

Abstract
We isolated the complete Foxl2 (Foxl2a) cDNA from the Monopterus albus ovary. An alignment of known Foxl2 amino-acid sequences confirmed the conservation of the Foxl2 open reading frame, especially the forkhead domain and C-terminal region. The expression of Foxl2 was detected in the brain, eyes, and gonads. A high level of Foxl2 expression in the ovary before sex reversal, but its transcripts decreased sharply when the gonad developed into the ovotestis and testis. The correlation between the Foxl2 expression and the process of sex development revealed the important function of Foxl2 during the sex reversal of M. albus. Immunohistochemical analysis showed that Foxl2 was expressed abundantly in granulosa cells and in the interstitial cells of the ovotestis and testis. These results suggest that Foxl2 plays a pivotal role in the development and maintenance of ovarian function. Foxl2 may be also involved in the early development of testis and the development of ocular structures of M. albus.
Forkhead transcription factors play a critical role in the regulation of cellular proliferation and differentiation1.They are also involved in several other biological processes, including tissue development, establishment of the body axis, metabolic processes, and the determination and differentiation of cell types2,3. In many cases, forkhead transcription factors are also responsible for various processes during embryonic development and adult metabolism2.
Forkhead box L2 (Foxl2) is a member of the forkhead family of transcription factors, characterized by a conserved 100-amino-acid domain called “forkhead box”4. Although the Foxl2 is highly conserved across divergent taxonomic groups5, the C-terminal region is more conserved than the N-terminal region outside the DNA-binding forkhead domain6. Mammalian Foxl2 contains a polyalanine tract and other low-complexity repeats absent from the fish sequences5. Foxl2 mutation in human leads to the blepharophimosis ptosis epicanthus inversus syndrome (BPES), which is characterized by eyelid malformations and premature ovarian failure (POF)7. Foxl2 was observed in the developing eyelids and perioptic mesenchyme of mouse7. It is reported that males and females of mouse lacking Foxl2 are small and show distinctive craniofacial morphology with upper eyelids absent8. Mouse Foxl2 has been also detected in the pituitary9 and developing ovary10. Recently, many studies demonstrated that Foxl2 is a putative transcription factor in the early development of the female vertebrate gonad and is involved in adult ovarian function6,11,12. In genetic program, somatic testis determination was activated in an XX gonads mouse lacking Foxl2 from meiotic prophase oocytes, implying the pivotal function of Foxl2 to repress the male gene pathway at several stages of female gonadal differentiation13. Foxl2 is also involved in the differentiation of granulosa cells of gonad and the maintenance of ovarian function, as well as the transcriptional regulation of other genes during gonadal differentiation in fish14,15,16.
Estrogens have important roles in sexual differentiation and sex changes in fish17,18. Aromatase, encoded by cyp19a1a, is responsible for 17β-estradiol (E2) synthesis by catalyzing the conversion of androgens to estrogens19. Aromatase and Foxl2 co-localize in the adult ovaries of medaka (Oryzias latipes)14 and Japanese flounder (Paralichthys olivaceus)16. The expression profile of Foxl2 also correlates strongly with aromatase activities in the gonads during the sexual differentiation of the chicken (Gallus gallus)20. These results suggest that Foxl2 is related to the sexual differentiation of fish and other vertebrates through the transcriptional regulation of the cyp19a1a gene.
Rice-field eel Monopterus albus, which belongs to the family Synbranchidae in the order Synbranchiformes (Neoteleostei, Teleostei, Vertebrata), is a hermaphroditic protogynous freshwater fish species that undergoes sexual reversal from a functional female to a male21. Some genes related to sex determination and differentiation have been identified in M. albus, including cyp19a1a22, Sox923, Dmrt124, and Jnk125. The complete genomic sequences of Foxl2 is available for the pufferfish (Takifugu rubripes), green spotted puffer (Tetraodon nigroviridis), and zebrafish (Danio rerio)15. Given the detections of Foxl2 transcripts in ovary, it is probably involved in ovarian development in mammal7,10, birds20, and teleost14,15,16. Although there are many studies which investigated the roles of other sex-related genes in M. albus, few studies of Foxl2 function have been reported in this hermaphrodite fish species. To investigate whether Foxl2 is involved in ovarian differentiation and development in M. albus, as in other vertebrates, we isolated the full-length Foxl2 cDNA from M. albus, and analyzed its expression pattern and protein localization in the gonads during sex reversal using real-time RT–PCR and immunohistochemistry.
