

Abstract
Aesthetic appreciation is part of our everyday life: it is a subjective judgment we make when looking at a painting, a landscape, or—in fact—at another person. Neuroimaging and electrophysiological evidence suggests that the left dorsolateral prefrontal cortex (DLPFC) plays a critical role in aesthetic judgments. Here, we show that the experience of beauty can be artificially enhanced with brain stimulation. Specifically, we show that aesthetic appreciation of representational paintings and photographs can be increased by applying anodal (excitatory) transcranial direct current stimulation on the left DLPFC. Our results thus show that beauty is in the brain of the beholder, and offer a novel view on the neural networks underlying aesthetic appreciation.
Keywords: neuroaesthetics, liking, art, colour, representational, abstract, tDCS, dorsolateral prefrontal cortex, brain stimulation
INTRODUCTION
‘Beauty—be not caused—It Is’ (E. Dickinson). For centuries, the domain of aesthetics has occupied a central place in philosophy, literature and art. It is hence not surprising that brain mechanisms underlying aesthetic appreciation have been object of increasing research interest in the last decades (Chatterjee, 2011). Aesthetic appreciation is not confined to artworks: ‘the connection between art and aesthetics is a matter of historical contingency, and not part of the essence of art’ (Danto, 1997). In fact, the range of objects that can be appreciated aesthetically includes any natural object or phenomenon, human production or person (Valentine, 1962). Moreover, aesthetic appreciation may also apply to ‘ugly’ objects that still elicit a gratifying aesthetic experience in the observer (Eco, 2007; Silvia and Brown, 2007; Silvia, 2009).
The use of neuroimaging has been instrumental in advancing our understanding of the cognitive and neural underpinnings of aesthetic appreciation, spurring the development of the cognitive neuroscience of aesthetics (Cela-Conde et al., 2011). This field has emerged at the intersection of psychology, neuroscience and aesthetics, and its diverse aims and methods reflect its multidisciplinary nature (Nadal and Pearce, 2011). Despite this, consistent neuroimaging and electrophysiological evidence suggests that the left prefrontal cortex (PFC) plays a critical role in aesthetic appreciation. Using magnetoencephalography (MEG), Cela-Conde et al. (2004) revealed a significantly higher activation in the left dorsolateral PFC (lDLPFC) for artworks and photographs that participants found beautiful, compared to artworks and photographs they judged as not beautiful. Lengger et al. (2007), who studied the relation between slow cortical potentials and the role of stylistic information in aesthetic appreciation, also reported an increase in the activity in the lDLPFC when participants rated the aesthetic qualities of visual art. The functional MRI (fMRI) study of Vessel et al. (2012) on the emotional aspect of aesthetic appreciation likewise showed that activity in this region was greater when participants viewed artworks that moved them aesthetically. These studies argued that the lDLPFC plays a crucial role in aesthetic appreciation related to executive functions in general, and to orienting and sustaining attention in particular. Vessel et al. (2012) even suggested that the pattern of activity observed in this and related frontal regions constitutes a signature of an aesthetic response.
Indeed, there are grounds to postulate that the increase in DLPFC activity observed during aesthetic appreciation is specifically related to the adoption of an aesthetic orientation towards stimuli. While viewing images, individuals without a strong background in visual arts or design tend to focus primarily on the depicted objects, aiming to identify them and understand the scene (Winston and Cupchik, 1992; Nodine et al., 1993). However, Cupchik et al. (2009) showed that people could be prompted to orient aesthetically towards visual stimuli. Their results revealed, moreover, that this orientation was associated with an increase in the activity in the lDLPFC, which was not observed when participants were encouraged to view the stimuli under a ‘common’ or ‘pragmatic’ mode (Cupchik et al., 2009). Aesthetic appreciation, thus, does not emerge from a purely bottom-up processing of stimuli, but relies also on a complex set of top-down cognitive operations aimed at selecting and evaluating certain information—the aesthetic features—present in the external stimulus (Ridderinkhof et al., 2004; Cupchik et al., 2009). The fact that this deliberate direction of attention towards—and evaluation of—information presented in the stimuli relies on activity in the DLPFC is in agreement with this region’s critical role in the processing and manipulation of externally presented information (Christoff and Gabrieli, 2000).
There is also evidence to suggest that the lDLPFC plays a more critical role in the aesthetic evaluation of representational than abstract art. Lengger et al. (2007) found greater lDLPFC activity with representational images than abstract ones. The stimuli set of Cupchik et al. (2009) included only representational stimuli, and Vessel et al. (2012) used over 90% of representational images. The aesthetic appreciation of abstract stimuli, conversely, appears to rely on a different kind of processing, based on internally generated information (Jacobsen and Höfel, 2003). This internally guided processing could be related with naïve participants’ lack of sufficient knowledge of the relevant features on which to ground an aesthetic decision about abstract works, given the absence of meaningful objects with which to parse the image (Ishai et al., 2007).
Because neuroimaging and electrophysiological studies have provided only correlational evidence and relied heavily on reverse inference, knowledge about the specific role of the lDLPFC in aesthetic appreciation is, at present, mostly conjectural. This study aims to overcome these limitations using for the first time transcranial direct current stimulation (tDCS) to directly test the effects of lDLPFC activity on the aesthetic appreciation of representational and abstract images. Brain stimulation, by modifying activation in the targeted region, allows drawing causal inferences on the role of a specific cortical area in the considered process (Pascual-Leone et al., 2000). More specifically, tDCS consists of the non-invasive, transcranial and painless induction of weak direct currents able to induce focal, prolonged and fully reversible shifts of cortical excitability, whose duration after the end of the stimulation and direction depend on stimulation parameters, such as the stimulation duration and polarity (anodal vs cathodal) (for recent reviews, see Medeiros et al., 2012; Miniussi et al., 2013). Consistent evidence suggests that anodal tDCS (i.e. anode placed over the region of interest) causes an enhancement of cortical excitability during stimulation, which lasts several minutes after the end of the stimulation, and that is usually accompanied by enhancement in cognitive performance (e.g. Batsikadze et al., 2013; for a review, see Jacobson et al., 2012).
