

Abstract
Antibiotic usage promotes intestinal colonization of antibiotic-resistant bacteria. However, whether resistant bacteria gain dominance in enteric microflora or disseminate to extraintestinal viscera remains unclear. Our aim was to investigate temporal diversity changes in microbiota and transepithelial routes of bacterial translocation after antibiotic-resistant enterobacterial colonization. Mice drinking water with or without antibiotics were intragastrically gavaged with ampicillin-resistant (Amp-r) nonpathogenic Escherichia coli (E. coli) and given normal water afterward. The composition and spatial distribution of intestinal bacteria were evaluated using 16S rDNA sequencing and fluorescence in situ hybridization. Bacterial endocytosis in epithelial cells was examined using gentamicin resistance assay and transmission electromicroscopy. Paracellular permeability was assessed by tight junctional immunostaining and measured by tissue conductance and luminal-to-serosal dextran fluxes. Our results showed that antibiotic treatment enabled intestinal colonization and transient dominance of orally acquired Amp-r E. coli in mice. The colonized Amp-r E. coli peaked on day 3 postinoculation and was competed out after 1 wk, as evidenced by the recovery of commensals, such as Escherichia, Bacteroides, Lachnospiraceae, Clostridium, and Lactobacillus. Mucosal penetration and extraintestinal dissemination of exogenous and endogenous enterobacteria were correlated with abnormal epithelial transcytosis but uncoupled with paracellular tight junctional damage. In conclusion, antibiotic-induced enteric dysbiosis predisposes to exogenous infection and causes systemic dissemination of both antibiotic-resistant and commensal enterobacteria through transcytotic routes across epithelial layers. These results may help explain the susceptibility to sepsis in antibiotic-resistant enteric bacterial infection.
Keywords: gut-derived sepsis, superbug, intestinal microbiota, epithelial barrier, bacterial endocytosis
high incidence of bloodstream infection with antibiotic-resistant bacteria or superbug has been reported in critical care units, of which the clinical isolates are mostly members of the Enterobacteriaceae family [e.g., Escherichia coli (E. coli) and Klebsiella pneumoniae] (29, 30, 34). Bacteria of the Enterobacteriaceae family are abundant in human feces and are part of the gut commensal microbiota (13, 33). It is generally believed that intestinal colonization of antibiotic-resistant bacteria may increase the risk of sepsis by selective overgrowth and dissemination in individuals who are clinically compromised.
One of the risk factors of antibiotic-resistant bacterial infection is previous history of antibiotic usage (34). Antibiotic treatment alters the intestinal microbiota, causing enteric dysbiosis (quantitative, qualitative, metabolic, or locational imbalances of gut commensals), which can take weeks to recover from after antibiotic withdrawal (16, 41). Previous studies in animal models showed that oral administration of antibiotics increase the susceptibility to pathogenic infection (9, 15, 26). Recent studies have indicated that usage of antibiotic cocktails induces clonal expansion and promotes colonization of multidrug-resistant enteric bacteria (1, 21, 35, 47), suggesting dominance of resistant organisms over commensals under antibiotic pressure. However, the time frame of antibiotic-resistant bacterial colonization and its relationship with commensal recovery after antibiotic withdrawal remain unclear.
Intestinal epithelial cells linked by tight junctions normally form a physical barrier to confine luminal bacteria (52). Once the barrier is breached, enteric bacteria may gain access to lamina propria through either transcellular or paracellular routes, causing microbial translocation to extraintestinal organs and systemic inflammation (17, 50). Abnormal adherence and internalization of commensal bacteria in epithelial cells have been documented in patients and experimental models with infection of Campylobacter jejuni (19) and Giardia lamblia (4), Crohn's disease (22, 46), celiac disease (12, 43), chronic psychological stress (14), surgical manipulation (11), and intestinal obstruction (39). However, the underlying mechanism of enterobacterial dissemination in antibiotic-induced dysbiosis is largely unknown. The double impact of commensal dysbiosis and exposure to high numbers of resistant bacteria on intestinal barrier functions has also yet to be explored.
Our specific aims were to investigate the colonization time course of exogenously administered resistant enterobacteria in normal flora and under antibiotic-induced dysbiosis, temporal diversity changes in the intestinal microbiota following exogenous bacteria colonization, and the mechanisms of enterobacterial dissemination to the mucosa and extraintestinal organs.
MATERIALS AND METHODS
Animals.
Specific pathogen-free BALB/c and C57BL/6 mice (5–7 wk of age, male) obtained from the Animal Center of the National Taiwan University were used. Myosin light-chain kinase-knockout (MLCK-KO) mice (B6 background) lacking the long 210-kDa MLCK were kind gifts from Dr. J. R. Turner, University of Chicago (8). Animals were raised in a temperature-controlled room (20 ± 2°C) with 12-h:12-h light/dark cycles and fed with regular mice chow and water. All experimental procedures were approved by the Animal Care and Use Committee of the National Taiwan University.
Antibiotic-resistant bacterial strains.
E. coli BL21 were transformed with pRSET containing an ampicillin-resistant (Amp-r) gene (a gift from Dr. Meng-Chun Hu, Graduate Institute of Physiology, National Taiwan University College of Medicine). E. coli BL21, a nonpathogenic laboratory strain derived from normal commensals of human gut, was originally isolated from human feces (10). Transformed E. coli were grown to log phase in Luria-Bertani (LB; Sigma, St. Louis, MO) broth containing 50 μg/ml ampicillin with vigorous shaking at 37°C. Colony-forming units (CFU) per milliliter were determined by spectrophotometry, and the required concentration of Amp-r E. coli was prepared in phosphate-buffered saline (PBS) for oral gavage.
Experimental design.
Mice were drinking normal water (NW) or antibiotic water (AW) containing mixtures of vancomycin (500 mg/l), neomycin (1 g/l), metronidazole (1 g/l), and ampicillin (1 g/l) (36) for 1 wk before oral gavage with Amp-r E. coli. Antibiotics were all purchased from Sigma. Mice were inoculated with 109 CFU of Amp-r E. coli in 0.2 ml PBS on day 0 and were given normal water for drinking afterward. On day 0 (before infection) and postinoculation (PI) days 1, 3, 7, and 14, mice were killed for sample collection.
Quantification of total and Amp-r bacteria in intestines.
To evaluate gut-associated bacterial colonization, 1-cm segments of mid-jejunum, cecum, and colon were excised and cut longitudinally, and the luminal contents were rinsed off. Tissues were weighed aseptically and transferred to sterile PBS at a ratio of 1 mg to 10 μl for homogenization and sonication. The lysate was plated on LB agar plates with or without 50 μg/ml ampicillin for aerobic and anaerobic bacterial culture overnight at 37°C, and the total and Amp-r bacterial counts were normalized per gram of intestinal tissues (CFU/g), representing the colonized bacterial numbers.
The bacterial numbers in jejunal chyme as well as in fecal samples of the cecum and colon were also examined. The luminal contents of each intestinal segment were collected and weighed, followed by homogenization and sonication in sterile PBS at a ratio of 1 mg to 10 μl. Diluted samples were plated onto LB agar plates with or without 50 μg/ml ampicillin for aerobic and anaerobic culturing overnight, and the total and Amp-r bacterial CFUs were normalized per gram of luminal content (CFU/g), representing luminal bacterial numbers.
Analysis of bacterial translocation to extraintestinal organs.
The livers and spleens were removed from animals with sterile instruments, and tissue weights were determined. The tissues were homogenized and sonicated in sterile PBS at a ratio of 1 mg to 10 μl. Undiluted lysate (200 μl) was inoculated onto LB agar plates with or without 50 μg/ml ampicillin for aerobic bacterial culturing. Following incubation at 37°C for 24 h, bacterial colonies were counted and normalized to CFU/g (28, 51).