Results
Isolation and characterization of M. albus Foxl2
The M. albus Foxl2 (Foxl2a, KC823043) cDNA is 2037 bp long, which comprises a 263-bp 5′-untranslated region (5′-UTR), an 853-bp 3′-untranslated region (3′-UTR) containing two typical poly(A) signals, and an open reading frame (ORF) (921 bp). The ORF encodes a putative 306-amino-acid containing the 100-amino-acid conserved sequence of the forkhead family (Fig. 1). The similarity of the M. albus Foxl2 nucleotide sequence to those of other vertebrates was determined with BLASTN at the National Center for Biotechnology Information (NCBI) web server (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The result exhibited high levels of identity, especially with the teleosts, including the honeycomb grouper (Epinephelus merra, 86%), spotted butterfish (Scatophagus argus, 85%), three-spot wrasse (Halichoeres trimaculatus, 82%), and peacock bass (Cichla monoculus, 82%). A high degree of similarity, E. merra (95.1%), Nile tilapia (Oreochromis niloticus, 94.4%), and H. trimaculatus (94.1%), was also apparent when the percentage identities of the corresponding amino-acid sequences were calculated with Clustal W (Fig. 2). Except the forkhead domain, which displayed almost complete conservation among these fish species, the C-terminal region of Foxl2 was more strongly conserved than the N-terminal region. Like the Foxl2 proteins of other fishes, there is no polyalanine tract or proline or glycine repeat in the M. albus (Fig. 3). A phylogenetic tree was constructed with 11 Foxl2 nucleotide sequences, which clustered into two separate branches when mammalian Foxl2 were used as the outgroup. M. albus Foxl2 clusters with E. merra displayed the close relationship between them, with 74 bootstrap support (Fig. 4).
Figure 1. Nucleotide and deduced amino-acid sequences of M. albus Foxl2.

The amino-acid sequence is shown with the standard one-letter code below the nucleotide sequence. The positions of the amino-acid residues are given in parentheses. The two typical polyadenylation signals are boxed.
Figure 2. Alignment of the Foxl2 amino-acid sequences of 11 species.
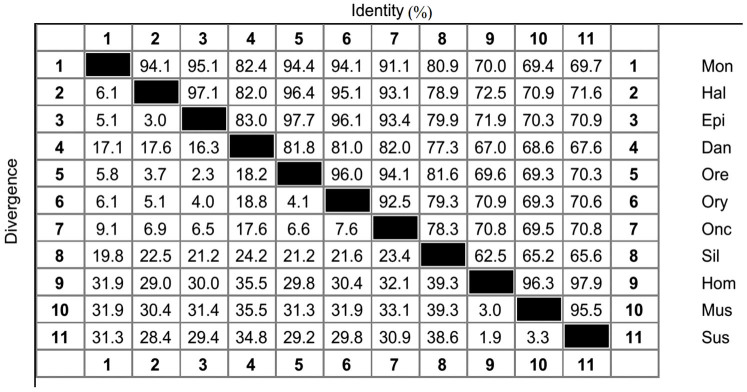
Mon, Monopterus albus; Hal, Halichoeres trimaculatus, BAJ15129.1; Epi, Epinephelus merra, ACD62374.1; Dan: Danio rerio, XP_698915.1; Ore, Oreochromis niloticus, AAT36328.1; Ory, Oryzias latipes, BAF42653.1; Onc, Oncorhynchus mykiss, AAS87040.2; Sil, Silurus meridionalis, ABK76309.1; Hom, Homo sapiens, AAK01352.1; Mus, Mus musculus, AAN04088.1; Sus, Sus scrofa, AAQ91845.1.
Figure 3. Comparison of Foxl2 amino-acid sequence of M. albus with known orthologues.

Abbreviations of the species are the same as in the legend to Fig. 2. The filled bar indicates the forkhead domain. The glycine-rich repeats (G), proline repeats (P), and polyalanine tracts (A) are boxed.
Figure 4. Phylogenetic tree based on the Foxl2 nucleotide sequences of 11 species.
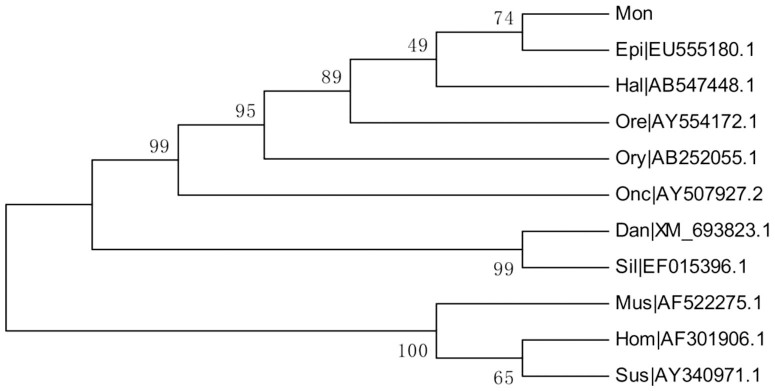
Abbreviations of the species are the same as in the legend to Fig. 2. The numbers at each branch represent the bootstrap values obtained with 1000 replicates.
Tissue distribution of Foxl2 expression
RT–PCR showed that Foxl2 was mainly expressed in the gonads. The level in the ovary was much higher than that in the testis and ovotestis, and there was no obvious difference between the testis and ovotestis. High levels of Foxl2 expression were also observed in the whole eyes and brain. However, the expression of Foxl2 was barely detected in the other tested tissues (Fig. 5).