Our initial hypothesis was that, in the case of naïve participants, lDLPFC activity is related to the adoption of an aesthetic orientation towards visual stimuli, with higher activity for the most liked items. It follows, thus, that enhancing activity in the lDLPFC via tDCS should lead to a greater appreciation for the presented pictures. This hypothesis pieces together the evidence provided by the studies noted above, and, if supported, will offer a unified explanation for some of their seemingly unrelated results. Aiming to test this hypotheses, we carried out a tDCS experiment (Experiment 2), in which a group of participants were asked to rate how much they liked a series of pictures before and after real or sham tDCS applied over the lDLPFC (with the real and sham sessions taking place on different days). To control for selective effects of anodal tDCS on aesthetic ratings, another group of participants was required to evaluate the same images in terms of their colourfulness (again prior to or following real vs sham tDCS). To achieve an optimal tDCS design, we needed two equivalent sets of stimuli to contrast our initial hypothesis, so the objective of Experiment 1 was to test the equivalence between two sets of selected stimuli in terms of their perceived beauty and colourfulness.
EXPERIMENT 1
Method
Participants
Twelve participants (6 F, mean age = 24.75 years, s.d. = 2.60, range = 20–29 years) with no previous training or special interest in art volunteered to participate in this study. They were all right handed (Oldfield, 1971) and all had normal or corrected to normal vision and normal colour vision. Written informed consent was obtained from all participants. The experiment was approved by the local ethical committee of the University of Milano-Bicocca and subjects were treated in accordance with the Declaration of Helsinki.
Stimuli
Stimuli consisted of reproductions of artistic paintings and photographs of natural objects similar to those used in previous MEG work (Cela-Conde et al., 2004, 2009) and belonged to two main categories: representational images (paintings and real photographs) and abstract images. There were 140 abstract images: 100 pictures of abstract paintings and 40 abstract coloured decorative sketches. There were 160 representational images: 50 realist artworks, 50 impressionist and postimpressionist artworks, and 60 photographs of landscapes, artefacts, urban scenes, and so forth [true-life pictures from the Master Clips Premium Image Collection (ISMI, San Rafael, CA), used in industrial design, book illustrating, etc.; images taken from Parr (2000); and photographs taken by the authors]. Figure 1A shows an example of each of the categories. Different artistic styles were included to present participants with a broad range of possibilities of aesthetic appreciation. To avoid the activation of facial-recognition brain mechanisms, pictures containing close views of humans were not included. Stimuli were adjusted for level of luminance and colour spectrum (see Cela-Conde et al., 2004 for details). Two sub-sets of pictures were created: each set contained 70 abstract images (50 abstract paintings and 20 decorative sketches) and 80 figurative images (25 realist art, 25 impressionist and postimpressionist art, 30 photographs of natural and urban scenes or real objects). Stimuli were assigned to the two different sets (Set A and Set B) following the criterion to have as many similar exemplars as possible in each set (e.g. same number of landscape photographs, and so forth) for each category. Six additional pictures were used in a first practice session that preceded the experiment itself.
Fig. 1.

(A) Examples of pictures used in the experiments. The upper panel shows representational images (from left to right: a realism style and an impressionist style painting, reproduced with kind permission of the Museo Nacional del Prado, Madrid (Spain); and a real photograph of an urban scene taken from one of the authors). The lower panel shows abstract exemplars (from left to right: an abstract painting and an abstract design. The examples presented here, free of copyright restrictions, are meant only to illustrate the categories and were not actually used in the experiment). (B) Example of an experimental trial. In each trial, a picture was presented in the middle of the screen and participants had to indicate, by moving the mouse cursor along a rating bar, how much they liked the image (aesthetic evaluation) or how much they considered it colourful (colour evaluation). The left end of the rating bar corresponded to 0% score (i.e. ‘I do not like it at all’/‘not colourful at all’). The right end of the rating bar corresponded to 100% score (‘I absolutely like it’/‘absolutely colourful’). Within the experiment, the mouse arrow appeared randomly an equal number of times below the left or the right end of the rating bar.
Procedure
Participants were seated in front of a 15.5″ PC (1280*800 pixels) screen at an approximate distance of 57 cm, in a normal-lightened and silent room, and asked to perform a computerized rating task. Before starting the experiment, participants were informed that they would be viewing the same sets of artistic and natural pictures twice: in one case, they would have required to express an aesthetic judgment (‘How much do you like this image?’), in the other case to indicate how colourful that specific image was (‘How colourful is this image?’).
Figure 1B shows the timeline of an experimental trial. Each trial started with the presentation of one of the images that appeared on a gray (R,G,B:120,120,120) background and subtended a visual angle of 10° × 10°. A blue horizontal rating bar appeared below each picture (see Figure 1B). Participants were informed that the bar was meant to express a 0–100% scale: the left end of the bar corresponded to a zero level of liking (or colourfulness, depending on the task), whereas the right end of the bar corresponded to the maximum level of liking (or colourfulness). Participants were instructed to express their judgment by clicking with the mouse using their right hand. The mouse cursor was a fully vertical arrow that appeared underneath either the left or the right extreme of the line and moved only horizontally. The initial position (left or right) of the cursor was randomly assigned for each trial. The image remained visible until participants expressed their judgment. After responding, the screen was cleared-out for 300 ms, after which a new image was presented. Before starting each task, participants were presented with six practice trials. There was no time limit but participants were encouraged to respond within 1 min after the appearance of each stimulus. The order of task presentation (aesthetic appreciation and colour judgment) was counterbalanced across participants and within male and female participants. Images in each set were presented in random order, but Set A was always presented first, and Set B second for both the aesthetic and the colour judgment tasks. The whole experiment lasted approximately 45 min. The software E-prime 2.0 (Psychology Software Tools, Inc., Pittsburgh, PA, USA) was used for stimuli presentation and data recording.