Quantification of epithelial endocytosed bacteria.
Segments of mid-jejunum (10 cm) and colon (2 cm) were excised, and the luminal contents were rinsed out with sterile PBS as previously described (4, 51). The tissues were placed inside-out on a steel rod and incubated in PBS containing 0.5 mM dithiothreitol (Sigma) for 10 min at room temperature to remove mucus. The segments were transferred into PBS with 30 mM ethylenediaminetetraacetic acid (EDTA) and 5.55 mM glucose and incubated for 20–30 min at 37°C and shaken gently. The solution was then passed through a nylon mesh with a pore size of 40 μm (BD Bioscience, San Jose, CA) to retain epithelial sheets. The epithelial sheets were washed out from the inverted mesh with PBS and separated into single cells by gentle pipetting. The viability and purity of enterocytes were determined in a pilot study, showing Trypan blue negativity (91.7 ± 0.6%), BerEP4 (an epithelial marker) positivity (91.1 ± 1.2%), and a lack of CD68 (a macrophage marker) transcripts in the cell preparation.
The cells (2 × 106) were then incubated with 300 μg/ml gentamicin (Invitrogen, Carlsbad, CA) for 1 h with gentle shaking. After being washed twice with PBS, cells were incubated with 1% Triton X-100 in PBS for 10 min on ice. Our pilot study confirmed that none of the intestinal bacteria was resistant to gentamycin. The lysate (200 μl) was plated onto agar plates for aerobic bacterial culturing overnight at 37°C. The numbers of bacterial colonies are presented as CFU per 106 cells (4, 51).
Temporal temperature gradient gel electrophoresis and 16S rDNA sequencing.
Colonic and cecal stool samples of the mice (0.1–0.5 g wet weight) were immediately diluted 10-fold in peptone/water containing 20% glycerol and stored at 20°C until analysis. DNA (in a 0.2-ml volume) was isolated from the diluted feces samples using the QIAamp DNA stool mini kit. Then, stool DNA was amplified by PCR using a T3000 Thermocycler (Biometra, Gottingen, Germany) with universal bacterial primers for 16S rDNA (341F with 40 bp GC at the 5′ end and 534R of E. coli) (2, 4). The PCR products were separated by temporal temperature gradient gel electrophoresis (TTGE) in a 10% polyacrylamide gel (37.5:1 acrylamide-bisacrylamide) and 7 M urea (J.T. Baker, Phillipsburg, NJ). Premigration was conducted at 20 V and 55°C over a 15-min period. Gels were run for 6 h at 150 V with the temperature increasing at 2.5°C/min from 55 to 70°C. The DNA fragments were visualized using SYBR Green staining (Life Technologies, Carlsbad, CA), and the gel was scanned using a Gel System MultiGel-21 (Topbio, Taipei, Taiwan).
TTGE bands were excised from the gel and reamplified with the same 341F and 534R primers but without the GC clamp. The PCR products were digested and subcloned into the pGEM-T Easy Vector Systems (Promega, Madison, WI) and transformed into cells. The TTGE fragment insert was then sequenced (Applied Biosystems, Foster City, CA) and classified using the Classifier Tool provided by the Ribosomal Database Project (RDP; http://rdp.cme.msu.edu/classifier/classifier.jsp). The number of bands, indicative of bacterial diversity, was determined.
Fluorescence in situ hybridization.
Microbiological analysis in intestinal tissues was conducted as described previously (4, 51). Briefly, intestinal segments (1 cm) were excised and cut longitudinally, and the luminal contents were rinsed off. Tissues were fixed in Carnoy's solution (Ricca Chemical, Arlington, TX), embedded in paraffin, and sectioned at 5 μm. The sections were dewaxed, placed in 1% Triton X-100, and washed in PBS three times before incubation in 5 mg/ml lysozyme at 37°C for 20 min. After PBS washing, the sections were incubated in prewarmed hybridization solution containing 0.02–0.1 μM oligonucleotide probes targeting bacterial 16S rDNA (see below) at 46°C overnight. The sections were then rinsed in washing buffer and air-dried before being stained with a Hoechst dye. The slides were mounted and viewed under a fluorescent microscope (Axio Imager A1; Carl Zeiss, Jena, Germany) equipped with a CCD camera and imaging software.
The probes included 5′-end FITC-labeled universal bacterial probe (EUB338) (5′-GCTGCCTCCCGTAGGAGT-3′) and negative control probe (non-EUB338) (5′-ACATCCTACGGGAGGC-3′), as well as 5′-end Cy3-labeled probes for Lactobacillus/Enterococcus (Lab158) (5′-GGTATTAGCACCTGTTTCCA-3′), E. coli (Ecol1513) (5′-CACCGTAGTGCCTCGTCATCA-3′), and Bacteroides (Bac 303) (5′-CCAATGTGGGGGACCTT-3′) (Genomics BioScience and Technology, Taipei, Taiwan).
Transmission electron microscopy.
Intestinal tissues were fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.4) for 2 h at 22°C and rinsed with 0.05 M Tris buffer (pH 7.4) for 4 h at 4°C. Tissues were osmicated, dehydrated in a graded ethanol series, and embedded in epoxy resin. Thick vibrotome sections (100 μm) were cut and washed in Tris buffer three times for 5 min each. Thin sections (70 nm) were then cut and examined in a Hitachi-7100 electronmicroscope equipped with a digital system. Electromicrographs of epithelial cells were taken at magnifications of ×6,000 and ×15,000 (50).
Histological examination.
Intestinal segments were fixed in 4% paraformaldehyde (PFA), and care was taken to ensure proper orientation of the crypt-to-villus axis during embedding. Sections of 4-μm thickness were stained with hematoxylin and eosin and observed under a light microscope.
Ussing chamber studies and macromolecular flux assay.
Intestinal segments were opened along the mesenteric border and mounted in Ussing chambers (WPI Instruments, Worcester, MA) as previously described (17, 50). The opening of the chamber (1 cm2) exposed the tissue to 5-ml circulating oxygenated Krebs buffer. The serosal buffer contained 10 mmol/l glucose that was osmotically balanced with 10 mmol/l mannitol in the mucosal buffer. A circulating water bath maintained the temperature of the buffer at 37°C. The potential difference (PD) between the two compartments was clamped at 0 V using a voltage clamp amplifier, and the short-circuit current (Isc) of the tissue was then determined. The tissue conductance (G) was calculated according to Ohm's law by dividing the value of Isc by that of PD.
Intestinal permeability was determined by the level of mucosal-to-serosal flux of dextran conjugated to FITC (dextran-FITC, molecular weight = 4 kDa; Sigma). Tissues mounted on the Ussing chambers were allowed to equilibrate until the Isc stabilized before dextran probe was added to the mucosal buffer at a final concentration of 500 μM. Samples (250 μl) of serosal buffer were collected at 0, 30, 60, and 90 min after addition of dextran probe and were replaced with Krebs buffer/glucose. The fluorescence units of dextran-FITC in serosal buffer were determined at excitation/emission = 490/530 nm using a multi-mode plate reader (Beckman Coulter, Fullerton, CA), and the concentration (nM) was calculated according to a standard curve.
Staining of tight junctional structures on intestinal tissues.
