

Abstract
Mirroring neurons fire both when an individual moves and observes another move in kind. This simulation of others’ movements is thought to effortlessly and ubiquitously support empathetic connection and social understanding. However, at times this could be maladaptive. How could a boxer mirror a losing opponent’s expressions of fatigue, feeling his weariness, precisely when strength is required? Clearly, the boxer must emotionally disconnect from his opponent and those expressions of fatigue must become irrelevant and not mirrored. But, movements that inform of his opponent’s intentions to deliver an incoming blow are quite relevant and still should require mirroring. We tested these dimensions of emotional connectedness and relevance of movement in an electroencephalography experiment, where participants’ desires to socially connect with a confederate were manipulated. Before manipulation, all participants mirrored the confederate’s purely kinematic (a hand opening and closing) and goal-directed (a hand opening and closing around a token that the participant desired) hand movements. After manipulation, unfairly treated subjects ceased to mirror the purely kinematic movements but continued to mirror goal-relevant movements. Those treated fairly continued to mirror all movements. The results suggest that social mirroring can be adaptive in order to meet the demands of a varied social environment.
Keywords: mirror neurons, empathy, affiliation, social cognition, emotion, goals
INTRODUCTION
We are built to connect and to understand
We have the ability to connect with others and to understand their internal states, and we have neural systems that support this connection and understanding. These neural systems work across time, first perceiving what action is being performed, then the implementation of how the action is performed, and finally the mentalization of why it is performed or ‘what does it mean?’ (Thioux et al., 2008). Recent empirical support for this sequential model has identified specific neural substrates responsible for these different levels of abstraction (Spunt et al., 2011; Spunt and Lieberman, 2012). More specifically, the mirror neuron system is involved in the early stages of this processing (the what and the how), with mirror neurons reacting to observed movement in the same way as though the observer had moved himself (di Pellegrino et al., 1992), giving the perceiver an intimate understanding of perceived movements. The mirror neuron system informs the why (mentalizing) system, enabling the ability to infer mental states. These systems, working together, are thought to enable capacities such as empathy (Gazzola et al., 2006), theory of mind (Iacoboni et al., 2005), the understanding of observed touch (Keysers et al., 2004) and imitation (Williams et al., 2006; Belot et al., 2013).
Literature to date has focused on the integral role that mirroring neurons play in the course of understanding. This process is thought to be ubiquitous, connecting us with one another at every turn. While there is certainly value in this research, questions arise when considering how it serves us in real life situations. For instance, what about occasions in which we would benefit most from disconnecting from another person’s feelings, such as encountering a crying child sitting alone on a street curb. Here, it would do little good if we assimilated to (adopted that other’s state) the child’s distraught behavior and emotions; instead, we take on a complementary stance (Eisenberg et al., 1989; Esteves et al., 1994). Instead of crying ourselves we seek to take charge, to comfort and to help the child.
At times we must disconnect
The distinction made in our example implies that mirroring should work differently in cases in which we must connect vs disconnect emotionally. For example, it would be completely counterproductive for a boxer to mirror his opponents’ weariness precisely at the moment when he would need to go in for the win. A ubiquitously operating mirroring mechanism would not by definition differentiate between situations requiring a complementary vs an assimilative behavioral and emotional orientation. It is not hard to generate other situations and contexts in which exclusively assimilative mirroring would be detrimental to the success of the interaction: a fireman rescuing a frightened victim, a soldier facing his enemy and a doctor caring for her anxious patient. In each case, the common denominator is that it would be counterproductive to assimilate to the other’s emotional states.
Indeed, previous research has shown that modulation of participants’ neural-level responses is dependent on this connecting factor. By connecting we mean having a similar target, a shared goal with respect to the target, and by disconnecting we mean dissimilarity, or an antagonistic goal with respect to the target. These investigations compared reactions to friend vs foe, in-group vs out-group, friend vs stranger, the humanized vs the dehumanized (Harris and Fiske, 2006; Singer et al., 2006; Gutsell and Inzlicht, 2010; Masten et al., 2011; Meyer et al., 2012; Sobhani et al., 2012). The common denominator is that we may easily assimilate emotionally with friends, in-group members and fellow humans, but not in the case of our foes, the out-groups and the dehumanized. Gutsell and Inzlicht (2010) found that when participants viewed videotaped out-group members’ hand movements they showed reduced activation in the mirror neuron system. This response was graded such that the more dissimilar the viewed out-group target, the more disconnection found in mirroring. Likewise, Harris and Fiske (2006) found reduced activation in the medial prefrontal cortex, in social-cognition regions usually recruited when attributing a mind to a target person, when they viewed photographs of dehumanized individuals such as homeless or drug addicted individuals.
Furthermore, the pain matrix of the brain reacts not only to our own pain but also to the pain of others. Singer et al. (2006) had two confederates play participants in an economic game. One confederate treated the participants fairly, and the other treated the participants unfairly. After the manipulation, participants watched both confederates (fair and unfair) receive a painful electric shock. Participants’ neural activity mirrored the pain of the fair confederates, but did not mirror the pain of the unfair confederates. We suggest that much like the mirroring mechanisms in the pain matrix, that mirroring of actions should optimally cease or be reduced when observing someone with whom we are emotionally disconnected.