Results
Rating scores
The position of the mouse cursor along the bar was automatically converted by the software to percentage rating scores, where a 0% score corresponded to the mouse cursor positioned at the left end of the rating bar and a 100% score corresponded to the mouse cursor positioned at the right end of the rating bar. Figure 2A show the mean percentage rating scores for the aesthetic evaluation and the colour evaluation as a function of Image type (representational vs abstract) and Set (A and B).
Fig. 2.

(A) Participants’ mean rating percentage scores and (B) mean RT for representational and abstract images for Set A and Set B in the aesthetic evaluation and colour evaluation. The two sets did not significantly differ in terms of liking or the perceived amount of colour. Participants were overall faster in Set B than in Set A, an effect that was significant for aesthetic evaluation only, possibly reflecting task familiarization effects. Error bars represent ±1 s.e.m.
Aesthetic evaluation
A repeated-measures analysis of variance (ANOVA) with Set (A and B) and Image type (representational vs abstract) as within-subjects variable and task order (aesthetic first vs colour first) as between-subjects variable was carried out on the mean percentage rating scores for the aesthetic evaluation. The analysis revealed a significant main effect of Image type, F(1, 10) = 9.14, P = 0.013, 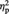 = 0.48, indicating overall higher liking for representational images than abstract ones (nine out of 12 participants showed this pattern). Critically, the main effect of Set was not significant (P = 0.86), confirming that Set A and Set B were comparable in terms of liking for the images. Task order was not significant (P = 0.80) nor were any of the possible interactions (all Ps > 0.27).
= 0.48, indicating overall higher liking for representational images than abstract ones (nine out of 12 participants showed this pattern). Critically, the main effect of Set was not significant (P = 0.86), confirming that Set A and Set B were comparable in terms of liking for the images. Task order was not significant (P = 0.80) nor were any of the possible interactions (all Ps > 0.27).
Colour evaluation
A similar ANOVA was performed on the mean percentage rating scores for the colour evaluation. The analysis revealed a significant main effect of Image type, F(1, 10) = 23.13, P = 0.001, 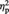 = 0.70, with abstract images being judged as more colourful than representational ones. Critically, the main effect of Set was not significant (P = 0.54), confirming that Set A and Set B were comparable in terms of perceived amount of colour in the images. Neither the main effect of task order (P = 0.98) nor any of the possible interactions (all Ps > 0.20) reached significance.
= 0.70, with abstract images being judged as more colourful than representational ones. Critically, the main effect of Set was not significant (P = 0.54), confirming that Set A and Set B were comparable in terms of perceived amount of colour in the images. Neither the main effect of task order (P = 0.98) nor any of the possible interactions (all Ps > 0.20) reached significance.
Correlational analyses (Pearson r, two-tailed) were carried out to assess whether in each type of evaluation (aesthetic and colour), rating scores were correlated between Set A and Set B for representational and abstract images. For aesthetic evaluation, participants’ rating scores were found to be highly positively correlated between the two sets for both representational, r = 0.932, P < 0.001, and abstract exemplars, r = 0.991, P < 0.001. Similarly, for colour evaluation, participants’ scores were highly positively correlated between the two sets for both representational, r = 0.909, P < 0.001, and abstract images, r = 0.863, P < 0.001.
Additional correlational analyses (Pearson r, two-tailed) were performed to assess whether colour and aesthetic evaluation were correlated. To this end, we considered the overall rating scores for representational and abstract images (regardless of set). A positive significant correlation was found between liking and colour judgments for both representational images, r = 0.656, P = 0.020, and abstract images, r = 0.734, P = 0.007, indicating that the more colourful an image was rated, the more it was liked. Notably, this analysis also suggests that colour is an important but not the critical dimension in aesthetic appreciation: if colour played a major role, abstract images (rated as significantly more colourful, see above) should have also been rated as more liked, but results showed the opposite pattern.
Response latencies
Figure 2B shows the mean response latencies (RT) for the aesthetic evaluation and the colour evaluation as a function of Image type (representational vs abstract) and Set (A and B).
Aesthetic evaluation
A repeated-measures ANOVA with Set (A and B) and Image type (representational vs abstract) as within-subjects variable and task order (aesthetic first vs colour first) as between-subjects variable revealed a significant main effect of Image type, F(1, 10) = 6.80, P = 0.026, 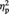 = 0.41, showing that, overall, participants took longer for representational than for abstract images when rating how much they liked them. The ANOVA also revealed a significant main effect of Set, F(1, 10) = 7.67, P = 0.020,
= 0.41, showing that, overall, participants took longer for representational than for abstract images when rating how much they liked them. The ANOVA also revealed a significant main effect of Set, F(1, 10) = 7.67, P = 0.020, 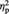 = 0.43, with faster RT in Set B than in Set A (as Set B was always rated after Set A, this speed up of responses possibly reflected stabilization of the criterion used). Task order was not significant (P = 0.31), nor were any of the possible interactions (all Ps > 0.12).
= 0.43, with faster RT in Set B than in Set A (as Set B was always rated after Set A, this speed up of responses possibly reflected stabilization of the criterion used). Task order was not significant (P = 0.31), nor were any of the possible interactions (all Ps > 0.12).
Colour evaluation
A similar ANOVA performed on RT in colour evaluation revealed no significant main effect for Image type (P = 0.20), Set (P = 0.29) or Task order (P = 0.89). None of the possible interactions reached significance (all Ps > 0.22).