A 1-cm gut segment was fixed with 4% PFA for 1 h at 4°C and then snap-frozen in optimal cutting temperature solution (Thermo Fisher Scientific, Rockford, IL). Frozen tissues were cut into 6-μm sections with a cryostat and mounted on precoated glass slides for immunofluorescence staining following previously described protocols. Frozen tissue sections were prefixed in cold acetone for 10 min, air dried, and fixed again with 4% PFA for 2 min. The tissues were then permeabilized with 1% Triton X-100 for 1 min and blocked with fetal bovine serum for 1.5 h at room temperature. Tissue sections were then incubated with rabbit anti-zonula occludens (ZO)-1 (1:50; Zymed, San Francisco, CA) overnight at 4°C. After being washed with PBS, tissues were incubated with goat anti-rabbit IgG conjugated to Alexa Fluor 488 for 1 h at room temperature. Tissues were then stained for F-actin by labeling with phalloidin conjugated to Alexa Fluor 633 (5 U/ml, Invitrogen) for 20 min. The slides were washed and stained with a Hoechst dye to visualize cell nuclei. Images were captured under a Zeiss fluorescence microscope equipped with a camera (18, 50).
Western blotting for occludin cleavage and proinflammatory signaling molecules.
Scraped mucosa from mouse intestines were lysed with complete radio immunoprecipitation assay buffer for 60 min on ice. The lysates were centrifuged (14,000 g, 15 min), and the supernatants were collected for a protein assay. Protein samples were adjusted to 5 mg/ml, dissolved, and boiled in 2× electrophoresis sample buffer. The samples were frozen at 20°C until used for Western blotting. Extracted protein samples were subjected to SDS/PAGE (4-13% polyacrylamide), and the resolved proteins were electrotransferred onto Hybond-P PVDF or nitrocellulose membranes in a semi-dry blotter. Blots were blocked with 5% (wt/vol) nonfat dry milk in Tris-buffered saline (TBS) or 5% (wt/vol) bovine serum albumin in TBS with Tween 20 (TBS-T) for 1 h at room temperature, washed three times with TBS-T [0.1% (vol/vol) Tween-20 in TBS], and incubated with primary antibody at 4°C overnight with gentle shaking. The membrane was washed with TBS-T and incubated with a secondary antibody for 1 h at room temperature and then washed with TBS-T. The membranes were incubated with chemiluminescent solution (ECL; Millipore, Billerica, MA) for 1 min with gentle agitation. The signal was detected on a UVP AutoChemi system (UVP, Upland, CA). Band density was determined using the Gel-pro Analyzer 4.0 software (Media Cybernetics, Rockville, MD) (17, 50).
Primary antibodies included those targeting occludin (1:3,000, Invitrogen), phospho (p)- and total (t)-Iκ-Bα (1:1,000; Cell Signaling, Beverly, MA), p- and t-ERK1/2 (1:2,000, Cell Signaling), p- and t-JNK (1:1,000, Cell Signaling), p- and t-p38 (1:1,000, Cell Signaling), and anti-β-actin (1:10,000, Sigma). The secondary antibodies used were horseradish peroxidase-conjugated goat anti-mouse IgG or anti-rabbit IgG (1:1,000, Cell Signaling).
ELISA for IFN-γ and TNF-α.
Intestinal segments (1 cm) were homogenized and sonicated in PBS, and the lysate was centrifuged. Levels of IFN-γ and TNF-α were measured using ELISA development kits (Peprotech, Rocky Hill, NJ), according to the manufacturer's instructions.
Statistical analysis.
All data except for band numbers are expressed as means ± SE, and the mean values were analyzed by one-way ANOVA, followed by a Student Newman-Keuls test. When comparing the difference between two groups of data that failed to satisfy the normality assumption, a nonparametric Mann-Whitney U-test was used. Significance was set at a value of P < 0.05.
RESULTS
Antibiotic treatment decreases intestinal bacterial numbers in mice.
To verify absence of resistant strains in intestines and antibiotic killing of commensals, the gut-associated and luminal bacterial counts were determined by culturing tissue lysates and luminal contents, respectively, after 1 wk of NW and AW treatment. A two- to four-log decrease in gut-associated bacterial counts was seen in the jejunum, cecum, and colon of AW mice, compared with NW controls (Table 1). Luminal bacterial numbers were also significantly lower in AW mice than NW mice (Table 1). After antibiotic removal for 14 days, the bacterial counts in AW mice returned to normal levels, comparable to those drinking NW (Table 1). Moreover, no bacterial colony was found on ampicillin-containing agar plates, suggesting that antibiotic-resistant bacteria were not present in the intestines of either group.
Table 1.
Gut-associated and luminal bacterial counts in mice drinking NW and AW for 1 wk, and in those after withdrawal of AW for 14 days
| Bacterial count, log10CFU/g | NW | AW | Withdrawal |
|---|---|---|---|
| Gut-associated bacteria | |||
| Aerobic | |||
| Jejunum | 4.41 ± 0.48 | 1.60 ± 0.67 | 5.49 ± 0.80 |
| Cecum | 6.42 ± 0.11 | 3.02 ± 0.61* | 6.09 ± 0.34 |
| Colon | 6.27 ± 0.19 | 2.24 ± 0.60* | 6.33 ± 0.85 |
| Anaerobic | |||
| Jejunum | 4.19 ± 0.45 | 1.33 ± 0.71 | 5.58 ± 1.21 |
| Cecum | 6.54 ± 0.24 | 3.12 ± 0.80* | 6.85 ± 1.30 |
| Colon | 6.30 ± 0.20 | 2.10 ± 0.60* | 6.74 ± 0.95 |
| Luminal bacteria | |||
| Aerobic | |||
| Jejunum | 6.24 ± 5.71 | 3.26 ± 3.22* | 6.81 ± 5.48 |
| Cecum | 8.55 ± 8.06 | 4.08 ± 3.70* | 7.76 ± 7.31 |
| Colon | 7.89 ± 7.36 | ND* | 8.12 ± 7.30 |
| Anaerobic | |||
| Jejunum | 6.06 ± 5.65 | 1.90 ± 1.90* | 5.33 ± 4.71 |
| Cecum | 6.94 ± 6.19 | 3.62 ± 3.45* | 6.84 ± 6.07 |
| Colon | 7.04 ± 6.23 | ND* | 6.92 ± 6.14 |
Values represent means ± SE.
The tissue homogenate and luminal content of each intestinal segment were plated on agar plates under aerobic and anaerobic conditions. NW, normal water; AW, antibiotic water; ND, not detected. CFU, colony-forming units.
P < 0.05 vs. NW. n = 8–10/group.
Intestinal colonization of Amp-r bacteria is seen only in mice after antibiotic treatment.
In the following experiments, mice were orogastrically inoculated with Amp-r E. coli to evaluate exogenous colonization of resistant bacteria. The total and Amp-r bacterial counts in jejunal, cecal, and colonic tissues were determined on day 0 (before inoculation), or PI days 1, 3, 7, and 14. In NW mice, none of the intestinal segments showed presence of Amp-r bacteria at any time point (Fig. 1A). The total bacterial numbers remained constant in cecal and colonic segments after inoculation of Amp-r E. coli; a decrease without statistical significance was seen in jejunal counts on PI day 3 (Fig. 1A). The anaerobic bacterial counts showed a similar trend to those of aerobic culturing, with no sign of Amp-r E. coli colonization in NW mouse intestines (data not shown).
Fig. 1.