Figure 5. RT–PCR analysis of Foxl2 mRNA in various tissues of M. albus.
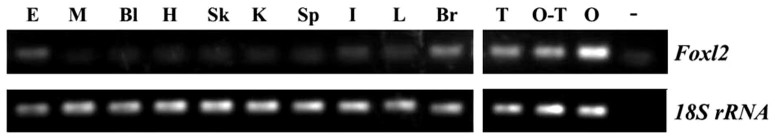
E, eye; M, muscle; Bl, blood; H, heart; Sk, skin; K, kidney; Sp, spleen; I, intestine; L, liver; Br, brain; T, testis; O-T, ovotestis; O, ovary; −, negative control; 18S rRNA, internal control. The figure was cropped, for uncropped figure, see figure S1 &S2, supplementary file.
Expression profile of Foxl2 during sex reversal of the gonads
To investigate the relationship between Foxl2 expression and sex reversal in M. albus, we analyzed the expression of Foxl2 in the seven different developmental stages of gonads using real-time RT–PCR. The highest level of Foxl2 was detected in the ovaries, especially in the fourth-stage ovaries (♀IV). However, its expression decreased sharply in the first-stage ovotestis (♀/♂I), and decreased continuously in the second- and third-stage ovotestis (♀/♂II and ♀/♂III), as well as the testis (Fig. 6).
Figure 6. Copies of Foxl2 transcripts in different phases of M. albus gonadal development.
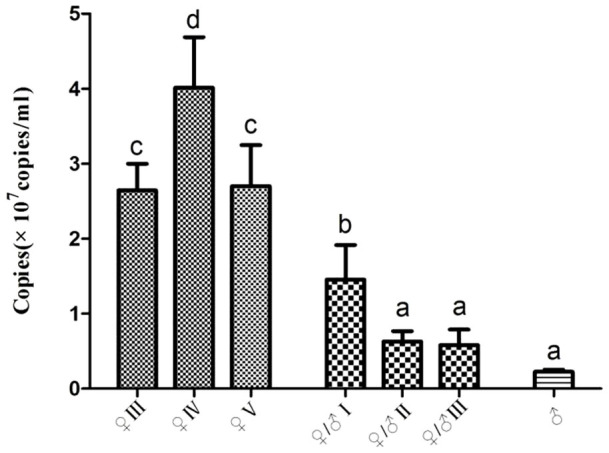
♀, ovaries; ♀/♂, ovotestis; ♂, testis.
Localization of Foxl2 protein in the gonads
In the ovary, Foxl2 immunoreactivity was detected abundantly in the granulosa cells around the oocytes and immature oocytes, but not in the mature oocytes (Fig. 7A, B). In the ovotestis, positive signals were observed in each type of cell, especially in the immature oocytes and interstitial cells (Fig. 7C, D). Foxl2 expression was also detected in the interstitial cells in testes, but not in sperm (Fig. 7E, F). No positive signals were observed in the negative control (Fig. 7G–I).
Figure 7. Immunohistochemical analysis of Foxl2 in different phases of M. albus gonadal development.

(A–F) Immunohistochemical analysis of Foxl2 in the (A and B) ovaries, (C and D) ovotestis, and (E and F) testis. (B), (D), and (F) are enlarged areas of (A), (C), and (E), respectively. The positive antigen was dyed brown with 3′,3′-diaminobenzidine (DAB) (arrows). (G–I) Negative controls.
Discussion
Two Foxl2 paralogs which are named Foxl2a (Foxl2) and Foxl2b (Foxl3) have been reported in some teleost species5,26. They are also identified by blast against the available fish genome database, which is in agreement with the genome duplication event in fish27. Foxl2a and Foxl2b in rainbow trout (Oncorhynchus mykiss) were expressed specifically in the ovary, but displayed different temporal expression patterns5. However, the mRNA levels of Foxl2 in ovary and Foxl3 in testis point to a strong sexual dimorphism and vary significantly during the reproductive cycle in European sea bass (Dicentrarchus labrax)26. Unlike above species, only Foxl2 was cloned in the ovary of M. albus in this study. The same phenomena have been reported in some fish species14,15,28. Due to the complexity and shortage of date in expression pattern of fish Foxl2b (Foxl3), a systematic investigation on Foxl3 need to be carried out.
The C-terminal region and the forkhead domain of Foxl2 are strongly conserved among different vertebrates. The functions of these two regions may have been conserved throughout evolution, whereas the N-terminal region has evolved under weaker conservation pressure29. The forkhead domain of Foxl2 is responsible for the nuclear import of this protein, and contains a putative nuclear localization signal (NLS, typically RRRRRMKR) at C-terminus, like other FOX proteins6,30,31. Homopolymeric runs of amino acids, such as A, G, and P, were present in the mammalian proteins but not in those of non-mammalian vertebrates. A phylogenetic analysis supported the strong evolutionary conservation of Foxl2 in fish and the close genetic relationship between M. albus and E. merra.
