 | 36.1 | 0.0 | |
| g0(.), σ(sex) | 4 | −272.7 | 555.4 | 36.4 | 0.0 | |
| g0(b), σ(.) | 4 | −273.2 | 556.4 | 37.4 | 0.0 | |
| g0(b), σ(sex) | 5 | −271.8 | 556.7 | 37.8 | 0.0 | |
| g0(sex), σ(.) | 4 | −273.7 | 557.3 | 38.3 | 0.0 | |
| g0(t), σ(.) | 8 | −273.6 | 572.1 | 53.1 | 0.0 | |
| g0(t), σ(sex) | 9 | −272.3 | 574.6 | 55.6 | 0.0 | |
| Intensive | g0(bk), σ(.) | 4 | −1175.8 | 2360.0 | 0.0 | 0.8 |
| g0(bk), σ(sex) | 5 | −1175.8 | 2362.2 | 2.2 | 0.2 | |
| g0(Bk), σ(.) | 4 | −1215.7 | 2439.9 | 79.9 | 0.0 | |
| g0(Bk), σ(sex) | 5 | −1215.2 | 2441.0 | 81.0 | 0.0 | |
| g0(b), σ(.) | 4 | −1246.6 | 2501.8 | 141.8 | 0.0 | |
| g0(sex), σ(.) | 4 | −1246.9 | 2502.3 | 142.3 | 0.0 | |
| g0(.), σ(.) | 3 | −1249.0 | 2504.3 | 144.3 | 0.0 | |
| g0(t), σ(.) | 8 | −1245.0 | 2507.7 | 147.7 | 0.0 | |
| g0(sex), σ(sex) | 5 | −1299.5 | 2609.6 | 249.7 | 0.0 | |
| g0(.), σ(sex) | 4 | −1301.0 | 2610.6 | 250.6 | 0.0 | |
| g0(b), σ(sex) | 5 | −1300.9 | 2612.5 | 252.5 | 0.0 | |
| g0(t), σ(sex) | 9 | −1296.7 | 2613.6 | 253.6 | 0.0 |
We fitted models using the half-normal detection function with baseline capture probability (g0) and scale parameter (σ). Effects on g0 and σ included time as a factor (t), global learned response (b), snare-specific learned response (bk), and a snare-specific Markovian response (Bk), and sex. Parameters with “.” indicate no effect.
We defined the state space (i.e., area of integration) as the area encompassing snares and all individuals potentially exposed to capture [30]. This area defines the extent of the distribution of home range centers in the population. We used three times the estimated σ to calculate the state space radius around snares [32] and tested if this was large enough using the mask check function in package secr. The state space radius was 45 km for the extensive design and 30 km for the intensive design, resulting in 41,121 km2 and 16,812 km2 areas, respectively. To estimate population size, we used the expected population size (E(N)) derived from the top supported model [34]. For the extensive design, we estimated population size using a 30 km radius (29,898 km2), as this represented our area of interest. We compared precision of parameter estimates using coefficients of variation (CV). We selected the top supported model for each configuration using Akaike’s Information Criterion corrected for small samples (AICc) and considered models competing if within 2 AICc units from the top supported model [55].
Simulations
We used our field sampling designs to simulate spatial capture-recapture datasets to evaluate accuracy of density estimates under each sampling configuration. We chose density and detection parameter values to represent both the observed values in our study and values commonly observed in other black bear studies in the United States [12], [56]–[58]. For density, we used values of 1.0 and 2.5 individuals per 100 km2, and capture probability (g0) values of 0.1 and 0.2. We also tested the scale parameter (σ) at 5, 10, and 15 km to investigate the effect of varying σ on density between sampling designs. We used a state space radius of 30 km when σ was 5 and 10 km and a radius of 45 km when σ was 15 km, with default point spacing of 64×64 points. Number of sampling intervals was set at six sessions. We then ran 100 replicates for each combination of density and detection parameter values (n = 12) under both configurations. For each scenario, we fit SCR models using the half-normal detection function in program DENSITY v5.0 [59]. To compare accuracy of density estimates to the true values from each sampling configuration we assessed average percent relative bias (%RB) and proportional coverage of confidence intervals (%COV).
Results
Microsatellite genotyping
To reduce genotyping error, we genotyped each hair sample three times before calling a consensus genotype [49] and we calculated an allelic dropout of 2.4% in Gimlet v1.3.3 [50] on 20 randomly selected samples. The probability of identity between siblings was 7.18×10−4; this level of PIDsib required samples to be genotyped at eight loci for inclusion in the study and required a genotyping rate of 61.5% for the extensive design and 53.3% for the intensive design to be included in the sample. Following removal of samples that genotyped at fewer than eight loci, in the extensive design there were 42 unique hair samples with an average genotyping rate of 93.8% across loci. In the intensive design, 224 unique hair samples were genotyped with an 83.8% genotyping rate.
Extensive sampling design
We collected 98 black bear hair samples suitable for DNA extraction over six sessions from 30 unique snares (8% of the total number of snares). Number of black bear hair samples declined over time ( = 16.3, 28 samples in session one to eight samples in session six). Mean distance of each snare to nearest neighbor was 3.6 km (SD = 0.04 km). Mean distance between consecutive detection locations was 9.6 km.