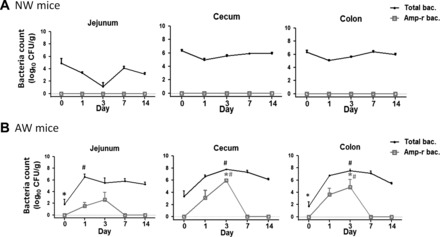
Total and ampicillin-resistant (Amp-r) bacterial (bac) counts in mouse intestines by aerobic culturing. Intestinal homogenates of normal water (NW) (A) and antibiotic water (AW) (B) mice postinoculated with Amp-r Escherichia coli (E. coli) were aerobically cultured on agar plates with or without ampicillin. Data are expressed as means ± SE. The dashed line indicates that no colony was detected on agar plates. *P < 0.05 vs. NW; #P < 0.05 vs. day 0. n = 5–6/group. The experiments were repeated twice.
In contrast, increased numbers of Amp-r bacteria were seen colonizing the cecal and colonic segments of AW mice compared with NW mice on PI day 3 (Fig. 1B). The numbers of colonized-resistant strains were higher in the cecum and colon than in the jejunum on PI day 3, whereas no Amp-r bacteria were detected after 7 days in any segment (Fig. 1B). The Amp-r bacteria were identified to be Escherichia genus by 16S rDNA sequencing. The total bacterial counts in intestinal tissues increased two- to four-log on PI days 1–3 and plateaued afterward, compared with day 0, in AW mice (Fig. 1B). Additionally, the luminal numbers of Amp-r bacteria in fecal samples of colon and cecum also peaked on PI day 3 in AW mice, whereas none was found in jejunal luminal contents (data not shown).
Clearance of Amp-r bacteria is associated with recovery of commensal microbiota.
The diversity of enteric microbiota was examined using TTGE and 16S rDNA sequencing, whereby DNA extracted from colonic stool was amplified using universal bacterial primers and separated into multiple bands on gradient gels. The band numbers amplified from colonic samples of NW mice were similar on day 0 (before inoculation) and 1, 3, 7, and 14 days after oral gavage of Amp-r E. coli (Fig. 2, A and B), indicating no alteration in bacterial diversity. Several bacterial taxa were identified in the NW colonic stool, such as Bacteroides, Porphyromonadaceae, Escherichia, Mucispirillum, Lactobacillus, Lachnospiraceae, Clostridium, and Ruminococcaceae.
Fig. 2.
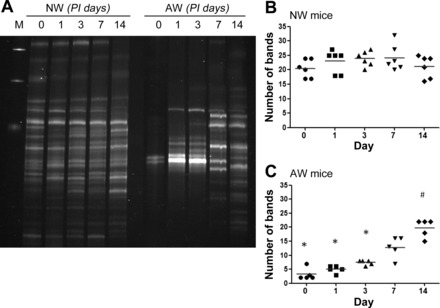
Analysis of bacterial diversity and strains in colonic stool by temporal temperature gradient gel electrophoresis and 16S rDNA sequencing. Colonic stool samples were collected from NW and AW mice on various postinoculation (PI) days after orogastric gavage of Amp-r E. coli. A: extracted fecal DNA were amplified with universal bacterial primers and separated into multiple bands on gradient gels. The bands on gels were later excised and sequenced. B and C: numbers of bands derived from stool samples of NW and AW mice represent the bacterial diversity. Each data point represents the values from 1 animal, and the median values are shown as bars. *P < 0.05 vs. NW; #P < 0.05 vs. day 0. n = 5–6/group.
Decreased band numbers were found in the colonic stool of AW mice compared with NW mice on day 0, indicating a loss of microbial diversity after antibiotic treatment (Fig. 2, A–C). The degree of bacterial diversity increased gradually and was close to normal levels by PI day 14 in AW mice (Fig. 2, A and C), suggesting recovery of commensal microbiota in about 2 wk after antibiotic withdrawal. The recovered bacterial strains on PI day 14 in AW mice included Escherichia, Bacteroides, Lactobacillus, Lachnospiraceae, and Clostridium.
Mucosal penetration and extraintestinal translocation of both resistant and commensal bacteria in antibiotic-treated mice.
Normal morphological and histological structures of gastrointestinal tracts were seen in NW mice before and after inoculation of Amp-r E. coli and also in AW mice on PI day 0 (before inoculation) (data not shown), suggesting that antibiotics treatment per se did not cause structural damages. Cecal flatulence or distension associated with tissue hyperemia and edema were evident in AW mice on PI days 1 and 3, of which the structures returned to normal on PI days 7–14 (data not shown). The jejunal and colonic histology in AW mice remains intact after colonization of resistant bacteria (data not shown).
We next evaluated the presence of bacteria in gut mucosa by in situ hybridization. To avoid contamination with luminal bacteria, chyme or fecal contents were washed off before tissue processing. No sign of bacteria was seen in intestinal mucosa (Fig. 3A) and extraintestinal organs (liver and spleen; Fig. 3D) of NW mice after inoculation of Amp-r E. coli. In contrast, bacteria were found in the intestinal crypts, lamina propria, and epithelial layer of AW mice (Fig. 3B). By hybridization with probes targeting specific bacterial strains, penetration of E. coli, Bacteroides, and Lactobacillus was noted in the gut mucosa of AW mice (Fig. 3C). Moreover, augmentation of bacterial translocation to the liver and spleen was seen on PI day 3 in AW mice compared with NW mice (Fig. 3, D and E).
Fig. 3.
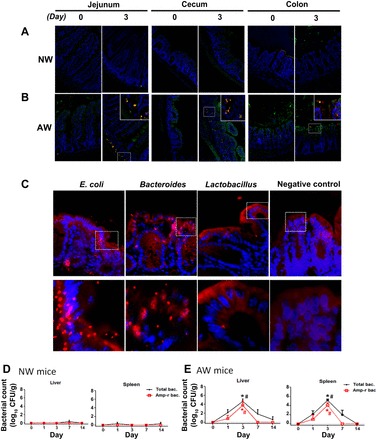
Bacterial penetration into gut mucosa and extraintestinal organs of AW mice after infection with Amp-r E. coli. A and B: jejunal, cecal, and colonic segments of NW and AW mice were subjected to fluorescence in situ hybridization with probes targeted to universal bacteria (green) and E. coli (red). Cell nuclei (blue) have been superimposed on the images. Representative photomicrographs (magnification ×200) were obtained from at least 5 mice per group. Inset: higher magnification of bacteria. C: penetration of E. coli, Bacteroides, and Lactobacillus to intestinal mucosa of AW mice on PI day 3. D and E: extraintestinal bacterial counts in liver and spleen homogenates of NW and AW mice. *P < 0.05 vs. NW; #P < 0.05 vs. day 0. n = 5/group. CFU, colony-forming units.
Presence of transcellular but not paracellular epithelial barrier dysfunction.
To investigate the routes for enteric bacterial translocation, transcellular and paracellular epithelial permeability was assessed. First, intracellular bacterial counts were measured in purified epithelial cells isolated from jejunal, cecal, and colonic segments using a gentamycin assay. The epithelial endocytosed bacterial count was minimal in NW mice (Fig. 4A), whereas a significant increase in bacterial numbers was found in the small and large intestinal epithelial cells in AW mice during peak colonization on PI day 3 (Fig. 4B). The presence of Amp-r bacteria was noted in the enterocytes of the cecum and colon of AW mice (Fig. 4B), of which the resistant bacteria were identified to be Escherichia genus by 16S rDNA sequencing. Moreover, electromicrographic images revealed bacterial internalization to jejunal and colonic epithelial cells in AW mice on PI day 3 after inoculation, whereas no sign of intracellular bacteria was seen in NW mice (Fig. 4C).
Fig. 4.