When we disconnect, it cannot be completely
The temporary cessation of activity in the mirror neuron system should apply only to cases in which movement is not relevant to the perceiver’s best interests. The boxer and other scenarios suggest that it should still be important to be able to predict the relevant movements of others (i.e. an incoming blow from the opponent in the ring), be they friend or foe, in-group or out-group, human or dehumanized. In all our prior examples, the relevance of the movement of the target was not manipulated—nor could it be clearly construed as relevant to the perceiver. In these cases, the target was passive—such as receiving a shock (Singer et al., 2006)—or executing a seemingly irrelevant, movement such as repeatedly reaching for the same glass of water (Gutsell and Inzlicht, 2010).
Our idea is premised on a discriminating action representation system that would mirror ubiquitously for relevant, goal-directed movements, and cease to mirror (when one cannot connect) un-purposed, purely kinematic, or extraneous movements. We are suggesting that the discrimination between relevant and irrelevant movements should occur at early stages of the action representation system, in the mirror neuron system—before this information is sent forward to the mentalizing system. This strong claim of early modulation is supported by past literature that clearly indicates that the relevance of movements modulates activity in the mirror neuron system. Responses in the mirror neuron system are stronger when the movements observed are relevant to the perceiver. Pineda and Oberman (2006) found that when participants who were smokers observed goal-directed relevant movements, such as picking up a cigarette, they showed more mirroring of those movements than participants who did not smoke. Similarly Cheng et al. (2007) found that hungry individuals showed stronger mirroring of goal-directed relevant movements towards food, compared with those who were sated.
Present aims
Here, we investigate the idea that the mirroring of movements is modulated by the perceiver’s connectedness or disconnectedness from another’s emotional states (in this case a friend vs foe scenario), as well as by the nature of the movement (goal-directed, relevant movements vs purely kinematic movements). We use the term ‘purely-kinematic’ to mean that these movements are not acting on or towards any other person (or thing) in a physically goal-relevant way. For instance, lips that are grasping at a straw to drink a soda are in goal-directed relevant movement. After gulping down some of the fizzy drink, lips that turn up at the corners to communicate pleasure are in purely kinematic movement. These purely kinematic expressions come from facial as well as body movements (de Gelder, 2006).
To test this hypothesis, we assess activity of the mirror neuron system in humans using electroencephalography (EEG). Observing and executing movement produces event-related desynchronization, also known as suppression, of the mu rhythm. The mu rhythm is an approximate 8–13 Hz oscillation produced over the sensorimotor cortex (Gastaut and Bert, 1954; Pineda, 2005). Mu suppression responses have been consistently recorded to both self and observed movements of the hand and are believed to reflect mirror neuron activity (e.g. Babiloni et al., 1999; Oberman et al., 2005; Pineda, 2005; Perry and Bentin, 2009; Gutsell and Inzlicht, 2010).
We specifically hypothesize that when observing someone with whom we wish to socially connect, mirroring responses will be found for both purely kinematic movements, such as a hand simply opening and closing, as well as to goal-relevant movements such as reaching for something that the participant desires. On the other hand, after being treated unfairly by someone, which severs the social connection to that person, we would expect that mirroring responses will cease to purely kinematic movements, but will continue to occur when movements are relevant to the perceiver, such as the grasping of an object that they themselves had just been denied.
METHOD
Participants
Twenty-five Caucasian male undergraduate students were recruited. Race and gender of participants were held constant to control for cross race or gender effects (see Discussion). The Yale University Human Subjects Internal Review Board approved this study. Data from two participants were excluded (one for improper application of the economic game manipulation, the second for loss of the raw EEG data). Thus, analysis was performed on a final sample of 23 participants (mean = 20.39 years, s.d. = 1.08 years): with 10 in the fair condition, and 13 in the unfair condition. No participants indicated knowing the confederate prior to the onset of the study.
Materials
Participants’ mood was assessed as they arrived at the lab using the 20-item Positive (α = 0.84, 10 items) and Negative (α = 0.86, 10 items) Affect Scale (PANAS; Watson et al., 1988) because present mood is an influence on how others are perceived (e.g. Clark and Isen, 1982; Bodenhausen et al., 1994). Anxiety was also measured using the State-Trait Anxiety Inventory (STAI; Spielberger et al., 1970; pre: α = 0.85, 10 items; post: α = 0.91, 10 items), both before and after the experiment. This ensured that we were manipulating the desire to connect and not more basic arousal or anxiety dimensions.
For theoretical reasons and in light of measures previously found to correlate with brain activity in the mirror neuron system (Gazolla et al., 2006) and in the pain matrix (Singer et al., 2006), we collected two measures of empathy, the Davis (1983) Interpersonal Reactivity Index (IRI), and the Empathizer Quotient Short Form (EQ; Wakabayashi et al., 2006). Considering that we were involving these individuals in a form of social interaction, we also collected measures of individual differences that could influence those interactions: the Cheek and Buss Shyness Scale (1981) and a short measure of the Big-Five personality dimensions (Gosling et al., 2003).
Participants were exposed to two blocks of five stimuli each. One block was administered before the manipulation and one after. Stimuli were presented approximately 24″ directly in front of participants. A researcher was present in the room to be sure that participants were continuously attending to the stimuli. Participants watched (i) a black background (created by a large black foam-core board) with no activity (resting trial—not included in the analysis); (ii) the confederate’s hand opening and closing, fingers to thumb in front of the black background (purely kinematic movement trial); (iii) the confederate’s hand opening and closing to grasp at a blue token (it was mounted to the reverse side of the black foam-core board so that the board could be easily flipped around by the experimenter between trials; goal-relevant trial); (iv) two white balls moving on a black background (17″ computer monitor) designed to be the same distance and trajectory as the finger to thumb movements witnessed live in the lab (Oberman et al., 2005; ball trial); and (v) the participant’s own right hand opening and closing in front of the foam-core black background (self-movement trial) (see Figure 1).