= 16.3, 28 samples in session one to eight samples in session six). Mean distance of each snare to nearest neighbor was 3.6 km (SD = 0.04 km). Mean distance between consecutive detection locations was 9.6 km.
Microsatellite marker analysis of hair samples revealed 25 unique individuals (11 F, 14 M) detected at 7% of all snares monitored (Dataset S2). Total detections per session ranged from six to nine (SD = 1.1), with 42 total detections, including within-session recaptures (Table 2). Individuals were detected on average 1.7 times (range = 1–5, SD = 1.0). Females were detected on average 1.9 times (range = 1–5, SD = 1.3) and males 1.5 times (range = 1–3, SD = 0.7). Fourteen individuals were not recaptured, including 55% of females and 57% of males. We detected individuals at an average of 1 snare (SD = 0.5, max = 2). Snares with ≥1 detection per session remained about constant (range = 6–8) across sampling sessions.
Table 2. Summary of sampling statistics for extensive and intensive (arrays A–E) black bear survey configurations in south-central Missouri, USA.
| Design | Array | Snaresa | ub | nc | Detectionsd | Snares Visitede | No. Hair Samples |
| Extensive | 378 | 4.2 (2.4, 25) | 6.5 (1.4, 39) | 7.0 (1.1, 42) | 6.8 (0.8, 26) | 16.3 (7.2, 98) | |
| Intensive | A | 81 | 0.7 (0.8, 4) | 0.8 (0.8, 5) | 1.0 (0.9, 6) | 1.0 (0.9, 4) | 2.2 (2.3, 13) |
| B | 79 | 8.0 (6.7, 48) | 14.7 (5.1, 88) | 18.5 (7.0, 111) | 12.8 (3.6, 36) | 39.5 (18.4, 237) | |
| C | 81 | 1.2 (0.8, 7) | 1.8 (0.8, 11) | 2.8 (1.2, 17) | 2.7 (1.2, 10) | 7.3 (5.7, 44) | |
| D | 81 | 3.7 (2.9, 22) | 8.0 (2.3, 48) | 11.8 (2.7, 71) | 9.2 (1.8, 29) | 32.8 (12.0, 197) | |
| E | 81 | 1.8 (1.8, 11) | 2.7 (2.0, 16) | 3.2 (2.2, 19) | 2.5 (1.9, 12) | 6.2 (4.4, 37) | |
| Intensive Total | 403 | 3.1 (2.6, 92) | 5.6 (2.2, 168) | 7.5 (2.8, 224) | 5.6 (1.9, 91) | 17.6 (8.6, 528) |
Order of values are mean (standard deviation, total) over six sessions. Note the sum of new detections (u) was 92 total individuals for the intensive design due to two individuals being detected in two arrays (i.e., total individuals was actually 90).
Number of lured snares in each session.
Number of individuals detected for the first time on each session.
Number of individuals detected on each session.
Number of detections, including within-session recaptures.
Number of snares having at least one detection per session.
The snare-specific learned response model (bk) was most supported and 1.7 times more supported than the single competing model (g0[bk], σ[sex]; Table 1). However, we used parameter estimates from the top model as results between competing models were similar. Capture probability (g0) under the top model increased following initial detection at the same snare (Table 3). The estimated scale parameter of the detection function (σ) was 14.8 km (95% CI = 10.2–21.6 km). Density was 0.3 bears per 100 km2 (95% CI = 0.2–0.6). Expected population size (E(N)) was 91 individuals (SE = 31, 95% CI = 47–175) in our 29,898 km2 area of interest. Coefficients of variation (CV) ranged from 19% to 43% for all parameters. Activity centers estimated from the model were located primarily in the north-central and eastern portions of the trap array (Figure 2).
Table 3. Real parameter estimates and their precision (CV) for the most supported models to estimate black bear density (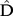 ; bears per 100 km2) for extensive and intensive array configurations in south-central Missouri, USA.
; bears per 100 km2) for extensive and intensive array configurations in south-central Missouri, USA.
| Density | g0 | σ | ||||||||
| Design |

|
SE | 95% CI | CV |

|
SE | CV |

|
SE | CV |
| Extensive | 0.3 | 0.1 | 0.2–0.6 | 34 | bk0: 0.003 | 0.001 | 43 | 14.8 | 2.9 | 19 |
| bk1: 0.168 | 0.059 | 35 | ||||||||
| Intensive | 1.7 | 0.3 | 1.1–2.4 | 19 | bk0: 0.011 | 0.002 | 15 | 8.5 | 0.9 | 10 |
| bk1: 0.166 | 0.025 | 15 | ||||||||
Capture probability ( ) given for initial capture (bk0) and for previously captured individuals (bk1). Scale parameter of the detection function (
) given for initial capture (bk0) and for previously captured individuals (bk1). Scale parameter of the detection function ( ) reported in kilometers.
) reported in kilometers.
Figure 2. Estimated black bear activity centers.

Location of hair snares and estimated activity centers (i.e., home range center) of identified bears with the extensive and intensive configurations in south-central Missouri, USA.