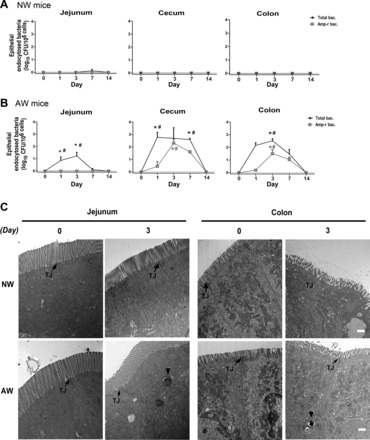
Increased bacterial endocytosis in intestinal epithelial cells of AW mice after inoculation of Amp-r E. coli. The number of intracellular bacteria was determined in jejunal and colonic epithelial cells of NW mice (A) and AW mice (B), using a gentamycin resistance assay. *P < 0.05 vs. NW; #P < 0.05 vs. day 0. n = 5/group. The experiments were repeated at least twice. C: electromicrographs showing bacterial internalization to epithelial cells in AW mice. Representative images showing jejunal and colonic epithelial cells of NW and AW mice on PI day 0 (before infection) and day 3 after orogavage of antibiotic-resistant bacteria. Intracellular presence of bacteria (arrowheads) was seen in jejunal and colonic segments of AW mice after infection but not in NW mice. Intact tight junctions (TJ) were seen in epithelial cells in all groups. Scale bar = 1 μm. n = 5/group.
The tissue conductance, luminal-to-serosal dextran flux, and tight junctional structures were next examined to verify changes in paracellular permeability. In both NW and AW mice, inoculation of Amp-r E. coli (PI day 3) did not alter the tissue conductance or dextran flux in jejunal and colonic segments, compared with day 0 (Fig. 5, A and B). Furthermore, normal honeycomb pattern of the ZO-1 and full-length occludin without cleavage were seen in the intestinal mucosa of both NW and AW mice (Fig. 5, C and D). Intact tight junctions were also observed in electromicrographs of intestinal epithelial cells in both NW and AW mice on PI days 0 and 3 (Fig. 4C). Taken together, the results suggested that neither antibiotic treatment nor bacteria inoculation caused tight junctional damage.
Fig. 5.
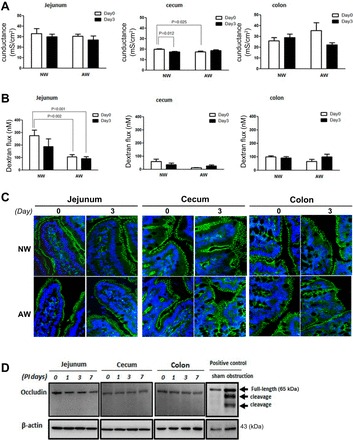
No increase in paracellular tight junctional permeability was seen in NW and AW mice postinoculation of resistant bacteria. Mouse jejunal, cecal, and colonic segments were used for measurement of tissue conductance and macromolecular flux and assessed for tight junctional structures. A: intestinal tissue conductance in NW and AW mice on days 0 and 3 after inoculation of Amp-r E. coli. B: luminal-to-serosal dextran flux in intestinal segments of NW and AW mice on days 0 and 3 after inoculation of Amp-r E. coli. n = 7–9/group. C: representative immunofluorescent staining of zonnula occludens (ZO)-1 revealed a honeycomb pattern of tight junctions on villus surfaces in both groups of mice. The green color indicates ZO-1 staining; the blue color indicates nuclear staining by a Hoechst dye. D: Western blots showed no sign of occludin cleavage in intestinal mucosal samples of AW mice following gavage of Amp-r E. coli. The positive control for cleaved occludin was mucosal samples from mice with bowel obstruction vs. sham operation. Representative blots from at least 2 individual experiments. n = 5/group.
Additional experiments were conducted to elucidate the mechanisms of transcellular passage of enteric bacteria. Our previous study had shown that long MLCK-210 kDa was involved in bacterial endocytosis by small intestinal epithelial cells in a mouse model of bowel obstruction (51). The activation of MLCK-210 caused terminal web myosin phosphorylation and brush border fanning, allowing bacterial penetration through intermicrovillus clefts in enterocytes. To assess the role of MLCK-210 in the mechanism of bacterial endocytosis following antibiotic-resistant bacterial colonization, wild-type (WT) mice and MLCK-210-KO mice were orally inoculated with Amp-r E. coli after they drank NW or AW. Increased numbers of gut-associated total and Amp-r bacterial counts were found in the large intestines of both mouse strains drinking AW compared with NW, suggesting comparable levels of antibiotic-resistant bacterial colonization in the two mouse strains (Fig. 6, A and B). In parallel with Amp-r bacterial colonization, the epithelial endocytosed bacterial counts in the large intestines of both mouse strains were also higher in groups drinking AW compared with NW (Fig. 6, C and D). The intracellular total bacterial counts in intestinal epithelial cells of MLCK-KO mice were slightly lower than WT mice (64 ± 23 vs. 287 ± 136 CFU/106 cells, P > 0.05) (Fig. 6C), albeit without statistical significance. An insignificant decrease of the numbers of endocytosed Amp-r bacteria was found in MLCK-KO mice compared with those of WT mice (12 ± 3 vs. 33 ± 13 CFU/106 cells, P > 0.05) (Fig. 6D).
Fig. 6.
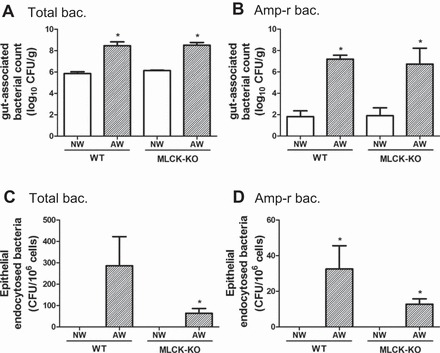
Role of myosin light-chain kinase (MLCK)-210 in mechanisms of bacterial endocytosis by intestinal epithelial cells after Amp-r E. coli colonization. Wild-type (WT) mice and MLCK- knockout (KO) mice were orally inoculated with Amp-r E. coli after drinking NW or AW, and large intestinal tissues were collected for analysis. A and B: numbers of gut-associated total and Amp-r bacteria colonized in large intestines were determined by culturing tissue homogenates. C and D: intracellular total and Amp-r bacterial counts in epithelial cells were also determined. *P < 0.05 vs. NW. n = 6–8/group.
Increase in proinflammatory signals and cytokines in gut tissues.
Lastly, the phosphorylation levels of proinflammatory signaling pathways (JNK, ERK, p38, Iκ-Bα) were examined in gut mucosa following antibiotic-resistant bacterial colonization. Comparable levels were seen before (day 0) and after (day 3) inoculation of Amp-r E. coli in mice drinking NW (data not shown). On the other hand, upregulated phosphorylation of Iκ-Bα and ERK in colonic mucosa was seen after Amp-r E. coli colonization in mice drinking AW (Fig. 7, A and B). No difference in phosphorylated levels of JNK and p38 was seen after resistant bacterial colonization in AW mice (Fig. 7, C and D). Moreover, increased IFN-γ levels in gut tissues were noted after infection in AW mice compared with NW mice (Fig. 7E). The TNF-α level was not different among groups (Fig. 7F).
Fig. 7.
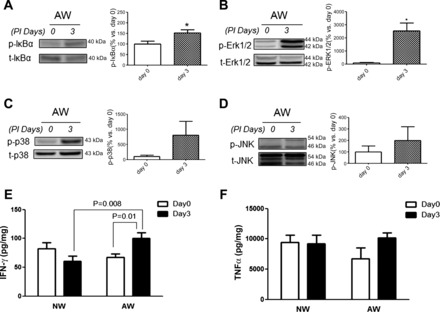
Colonic inflammation was noted after Amp-r E. coli inoculation in mice drinking AW but not NW. Western blots showing the phosphorylation levels of Iκ-Bα (A), ERK1/2 (B), p38 (C), and JNK (D) in colonic mucosa before (day 0) and after (day 3) inoculation of Amp-r E. coli in mice drinking AW. Band densities are expressed as means ± SE. Representative blots of at least 2 individual experiments. *P < 0.05 vs. day 0. n = 5–6/group. Levels of IFN-γ (E) and TNF-α (F) in colonic tissues of NW and AW mice were also evaluated. n = 5–6/group.