EXPERIMENT 2
Method
Participants
Twelve participants (5 F, mean age = 23.42 years, s.d. = 3.15, range = 19–28 years) took part in the aesthetic evaluation study and 12 participants (8 F, mean age = 23.83 years, s.d. = 1.94, range = 21–26 years) took part in the colour evaluation study. None of the participants had previous training or special interest in art; none had taken part in Experiment 1. All participants were right handed (Oldfield, 1971) and all had normal or corrected to normal vision and normal colour vision. All participants had no history of chronic or acute neurologic, psychiatric, or medical disease, no family history of epilepsy, no current pregnancy, no cardiac pacemaker, no previous surgery involving implants to the head (cochlear implants, aneurysm clips, brain electrodes), and did not take acute or chronic medication. Written informed consent was obtained from all participants. The experiment was approved by the local ethical committee of the University of Milano-Bicocca and subjects were treated in accordance with the Declaration of Helsinki.
Transcranial direct current stimulation
tDCS was delivered by a battery driven, constant current stimulator (Eldith, Neuroconn, Ilmenau, Germany) through a pair of saline-soaked sponge electrodes (7 × 5 cm: 35 cm2) kept firm by elastic bands. The excitability-enhancing anodal electrode was placed over the left DLPFC localized as the middle point between F3 and F5 in the 10-20 electroencephalography (EEG) system (see Rusjan et al., 2010), whereas the cathodal electrode was placed over the right supraorbitary region. This electrode arrangement (anodal electrode over one DLPFC with the cathodal electrode over the contralateral supraorbital area) is thought to induce unilateral modulation of one DLPFC and has been shown effective in various studies (Kincses et al., 2004; Fregni et al., 2005; Fecteau et al., 2007).
Each participant underwent two stimulation sessions: a real one and a sham one. In each session, participants performed the task twice: once before stimulation, and once after stimulation. The images they evaluated were different in the pre- and post-tDCS evaluation (see below). Sessions were separated by an average of 3.5 days (range: 2–5 days). The order of stimulation session was counterbalanced across participants, so that half started with the sham session and the other half with the real session. In the real tDCS session, stimulation intensity was set at 2 mA and the duration of stimulation was 20 min. Previous studies have shown that this intensity of stimulation is safe and can be more effective than a 1 mA stimulation (Moos et al., 2012). Moreover, 20 min of 2 mA anodal stimulation results in an excitability enhancement that is still observable 90 min after the end of the stimulation (e.g. Batsikadze et al., 2013). For the sham stimulation, the electrodes were placed at the same positions as for active stimulation, but the stimulator was turned on only for 30 s. Thus, participants felt the initial itching sensation associated with tDCS, but received no active current for the rest of the stimulation period. This procedure ensured that participants felt the initial itching sensation at the beginning of the sham stimulation, but prevented any effective modulation of cortical excitability by sham tDCS, thus allowing for a successful blinding of participants for the real vs sham stimulation condition (Russo et al., 2013). The study was a single-blind experiment: participants were not aware of the type of stimulation they received, whereas the experimenter was fully informed (see Cattaneo et al., 2011; Pisoni et al., 2012 for a similar procedure).
Stimuli and procedure
The same stimuli and apparatus used in Experiment 1 were used here. In the aesthetic judgment task, participants had to rate how much they liked the presented image. In the colour judgment task (control task), participants had to rate the colourfulness of the presented image. In both cases, responses were given using the mouse as described for Experiment 1. In each experimental session (sham and real), participants were first informed about the task and their left DLPFC was localized. Set A was hence presented. After completion of the ratings for Set A, electrodes were placed over the participants’ head and the stimulation was started. Concurrently with the beginning of the stimulation, a cartoon movie was projected on the computer screen. This was done in order to reduce inter-subjects variability by exposing participants to the same visual experience during the stimulation period (see Cattaneo et al., 2011; Pisoni et al., 2012 for a similar procedure). After 18 min since the beginning of the stimulation, the cartoon movie was stopped and subjects were told that in 2 min they would have to perform the rating task for a new set of images (Set B). The rating task was administered within 1 min from the end of the tDCS stimulation. In all participants, the task was completed within 10 min from the end of the tDCS stimulation.
Results
Although participants were not systematically asked to report their sensations, no participants spontaneously reported different sensations in the real and the sham stimulation conditions. Analyses were performed on rating scores and mean RT. In particular, for both the aesthetic and the colour tasks a repeated-measures ANOVA was carried out on mean rating scores and RT with Session (prior to vs post tDCS), Image type (abstract vs representational) and tDCS type (real vs sham) as within-subjects factors.
Aesthetic evaluation
Rating scores
Figure 3A shows the mean percentage rating scores for the aesthetic evaluation in the different experimental conditions. Figure 3B shows the difference in aesthetic appreciation scores between post-stimulation and pre-stimulation (=‘post-tDCS scores − pre-tDCS scores’) for representational and abstract images for real and sham tDCS. The ANOVA revealed a significant main effect of Image type, F(1, 11) = 14.41, P = 0.003, 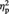 = 0.57, indicating higher aesthetic appreciation of representational than of abstract images, and a significant interaction Session by Image type by tDCS type, F(1, 11) = 4.83, P = 0.05,
= 0.57, indicating higher aesthetic appreciation of representational than of abstract images, and a significant interaction Session by Image type by tDCS type, F(1, 11) = 4.83, P = 0.05, 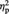 = 0.31. None of the other main effects or interactions reached significance (all Ps > 0.12). The three-way significant interaction was further analysed by looking at the simple main effects of Image type and tDCS type within each session (pre and post) separately.
= 0.31. None of the other main effects or interactions reached significance (all Ps > 0.12). The three-way significant interaction was further analysed by looking at the simple main effects of Image type and tDCS type within each session (pre and post) separately.
Fig. 3.

(A) Participants’ mean rating percentage scores in the aesthetic evaluation for representational and abstract images in the different stimulation conditions (i.e. prior to/following real vs sham tDCS). (B) Difference between post- and pre-stimulation participants’ mean rating percentage scores in the aesthetic evaluation for representational and abstract images for real and sham tDCS. (C) Participants’ mean rating percentage scores in the aesthetic evaluation for representational paintings and real photographs prior to and following real or sham tDCS. Error bars represent ±1 s.e.m. Asterisks indicate a significant difference between sham and real tDCS.