Fig. 1.

Experimental stimuli. These photographs illustrate the experimental stimuli. The resting state stimulus (top left) consisted solely of observing a black background. The purely kinematic movement (top right) was performed by the opening and closing of the hand fingers to thumb, by the participant (self-movement) and also by the confederate (purely kinematic movement). The goal-relevant movement (bottom left) illustrates the same hand motion but this time grasping a token of the same type that was used in the economic game. The ball stimuli (bottom right) consisted of a video showing two balls that moved in the trajectory and speed at which the participant and confederate moved their hands.
The confederate was trained to the rhythm of opening and closing his hand at the rate of one movement per second. It was considered to have a metronome present in the experimental setting, but in the end it was decided that it would be best to not have the additional stimulus of the ticking noise present. Great care was therefore taken, in training, so that the confederate was able to produce the same movements and to keep time at the rate of one complete movement per second. This training was refreshed over the duration of the study, and the researcher who was responsible for the coordination of each experimental session in the EEG chamber checked with the second hand of her watch to be sure that the confederate was keeping the time of one revolution per second. The confederate sat with the participant and executed the hand movements with his arm extended so that his hand would be in front of a large black board that served as a background. Participants were instructed to direct their attention to the confederate’s hand movements and not to move during observation. The researcher observed to be sure that although the participants were watching the confederate move they were not moving themselves.
All movements of the stimuli were continuous at an approximate rate of one repetition per second, or in other words, 60 repetitions in all over the 1-min presentation. The order of stimuli presentation (resting, purely kinematic, goal-relevant, ball and self-movement) was held constant across conditions, and thus cannot be responsible for the pattern of results between conditions.
The confederate was a Caucasian male from the Yale University undergraduate student body, who had studied acting. He was also prepared with a 32-electrode EEG cap and a mock connection to the amplifier. He filled out questionnaires and consent forms and sat in the same room as the participant. Throughout the study experimenters treated the confederate as a second participant. He was instructed and trained prior to the experimental sessions to maintain consistency in tone and demeanor, to interact minimally with the participant, and to respond naturally but briefly if the participant initiated communication.
The economic game was described to participants as a five-round shared resources game in which the ‘dictator’ would choose how to allocate between himself and the other individual a six-token pool of resources, and these tokens would later be traded in for a proportion of a $6.00 pot. To decide who would be the dictator, the experimenter flipped a double-headed coin rigged so that the confederate would always win the role of the dictator. In the fair condition, the confederate had been instructed to split the tokens—three for himself and three for the participant. In the unfair condition, the confederate had been instructed to split the tokens—five for himself and one for the participant. Two dictator rounds were played.
Post-experimental manipulation checks asked participants ‘Before today, did you know the other participant with whom you interacted?’ with response options: 1 = Yes, 2 = No, 3 = I've seen that person but never spoken to him or her and 4 = I am not sure. Questions were designed to capture a desire for a social connection with the confederate (α = 0.81, six items): ‘I liked the person that I interacted with today. If given the chance I would like to interact with this person again. I could see myself becoming friends with the other participant who I interacted with today. I did not care for the other participant's behavior (reversed when coded). I would not like to interact with the other participant again if it were up to me (reversed when coded). I would not make any gestures to become friends with the other participant with whom I interacted with today (reversed when coded)’. We provided participants the following response options: (1 = Very Strongly Disagree to 7 = Very Strongly Agree).
Participants were also asked about the inferences that they made about the confederate with respect to trustworthiness and warmth, as we would expect trustworthiness and warmth to be perceived as higher in the fair condition. We also measured inferences about whether the confederate was happy, sad, tense, competent, irritable, not happy or distracted. We did not expect differences on these other aspects, as we took great care to hold these other aspects constant through the confederate’s trained behavior. The confederate was to have similar demeanor and expressions of emotion whether he fairly or unfairly divided the resources. Participants first saw the stem of a sentence: ‘I found this person to be…’, the list of adjectives ran vertically, and a 5-point response scale (1 = Do Not Agree to 5 = Completely Agree) ran horizontally.1
For the dictator game, we used blue tokens made of glass; approximately 1 inch in diameter, flat on one side, and slightly rounded on the other. These glass tokens were unique and highly unlikely to hold relevant value and meaning to the participants outside of the experimental context. It was through our manipulation that the tokens were given relevant meaning as being the resource that was shared with or denied to the participant. Thus, we can interpret that reaching toward the glass token is a relevant action in the context of our experiment. One glass token was affixed to a large black piece of foam-core board by the experimenter to be used for the goal-relevant trials. The loose tokens were present during the dictator game, and then removed for the balance of the experiment.
Experimental procedure
Informed consent was obtained prior to the initiation of the experiment. After arriving individually at the lab, participants were randomly assigned to either the fair or unfair condition. The participant was prepared with an EEG cap and told we were studying individual differences in neural responses to simple hand motions and that at some point we needed to complete a quick economic game for other purposes. Participants were seated in a comfortable chair inside an electrically shielded, sound attenuated chamber beside the already similarly situated confederate and asked to fill out the two questionnaires, the PANAS and STAI.