Intensive sampling design
We collected 528 black bear hair samples suitable for DNA extraction over six sessions from 110 unique snares (27% of total). Number of black bear hair samples collected in each array per session was variable (mean range = 2.2–39.5, grand mean = 17.6) and the total number of black bear hair samples collected declined over time (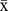 = 88.0; 107 samples in session one to 53 in session six). Mean distance of each snare to nearest neighbor within arrays was 1.0 km (SD = 0.04 km). Mean distance between consecutive detection locations was 2.5 km.
= 88.0; 107 samples in session one to 53 in session six). Mean distance of each snare to nearest neighbor within arrays was 1.0 km (SD = 0.04 km). Mean distance between consecutive detection locations was 2.5 km.
Microsatellite marker analysis of hair samples from snares revealed 90 unique individuals (59 F, 31 M; 4–48 individuals/array) detected at 23% of all snares monitored (Table 2; Dataset S2). Total detections per session ranged from 25 to 43 (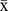 = 37.3, SD = 7.5), with 224 total detections, including within-session recaptures. Individuals were detected on average 2.5 times (range = 1–10, SD = 2.5). Females were detected on average 2.6 times (range = 1–10, SD = 2.7) and males 2.3 times (range = 1–10, SD = 2.2). Fifty-five individuals were not recaptured, including 63% of females and 61% of males. Two individuals were detected in two arrays, a male with an inter-trap distance of 57 km and a female with an inter-trap distance of 31 km. We detected individuals at an average of 2.0 snares (SD = 1.6, max = 8), and number of snares having ≥1 detection varied among arrays (range = 4–36; Table 2).
= 37.3, SD = 7.5), with 224 total detections, including within-session recaptures. Individuals were detected on average 2.5 times (range = 1–10, SD = 2.5). Females were detected on average 2.6 times (range = 1–10, SD = 2.7) and males 2.3 times (range = 1–10, SD = 2.2). Fifty-five individuals were not recaptured, including 63% of females and 61% of males. Two individuals were detected in two arrays, a male with an inter-trap distance of 57 km and a female with an inter-trap distance of 31 km. We detected individuals at an average of 2.0 snares (SD = 1.6, max = 8), and number of snares having ≥1 detection varied among arrays (range = 4–36; Table 2).
The snare-specific learned response model (bk) was most supported and four times more supported than the next best model (g0[bk], σ[sex]; Table 1). Capture probability under the top model increased following initial detection at the same trap, and σ was 8.5 km (95% CI = 7.0–10.4 km; Table 3). Density of the pooled arrays was 1.7 bears per 100 km2 (95% CI = 1.1–2.4). Expected population size (E(N)) was 279 individuals (SE = 54, 95% CI = 193–406) in the 16,812 km2 state space. Coefficients of variation ranged from 10% to 19% for all parameters. Estimated activity centers were primarily clustered within and around arrays B and D (Figure 2).
Since we only added a lower strand of barbed wire to hair snares in the intensive configuration, we also describe detections after excluding hair samples captured on the lower strand to compare sample size and distribution of detections to the full data set. Of the 224 total detections, 35 (16%) resulted from hair samples collected from the bottom strand and represented 30 individuals (23 females, 7 males). Exclusion of these detections resulted in the loss of 14 unique bears (10 females, 4 males) from the data set. Bottom strand detections also resulted in the loss of 11 observed movements (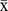 = 7.7 km, SD = 16.3) between successive recaptures at different snares, including the largest distance (57 km) moved between arrays (B and D) by a male bear, which was also the largest movement in the full data set.
= 7.7 km, SD = 16.3) between successive recaptures at different snares, including the largest distance (57 km) moved between arrays (B and D) by a male bear, which was also the largest movement in the full data set.
Simulations
For our low density simulations (1.0 bear per 100 km2) with σ<15 km, both array configurations had relative bias less than ±2.0% and 95% confidence interval coverage ≥92% (Table 4). Similarly, at 2.5 bears per 100 km2 both configurations produced almost unbiased density estimates at both g0 levels and when σ<15 km. At σ = 15 km, relative bias for the extensive design remained similar to other scenarios, but showed a positive increase for the intensive design. Both configurations performed well when simulation scenarios reflected field results of each configuration.
Table 4. Percent relative bias (%RB) and percent coverage of 95% confidence intervals (%COV) of mean density estimates 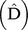 for simulations of spatial capture recapture models under extensive and intensive trap configurations.
for simulations of spatial capture recapture models under extensive and intensive trap configurations.