DISCUSSION
Emergence of antibiotic-resistant bacteria has become a worldwide public health problem, challenging clinical microbiologists and infection control professionals. Previous studies showed that antibiotic usage promotes colonization and selective expansion of antibiotic-resistant enterobacterial species in the intestinal tract (1, 21, 35, 47). Novel findings from our study indicate that transient dominance of exogenous enterobacteria caused by antibiotic manipulation may lead to microbial dissemination to extraintestinal viscera through transcytotic passage. We demonstrated that transcellular microbial entry was uncoupled with tight junctional damage, and the invasiveness was limited to antibiotic-resistant strains and also to commensals.
Orogastric gavage of antibiotic-resistant E. coli did not lead to intestinal colonization in mice with normal flora. It is noteworthy that a decrease, albeit with statistical insignificance, of total bacterial counts was seen in the NW mouse jejunum 3 days after inoculation. We speculate that orogastric gavage of Amp-r E. coli may have an impact on the numbers of normal commensals, possibly by stimulation of antimicrobial peptides. However, neither microbiota population change nor exogenous infection was seen in NW mice. On the other hand, colonization of antibiotic-resistant bacteria was observed after commensals were depleted by antibiotics. This finding is in keeping with previous studies (40, 47), supporting clinical observations that antibiotic usage may increase susceptibility to secondary infection. We also demonstrated that clearance of exogenous bacteria from the intestinal tract paralleled the recovery of commensal diversity after antibiotic withdrawal. The findings here bring novel information for strategies of vaccine design employing mucosal colonization of constructed bacteria for antigen delivery (5, 20). Future studies to tease out which commensals are important in competitive elimination of exogenous colonization and their mode of action are warranted.
Our results showed that intestinal colonization of antibiotic-resistant bacteria upon dysbiosis triggered the translocation of resistant and commensal bacteria to gut mucosa and extraintestinal organs. A previous study showed that oral antibiotics induce the expansion of multi-drug resistant enterobacterial strains from the population of commensal microbiota, of which the resistant strain may invade gut mucosa after chemical-induced colitis (1). We provided evidence here that both exogenous resistant strains and endogenous enterobacteria may translocate across the epithelial layer, and chemical disruption of epithelial barrier is not a prerequisite for enteric microbial penetration. In our study, bacterial influx was accompanied by increased proinflammatory signals and upregulated IFN-γ levels in gut tissues. The phenomenon of bacterial translocation and the subsequent gut-derived septic complications would be detrimental to immunocompromised individuals, such as patients with acquired immunodeficiency virus infection and those receiving immunosuppressant therapy, who are often already on multiple long-term antibiotic regimens (38).
Translocation of resistant and commensal bacteria to extraintestinal organs was observed despite normal epithelial tight junctional structures in antibiotic-treated mouse intestines. The results indicated that antibiotics alone did not render any of the gut segments permeable. Thus colonization and exposure to large numbers of resistant enterobacteria may be the main cause triggering enhanced transepithelial passage of microbes. We found a transient increase in intracellular bacteria counts in jejunal, cecal, and colonic epithelial cells after antibiotic-resistant bacterial infection. Abnormal epithelial endocytosis of commensals or nonpathogenic bacteria has been reported in animal models of infection with Campylobacter jejuni (19) and Giardia lamblia (4), psychological stress (44, 53), and chemically induced colitis (32). Moreover, presence of mucosal adherent and invasive bacteria or E. coli was documented in patients with inflammatory bowel disease, surgical stress, and necrotizing enterocolitis (3, 23, 37). In vitro epithelial cell cultures studies also showed the internalization and endocytosis of nonpathogenic bacteria under inflammatory conditions and metabolic stress, such as low-dose IFN-γ (6), TNF-α during glutamine deprivation (7), uncoupling of mitochondrial oxidative phosphorylation (27), and hypoxia (49). Although we cannot rule out the possibility of cells with phagocytic functionality (e.g., M cells and dendritic cells) for internalizing bacteria (45), our in situ hybridization data did show augmented adherence and colocalization of bacteria with the large surface area of epithelial layers.
Our previous study had shown that long MLCK-210 kDa was involved in terminal web myosin phosphorylation and brush border fanning, allowing bacterial penetration through intermicrovillus clefts in small intestinal epithelial cells, in a mouse model of bowel obstruction (51). In the current model of antibiotic-resistant bacterial colonization, the lack of MLCK-210 appears to decrease the level of microbial internalization by large intestinal epithelial cells, albeit without statistical significance. We speculate that the dependency on MLCK-210 for bacterial endocytosis may differ between epithelial cells with short vs. long brush border. Because antibiotic-resistant bacteria colonize mainly the large intestine rather than the small intestine, alternative mechanisms other than MLCK-210 may be responsible for bacterial internalization.
The enterobacterial penetration and dissemination paralleled the colonization time frame of Amp-r E. coli, both of which peaked on PI day 3 and were eliminated by day 14, indicating that recovery of commensal bacterial diversity, not only cleared out exogenous microbes, but also reestablished the epithelial defenses against luminal commensals. Several lines of evidence show that the normal microbiota is involved in the regulation of epithelial turnover and barrier fortification (24, 42). Previous studies have shown that administration of a single or mixtures of probiotics, such as Lactobacillus spp., E. coli Nissle 1917, Enterococcus faecalis, and Bifidobacterium brevis, may prevent the epithelial permeability increase and bacterial translocation in various disease models (25, 31, 48, 53). Taken together, interventions fortifying the symbiont/probiotic population or epithelial barriers may be considered as adjunct therapy for patients receiving long-term antibiotic therapy.
In conclusion, disruption of enteric microflora predisposes colonization of orally acquired antibiotic-resistant organisms, leading to systemic dissemination of both commensal and resistant bacteria through transcytotic routes in the absence of paracellular tight junctional damage. The subsequent gut-derived septic complications would be detrimental to immunocompromised individuals, who are often already on multiple long-term antibiotic regimens.
GRANTS
This work was supported by grants from the National Science Council (NSC100-2321-B002-022; NSC99-2628-B-002-008-MY3; NSC102-2628-B-002-009-MY3) and National Taiwan University (102R39012).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
L.C.H.Y. and Y.H.N. conception and design of research; L.C.H.Y., K.S.L., S.C.W., J.R.T., and Y.H.N. interpreted results of experiments; L.C.H.Y. and Y.H.N. drafted manuscript; L.C.H.Y. and Y.H.N. edited and revised manuscript; L.C.H.Y., Y.A.S., L.L.W., Y.D.L., W.T.K., W.H.P., K.S.L., S.C.W., J.R.T., and Y.H.N. approved final version of manuscript; Y.A.S., L.L.W., Y.D.L., W.T.K., and W.H.P. performed experiments; Y.A.S., L.L.W., Y.D.L., W.T.K., W.H.P., K.S.L., and S.C.W. analyzed data; Y.A.S., L.L.W., Y.D.L., and W.H.P. prepared figures.
ACKNOWLEDGMENTS
We thank the staff of the imaging and sequencing facility at the First Core Lab and the AM2 Pharmacological Studies of Drugs in Animal Models for Human Diseases, Disease Animal Research Center, National Taiwan University College of Medicine, for their technical assistance.