Foxl2 was mainly expressed in the brain, eyes, and gonads of M. albus, with the highest level of expression in the ovary. The gonadotropin-releasing hormone receptor gene (Gnrhr), which contains a site bound by Foxl2, AP-1, and Smads, can be activated by Foxl2 in mouse11. The high level of Foxl2 transcript was found in the M. albus brain, indicating that the Gnrhr may be regulated by Foxl2 on the transcriptional level. Fish Foxl2 is involved in the hypothalamus-pituitary-gonadal axis due to the detection of Foxl2 expression in the hypothalamus, pituitary, and gonad15,32. In M. albus, the transcription of Foxl2 was much higher in gonad than that in brain. It is implied that Foxl2 probably executes its functions via the transcriptional regulation of the gonadotropin-releasing hormone-gonadotropin-sex steroid pathway.
The mutation of Foxl2 leads to BPES, characterized by malformations of the eyelid and premature ovarian failure in human7. Compared to BPES, mouse lacking Foxl2 were born with open and necrotizing eyes, coupled with severe eyelid hypoplasia8. Foxl2 expression was concentrated in the perioptic mesenchyme7. Lower levels of Foxl2 expression were also detected in the lens fibers of the eyes7. Consequently, such expression pattern of Foxl2 could be required for the eyelid formation and the development of other ocular structures in mouse7. Foxl2 expression in dogfish (Scyliorhinus canicula) was firstly detected in the mesenchyme around the eyes and then restricted to the underlying mesenchyme at the outer edges of the developing eyelids33. As in other fish27,32,34, Foxl2 was highly expressed in the eyes of M. albus. This expressive pattern manifests the conserved functions of Foxl2 among different species. It is implied that Foxl2 is probably involved in the development of other ocular structures because of the apparent absence of eyelids from most fish.
In general, Foxl2 is used to be considered as a good marker of ovarian differentiation because of its high-level expression in the ovary reported in several vertebrates29. Although Foxl2 expression has been detected in the adult testis of several species, the levels of Foxl2 in the testis were all significantly lower than that in the ovary in G. gallus20, the southern catfish (Silurus meridionalis)35, Oncorhynchus mykiss5, and frog (Rana rugosa)36.On the contrary, a study in protogynous H. trimaculatus demonstrated that Foxl2 expression has no sexual dimorphism between the testis and ovary32. Foxl2 expression was also detected in testis of M. albus, though it was lower in the testis than that in the ovary. Thus, whether Foxl2 is a good marker of ovarian differentiation in sex reversal species or not should be deliberated.
The highest levels of Foxl2 expression were observed in the ovary before sex reversal in M. albus. Foxl2 transcript decreased sharply when individuals developed to the intersex stage I (♀/♂I). Foxl2 expression also declined dramatically as the oocytes continued to degenerate, paralleling with the initiation of germ-cell proliferation into spermatogonia (♀/♂II and ♀/♂III). Then, Foxl2 expression was maintained at a low level in the testis. The correlation between the Foxl2 expression pattern and the process of sex development reveals the crucial function of Foxl2 during the sex reversal of M. albus. Foxl2 was highly expressed in brain and ovary during pre-spawning phase implying the important role for this correlate in ovarian recrudescence in catfish (Clarias gariepinus)37. As the same, high expression of Foxl2 was also observed in brain and ovary of M. albus, especially in fourth-stage ovaries (♀IV). This result suggests that Foxl2 is involved in the ovarian development and maturity. Foxl2 mRNA in the ovary is restricted to the granulosa (follicular) cells surrounding the oocytes in Oreochromis niloticus15 and O. latipes14. However, no signals have been observed in the oocytes, as the situation in mammals29,38. In this study, an immunohistochemical analysis showed that the Foxl2 protein was observed abundantly in the granulosa cells around the oocytes and immature oocytes, but not in the mature oocytes. This conserved expression patterns suggest that Foxl2 has a basic function in the differentiation of granulosa cells and a crucial role in the maintenance of oocytes. Lower levels of Foxl2 were detected in the developing testis of mouse from 14.5 dpc (day post conception) onward and in the developing epididymis at later stages by in situ hybridization12. In the hermaphrodite H. trimaculatus32, Foxl2 protein was concentrated in the interstitial cells, including in the tubules and Leydig cells of the testis. A small amount of Foxl2 mRNA was expressed in testis, and its protein was also observed in the interstitial cells of the ovotestis and testis of M. albus. It is inferred that the function of Foxl2 in the testis of sex-reversal species could be quite different from other gonochorous teleost. Foxl2 might be also involved in the early development and maintenance of testis.