| Array Configuration | ||||||||||
| Scenario | Extensive | Intensive | ||||||||
| D | g0 | σ |

|
SE | %RB | %COV |

|
SE | %RB | %COV |
| 1.0 | 0.1 | 5 | 1.01 | 0.09 | 1.21 | 92 | 1.00 | 0.16 | −0.01 | 93 |
| 10 | 0.99 | 0.07 | −0.72 | 96 | 0.99 | 0.09 | −0.84 | 94 | ||
| 15 | 1.00 | 0.13 | 0.07 | 93 | 1.01 | 0.07 | 0.66 | 97 | ||
| 0.2 | 5 | 1.00 | 0.08 | −0.08 | 97 | 1.00 | 0.14 | −0.28 | 94 | |
| 10 | 1.00 | 0.07 | −0.18 | 97 | 1.02 | 0.09 | 1.94 | 96 | ||
| 15 | 1.00 | 0.13 | −0.26 | 96 | 1.03 | 0.07 | 3.43 | 93 | ||
| 2.5 | 0.1 | 5 | 2.47 | 0.01 | −1.01 | 95 | 2.50 | 0.03 | −0.01 | 90 |
| 10 | 2.49 | 0.01 | −0.45 | 95 | 2.51 | 0.02 | 0.28 | 91 | ||
| 15 | 2.49 | 0.09 | −0.22 | 98 | 2.53 | 0.11 | 1.36 | 96 | ||
| 0.2 | 5 | 2.51 | 0.13 | 0.58 | 92 | 2.49 | 0.02 | −0.52 | 97 | |
| 10 | 2.50 | 0.01 | −0.01 | 92 | 2.53 | 0.01 | 1.22 | 92 | ||
| 15 | 2.51 | 0.12 | 0.59 | 96 | 2.57 | 0.11 | 2.62 | 91 | ||
Estimates are based on averages over 100 replicates for each scenario of density (1.0, 2.5 bears per100 km2), g0 (0.1, 0.2), and σ (5, 10, 15 km).
Discussion
We found that multiple arrays spaced across a landscape using intensive snare coverage yielded more captures and recaptures of more individuals than extensive coverage spaced over an area approximately 13 times larger. Consequently, estimated density using the intensive configuration was 5.5 times greater than that under the extensive configuration. By pooling detections among our arrays with closer snare spacing and using SCR models to explicitly account for variable exposure to traps, we were able to increase precision while retaining the ability to estimate average density over a landscape [60]–[62]. However, placement of intensive arrays was informed largely by the distribution of detections from the extensive sampling effort. Therefore, although results support the intensive design, prior knowledge of bear distribution was critical to increasing detections in the intensive configuration. When population distribution and space use are poorly understood, adjusting sampling design over multiple surveys may be required [28].
With intensive sampling, CV of parameter estimates decreased on average by 53% compared to extensive sampling. Pollock et al. [63] recommended a CV<20% for reasonable precision of estimates, which we achieved for all parameters with our intensive configuration. Boulanger et al. [28] also demonstrated increased capture probability and precision under intensive sampling of a grizzly bear (Ursus arctos) population. However, over 50% of individuals in our study were not recaptured under both sampling designs and capture probability remained below recommended levels (i.e., >0.2; [28]). The lack of food reward at snares, summer migration to find food, mating opportunities, or dispersal [64], [65], may partially explain low recaptures observed during our summer (June–August) surveys. We suggest greater precision under the intensive design was largely due to detecting a greater proportion of individuals and increased detections at multiple snares [28], [29]. Though we cannot discern potential demographic changes between years, it is important to consider potential year effects on parameter estimates. For example, seasonal food abundance can affect movements and responses to baited sites [64], [66]. However, model selection results suggest bears responded to lured snares similarly between years. Therefore, we suggest our comparisons are appropriate given the constraints of implementing such large scale capture-recapture studies.
Although our two sampling designs are not comparable experimentally, results demonstrate the interplay among spatial sampling design, population distribution, and precision of detection parameters [35], [60]. The extensive configuration covered a wide geographic area, but snare distribution either covered large areas of unoccupied habitat or snare spacing was too wide given individual movements [29]. Low precision of parameter estimates with extensive sampling may illustrate the reality of simulations by Tobler and Powell [27], where precision decreased as trap spacing increased with larger array size. The logistical constraints of implementing such a large survey and risk of obtaining insufficient detections makes this approach unattractive for low density populations, especially those unevenly distributed over the landscape. This sampling design has proved effective in populations with higher bear density and larger home range size when population estimation was combined with independent data from hunter harvests [18]. In less studied and non-harvested populations, such as in Missouri, auxiliary information is often unavailable or too cursory to accurately inform study design or analyses.
Detections over the extensive and intensive arrays were not uniformly distributed, with most detections concentrated in two distinct areas during both years (Figure 2). If the low detection areas of the extensive design resulted from insufficient sampling alone, we would expect detections to increase in these areas when using intensive sampling [67], assuming minimal demographic changes between years. Although overall detections were greater using the intensive design, two of the four arrays that overlapped the extensive design area still received low detections. Though ancillary, this spatial pattern of detections during both years suggests a low, heterogeneous density as opposed to insufficient sampling design. Moreover, Karanth et al. [68] demonstrated a positive relationship between spatial coverage of traps and total animals detected. With extensive sampling, we detected 25 bears over a nominal array area of about 13,500 km2 and with intensive sampling we detected 90 bears over about 1,000 km2. That our results were not consistent with findings by Karanth et al. [68] further suggests a population where most individuals occurred in clustered regions with few bears interspersed between these areas. Heterogeneous densities are common among large carnivore populations in a varied landscape [58], [62], particularly among recently recolonizing populations [12], [49], [69].