REFERENCES
- 1.Ayres JS, Trinidad NJ, Vance RE. Lethal inflammasome activation by a multidrug-resistant pathobiont upon antibiotic disruption of the microbiota. Nat Med 18: 799–806, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Balamurugan R, Magne F, Balakrishnan D, Suau A, Ramani S, Kang G, Ramakrishna BS. Faecal bifidobacteria in Indian neonates and the effect of asymptomatic rotavirus infection during the first month of life. Indian J Med Res 132: 721–727, 2010 [PMC free article] [PubMed] [Google Scholar]
- 3.Bucher BT, McDuffie LA, Shaikh N, Tarr PI, Warner BB, Hamvas A, White FV, Erwin CR, Warner BW. Bacterial DNA content in the intestinal wall from infants with necrotizing enterocolitis. J Pediatr Surg 46: 1029–1033, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chen TL, Chen S, Wu HW, Lee TC, Lu YZ, Wu LL, Ni YH, Sun CH, Yu WH, Buret AG, Yu LC. Persistent gut barrier damage and commensal bacterial influx following eradication of Giardia infection in mice. Gut Pathog 5: 26, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Choi AH, Basu M, McNeal MM, Bean JA, Clements JD, Ward RL. Intranasal administration of an Escherichia coli-expressed codon-optimized rotavirus VP6 protein induces protection in mice. Prot Expr Purif 38: 205–216, 2004 [DOI] [PubMed] [Google Scholar]
- 6.Clark E, Hoare C, Tanianis-Hughes J, Carlson GL, Warhurst G. Interferon gamma induces translocation of commensal Escherichia coli across gut epithelial cells via a lipid raft-mediated process. Gastroenterology 128: 1258–1267, 2005 [DOI] [PubMed] [Google Scholar]
- 7.Clark EC, Patel SD, Chadwick PR, Warhurst G, Curry A, Carlson GL. Glutamine deprivation facilitates tumour necrosis factor induced bacterial translocation in Caco-2 cells by depletion of enterocyte fuel substrate. Gut 52: 224–230, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Clayburgh DR, Barrett TA, Tang Y, Meddings JB, Van Eldik LJ, Watterson DM, Clarke LL, Mrsny RJ, Turner JR. Epithelial myosin light chain kinase-dependent barrier dysfunction mediates T cell activation-induced diarrhea in vivo. J Clin Invest 115: 2702–2715, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Croswell A, Amir E, Teggatz P, Barman M, Salzman NH. Prolonged impact of antibiotics on intestinal microbial ecology and susceptibility to enteric Salmonella infection. Infect Immun 77: 2741–2753, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Daegelen P, Studier FW, Lenski RE, Cure S, Kim JF. Tracing ancestors and relatives of Escherichia coli B, and the derivation of B strains REL606 and BL21(DE3). J Mol Biol 394: 634–643, 2009 [DOI] [PubMed] [Google Scholar]
- 11.Engel DR, Koscielny A, Wehner S, Maurer J, Schiwon M, Franken L, Schumak B, Limmer A, Sparwasser T, Hirner A, Knolle PA, Kalff JC, Kurts C. T helper type 1 memory cells disseminate postoperative ileus over the entire intestinal tract. Nat Med 16: 1407–1413, 2010 [DOI] [PubMed] [Google Scholar]
- 12.Forsberg G, Fahlgren A, Hörstedt P, Hammarström S, Hernell O, Hammarström ML. Presence of bacteria and innate immunity of intestinal epithelium in childhood celiac disease. Am J Gastroenterol 99: 894–904, 2004 [DOI] [PubMed] [Google Scholar]
- 13.Frank DN, St Amand AL, Feldman RA, Boedeker EC, Harpaz N, Pace NR. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci USA 104: 13780–13785, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gareau MG, Jury J, MacQueen G, Sherman PM, Perdue MH. Probiotic treatment of rat pups normalises corticosterone release and ameliorates colonic dysfunction induced by maternal separation. Gut 56: 1522–1528, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Garner CD, Antonopoulos DA, Wagner B, Duhamel GE, Keresztes I, Ross DA, Young VB, Altier C. Perturbation of the small intestine microbial ecology by streptomycin alters pathology in a Salmonella enterica serovar typhimurium murine model of infection. Infect Immun 77: 2691–2702, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hill DA, Hoffmann C, Abt MC, Du Y, Kobuley D, Kirn TJ, Bushman FD, Artis D. Metagenomic analyses reveal antibiotic-induced temporal and spatial changes in intestinal microbiota with associated alterations in immune cell homeostasis. Mucosal Immunol 3: 148–158, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Huang CY, Hsiao JK, Lu YZ, Lee CT, Yu LCH. Anti-apoptotic PI3K/Akt signaling by sodium/glucose transporter 1 reduces epithelial barrier damage and bacterial translocation in intestinal ischemia. Lab Invest 91: 294–309, 2011 [DOI] [PubMed] [Google Scholar]
- 18.Huang CY, Kuo WT, Huang YC, Lee TC, Yu LC. Resistance to hypoxia-induced necroptosis is conferred by glycolytic pyruvate scavenging of mitochondrial superoxide in colorectal cancer cells. Cell Death Dis 4: e622, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kalischuk LD, Inglis GD, Buret AG. Campylobacter jejuni induces transcellular translocation of commensal bacteria via lipid rafts. Gut Pathog 1: 2, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kesik M, Saczynska V, Szewczyk B, Plucienniczak A. Inclusion bodies from recombinant bacteria as a novel system for delivery of vaccine antigen by the oral route. Immunol Lett 91: 197–204, 2004 [DOI] [PubMed] [Google Scholar]
- 21.Kinnebrew MA, Ubeda C, Zenewicz LA, Smith N, Flavell RA, Pamer EG. Bacterial flagellin stimulates Toll-like receptor 5-dependent defense against vancomycin-resistant Enterococcus infection. J Infect Dis 201: 534–543, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kleessen B, Kroesen AJ, Buhr HJ, Blaut M. Mucosal and invading bacteria in patients with inflammatory bowel disease compared with controls. Scand J Gastroenterol 37: 10034–10041, 2002 [DOI] [PubMed] [Google Scholar]
- 23.Kosovac K, Brenmoehl J, Holler E, Falk W, Schoelmerich J, Hausmann M, Rogler G. Association of the NOD2 genotype with bacterial translocation via altered cell-cell contacts in Crohn's disease patients. Inflamm Bowel Dis 16: 1311–1321, 2010 [DOI] [PubMed] [Google Scholar]
- 24.Kozakova H, Kolinska J, Lojda Z, Rehakova Z, Sinkora J, Zakostelecka M, Splichal I, Tlaskalova-Hogenova H. Effect of bacterial monoassociation on brush-border enzyme activities in ex-germ-free piglets: comparison of commensal and pathogenic Escherichia coli strains. Microbes Infect 8: 2629–2639, 2006 [DOI] [PubMed] [Google Scholar]
- 25.Laudanno OM, Cesolari JA, Godoy A, Sutich E, Sarangone S, Catalano J, San MP. Bioflora probiotic in immunomodulation and prophylaxis of intestinal bacterial translocation in rats. Dig Dis Sci 53: 2667–2670, 2008 [DOI] [PubMed] [Google Scholar]
- 26.Leatham MP, Banerjee S, Autieri SM, Mercado-Lubo R, Conway T, Cohen PS. Precolonized human commensal Escherichia coli strains serve as a barrier to E. coli O157:H7 growth in the streptomycin-treated mouse intestine. Infect Immun 77: 2876–2886, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lewis K, Lutgendorff F, Phan V, Soderholm JD, Sherman PM, McKay DM. Enhanced translocation of bacteria across metabolically stressed epithelia is reduced by butyrate. Inflamm Bowel Dis 16: 1138–1148, 2010 [DOI] [PubMed] [Google Scholar]