It has been reported that Foxl2 regulates the expression of aromatase, the product of cyp19a1a39,40, which is the key enzyme in the synthesis of estrogen and important for sexual differentiation in fish17,41. Foxl2 in G. gallus is involved in the regulation of aromatase transcription during early sexual differentiation20,42. Foxl2 and aromatase in Oreochromis niloticus43 and O. latipes14 are co-expressed in some somatic cells located on the ventral side of the XX gonads. This data suggest the important role of Foxl2 in early ovarian differentiation by activating cyp19a1a transcription. Earlier studies of M. albus demonstrated that cyp19a1a was expressed specifically in the brain and gonads, and then its expression declined significantly with the gonadal development22,44. In this study, M. albus Foxl2 was expressed in the brain and gonads. It was also co-expressed with cyp19a1a during gonadal development. Taken together, the correlation expression pattern between Foxl2 and cyp19a1a in ovary and brain indicates that Foxl2 may play an important role in ovarian development.
In conclusion, Foxl2 expression in the gonads is in line with the process of sex development, revealing the important function of Foxl2 during the sex reversal of M. albus. Foxl2 protein and Foxl2 mRNA were detected in the ovaries, ovotestis and testis. Foxl2 was primarily located in granulosa cells, immature oocytes, and interstitial cells. The Foxl2 mRNA was also observed in brain. These results illustrate that Foxl2 is involved in the early development and maintenance of both ovary and testis. In addition, the detection of Foxl2 expression in eyes suggests the possible function of Foxl2 in the development of ocular structures of M. albus.
Methods
Fish
Wild M. albus were purchased from markets in Wuhan, China. The length of the individuals varied from 34 to 54 cm and their bodyweight ranged from 50 to 120 g. They were kept in fresh water for one week before processing. A small segment of the gonads was fixed in Holland–Bouin's fluid for histological assessment of the sexual status and immunohistochemical analysis. The remainder was frozen immediately in liquid nitrogen and then stored at −80°C. All experiments were performed with the approval from the Institutional Animal Care and Use Committee of Huazhong Agricultural University (Wuhan, China), strictly according to the guidelines set by this committee for the treatment of animals. The experiments were also performed in accordance with the International Guiding Principles for Biomedical Research Involving Animals, promulgated by the Society for the Study of Reproduction.
Gonadal histology
The fixed gonads were dehydrated in a graded series of ethanol and then embedded in paraffin. Sections (5–8 μm thick) were cut and stained with hematoxylin and eosin. The sexual phase of each fish was confirmed by observation under a light microscope45.
RNA extraction
After the developmental phase of the gonads was determined following our previous study46, total RNA was extracted from the gonads (two samples were selected and RNA was extracted in each phase) and from the blood, muscle, skin, liver, eye (whole eye), spleen, intestines, kidney, heart, and brain (whole brain), according to the RNAiso Plus manufacturer's recommendations (Takara, Dalian, China). First-strand cDNA was synthesized from 1 μg of total RNA in a 20 μL reaction volume following the manufacturer's recommendations of PrimeScript® RT Reagent Kit with gDNA Eraser (Perfect Real Time) (Takara).
Molecular cloning of Foxl2
To clone Foxl2 (Foxl2a) from M. albus, a pair of degenerate primers was designed from the conserved regions of Foxl2 in other fish species. PCR was performed in a 25 μL reaction volume containing: 2 μL of ovarian cDNA as template, 0.5 μL of each 10 mM primer (see Table S1, supplementary file), 12.5 μL of Premix Taq DNA polymerase (Takara), and double-distilled water to a final volume of 25 μL. The 5′ and 3′ ends of the Foxl2 cDNA were obtained according to the manufacturer's instructions of the SMART RACE Kit (Clontech, USA). Four gene-specific primers (see Table S1, supplementary file) were designed for the rapid amplification of cDNA ends (RACE). The PCR products were subjected to electrophoresis in 1% (w/v) agarose gel and purified using the TIANquick Midi Purification Kit (Tiangen, China). The purified product was recovered and cloned into the PMD-19T vector (Takara), and then sequenced at the Beijing Genomics Institute.
Sequence and phylogenetic analyses
Nucleotide sequences were identified with BLASTN at the NCBI web server. We deduced the amino-acid sequence using the BioEdit software and calculated the percentage identity with the MegAlign program. Clustal W was used to construct a multiple alignment of amino-acid sequences. The phylogenetic trees were constructed using the neighbor-joining method in MEGA version 5.05. The credibility of each branch was supported by the bootstrap scores (1000 replicates). All of the sequences used in multiple alignment and phylogenetic analyses were obtained from GenBank.
Tissue distribution of Foxl2 expression examined with RT–PCR
To identify the expression pattern of Foxl2 in various M. albus tissues, PCR amplification was performed with the gene-specific primers (see Table S1, supplementary file), which were designed based on the nucleotide sequence we cloned (see above). The PCR protocol was: preheating at 95°C for 3 min, followed by 35 cycles at 95°C for 20 s, 60°C for 20 s, and 72°C for 20 s, with a final extension at 72°C for 10 min. The mixture contained 2 μL of 10-fold-diluted cDNA in 25 μL PCR reaction volume (as described above). 18s rRNA (EU120033.1) was amplified with 18 PCR cycles as the internal control to calibrate the expression of Foxl2. All PCR products were subjected to electrophoresis in 1% (w/v) agarose gel and the bands were visualized by staining with GelRed™.