Changes to the intensive design, including the addition of a lower strand of barbed wire, increased the number of unique individuals and overall detections. Excluding lower strand detections greatly affected the number and spatial distribution of detections. One of the male detection losses represented the largest detected movement (57 km) between snares. Sex-specific space use can bias detection in carnivore population surveys [15], and our results illustrate the potential importance of spatial sampling design and snare design to increasing overall detections and sex-specific movements among snares. Some studies have attempted to quantify the effectiveness of using a second, lower strand of barbed wire to increase capture probability or identify family groups [18], [28]. Whereas Boulanger et al. [70] found that a lower strand did not greatly affect estimates for a grizzly bear population, we contend that for low density populations a second strand may sufficiently increase data on encountered individuals and movements.
Simulations of our extensive and intensive configurations showed low bias and adequate confidence interval coverage for all scenarios. Although bias was low, the positive bias for the intensive design when σ = 15 km suggests increasing distance between snares to extend spatial coverage may increase the likelihood of detecting large movements [35]. Increasing spacing in the intensive design likely wouldn’t affect precision of σ as our effective trap spacing (i.e., spacing/σ; [29]) was much narrower than the <2σ suggested by Sun et al. [29]. Whereas both designs had low bias in simulations, few detections and inter-trap recaptures precluded precise density estimation for the extensive design field study, though precision of σ remained adequate (i.e., CV<20%). These results show that although SCR models are robust to variable spatial sampling designs [29], [35], in low density populations, or populations with non-uniform space use, optimizing the tradeoff between snare spacing, coverage, and sample size is critical for estimating σ and density with high precision and accuracy.
Management decisions for large mammals are typically made over large spatial scales [18], [71], [72], and inferences informing these decisions should cover a similar area [73]. However, logistical constraints and carnivore ecology often preclude large scale inference [74], [75]. For example, although SCR may be robust to our extensive sampling design, low detections still hindered precise density estimation over such a large region [35], [63]. Moreover, increasing trap intensity over a smaller region poses limits to the extent of density extrapolation to a larger area [74]. Thus, our extrapolation of density to a population estimate of 279 (95% CI = 193–406) black bears in a 16,812 km2 area must be treated with caution [74]. This estimate also cannot be compared to that from extensive sampling as they were derived from different areas. However, given knowledge of bear presence and movements within this region (J. Beringer, unpublished data), the location of our intensive arrays likely sampled a representative range of bear densities and focused on areas with known populations [60].
Conclusion
Our study highlights important considerations in sampling design for attaining precise estimates using SCR models for wide-ranging mammals. Although SCR models are flexible to various spatial designs [34], they remain sensitive to the number of detections and inter-trap recaptures across the range of individual movements [35]. Complete spatial coverage with sufficient trap spacing is challenging when animals with large home ranges exist at low densities and are not uniformly distributed [35], [62], [76]. Given these conditions, multiple arrays with intensive trap spacing similar in extent to individual movements should increase precision of detection parameters. We demonstrated support for sampling recommendations from simulated SCR analyses of black bear data sets [29], [35] and illustrated realistic challenges of tailoring large scale spatial trap designs to a species’ behavior and spatial ecology. Although our study was specific to black bears, we suggest our findings are applicable to other wide ranging and low density species. The flexibility of SCR models to various sampling designs and techniques provide increased opportunities to accurately survey rare and elusive animals of high management or conservation priority.
Supporting Information
Microsatellite genotypes of individual bears sampled at hair snares.
(XLSX)
DNA encounter histories from extensive and intensive design field seasons.
(XLSX)
Acknowledgments
We thank J. Barnett, J. Blair, M. Cancellare, I. Davis, D. Dobesh, D. Geurts, R. Langendorf, C. Lockerby, S. Lynch, T. Moody, M. Salyer, K. Wiskirchen, and many Missouri Department of Conservation staff for field assistance and B.K. Strickland for helpful comments on earlier drafts.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