The first experimenter, who was in the room with the participant and the confederate, coordinated the pre-manipulation stimuli presentation. Once the EEG data were collected a second experimenter entered the room, interrupted and explained that they needed to move on to the economic game. This experimenter then left the room to return to the control room. The first experimenter implemented the dictator game. In both rounds, the confederate won the coin toss and was deemed the dictator. After two rounds of the economic game the second experimenter interrupted again apologizing, and saying that instead she needed the collection of the hand movement data again and that they could resume the economic game later. At that point, the experimenter removed the loose tokens and the post-manipulation EEG data set was collected.
After EEG data collection the participant and confederate were then separated into different rooms. The participant was then asked to fill out questionnaires assessing his desire to socially connect, inferences made about the participant, the IRI, the EQ, the Big-Five Personality Index and handedness (Oldfield, 1971; collected for another investigation). The participants were asked about their suspicions of the true nature of the study. Only one participant indicated suspicion of the economic game, but he showed no awareness of the study’s hypothesis. After the experimental debriefing every participant was thanked and received a total of $16 for participation in the study.
EEG data acquisition procedure
The EEG was collected simultaneously from 32 to 9 mm tin electrodes embedded in a fabric cap using the 10–20 international method of electrode placement. Additional electrodes were applied to the face above and below the right eye and at the outer canthi of each eye as bipolar horizontal and vertical electro-oculogram (EOG), and an additional electrode applied to the chin was used as the reference electrode. The impedances on all electrodes were measured and confirmed to be between 5 and 10 kΩ. EEG was collected continuously at a sampling rate of 500 Hz with a bandpass of 0.5–100 Hz using a Compumedics amplifier with a fixed gain of 2010. Digital codes indicated that the stimulus presentation was recorded simultaneously with the continuous data. EEG was recorded and analyzed using a Neuroscan Synamps system.
Data processing
For each EEG trial, fifty-five 1 s intervals were extracted beginning 5 s after onset of the stimulus. EOG artifacts, such as eye blinks, muscle tension and movement artifacts were removed by visual inspection, and the data were then subjected to Fourier analyses using EEGLab to average the frequencies of interest, an approximate alpha wave band at 7.8–13.18 Hz. We created ratio scores of the power of the mu-rhythm across the electrodes over the hand regions of the somatosensory cortex: C3, C4, CP3 and CP4. To show that the expected activity was specific to the motor cortex electrodes, we also created the same ratio scores for the O1 and O2 electrodes that record activity from the occipital cortex. The occipital cortex modulates in alpha frequency (8–12 Hz) in accordance with attentional processing (Herrmann and Knight, 2001). We used this as a source-of-signal validation, where we would not expect to see the same pattern of results in these otherwise located electrodes. The ratio scores were: biological movement relative to non-biological movement (purely kinematic/ball trial), biological goal-relevant movement relative to non-biological movement (goal-relevant/ball trial) and lastly self-movement relative to non-biological movement (self-movement/ball trial). We then computed log10 functions of these ratio data in order to normalize the distributions, which tend to be right skewed, following protocol for mu-suppression index (Oberman et al., 2005; Oberman et al., 2008).
RESULTS
Preliminary analysis
Covariates
Participants in each condition did not differ by age, positive affect, negative affect or anxiety upon arrival at the lab. Nor did they differ on the four subscales of the IRI (fantasy, perspective taking, concern and personal distress), the Empathizer quotient, shyness or any of the NEO-5 subscales (openness, conscientiousness, extraversion, agreeableness and neuroticism), all Ps > 0.09. These factors were not different between our two groups and therefore could not then be considered an alternative explanation of our findings. They were excluded from the main analysis.
Manipulation checks
Desire to maintain a social connection to the confederate was the primary independent variable. As predicted, those in the unfair condition (M = 32.56, s.d. = 3.81) expressed less desire to maintain a social connection to the confederate than those in the fair condition (M = 36.58, s.d. = 3.20), t = −2.63, df = 19, P = 0.02, Cohen’s d = −1.14. Inferences made by participants of the confederate’s warmth and trustworthiness were as predicted and supported the efficacy of the manipulation. Participants found the confederate to be less trustworthy in the unfair condition (M = 2.59, s.d. = 0.54) than in the fair condition (M = 3.64, s.d. = 0.84), t = −3.60, P < 0.01, Cohen’s d = 1.49. Participants also found the confederate to be warmer in the fair condition (M = 3.57, s.d. = 1.07) than the in the unfair condition, (M = 2.77, s.d. = 0.68), t = −2.15, P = 0.04, Cohen’s d = 0.89. Consistent with the idea that the confederate’s demeanor was constant in both conditions, data show that no other differences were found concerning inferences of how happy, sad, tense, competent, irritable, not happy and distracted the confederate was, all Ps > 0.80.
There was a main effect of anxiety such that greater anxiety occurred before the experimental manipulation (M = 2.09, s.e. = 0.09) compared with after (M = 1.95, s.e. = 0.11), repeated measures F(1, 19) = 5.61, P = 0.03. However, the interaction between condition and time of measure was not significant (P = 0.34). There was no support for alternative explanations of the results in terms of anxiety or arousal.