Foxl2 expression pattern during sex reversal of the gonads examined with real-time RT–PCR
Absolute quantitative real-time RT–PCR experiments were performed in a final volume of 25 μL containing 2 μL of cDNA, 0.5 μL of each 10 mM primer, and 12.5 μL of SYBR® Premix Ex Taq™ II (Perfect Real Time, Takara). The protocol was: 30 s at 95°C, followed by 30 cycles of 95°C for 5 s, 60°C for 45 s, and 72°C for 30 s. The samples were analyzed in triplicate and the fluorescence released from the dye which was monitored by Rotor-Gene Q. A negative control was containing each assay but cDNA. The standard curve was constructed by the serially diluted plasmid containing the target gene. The expression of Foxl2 in the samples was calculated from the standard curve and expressed as copies/ml. Differences of gene expressions in different developmental stages of the gonads were determined with one-way analysis of variance (ANOVA) following by Tukey's HSD test. The differences were deemed statistically significant at P < 0.05.
Localization of Foxl2 protein in the gonads with immunohistochemistry
An anti-Foxl2 polyclonal antibody (PA1-802, Thermo, USA) was used to determine the cellular localization of Foxl2 protein in the M. albus gonads. Sections were prepared as described above. After paraffin removal and dehydration, the sections were washed with citric acid buffer (0.1 M citric acid and sodium citrate, pH 6.0), incubated in 3% (v/v) H2O2 and 10% (v/v) normal goat serum to block nonspecific binding, and then incubated overnight at 4°C with the primary antibody (diluted 1:100), which is a synthetic peptide corresponding to residues M(1)MASYPEPEDTAGT(14) of mouse FOXL2 and the host is rabbit. After incubation with the secondary antibody which is labeled by HRP (anti-mouse/rabbit, Maixin KIT-9901), the sections were exposed to 3′,3′-diaminobenzidine (DAB) and stained with hematoxylin to visualize the nuclei in the gonadal tissues, and then observed under a fluorescence microscope (Eclipse H600L, Nikon). As the negative control, the sections were treated in the same way but with Tris-buffered saline instead of the primary antibody.
Author Contributions
Q.H. wrote this manuscript text; R.T., D.L. and Q.H. designed the experiments; W.G. collected material for study; Q.H., W.G. and Y.G. carried out the experiments and analyzed the data. All authors reviewed the manuscript.
Supplementary Material
supplementary file
Acknowledgments
This work was supported by the National Department Public Benefit Research Foundation of China (201003076), the Fundamental Research Funds for the Central Universities (2013PY024), and the National Natural Foundation of China (Project no. 30970529). The authors thank Dr. Jiaoyan Yang, Wei Chi, and Ming Zou for their suggestion on the organization of this manuscript. The authors thank Shimin Lu and Zidong Liu for their technical assistances in this experiment.
References
- Cunningham M. A., Zhu Q., Unterman T. G. & Hammond J. M. Follicle-stimulating hormone promotes nuclear exclusion of the forkhead transcription factor FoxO1a via phosphatidylinositol 3-kinase in porcine granulosa cells. Endocrinology 144, 5585–5594 (2003). [DOI] [PubMed] [Google Scholar]
- Carlsson P. & Mahlapuu M. Forkhead transcription factors: key players in development and metabolism. Dev. Biol. 250, 1–23 (2002). [DOI] [PubMed] [Google Scholar]
- Uhlenhaut N. H. & Treier M. Foxl2 function in ovarian development. Mol. Genet. Metab. 88, 225–234 (2006). [DOI] [PubMed] [Google Scholar]
- Kaestner K. H., Knöchel W. & Martínez D. E. Unified nomenclature for the winged helix/forkhead transcription factors. Gene. Dev. 14, 142–146 (2000). [PubMed] [Google Scholar]
- Baron D. et al. An evolutionary and functional analysis of FoxL2 in rainbow trout gonad differentiation. J. Mol. Endocrinol. 33, 705–715 (2004). [DOI] [PubMed] [Google Scholar]
- Cocquet J. et al. Structure, evolution and expression of the FOXL2 transcription unit. Cytogenet. Genome Res. 101, 206–211 (2003). [DOI] [PubMed] [Google Scholar]
- Crisponi L. et al. The putative forkhead transcription factor FOXL2 is mutated in blepharophimosis/ptosis/epicanthus inversus syndrome. Nat. Genet. 27, 159–166 (2001). [DOI] [PubMed] [Google Scholar]