Funding was provided by the Missouri Department of Conservation; U.S. Fish and Wildlife Service Federal Aid in Wildlife Restoration; Department of Wildlife, Fisheries, and Aquaculture and Forest and Wildlife Research Center at Mississippi State University; and Safari Club International Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Eggert LS, Eggert JA, Woodruff DS (2003) Estimating population sizes for elusive animals: the forest elephants of Kakum National Park, Ghana. Mol Ecol 12: 1389–1402. [DOI] [PubMed] [Google Scholar]
- 2. Kendall KC, Stetz JB, Boulanger J, Macleod AC, Paetkau D, et al. (2009) Demography and genetic structure of a recovering grizzly bear population. J Wildl Manage 73: 3–17. [Google Scholar]
- 3. Gray TNE, Prum S (2012) Leopard density in post-conflict landscape, Cambodia: evidence from spatially explicit capture–recapture. J Wildl Manage 76: 163–169. [Google Scholar]
- 4.Williams BK, Nichols JD, Conroy MJ (2002) Analysis and management of animal populations: modeling, estimation, and decision making. San Diego, USA and London, UK: Academic Press. 817 p. [Google Scholar]
- 5. Garshelis DL, Hristienko H (2006) State and provincial estimates of American black bear numbers versus assessments of population trend. Ursus 17: 1–7. [Google Scholar]
- 6.Gross M, Simmons N, Mahony M (2008) Wildlife Conservation Society Annual Report 2008. Behler DA, editor. Bronx, New York, USA. [Google Scholar]
- 7. Stokes EJ, Strindberg S, Bakabana PC, Elkan PW, Iyenguet FC, et al. (2010) Monitoring great ape and elephant abundance at large spatial scales: measuring effectiveness of a conservation landscape. PLoS ONE 5(4): e10294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Master LL (1991) Threats and setting priorities for conservation. Conserv Biol 5: 559–563. [Google Scholar]
- 9. Neel MC, Leidner AK, Haines A, Goble DD, Scott JM (2012) By the numbers: how is recovery defined by the US Endangered Species Act? BioScience 62: 646–657. [Google Scholar]
- 10. Peacock E, Titus K, Garshelis DL, Peacock MM, Kuc M (2011) Mark–recapture using tetracycline and genetics reveal record-high bear density. J Wildl Manage 75: 1513–1520. [Google Scholar]
- 11. Damuth J (1981) Population density and body size in mammals. Nature 290: 699–700. [Google Scholar]
- 12. Frary VJ, Duchamp J, Maehr DS, Larkin JL (2011) Density and distribution of a colonizing front of the American black bear Ursus americanus . Wildlife Biol 17: 404–416. [Google Scholar]
- 13. Sunarto, Kelly MJ, Klenzendorf S, Vaughan MR, Zulfahmi, et al. (2013) Threatened predator on the equator: multi-point abundance estimates of the tiger Panthera tigris in central Sumatra. Oryx 47: 211–220. [Google Scholar]
- 14. Ebert C, Knauer F, Storch I, Hohmann U (2010) Individual heterogeneity as a pitfall in population estimates based on non-invasive genetic sampling: a review and recommendations. Wildlife Biol 16: 225–240. [Google Scholar]
- 15. Sollmann R, Furtado MM, Gardner B, Hofer H, Jácomo AT, et al. (2011) Improving density estimates for elusive carnivores: accounting for sex-specific detection and movements using spatial capture–recapture models for jaguars in central Brazil. Biol Conserv 144: 1017–1024. [Google Scholar]
- 16. O'Brien TG, Kinnaird MF (2011) Density estimation of sympatric carnivores using spatially explicit capture-recapture methods and standard trapping grid. Ecol Appl 21: 2908–2916. [Google Scholar]
- 17. Mowat G, Strobeck C (2000) Estimating population size of grizzly bears using hair capture, DNA profiling, and mark-recapture analysis. J Wildl Manage 64: 183–193. [Google Scholar]
- 18. Dreher BP, Winterstein SR, Scribner KT, Lukacs PM, Etter DR, et al. (2007) Noninvasive estimation of black bear abundance incorporating genotyping errors and harvested bear. J Wildl Manage 71: 2684–2693. [Google Scholar]
- 19. Belant JL, Van Stappen JF, Paetkau D (2005) American black bear population size and genetic diversity at Apostle Islands National Lakeshore. Ursus 16: 85–92. [Google Scholar]
- 20. Garshelis DL (2006) On the allure of noninvasive genetic sampling – putting a face to the name. Ursus 17: 109–123. [Google Scholar]
- 21. Boulanger J, McLellan BN (2001) Closure violation in DNA-based mark-recapture estimation of grizzly bear populations. Can J Zool 79: 642–651. [Google Scholar]
- 22. Apps CD, McLellan BN, Woods JG, Proctor MF (2004) Estimating grizzly bear distribution and abundance relative to habitat and human influence. J Wildl Manage 68: 138–152. [Google Scholar]
- 23. Wegge P, Pokheral CP, Jnawali SR (2004) Effects of trapping effort and trap shyness on estimates of tiger abundance from camera trap studies. Anim Conserv 7: 251–256 10.1017/S1367943004001441 [DOI] [Google Scholar]
- 24. Royle JA, Young KV (2008) A hierarchical model for spatial capture-recapture data. Ecology 89: 2281–2289. [DOI] [PubMed] [Google Scholar]
- 25. Bondrup-Nielsen S (1983) Density estimation as a function of live-trapping grid and home range size. Can J Zool 61: 2361–2365. [Google Scholar]
- 26. Dillon A, Kelly MJ (2007) Ocelot Leopardus pardalis in Belize: the impact of trap spacing and distance moved on density estimates. Oryx 41: 469–477. [Google Scholar]
- 27. Tobler MW, Powell GVN (2013) Estimating jaguar densities with camera traps: problems with current designs and recommendations for future studies. Biol Conserv 159: 109–118. [Google Scholar]