Baseline mu suppression
To ensure participants showed mu suppression initially, we tested the ratio factors for self-movement, observed purely kinematic movement and observed goal-relevant action in the pre-manipulation measures. The ratio factors were created with the following formula (condition/ball) − 1 (Oberman et al., 2005), to test against a difference from zero. If the tested value is significantly greater than zero that indicates mu power amplification, and if the tested value is significantly less than zero, that indicates mu power suppression. In repeated measures (four electrodes: C3, C4, CP3 and CP4) univariate models, we found significant mu suppression when participants observed purely kinematic movements, M = −0.06, s.e. = 0.01, F(1, 23) = 6.81, P = 0.02, when participants observed goal-relevant movements, M = −0.06, s.e. = 0.01, F(1,23) = 11.19, P < 0.01, and when participants moved themselves, M = −0.10, s.e. = 0.01, F(1, 23) = 19.34, P < 0.01.
Next, we tested to see that participants in both conditions did not differ to begin with in the pre-manipulation measures of mu suppression. Again we conducted a repeated measures (four electrodes: C3, C4, CP3 and CP4) test of pre-manipulation mu suppression, but this time we compared between the two conditions by adding in the fixed factor of condition. We found no difference in pre-manipulation mu suppression when participants observed purely kinematic movements in the fair condition (M = −0.04, s.e. = 0.03) and the unfair condition (M = −0.08, s.e. = 0.03), F(1, 23) = 0.94, P = 0.34. We found no difference in pre-manipulation mu suppression when participants observed goal-relevant movements in the fair condition (M = −0.04, s.e. = 0.02) and the unfair condition (M = −0.09, s.e. = 0.03), F(1, 23) = 1.59, P = 0.22. And, we found no difference in pre-manipulation mu suppression when participants moved themselves in the fair condition (M = −0.10, s.e. = 0.03) and the unfair condition (M = −0.11, s.e. = 0.04), F(1, 23) = 0.07, P = 0.80. Likewise, mu suppression to self-movements did not differ from pre- to post-manipulation, did not differ by condition, and there was no interaction between the two factors, all Ps > 0.30. These analyses assured us that participants were indeed showing mu suppression before the manipulation of the economic game, and that this did not differ by condition.
Source-of-signal validation
The same analysis was conducted using the ratio scores for the O1 and O2 electrodes, with the prediction that we would not see the same pattern of effects in the occipital lobe location. As predicted, we found no main effect of condition (fair or unfair; P = 0.79), time of collection (pre-manipulation, post-manipulation; P = 0.61), type of action (purely kinematic-irrelevant, goal oriented; P = 0.78). We found no significant two-way interactions, all Ps > 0.35, no three-way interaction, P = 0.17 and no significant post hoc pairwise comparisons, all Ps > 0.31.
Main analysis
In the main analysis we constructed a 2 × 2 × 2 × 4 repeated measures, nested design, mixed-model in SPSS. Our between factor was condition (fair or unfair) and our within factors were repeated effects of time of collection (pre-manipulation, post-manipulation) nested in action (purely kinematic-irrelevant, goal oriented) nested in electrode (C3, C4, CP3 and CP4). We tested for main effects of condition, time and action, and all interactions. There was no main effect of condition, F(1, 23) = 0.81, P = 0.38. There was a main effect of time of collection, F(1, 345) = 13.66, P < 0.01, with pre-manipulation mu suppression greater (M = −0.06, s.e. = 0.02) than post-mu suppression (M = −0.04, s.e. = 0.02). There was also a main effect of action type, F(1, 345) = 11.45, P < 0.01 with mu suppression greater for goal-relevant movements (M = −0.06, s.e. = 0.16) than purely kinematic movements (M = −0.04, s.e. = 0.02).
The two-way interactions are reported here, but are not relevant in this research endeavor, because in each case they require collapsing across one of our three variables of interest. The results are better explained in the granularity of three-way interaction reported below. There was a significant two-way interaction between condition and time, F(1, 345) = 5.04, P = 0.03 (this is collapsed across type of action). In the unfair condition, mu suppression was greater pre-manipulation (M = −0.08, s.e. = 0.02) than post-manipulation (M = −0.04, s.e. = 0.02), P < 0.001, confidence interval (CI) of the difference lower limit (LL) = −0.06, upper limit (UL) = −0.02. However in the fair condition, there were no differences between pre-manipulation (M = −0.04, s.e. = 0.02) and post-manipulation (M = −0.03, s.e. = 0.02), P = 0.27. There was also a significant two-way interaction between condition and action, F(1, 345) = 5.57, P = 0.02 (this is collapsed across pre- and post-scores). In the unfair condition, mu suppression was greater for goal-relevant actions (M = −0.08, s.e. = 0.02) than purely kinematic actions (M = −0.04, s.e. = 0.02), P < 0.01, CI of the difference LL = −0.06, UL = −0.02. In the fair condition, there was no difference in mu suppression for goal-relevant actions (M = −0.04, s.e. = 0.02) or purely kinematic actions (M = −0.03, s.e. = 0.02), P = 0.44. There was a significant two-way interaction between time and type of action, F(1, 345) = 6.61, P < 0.01 (this is collapsed across condition). In the post-manipulation measures, mu suppression was greater for goal-relevant movements (M = −0.06, s.e. = 0.02) than purely kinematic movements (M = −0.02, s.e. = 0.02), P < 0.001, CI of the difference LL = −0.063, UL = −0.023. However in the pre-manipulation measures, there were no differences between goal-relevant movements (M = −0.07, s.e. = 0.02) and purely kinematic movements (M = −0.06, s.e. = 0.02), P = 0.57.