- Uda M. et al. Foxl2 disruption causes mouse ovarian failure by pervasive blockage of follicle development. Hum. Mol. Genet. 13, 1171–1181 (2004). [DOI] [PubMed] [Google Scholar]
- Ellsworth B. S. et al. FOXL2 in the pituitary: molecular, genetic, and developmental analysis. Mol. Endocrinol 20, 2796–2805 (2006). [DOI] [PubMed] [Google Scholar]
- Schmidt D. et al. The murine winged-helix transcription factor Foxl2 is required for granulosa cell differentiation and ovary maintenance. Development 131, 933–942 (2004). [DOI] [PubMed] [Google Scholar]
- Ellsworth B. S. et al. The gonadotropin releasing hormone (GnRH) receptor activating sequence (GRAS) is a composite regulatory element that interacts with multiple classes of transcription factors including Smads, AP-1 and a forkhead DNA binding protein. Mol. Cell. Endocrinol. 206, 93–111 (2003). [DOI] [PubMed] [Google Scholar]
- Loffler K. A., Zarkower D. & Koopman P. Etiology of ovarian failure in blepharophimosis ptosis epicanthus inversus syndrome: FOXL2 is a conserved, early-acting gene in vertebrate ovarian development. Endocrinology 144, 3237–3243 (2003). [DOI] [PubMed] [Google Scholar]
- Ottolenghi C. et al. Foxl2 is required for commitment to ovary differentiation. Hum. Mol. Genet. 14, 2053–2062 (2005). [DOI] [PubMed] [Google Scholar]
- Nakamoto M., Matsuda M., Wang D. S., Nagahama Y. & Shibata N. Molecular cloning and analysis of gonadal expression of Foxl2 in the medaka, Oryzias latipes. Biochem. Bioph. Res. Co. 344, 353–361 (2006). [DOI] [PubMed] [Google Scholar]
- Kobayashi T. & Nagahama Y. Molecular cloning and gene expression of Foxl2 in the Nile tilapia, Oreochromis niloticus. Biochem. Bioph. Res. Co. 320, 83–89 (2004). [DOI] [PubMed] [Google Scholar]
- Yamaguchi T., Yamaguchi S., Hirai T. & Kitano T. Follicle-stimulating hormone signaling and Foxl2 are involved in transcriptional regulation of aromatase gene during gonadal sex differentiation in Japanese flounder, Paralichthys olivaceus. Biochem. Bioph. Res. Co. 359, 935–940 (2007). [DOI] [PubMed] [Google Scholar]
- Guiguen Y., Fostier A., Piferrer F. & Chang C. F. Ovarian aromatase and estrogens: a pivotal role for gonadal sex differentiation and sex change in fish. Gen. Comp. Endocr. 165, 352–366 (2010). [DOI] [PubMed] [Google Scholar]
- Nakamura M., Bhandari R. K. & Higa M. The role estrogens play in sex differentiation and sex changes of fish. Fish Physiol. Biochem. 28, 113–117 (2003). [Google Scholar]
- Simpson E. R. et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 15, 342–355 (1994). [DOI] [PubMed] [Google Scholar]
- Govoroun M. S. et al. Isolation of chicken homolog of the FOXL2 gene and comparison of its expression patterns with those of aromatase during ovarian development. Dev. Dynam. 231, 859–870 (2004). [DOI] [PubMed] [Google Scholar]
- Liu C. Rudimentary hermaphroditism in the symbranchoid eel, Monopterus javanensis. Sinensia 15, 1–8 (1944). [Google Scholar]
- Liu J. F., Guiguen Y. & Liu S. J. Aromatase (P450arom) and 11β-hydroxylase (P45011β) genes are differentially expressed during the sex change process of the protogynous rice field eel, monopterus albus. Fish Physiol. Biochem. 35, 511–518 (2009). [DOI] [PubMed] [Google Scholar]
- Zhou R. et al. Similar gene structure of two Sox9a genes and their expression patterns during gonadal differentiation in a teleost fish, rice field eel (Monopterus albus). Mol. Reprod. Dev. 66, 211–217 (2003). [DOI] [PubMed] [Google Scholar]
- Huang X. et al. Multiple alternative splicing and differential expression of dmrt1 during gonad transformation of the rice field eel. Biol. Reprod. 73, 1017–1024 (2005). [DOI] [PubMed] [Google Scholar]
- Xiao Y. M. et al. Contrast expression patterns of JNK1 during sex reversal of the rice-field eel. J. Exp. Zool. Part B. 314, 242–256 (2010). [DOI] [PubMed] [Google Scholar]
- Crespo B., Lan-Chow-Wing O., Rocha A., Zanuy S. & Gómez A. foxl2 and foxl3 are two ancient paralogs that remain fully functional in teleosts. Gen. Comp. Endocr. 194, 81–93 (2013). [DOI] [PubMed] [Google Scholar]
- Jiang W. et al. Effects of sexual steroids on the expression of foxl2 in Gobiocypris rarus. Comp. Biochem. Phys. B. 160, 187–193 (2011). [DOI] [PubMed] [Google Scholar]