- 28.Lu YZ, Wu CC, Huang YC, Huang CY, Yang CY, Lee TC, Chen CF, Yu LC. Neutrophil priming by hypoxic preconditioning protects against epithelial barrier damage and enteric bacterial translocation in intestinal ischemia/reperfusion. Lab Invest 92: 783–796, 2012 [DOI] [PubMed] [Google Scholar]
- 29.Maragakis LL. Recognition and prevention of multidrug-resistant Gram-negative bacteria in the intensive care unit. Crit Care Med 38: S345–351, 2010 [DOI] [PubMed] [Google Scholar]
- 30.Michalopoulos A, Falagas ME, Karatza DC, Alexandropoulou P, Papadakis E, Gregorakos L, Chalevelakis G, Pappas G. Epidemiologic, clinical characteristics, and risk factors for adverse outcome in multiresistant Gram-negative primary bacteremia of critically ill patients. Am J Infect Control 39: 396–400, 2011 [DOI] [PubMed] [Google Scholar]
- 31.Miyauchi E, Morita H, Tanabe S. Lactobacillus rhamnosus alleviates intestinal barrier dysfunction in part by increasing expression of zonula occludens-1 and myosin light-chain kinase in vivo. J Dairy Sci 92: 2400–2408, 2009 [DOI] [PubMed] [Google Scholar]
- 32.Mizoguchi E. Chitinase 3-like-1 exacerbates intestinal inflammation by enhancing bacterial adhesion and invasion in colonic epithelial cells. Gastroenterology 130: 398–411, 2006 [DOI] [PubMed] [Google Scholar]
- 33.Moles L, Gomez M, Heilig H, Bustos G, Fuentes S, de Vos W, Fernandez L, Rodriguez JM, Jimenez E. Bacterial diversity in meconium of preterm neonates and evolution of their fecal microbiota during the first month of life. PLoS One 8: e66986, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nasa P, Juneja D, Singh O, Dang R, Singh A. An observational study on bloodstream extended-spectrum beta-lactamase infection in critical care unit: incidence, risk factors and its impact on outcome. Eur J Intern Med 23: 192–195, 2012 [DOI] [PubMed] [Google Scholar]
- 35.Perez F, Pultz MJ, Endimiani A, Bonomo RA, Donskey CJ. Effect of antibiotic treatment on establishment and elimination of intestinal colonization by KPC-producing Klebsiella pneumoniae in mice. Antimicrob Agents Chemother 55: 2585–2589, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S, Medzhitov R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 118: 229–241, 2004 [DOI] [PubMed] [Google Scholar]
- 37.Reddy BS, MacFie J, Gatt M, Macfarlane-Smith L, Bitzopoulou K, Snelling AM. Commensal bacteria do translocate across the intestinal barrier in surgical patients. Clin Nutr 26: 208–215, 2007 [DOI] [PubMed] [Google Scholar]
- 38.Rosenberg AL, Seneff MG, Atiyeh L, Wagner R, Bojanowski L, Zimmerman JE. The importance of bacterial sepsis in intensive care unit patients with acquired immunodeficiency syndrome: implications for future care in the age of increasing antiretroviral resistance. Crit Care Med 29: 548–556, 2001 [DOI] [PubMed] [Google Scholar]
- 39.Samel S, Keese M, Kleczka M, Lanig S, Gretz N, Hafner M, Sturm J, Post S. Microscopy of bacterial translocation during small bowel obstruction and ischemia in vivo—a new animal model. BMC Surgery 2: 6, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Schjorring S, Struve C, Krogfelt KA. Transfer of antimicrobial resistance plasmids from Klebsiella pneumoniae to Escherichia coli in the mouse intestine. J Antimicrob Chemother 62: 1086–1093, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sekirov I, Tam NM, Jogova M, Robertson ML, Li Y, Lupp C, Finlay BB. Antibiotic-induced perturbations of the intestinal microbiota alter host susceptibility to enteric infection. Infect Immun 76: 4726–4736, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Shirkey TW, Siggers RH, Goldade BG, Marshall JK, Drew MD, Laarveld B, Van Kessel AG. Effects of commensal bacteria on intestinal morphology and expression of proinflammatory cytokines in the gnotobiotic pig. Exp Biol Med (Maywood) 231: 1333–1345, 2006 [DOI] [PubMed] [Google Scholar]
- 43.Sjöberg V, Sandström O, Hedberg M, Hammarström S, Hernell O, Hammarström ML. Intestinal T-cell responses in celiac disease—impact of celiac disease associated bacteria. PLoS One 8: e53414, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Soderholm JD, Yang PC, Ceponis P, Vohra A, Riddell R, Sherman PM, Perdue MH. Chronic stress induces mast cell-dependent bacterial adherence and initiates mucosal inflammation in rat intestine. Gastroenterology 123: 1099–1108, 2002 [DOI] [PubMed] [Google Scholar]
- 45.Swiatczak B, Rescigno M. How the interplay between antigen presenting cells and microbiota tunes host immune responses in the gut. Semin Immunol 24: 43–49, 2012 [DOI] [PubMed] [Google Scholar]
- 46.Swidsinski A, Ladhoff A, Pernthaler A, Swidsinski S, Loening-Baucke V, Ortner M, Weber J, Hoffmann U, Schreiber S, Dietel M, Lochs H. Mucosal flora in inflammatory bowel disease. Gastroenterology 122: 44–54, 2002 [DOI] [PubMed] [Google Scholar]
- 47.Ubeda C, Taur Y, Jenq RR, Equinda MJ, Son T, Samstein M, Viale A, Socci ND, van den Brink MR, Kamboj M, Pamer EG. Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J Clin Invest 120: 4332–4341, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ukena SN, Singh A, Dringenberg U, Engelhardt R, Seidler U, Hansen W, Bleich A, Bruder D, Franzke A, Rogler G, Suerbaum S, Buer J, Gunzer F, Westendorf AM. Probiotic Escherichia coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity. PLoS One 2: e1308, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wells CL, VandeWesterlo EM, Jechorek RP, Erlandsen SL. Effect of hypoxia on enterocyte endocytosis of enteric bacteria. Crit Care Med 24: 985–991, 1996 [DOI] [PubMed] [Google Scholar]
- 50.Wu LL, Chiu HD, Peng WH, Lin BR, Lu KS, Lu YZ, Yu LCH. Epithelial inducible nitric oxide synthase causes bacterial translocation by impairment of enterocytic tight junctions via intracellular signals of Rho-associated kinase and protein kinase C zeta. Crit Care Med 39: 2087–2098, 2011 [DOI] [PubMed] [Google Scholar]
- 51.Wu LL, Peng WH, Kuo WT, Huang CY, Ni YH, Lu KS, Turner JR, Yu LC. Commensal bacterial endocytosis in epithelial cells is dependent on myosin light chain kinase-activated brush border fanning by interferon-gamma. Am J Pathol 184: 2260–2274, 2014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yu LC, Wang JT, Wei SC, Ni YH. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World J Gastrointest Pathophysiol 3: 27–43, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zareie M, Johnson-Henry K, Jury J, Yang PC, Ngan BY, McKay DM, Soderholm JD, Perdue MH, Sherman PM. Probiotics prevent bacterial translocation and improve intestinal barrier function in rats following chronic psychological stress. Gut 55: 1553–1560, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]