- 28. Boulanger J, McLellan BN, Woods JG, Proctor MF, Strobeck C (2004) Sampling design and bias in DNA-based capture-mark-recapture population and density estimates of grizzly bears. J Wildl Manage 68: 457–469. [Google Scholar]
- 29. Sun CC, Fuller AK, Royle JA (2014) Trap configuration and spacing influences parameter estimates in spatial capture-recapture models. PLoS ONE 9(2): e88025 10.1371/journal.pone.0088025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Borchers DL, Efford MG (2008) Spatially explicit maximum likelihood methods for capture–recapture studies. Biometrics 64: 377–385. [DOI] [PubMed] [Google Scholar]
- 31.Royle JA, Gardner B (2011) Hierarchical spatial capture–recapture models for estimating density from trapping arrays. In: O’Connell AF, Nichols JD, Karanth KU, editors. Camera traps in animal ecology: Methods and analyses. New York: Springer-Verlag. 163–190. [Google Scholar]
- 32.Royle JA, Chandler RB, Sollmann R, Gardner B (2014) Spatial Capture-Recapture. Waltham, USA: Academic Press. 577 p. [Google Scholar]
- 33. Gardner B, Royle JA, Wegan MT (2009) Hierarchical models for estimating density from DNA mark-recapture studies. Ecology 90: 1106–1115. [DOI] [PubMed] [Google Scholar]
- 34. Efford MG, Fewster RM (2013) Estimating population size by spatially explicit capture–recapture. Oikos 122: 918–928. [Google Scholar]
- 35. Sollmann R, Gardner B, Belant JL (2012) How does spatial study design influence density estimates from spatial capture-recapture models? PLoS ONE 7: e34575 10.1371/journal.pone.0034575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Gardner B, Royle JA, Wegan MT, Rainbolt RE, Curtis PD (2010) Estimating black bear density using DNA data from hair snares. J Wildl Manage 74: 318–325. [Google Scholar]
- 37. Fry JA, Xian G, Jin S, Dewitz J, Homer C, et al. (2011) Completion of the 2006 National Land Cover Database for the Conterminous United States. Photogramm Eng Remote Sensing 77: 858–864. [Google Scholar]
- 38.Raeker G, Fleming J, Morris M, Moser K, Treiman T (2010) Missouri’s forest resource assessment and strategy, seeking a sustainable future for Missouri’s forest resources. Missouri Department of Conservation, Jefferson City, Missouri, USA and US Forest Service, Northern Research Station, Newtown Square, Pennsylvania, USA.
- 39.Nigh TA, Schroeder WA (2002) Atlas of Missouri ecoregions. Missouri Department of Conservation, Jefferson City, Missouri, USA.
- 40.United States Geological Survey (2009) Science in your backyard: Missouri. United States Geological Survey. Available: http://www.usgs.gov/state/state.asp?State=MO. Accessed 05 April 2011.
- 41. Woods JG, Paetkau D, Lewis D, McLellan BN, Proctor M, et al. (1999) Genetic tagging of free-ranging black and brown bears. Wildl Soc Bull 27: 616–627. [Google Scholar]
- 42. Wilton CW, Belant JL, Beringer J (2014) Distribution of American black bear occurrences and human-bear incidents in Missouri. Ursus 25: 53–60. [Google Scholar]
- 43.Missouri Spatial Data Information Service (2005) MSDIS Data Theme Listing. Available: http://msdis.missouri.edu/datasearch/ThemeList.jsp. Accessed 07 April 2011.
- 44. Gagneux P, Boesch C, Woodruff DS (1997) Microsatellite scoring errors associated with noninvasive genotyping based on nuclear DNA amplified from shed hair. Mol Ecol 6: 861–868. [DOI] [PubMed] [Google Scholar]
- 45.Eggert LS, Maldonado JE, Fleischer RC (2005) Nucleic acid isolation from ecological samples: animal scat and their associated materials. In: Elizabeth AZ, Eric HR, editors. Methods Enzymol. New York: Academic Press. 73–82. [DOI] [PubMed] [Google Scholar]
- 46. Paetkau D, Shields GF, Strobeck C (1998) Gene flow between insular, coastal, and interior populations of brown bears in Alaska. Mol Ecol 7: 1283–1292. [DOI] [PubMed] [Google Scholar]
- 47. Sanderlin JS, Faircloth BC, Shamblin B, Conroy MJ (2009) Tetranucleotide microsatellite loci from the black bear (Ursus americanus). Mol Ecol Resour 9: 288–291. [DOI] [PubMed] [Google Scholar]
- 48. Taberlet P, Camarra JJ, Griffin S, Uhres E, Hanotte O, et al. (1997) Noninvasive genetic tracking of the endangered Pyrenean brown bear population. Mol Ecol 6: 869–876. [PubMed] [Google Scholar]
- 49. Puckett EE, Kristensen TV, Wilton CW, Lyda SB, Noyce KV, et al. (2014) Influence of drift and admixture on population structure of American black bears (Ursus americanus) in the Central Interior Highlands, USA 50 years after translocation. Mol Ecol 23: 2414–2427. [DOI] [PubMed] [Google Scholar]
- 50. Valière N (2002) Gimlet: a computer program for analysing genetic individual identification data. Mol Ecol Notes 2: 377–379. [Google Scholar]
- 51. Carmichael LE, Krizan P, Polischuk Blum S, Strobeck C (2005) Genotyping of pseudohermaphrodite polar bears in Nunavut and advances in DNA sexing techniques. J Mammal 86: 160–169. [Google Scholar]
- 52.Efford MG (2013) secr: Spatially explicit capture–recapture models. R package version 2.5.0. http://CRAN.R-project.org/package=secr.
- 53.R Core Team (2013) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/.