- Alam M. A., Kobayashi Y., Horiguchi R., Hirai T. & Nakamura M. Molecular cloning and quantitative expression of sexually dimorphic markers Dmrt1 and Foxl2 during female-to-male sex change in Epinephelus merra. Gen. Comp. Endocr. 157, 75–85 (2008). [DOI] [PubMed] [Google Scholar]
- Cocquet J. et al. Evolution and expression of FOXL2. J. Med. Genet. 39, 916–921 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hancock W. W. & Özkaynak E. Three distinct domains contribute to nuclear transport of murine Foxp3. PloS one 4, e7890 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romanelli M. G., Tato L., Lorenzi P. & Morandi C. Nuclear localization domains in human thyroid transcription factor 2. BBA-Mol. Cell Res. 1643, 55–64 (2003). [DOI] [PubMed] [Google Scholar]
- Kobayashi Y., Horiguchi R., Nozu R. & Nakamura M. Expression and localization of forkhead transcriptional factor 2 (Foxl2) in the gonads of protogynous wrasse, Halichoeres trimaculatus. Biol. Sex Differ. 1, 1–9 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wotton K. R., French K. E. M. & Shimeld S. M. The developmental expression of foxl2 in the dogfish Scyliorhinus canicula. Gene Expr. Patterns. 7, 793–797 (2007). [DOI] [PubMed] [Google Scholar]
- Wang H., Wu T., Qin F., Wang L. & Wang Z. Molecular cloning of Foxl2 gene and the effects of endocrine-disrupting chemicals on its mRNA level in rare minnow, Gobiocypris rarus. Fish Physiol. Biochem. 38, 653–664 (2012). [DOI] [PubMed] [Google Scholar]
- Liu Z. et al. Molecular cloning of doublesex and mab-3-related transcription factor 1, forkhead transcription factor gene 2, and two types of cytochrome P450 aromatase in Southern catfish and their possible roles in sex differentiation. J. Endocrinol. 194, 223–241 (2007). [DOI] [PubMed] [Google Scholar]
- Oshima Y., Uno Y., Matsuda Y., Kobayashi T. & Nakamura M. Molecular cloning and gene expression of Foxl2 in the frog Rana rugosa. Gen. Comp. Endocr. 159, 170–177 (2008). [DOI] [PubMed] [Google Scholar]
- Sridevi P. & Senthilkumaran B. Cloning and differential expression of FOXL2 during ovarian development and recrudescence of the catfish, Clarias gariepinus. Gen. Comp. Endor. 174, 259–268 (2011). [DOI] [PubMed] [Google Scholar]
- Pannetier M. et al. Expression studies of the PIS-regulated genes suggest different mechanisms of sex determination within mammals. Cytogenet. Genome Res. 101, 199–205 (2003). [DOI] [PubMed] [Google Scholar]
- Fleming N. I. et al. Aromatase is a direct target of FOXL2: C134W in granulosa cell tumors via a single highly conserved binding site in the ovarian specific promoter. PloS one 5, e14389 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang D. S. et al. Foxl2 up-regulates aromatase gene transcription in a female-specific manner by binding to the promoter as well as interacting with ad4 binding protein/steroidogenic factor 1. Mol. Endocrinol. 21, 712–725 (2007). [DOI] [PubMed] [Google Scholar]
- Wu G. C., Tomy S., Nakamura M. & Chang C. F. Dual roles of cyp19a1a in gonadal sex differentiation and development in the protandrous black porgy, Acanthopagrus schlegeli. Biol. Reprod. 79, 1111–1120 (2008). [DOI] [PubMed] [Google Scholar]
- Hudson Q. J., Smith C. A. & Sinclair A. H. Aromatase inhibition reduces expression of FOXL2 in the embryonic chicken ovary. Dev. Dynam. 233, 1052–1055 (2005). [DOI] [PubMed] [Google Scholar]
- Ijiri S. et al. Sexual dimorphic expression of genes in gonads during early differentiation of a teleost fish, the Nile tilapia Oreochromis niloticus. Biol. Reprod. 78, 333–341 (2008). [DOI] [PubMed] [Google Scholar]
- Yu J. H., Tang Y. K. & Li J. L. Cloning, structure, and expression pattern of the P-450 aromatase gene in rice field eel (Monopterus albus). Biochem. Genet. 46, 267–280 (2008). [DOI] [PubMed] [Google Scholar]
- Liu C. & Ku K. Histological changes in the gonad of Monopterus during sex transformation. Acta. Hydrobiol. Sin. 2, 85–109 (1951). [Google Scholar]
- Hu Q., Guo W., Gao Y., Tang R. & Li D. Reference gene selection for real-time RT-PCR normalization in rice field eel (Monopterus albus) during gonad development. Fish Physiol. Biochem. 10.1007/s10695-014-9962-3 (2014). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
supplementary file