For the pre-tDCS session, a repeated-measures ANOVA with Image type (abstract vs representational) and tDCS type (real vs sham) as within-subjects factors showed a significant main effect of Image type, F(1, 11) = 14.86, P = 0.003, 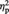 = 0.58, indicating higher aesthetic appreciation of representational than of abstract images (11 out of 12 participants showed this pattern). Neither the main effect of tDCS type (P = 0.25) nor the interaction Image type by tDCS (P = 0.39) was significant, indicating that the ‘baseline evaluation’ preceding tDCS was comparable between the real and the sham sessions. For the post-tDCS session, the ANOVA showed a significant main effect of Image type, F(1, 11) = 13.77, P = 0.003,
= 0.58, indicating higher aesthetic appreciation of representational than of abstract images (11 out of 12 participants showed this pattern). Neither the main effect of tDCS type (P = 0.25) nor the interaction Image type by tDCS (P = 0.39) was significant, indicating that the ‘baseline evaluation’ preceding tDCS was comparable between the real and the sham sessions. For the post-tDCS session, the ANOVA showed a significant main effect of Image type, F(1, 11) = 13.77, P = 0.003, 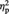 = 0.56, again reflecting higher scores for representational than for abstract exemplars. The main effect of tDCS was not significant (P = 0.26). Critically, the interaction tDCS by Image type was significant, F(1, 11) = 5.11, P = 0.045,
= 0.56, again reflecting higher scores for representational than for abstract exemplars. The main effect of tDCS was not significant (P = 0.26). Critically, the interaction tDCS by Image type was significant, F(1, 11) = 5.11, P = 0.045, 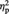 = 0.32. Tukey post hoc comparisons (Bonferroni correction applied) showed that liking scores were higher after real than after sham stimulation for representational images, t(11) = 3.22, P = 0.032, but not for abstract images, t(11) = 1.07, P = 0.309.
= 0.32. Tukey post hoc comparisons (Bonferroni correction applied) showed that liking scores were higher after real than after sham stimulation for representational images, t(11) = 3.22, P = 0.032, but not for abstract images, t(11) = 1.07, P = 0.309.
Planned pairwise t-tests showed that for representational images the difference between post-tDCS scores and pre-tDCS scores was significantly greater for real than for sham tDCS, t(11) = 2.29, P = 0.043, whereas for abstract images the difference in evaluation between the pre- and post-session was comparable for sham and real tDCS, t(11) < 1, P = 0.946 (see Figure 3B).
An additional analysis was carried out to verify whether the effect of real tDCS on aesthetic evaluation found with representational images extended to representational art (realist, impressionist and post-impressionist paintings) and photographs. Figure 3C shows aesthetic appreciation scores for representational paintings and real photographs in each experimental condition. Mean rating scores did not differ overall for representational paintings and photographs, t(11) = 0.415, P = 0.69. Critically, planned pairwise comparisons showed that aesthetic appreciation was significantly higher following real tDCS than sham tDCS for both representational art, t(11) = 2.37, P = 0.037, and photographs, t(11) = 2.24, P = 0.047.
Response latencies
Figure 4 shows the mean RT for the aesthetic evaluation in the different experimental conditions. The ANOVA revealed a significant main effect of Image type, F(1, 11) = 23.42, P = 0.001, 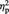 = 0.68, indicating faster evaluation of abstract than representational images (resembling the pattern found in Experiment 1). The main effect of session was significant, F(1, 11) = 17.51, P = 0.002,
= 0.68, indicating faster evaluation of abstract than representational images (resembling the pattern found in Experiment 1). The main effect of session was significant, F(1, 11) = 17.51, P = 0.002, 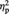 = 0.61, with evaluation being faster in the post-tDCS than in the pre-tDCS session (regardless of stimulation type, i.e. real vs sham), likely reflecting stabilization of criterion used/task familiarization effects resulting in faster responses. None of the other main effects or interactions reached significance (all Ps > 0.19).
= 0.61, with evaluation being faster in the post-tDCS than in the pre-tDCS session (regardless of stimulation type, i.e. real vs sham), likely reflecting stabilization of criterion used/task familiarization effects resulting in faster responses. None of the other main effects or interactions reached significance (all Ps > 0.19).
Fig. 4.

Participants’ mean RT for representational and abstract images in the aesthetic evaluation task for each experimental condition (pre/post real/sham tDCS). Error bars represent ±1 s.e.m.
Colour evaluation
Rating scores
Figure 5A shows participants’ mean percentage rating scores for the colour evaluation of representational and abstract images in the pre- and post-tDCS session (for both real and sham tDCS). The ANOVA revealed a significant main effect of Image type, F(1, 11) = 18.72, P = 0.001, 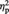 = 0.63, indicating that participants overall evaluated abstract exemplars as more colourful than representational exemplars. None of the other main effects or interactions reached significance (all Ps > 0.37; in particular for the interaction Session by Image type by tDCS type: F(1, 11) < 1, P = 0.81,
= 0.63, indicating that participants overall evaluated abstract exemplars as more colourful than representational exemplars. None of the other main effects or interactions reached significance (all Ps > 0.37; in particular for the interaction Session by Image type by tDCS type: F(1, 11) < 1, P = 0.81, 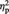 = 0.01).
= 0.01).
Fig. 5.

(A) Participants’ mean rating percentage scores and (B) mean RT in the colour evaluation for representational and abstract images as a function of experimental condition (pre-tDCS vs Post-tDCS; sham vs real). tDCS did not significantly affect colour judgments. Error bars represent ±1 s.e.m.
Response latencies
Figure 5B shows participants’ mean RT for the colour evaluation of representational and abstract images in the pre- and post-tDCS session, for both real and sham tDCS. The ANOVA showed a significant main effect of session, F(1, 11) = 18.13, P = 0.001, 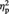 = 0.62, with evaluation being faster in the post-tDCS than in the pre-tDCS session (regardless of stimulation type), resembling the pattern found in the aesthetic evaluation and possibly reflecting task familiarization effects. None of the other main effects or interactions reached significance (all Ps > 0.10).