The interaction of interest was the three-way interaction, Condition × Time × Action. It tested whether mu suppression would be reduced after the manipulation, but only in the unfair condition, and only when those movements were of a purely kinematic nature. As predicted, the interaction was significant, F(1, 345) = 4.30, P = 0.04. Step-down comparisons found that in the unfair condition, after the manipulation, participants showed less mu suppression when viewing purely kinematic movements (M = −0.01, s.e. = 0.03) than they had before the manipulation (M = −0.08, s.e. = 0.03), P < 0.001, CI of the difference LL = −0.11, UL = −0.05. On the other hand, these same participants showed no difference between mu suppression of goal-relevant movements after the manipulation (M = −0.09, s.e. = 0.03) than before the manipulation (M = −0.08, s.e. = 0.03), P = 0.33. Furthermore, as the significant three-way interaction would suggest, this pattern of results was not the same in the fair condition. In the fair condition, there were no differences in mu suppression from when participants viewed purely kinematic movements after the manipulation (M = −0.03, s.e. = 0.02) or before (M = −0.04, s.e. = 0.05), P = 30. And there were no differences between mu suppression when participants viewed goal-relevant movements from after the manipulation (M = −0.04, s.e. = 0.02) and before (M = −0.04, s.e. = 0.02), P = 0.61 (see Figure 2).
Fig. 2.

Mu Suppression differences (from pre- to post-manipulation) between conditions (fair and unfair) and type of movement (goal-relevant and purely kinematic). This figure depicts the change in mu suppression from pre-manipulation to post-manipulation between conditions and across movement types. The error bars display the 95% confidence intervals. A score of zero indicates no change from pre- to post-scores, and negative values indicate a loss of mu suppression from pre to post. In the fair condition, there was no difference between pre-manipulation and post-manipulation mu suppression to either observed goal-relevant-relevant or purely kinematic movements. On the other hand in the unfair condition, participants had significantly reduced mu suppression to purely kinematic movements but no differences in mu suppression to goal-relevant movements.
In sum, participants in the unfair condition showed reduced mu suppression to purely kinematic movements after being treated unfairly by the confederate, but these participants continued to mirror the confederate’s goal-relevant movements of reaching for the token that they had just been denied. On the other hand, participants in the fair condition showed mu suppression of all hand movements both before and after they had been treated fairly by the confederate. These results support our theoretical position.
In addition, the data could be analyzed in another way. If our procedures in the economic game had successfully manipulated the participants’ desire to affiliate with the confederate, then a range of desires to affiliate with the confederate should have been created across participants. Thus, if our hypothesis is correct, these affiliation scores should predict mirroring activity, and we can test this hypothesis using a linear model. To examine the overall relationship of affiliation with mu suppression, we ran a repeated measures (repeated factor of four sites C3, C4, CP3 and CP4), multi-level linear model analysis predicting post-mu suppression index scores to purely kinematic hand movements from participants’ affiliation scores (standardized). Consistent with our hypothesis, self reported desire to affiliate with the confederate predicted greater mirroring activity (mu suppression), b = −0.03, β = −0.23, s.e. = 0.01, Wald = −2.97, P < 0.01 (see Figure 3).
Fig. 3.
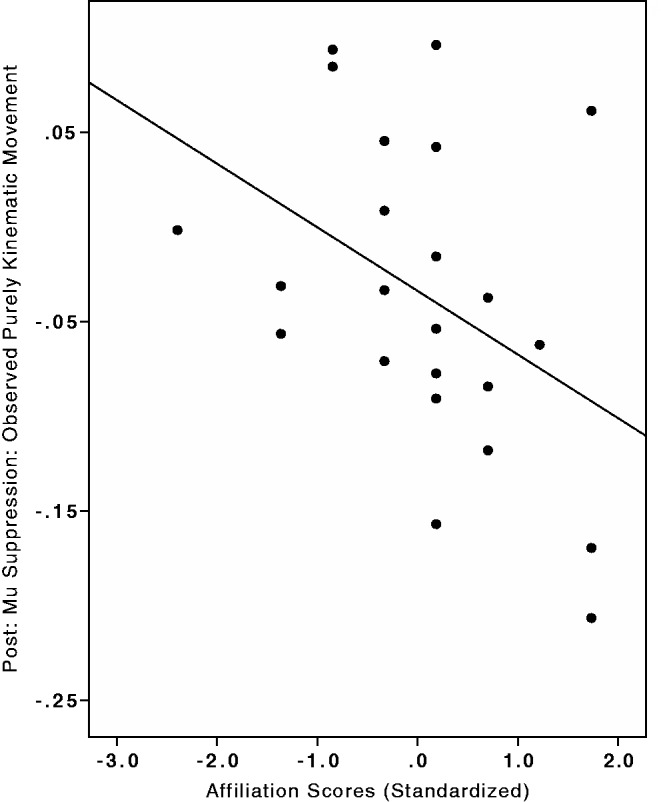
Scatterplot of desire to socially connect with the confederate and mu suppression. Self-reported scores of a desire to socially connect with the confederate predicted mirroring activity (mu suppression illustrated here is the mean score of C3, C4, CP3 and CP4), b = −0.027, β = −0.229, s.e. = 0.009, Wald = −2.97, P < 0.01.