- 54.Efford MG, Borchers DL, Byrom AE (2009) Density estimation by spatially explicit capture-recapture: likelihood-based methods. In: Thomson DL, Cooch EG, Conroy MJ, editors. Modeling Demographic Processes in Marked Populations. New York, USA: Springer Science+Business Media. 255–269. [Google Scholar]
- 55.Burnham KP, Anderson DR (2002) Model selection and multimodal inference: a practical information-theoretic approach. Second edition. New York: Springer-Verlag. 488 p. [Google Scholar]
- 56. Carter NH, Brown DG, Etter DR, Visser LG (2010) American black bear habitat selection in northern Lower Peninsula, Michigan, USA, using discrete-choice modeling. Ursus 21: 57–71. [Google Scholar]
- 57. Baldwin RA, Bender LC (2012) Estimating population size and density of a low-density population of black bears in Rocky Mountain National Park, Colorado. Eur J Wildl Res 58: 557–566. [Google Scholar]
- 58. Drewry JM, Van Manen FT, Ruth DM (2013) Density and genetic structure of black bears in coastal South Carolina. J Wildl Manage 77: 153–164. [Google Scholar]
- 59.Efford MG (2012) Density 5.0: software for spatially explicit capture-recapture. Department of Mathematics and Statistics, University of Otago, Dunedin, New Zealand. http://www.otago.ac.nz/density.
- 60. Efford MG, Warburton B, Coleman MC, Barker RJ (2005) A field test of two methods for density estimation. Wildl Soc Bull 33: 731–738. [Google Scholar]
- 61. Ngoprasert D, Reed DH, Steinmetz R, Gale GA (2012) Density estimation of Asian bears using photographic capture–recapture sampling based on chest marks. Ursus 23: 117–133. [Google Scholar]
- 62. Howe EJ, Obbard ME, Kyle CJ (2013) Combining data from 43 standardized surveys to estimate densities of female American black bears by spatially explicit capture–recapture. Popul Ecol 55: 595–607. [Google Scholar]
- 63. Pollock KH, Nichols JD, Brownie C, Hines JE (1990) Statistical Inference for Capture-Recapture Experiments. Wildlife Monogr 107: 3–97. [Google Scholar]
- 64. Noyce KV, Garshelis DL (2011) Seasonal migrations of black bears (Ursus americanus): causes and consequences. Behav Ecol Sociobiol 65: 823–835. [Google Scholar]
- 65. Sawaya MA, Stetz JB, Clevenger AP, Gibeau ML, Kalinowski ST (2012) Estimating grizzly and black bear population abundance and trend in Banff National Park using noninvasive genetic sampling. PLoS ONE 7(5): e34777 10.1371/journal.pone.0034777 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Noyce KV, Garshelis DL, Coy PL (2001) Differential vulnerability of black bears to trap and camera sampling and resulting biases in mark-recapture estimates. Ursus 12: 211–225. [Google Scholar]
- 67. Otis DL, Burnham KP, White GC, Anderson DR (1978) Statistical inference from capture data on closed animal populations. Wildlife Monogr 62: 1–135. [Google Scholar]
- 68.Karanth KU, Nichols JD, Kumar NS (2011) Estimating tiger abundance from camera trap data: field surveys and analytical issues. In: O’Connel AF, Nichols JD, Karanth KU, editors. Camera traps in animal ecology: Methods and analyses. New York: Springer-Verlag. 97–118. [Google Scholar]
- 69. Bales SL, Hellgren EC, Leslie DM, Hemphill J (2005) Dynamics of a recolonizing population of black bears in the Ouachita Mountains of Oklahoma. Wildl Soc Bull 33: 1342–1351. [Google Scholar]
- 70. Boulanger J, Proctor M, Himmer S, Stenhouse G, Paetkau D, et al. (2006) An empirical test of DNA mark-recapture sampling strategies for grizzly bears. Ursus 17: 149–158 10.2307/3873092 [DOI] [Google Scholar]
- 71. Lofroth EC, Krebs J (2007) The abundance and distribution of wolverines in British Columbia, Canada. J Wildl Manage 71: 2159–2169. [Google Scholar]
- 72. Russell RE, Royle JA, Desimone R, Schwartz MK, Edwards VL, et al. (2012) Estimating abundance of mountain lions from unstructured spatial sampling. J Wildl Manage 76: 1551–1561. [Google Scholar]
- 73. Turner MG, Arthaud GJ, Engstrom RT, Hejl SJ, Liu J, et al. (1995) Usefulness of spatially explicit population models in land management. Ecol Appl 5: 12–16. [Google Scholar]
- 74. Settlage KE, Van Manen FT, Clark JD, King TL (2008) Challenges of DNA-based mark-recapture studies of American black bears. J Wildl Manage 72: 1035–1042. [Google Scholar]
- 75. Kéry M, Gardner B, Stoeckle T, Weber D, Royle JA (2011) Use of spatial capture-recapture modeling and DNA data to estimate densities of elusive animals. Conserv Biol 25: 356–364. [DOI] [PubMed] [Google Scholar]
- 76. Soisalo MK, Cavalcanti S (2006) Estimating the density of a jaguar population in the Brazilian Pantanal using camera-traps and capture–recapture sampling in combination with GPS radio-telemetry. Biol Conserv 129: 487–496. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Microsatellite genotypes of individual bears sampled at hair snares.
(XLSX)
DNA encounter histories from extensive and intensive design field seasons.
(XLSX)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.