= 0.62, with evaluation being faster in the post-tDCS than in the pre-tDCS session (regardless of stimulation type), resembling the pattern found in the aesthetic evaluation and possibly reflecting task familiarization effects. None of the other main effects or interactions reached significance (all Ps > 0.10).
DISCUSSION
Overall, our results suggest that the lDLPFC plays a causal role in visual aesthetic appreciation. Specifically, enhancing activity in the left DLPFC by applying anodal tDCS over this region resulted in a moderate (around 3%) but statistically significant increase of aesthetic appreciation of representational images (both artworks and photographs). In turn, stimulation did not affect evaluation of abstract images, suggesting that the neural mechanisms underlying appreciation of figurative and abstract images may be different (at least in individuals with no strong background in fine arts tending to spontaneously prefer figurative images) (Fairhall and Ishai, 2008).
Critically, no effect of tDCS over the left DLPFC was observed when the same images had to be judged in terms of the amount of colour they contained. This indicates that the effect of tDCS was specific for the aesthetic evaluation (‘How much do you like this image’), and not general for any kind of visual judgment on the image’s attributes. This is even more remarkable when considering that aesthetic judgments and colour judgments were positively correlated (i.e. aesthetic appreciation increased with the increase of perceived colour extent of the images), as showed in Experiment 1. Notably, although abstract images were rated overall as significantly more colourful than representational images, representational paintings and photos, overall, were liked significantly more than abstract images. This indicates that although colour is likely to be a critical component in aesthetic judgment, liking is most probably the result of the interactive evaluation of multiple aspects of the stimulus and one’s affective response to it, such as form, content and elicited emotions (Leder et al., 2004; Nadal et al., 2008). Moreover, it could be objected that the selective effects of tDCS on aesthetic evaluation of representational images depended on tDCS affecting a specific (more central) portion of the evaluation bar (i.e. the central part, corresponding to an appreciation around 50%). However, this is unlikely. In fact, if the tDCS effects we reported were merely due to tDCS affecting scores in central/high ranges only (around 50% or more), regardless of the type of required visual judgment, tDCS should have affected colour judgments for abstract images as well (with mean rating scores of abstract pictures in the colour task being comparable and even higher than liking scores received by representational images in the aesthetic task).
In their integrative theory of PFC functions, Miller and Cohen (2001) suggest that the PFC exerts a top-down control by specifying the pattern of neural pathways that are currently needed, inhibiting—if needed—those that are more automatically activated by the external input but less relevant for the task at play. In the context of aesthetic appreciation, it has been hypothesized that the left PFC underlies the switching from a ‘pragmatic orientation’ to an ‘aesthetic orientation’ (Cupchik et al., 2009). Accordingly, our finding that modulating activity in the lDLPFC directly affected evaluation of representational images would reflect disengagement from a habitual mode of identifying objects to adopt an aesthetic perspective, also in line with previous brain stimulation evidence pointing to the role of the prefrontal cortices in cognitive control (e.g. Fecteau et al., 2007). This ‘switch’ entails focusing attention on the stimuli’s aesthetic qualities and evaluating them. Notably, the effects of tDCS on the left DLPFC were significant for aesthetic evaluation of both representational artworks and photographs. These findings suggest that the left DLPFC plays a role in aesthetic appreciation in general, and not in artistic aesthetic appreciation only (see also Graham et al., 2013 for behavioural evidence). In contrast, art naïve participants usually lack sufficient knowledge about the aesthetic features in abstract art and design to be able to orient toward them—such as a peculiar use of colour or combination of shapes and materials—that are critical in art experts’ evaluation. This might explain the lack of lDLPFC stimulation effects on appreciation of abstract images. This explanation also fits with the pattern of RT we reported. Overall, representational images took more time to be rated in terms of liking than abstract images, likely reflecting a shift toward aesthetic orientation, whereas decision times were the same for representational and abstract images when deciding about the colourfulness of the image, a task which does not require an aesthetic orientation.
In addition to its role in cognitive control, the left DLPFC is also known to be a critical region in emotional processing, being particularly important for down-regulation of negative emotional conditions (e.g. Peña-Gómez et al., 2011). Accordingly, enhancing activation in the left DLPFC through brain stimulation has been found to reduce depressive symptoms (e.g. Boggio et al., 2009). In light of this, we cannot exclude that the effects we reported in our study depended on a conjoined effect of tDCS over cognitive and affective mechanisms, both mediated by the left DLPFC (Ray and Zald, 2012), and both critical in aesthetic experience (Leder et al., 2004; Nadal et al., 2008; Nadal and Pearce, 2011).
A detailed account of the neural mechanisms mediating the effects we reported is beyond the scope of this investigation. Nonetheless, previous fMRI studies assessing the effect of anodal tDCS on the left DLPFC, and using the same montage and stimulation parameters as the ones we used here, found that anodal tDCS increased the resting state functional connectivity within networks that included the DLPFC (Keeser et al., 2011). Given that the left DLPFC is a crucial node in a rostro-caudal network of brain regions underlying the appraisal of aesthetic qualities (Cela-Conde et al., 2013), it is conceivable that our experimental procedure enhanced processes performed along the whole network.
Future research is required to ascertain whether the ratings of art experts, who rely more on long-term memory and the exploration of formal elements (Leder et al., 2004), would be affected in a similar way to naïve participants’, or whether similar effects would be observed in the case of other type of non-visual art (e.g. music). On the other hand, there is evidence that aesthetic responses can be preserved in the face of cognitive decline caused by several forms of dementia (e.g. Halpern et al., 2008; Graham et al., 2013; Halpern and O’Connor, 2013). Continued advances in our knowledge of the neural foundations aesthetic appreciation could inform rehabilitation protocols, not only in healthy or pathological ageing but also in such disorders as major depression or schizophrenia, characterized by a diminished interest or pleasure (anhedonia) (see Der-Avakian and Markou, 2012).