DISCUSSION
Results from this study indicate that when participants were treated fairly they inferred that the confederate was more trustworthy and warm, and also reported that they liked him more and expressed a desire to socially connect with him further. As hypothesized, for these individuals who desired to socially connect with the confederate, mirroring responses of the confederates’ movements were found for both purely kinematic movements and for goal-relevant movements. On the other hand, when participants were treated unfairly they reported the confederate to be less trustworthy and warm and they expressed wanting less of a social connection with him in the future. These participants who had been treated unfairly, and who disliked and did not socially connect with the confederate, ceased to mirror the confederate’s movements that were not relevant to their well-being (even though they had done so before the unfair treatment), but continued to mirror movements that were relevant to their own current goals, the latter operationalized as the confederate reaching for an object that he had just denied them in an economic game. Additionally, overarching analysis of mu suppression showed that mirroring of purely kinematic movements decreased when the desire to socially connect decreased. This effect was not due to differentially allocated attentional resources because the experimenter was vigilant to be sure that the participant was observing the confederate during each and every trial, and we saw no evidence of differential attentional biases in the analysis of the alpha wave activity from the occipital cortex.
Although these data provide an important demonstration of the flexibility of the human mirror neuron system in response to diverse contextual social demands, it is important to consider several limitations of the research. The design of the experiment was a variation of a study in which empathic pain responses were diminished when viewing pain in individuals who had previously treated the observer unfairly (Singer et al., 2006). However, empathic inhibition in that study was found only to occur in male participants, implying that there may be gender-specific effects of this fair and unfair treatment paradigm.2 For this reason, we limited our participant pool to males. The role of gender-specific differences of social connection and action relevance awaits further investigation. Second, a potential generalization issue is the confederate’s lack of blindness to the condition. To control for this, experimenters took note of his behavior and had instructed him to interact minimally with all participants. Moreover, multiple manipulation checks as to the confederate’s demeanor revealed no evidence of any differential or biased demeanor in the confederate. The most important factor to consider is that we intended to manipulate the participant’s desire to affiliate with the confederate in a systematic way, and the manipulation checks show that we were effective in doing so.
Our results demonstrate that the more an individual desires a social connection with another person, the more mu suppression they exhibit to both purely kinematic and goal-relevant movements of that person. We speculate that this distinction between purely kinematic and goal-relevant movements is important because connecting with another includes taking in not only goal-relevant movements, but more specifically regarding the function of emotional assimilation, the perceiver also takes in the emotion signals that are more often than not displayed through purely kinematic movements. The use of purely kinematic movements as a proxy for emotional engagement we believe to be reasonable when one considers that emotion signals and gestures are typically of this nature—purely kinematic movements. That is not to say that all purely kinematic movements are emotional expressions, but it seems that most (if not all) emotional expressions are purely kinematic movements. In this first pass, we thought it most valuable to be sure that the movements were as similar as possible and using the same effector in each case (the hand). Future research might build upon the ideas presented in this study using multiple effectors (perhaps the mouth) and various movements (e.g. a mouth grasping a straw vs smiling) to see if what we found will hold across stimulus types.
A distinction between goal-directed movement and relevance could not be parsed out in this investigation. Past research that demonstrated modulations in mirroring dependent on relevance also used stimuli that were both goal-directed and relevant (Pineda and Oberman, 2006; Cheng et al., 2007). The experience for our participant was that he was seated directly next to a real-life person who had just treated him unfairly. This unfair person was reaching for a token of the like that the participant had just been given or denied. Not only is it fair to say that these were relevant movements but also one might go so far as to say that particularly for those who had been treated unfairly any goal-directed movements might have been relevant, because vigilance is heightened when leery of another person. We put forth that in real-life scenarios, that relevance is typically part and parcel to goal-directedness, and irrelevance is much in the same way part and parcel to purely kinematic movements. And this should be especially so when we do not desire to take-in another person for the purpose of assimilating their emotional state. Overall, we interpret that the movements we created in the goal-relevant trial were relevant because of our manipulation. If they were also relevant by their nature of being goal-directed, the hypothesis would remain the same. We would expect no modulations in mirroring responses to relevant movements, and that is what we found. It was the purely kinematic movements that differed by social connectedness.
In this investigation we chose to manipulate whether someone is ‘with us or against us’, a primary and highly important dimension of social perception (Fiske et al., 2007). However we do not believe that this is simply the case of friend or foe, and we do not intend to limit the ideas presented in this study to such cases. In the earlier example given of a caretaker and an injured child, in those moments where the child’s well-being is at stake, it is not ideal for the caretaker to assimilate that child’s pain. In this case, we would also expect that the mirroring of the child’s purely kinematic movements would cease. Indeed, a future research endeavor would investigate such an outcome in mirroring when it is necessary to take on a complementary rather than the same emotional state of someone the perceiver is close to.
The present findings do test the scenario of friend or foe, and we show that mirroring activity modulates when we do and do not wish to socially connect with others. In support of this finding, are the ideas that activity in the mirror neuron system is thought to support behavioral mimicry and imitation, and all of its consequences for social interaction, such as interpersonal bonding, cooperation in shared activities and relationship formation (Chartrand and Lakin, 2013). This investigation thus conceptually replicates the social psychology literature demonstrating that connection and disconnection modulate behavioral mimicry and imitation (Lakin and Chartrand, 2003; Lakin et al., 2003; Sonnby-Borgstrom, 2002; Sonnby-Borgstrom et al., 2003). In those studies, participants report higher liking for confederates who had spontaneously mimicked their behaviors (Chartrand and Bargh, 1999), and show greater mimicry of those whom they had higher liking (McIntosh, 2006), or who were in-group rather than out-group members (Yabar et al., 2006). There was no spontaneous mimicry for disliked targets (see Chartrand and Lakin, 2013, for review). As a note, mimicry at times has been operationalized through actions such as foot shaking, which might be considered purely kinematic in that they are not goal-directed toward an object. Still, other mimicked actions such as rubbing of the chin are goal-directed. Possibly then, as we have put forth—the distinction to be made is maybe not the goal-directedness of a movement, but the relevance of the movement to the perceiver.