In conclusion, our results show that the judgments of beauty can be artificially enhanced in art naïve individuals using brain stimulation. Emiliy Dickinson was only partially right: beauty certainly is, but it can also be enhanced.
Acknowledgments
We would like to thank Carlo Toneatto for technical support, Marina Bassi and Tiziana Burla for their precious helping in data collection, and Rossana Actis Grosso and Daniele Zavagno for their thoughtful comments on the data. This work has been partly supported by a FIRB (Basic research investment fund) (number: RBFR12F0BD) funded by the Italian Ministry of Education, Universities and Research (MIUR) to ZC.
REFERENCES
- Batsikadze G, Moliadze V, Paulus W, Kuo MF, Nitsche MA. Partially non-linear stimulation intensity-dependent effects of direct current stimulation on motor cortex excitability in humans. The Journal of Physiology. 2013;591:1987–2000. doi: 10.1113/jphysiol.2012.249730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boggio PS, Zaghi S, Fregni F. Modulation of emotions associated with images of human pain using anodal transcranial direct current stimulation (tDCS) Neuropsychologia. 2009;47(1):212–7. doi: 10.1016/j.neuropsychologia.2008.07.022. [DOI] [PubMed] [Google Scholar]
- Cattaneo Z, Pisoni A, Papagno C. Transcranial direct current stimulation over Broca’s region improves phonemic and semantic fluency in healthy individuals. Neuroscience. 2011;183:64–70. doi: 10.1016/j.neuroscience.2011.03.058. [DOI] [PubMed] [Google Scholar]
- Cela-Conde CJ, Agnati L, Huston JP, Mora F, Nadal M. The neural foundations of aesthetic appreciation. Progress in Neurobiology. 2011;94:39–48. doi: 10.1016/j.pneurobio.2011.03.003. [DOI] [PubMed] [Google Scholar]
- Cela-Conde CJ, Ayala FJ, Munar E, et al. Sex-related similarities and differences in the neural correlates of beauty. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:3847–52. doi: 10.1073/pnas.0900304106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cela-Conde CJ, García-Prieto J, Ramasco JJ, et al. Dynamics of brain networks in the aesthetic appreciation. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(Suppl. 2):10454–61. doi: 10.1073/pnas.1302855110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cela-Conde CJ, Marty G, Maestú F, et al. Activation of the prefrontal cortex in the human visual aesthetic perception. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(16):6321–5. doi: 10.1073/pnas.0401427101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatterjee A. Neuroaesthetics: a coming of age story. Journal of Cognitive Neuroscience. 2011;23(1):53–62. doi: 10.1162/jocn.2010.21457. [DOI] [PubMed] [Google Scholar]
- Christoff K, Gabrieli JD. The frontopolar cortex and human cognition: evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology. 2000;28(2):168–86. [Google Scholar]
- Cupchik GC, Vartanian O, Crawley A, Mikulis DJ. Viewing artworks: contributions of cognitive control and perceptual facilitation to aesthetic experience. Brain and Cognition. 2009;70(1):84–91. doi: 10.1016/j.bandc.2009.01.003. [DOI] [PubMed] [Google Scholar]
- Danto AC. After the End of Art: Contemporary Art and the Pale of History. Princeton, NJ: Princeton University Press; 1997. [Google Scholar]
- Der-Avakian A, Markou A. The neurobiology of anhedonia and other reward-related deficits. Trends in Neuroscience. 2012;35(1):68–77. doi: 10.1016/j.tins.2011.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eco U. On Ugliness. London: Harvill Secker; 2007. [Google Scholar]
- Fairhall SL, Ishai A. Neural correlates of object indeterminacy in art compositions. Consciousness and Cognition. 2008;17:923–32. doi: 10.1016/j.concog.2007.07.005. [DOI] [PubMed] [Google Scholar]
- Fecteau S, Pascual-Leone A, Zald DH, et al. Activation of prefrontal cortex by transcranial direct current stimulation reduces appetite for risk during ambiguous decision making. The Journal of Neuroscience. 2007;27(23):6212–8. doi: 10.1523/JNEUROSCI.0314-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fregni F, Boggio PS, Nitsche M, et al. Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory. Experimental Brain Research. 2005;166(1):23–30. doi: 10.1007/s00221-005-2334-6. [DOI] [PubMed] [Google Scholar]
- Graham DJ, Stockinger S, Leder H. An island of stability: art images and natural scenes—but not natural faces—show consistent esthetic response in Alzheimer’s-related dementia. Frontiers in Psychology. 2013;4:107. doi: 10.3389/fpsyg.2013.00107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halpern AR, Ly J, Elkin-Frankston S, O’Connor MG. “I know what I like”: stability of aesthetic preference in Alzheimer’s patients. Brain and Cognition. 2008;66:65–72. doi: 10.1016/j.bandc.2007.05.008. [DOI] [PubMed] [Google Scholar]
- Halpern AR, O’Connor MG. Stability of art preference in frontotemporal dementia. Psychology of Aesthetics, Creativity, and the Arts. 2013;7:95–9. [Google Scholar]
- Ishai A, Fairhall SL, Pepperell R. Perception, memory and aesthetics of indeterminate art. Brain Research Bulletin. 2007;73(4–6):319–24. doi: 10.1016/j.brainresbull.2007.04.009. [DOI] [PubMed] [Google Scholar]
- Jacobsen T, Höfel L. Descriptive and evaluative judgment processes: behavioral and electrophysiological indices of processing symmetry and aesthetics. Cognitive, Affective Behavioral Neuroscience. 2003;3(4):289–99. doi: 10.3758/cabn.3.4.289. [DOI] [PubMed] [Google Scholar]
- Jacobson L, Koslowsky M, Lavidor M. tDCS polarity effects in motor and cognitive domains: a meta-analytical review. Experimental Brain Research. 2012;216:1–10. doi: 10.1007/s00221-011-2891-9. [DOI] [PubMed] [Google Scholar]
- Keeser D, Meindl T, Bor J, et al. Prefrontal transcranial direct current stimulation changes connectivity of resting-state networks during fMRI. Journal of Neuroscience. 2011;31(43):15284–93. doi: 10.1523/JNEUROSCI.0542-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kincses TZ, Antal A, Nitsche MA, Bártfai O, Paulus W. Facilitation of probabilistic classification learning by transcranial direct current stimulation of the prefrontal cortex in the human. Neuropsychologia. 2004;42(1):113–7. doi: 10.1016/s0028-3932(03)00124-6. [DOI] [PubMed] [Google Scholar]
- Leder H, Belke B, Oeberst A, Augustin M. A model of aesthetic appreciation and aesthetic judgments. British Journal of Psychology. 2004;95:489–508. doi: 10.1348/0007126042369811. [DOI] [PubMed] [Google Scholar]
- Lengger P, Fischmeister F, Leder H, Bauer H. Functional neuroanatomy of the perception of modern art: a DC-EEG study on the influence of stylistic information on aesthetic experience. Brain Research. 2007;1158:93–102. doi: 10.1016/j.brainres.2007.05.001. [DOI] [PubMed] [Google Scholar]
- Medeiros LF, de Souza IC, Vidor LP, et al. Neurobiological effects of transcranial direct current stimulation. Frontiers in Psychiatry. 2012;3:110. doi: 10.3389/fpsyt.2012.00110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annual Review of Neuroscience. 2001;24:167–202. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- Miniussi C, Harris JA, Ruzzoli M. Modelling non-invasive brain stimulation in cognitive neuroscience. Neuroscience and Biobehavioral Reviews. 2013;37(8):1702–12. doi: 10.1016/j.neubiorev.2013.06.014. [DOI] [PubMed] [Google Scholar]
- Moos K, Vossel S, Weidner R, Sparing R, Fink GR. Modulation of top-down control of visual attention by cathodal tDCS over right IPS. Journal of Neuroscience. 2012;32(46):16360–8. doi: 10.1523/JNEUROSCI.6233-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadal M, Munar E, Capó MA, Rosselló J, Cela-Conde CJ. Towards a framework for the study of the neural correlates of aesthetic preference. Spatial Vision. 2008;21(3–5):379–96. doi: 10.1163/156856808784532653. [DOI] [PubMed] [Google Scholar]
- Nadal M, Pearce MT. The Copenhagen Neuroaesthetics conference: prospects and pitfalls for an emerging field. Brain and Cognition. 2011;76:172–83. doi: 10.1016/j.bandc.2011.01.009. [DOI] [PubMed] [Google Scholar]
- Nodine CF, Locher PJ, Krupinski EA. The role of formal art training on perception and aesthetic judgment of art compositions. Leonardo. 1993;26:219–27. [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9(1):97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Parr M. Boring Postcards USA. London: Phaidon Press; 2000. [Google Scholar]
- Pascual-Leone A, Walsh V, Rothwell J. Transcranial magnetic stimulation in cognitive neuroscience—virtual lesion, chronometry, and functional connectivity. Current Opinion in Neurobiology. 2000;10(2):232–7. doi: 10.1016/s0959-4388(00)00081-7. [DOI] [PubMed] [Google Scholar]
- Peña-Gómez C, Vidal-Piñeiro D, Clemente IC, Pascual-Leone Á, Bartrés-Faz D. Down-regulation of negative emotional processing by transcranial direct current stimulation: effects of personality characteristics. PLoS One. 2011;6(7):e22812. doi: 10.1371/journal.pone.0022812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pisoni A, Papagno C, Cattaneo Z. Neural correlates of the semantic interference effect: new evidence from transcranial direct current stimulation. Neuroscience. 2012;223:56–67. doi: 10.1016/j.neuroscience.2012.07.046. [DOI] [PubMed] [Google Scholar]
- Ray RD, Zald DH. Anatomical insights into the interaction of emotion and cognition in the prefrontal cortex. Neuroscience and Biobehavioral Reviews. 2012;36(1):479–501. doi: 10.1016/j.neubiorev.2011.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science (New York, N.Y.) 2004;306(5695):443–7. doi: 10.1126/science.1100301. [DOI] [PubMed] [Google Scholar]
- Rusjan PM, Barr MS, Farzan F, et al. Optimal transcranial magnetic stimulation coil placement for targeting the dorsolateral prefrontal cortex using novel magnetic resonance image-guided neuronavigation. Human Brain Mapping. 2010;31(11):1643–52. doi: 10.1002/hbm.20964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo R, Wallace D, Fitzgerald PB, Cooper NR. Perception of comfort during active and sham transcranial direct current stimulation: a double blind study. Brain Stimulation. 2013;6(6):946–51. doi: 10.1016/j.brs.2013.05.009. [DOI] [PubMed] [Google Scholar]
- Silvia PJ. Looking past pleasure: anger, confusion, disgust, pride, surprise, and other unusual aesthetic emotions. Psychology of Aesthetics, Creativity, and the Arts. 2009;3(1):48–51. [Google Scholar]
- Silvia PJ, Brown EM. Anger, disgust, and the negative aesthetic emotions: expanding an appraisal model of aesthetic experience. Psychology of Aesthetics, Creativity, and the Arts. 2007;1(2):100–6. [Google Scholar]
- Valentine CW. The Experimental Psychology of Beauty. London: Methuen & Co; 1962. [Google Scholar]
- Vessel EA, Starr GG, Rubin N. The brain on art: intense aesthetic experience activates the default mode network. Frontiers in Human Neuroscience. 2012;6:66. doi: 10.3389/fnhum.2012.00066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winston AS, Cupchik GC. The evaluation of high art and popular art by naive and experienced viewers. Visual Arts Research. 1992;18:1–14. [Google Scholar]