The present findings thus indicate that the modulation in social connectedness is not a downstream suppression of neural mirroring activity but modulation of the activity of the mirror neuron system itself—that is, modulation of the early stages (the what and the how) of action representation. This is not to make the claim that we have pinpointed the moment at which this disconnection takes place. We are simply showing that it takes place as early as this early stage of the action representation system. In the larger scope of empathy, we are also showing that once an individual does not want to connect with another, it is not the case that they are mirroring those movements, assimilating another’s emotional states, and then quashing those feelings with a kind of cognitive emotion regulation (Gross et al., 2010). It appears that the very nature of the information sent forward to latter processing systems from the action representation system might be different when one is to assimilate vs complement.
This study begins to reconcile those instances in which it would be detrimental for the perceiver to adopt another’s emotional state, but rather more suitable to take a complementary emotional perspective. This study also conceptually replicates the social psychology literature with respect to spontaneous mimicry, which researchers consider to be related to activity in the mirror neuron system. Our findings suggest that the networks underlying action representation and simulation are flexible and sensitive to perceptual stimuli, depending on one’s desire to socially connect with the target person, and whether or not their current action has relevance to our own current goal pursuits (see Huang and Bargh, 2013).
Acknowledgments
The authors would like to thank Gregory McCarthy for the support in laboratory resources. They would also like to thank Andre Morales, Erica Reetz and Ah Joo Shin for assisting in the running of the study. This work was supported by the Yale University, Psychology Department, Graduate Dissertation Research Fund.
Footnotes
1Materials are available from the first author.
2See review paper of Taylor et al. (2000) for explanation of men’s fight or flight reactions vs women’s tend and befriend reactions to non-friendly others, which may provide insight into gender differences in response to being cheated.
REFERENCES
- Babiloni C, Carducci F, Cincotti F, et al. Human movement-related potentials vs. desynchronization of EEG alpha rhythm: a high-resolution EEG study. NeuroImage. 1999;10:658–65. doi: 10.1006/nimg.1999.0504. [DOI] [PubMed] [Google Scholar]
- Belot M, Crawford VP, Heyes C. Players of Matching Pennies automatically imitate opponents’ gestures against strong incentives. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(8):2763–8. doi: 10.1073/pnas.1209981110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bodenhausen GV, Sheppard LA, Kramer GP. Negative affect and social judgment: the differential impact of anger and sadness. European Journal of Social Psychology. 1994;24:45–62. [Google Scholar]
- Chartrand TL, Bargh JA. The chameleon effect: the perception-behavior link and social interaction. Journal of Personality and Social Psychology. 1999;76:893–910. doi: 10.1037//0022-3514.76.6.893. [DOI] [PubMed] [Google Scholar]
- Chartrand TL, Lakin JL. The antecedents and consequences of human behavioral mimicry. Annual Review of Psychology. 2013;64:285–308. doi: 10.1146/annurev-psych-113011-143754. [DOI] [PubMed] [Google Scholar]
- Cheek JM, Buss AH. Shyness and sociability. Journal of Personality and Social Psychology. 1981;41:330–9. [Google Scholar]
- Cheng Y, Meltzoff AN, Decety J. Motivation modulates the activity of the human mirror-neuron system. Cerebral Cortex. 2007;17(8):1979–86. doi: 10.1093/cercor/bhl107. [DOI] [PubMed] [Google Scholar]
- Clark MS, Isen AM. Toward understanding the relationship between feeling states and social behavior. Cognitive social psychology. 1982;73:108. [Google Scholar]
- Davis MH. Measuring individual differences in empathy: evidence for a multidimensional approach. Journal of Personality and Social Psychology. 1983;44(1):113–26. [Google Scholar]
- de Gelder B. Towards the neurobiology of emotional body language. Nature Reviews Neuroscience. 2006;7(3):242–9. doi: 10.1038/nrn1872. [DOI] [PubMed] [Google Scholar]
- di Pellegrino G, Fadiga L, Fogassi L, Gallese V, Rizzolatti G. Understanding motor events: a neurophysiological study. Experimental Brain Research. 1992;91:176–80. doi: 10.1007/BF00230027. [DOI] [PubMed] [Google Scholar]
- Eisenberg N, Fabes RA, Miller PA, et al. Relation of sympathy and personal distress to prosocial behavior: a multimethod study. Journal of Personality and Social Psychology. 1989;57(1):55. doi: 10.1037//0022-3514.57.1.55. [DOI] [PubMed] [Google Scholar]
- Esteves F, Dimberg U, Öhman A. Automatically elicited fear: conditioned skin conductance responses to masked facial expressions. Cognition & Emotion. 1994;8(5):393–413. [Google Scholar]
- Fiske ST, Cuddy AJC, Glick P. Universal dimensions of social perception: warmth and competence. Trends in Cognitive Science. 2007;11:77–83. doi: 10.1016/j.tics.2006.11.005. [DOI] [PubMed] [Google Scholar]
- Gastaut HJ, Bert J. EEG changes during cinematographic presentation. Electroencephalography and Clinical Neurophysiology. 1954;6:433–44. doi: 10.1016/0013-4694(54)90058-9. [DOI] [PubMed] [Google Scholar]
- Gazzola V, Aziz-Zadeh L, Keysers C. Empathy and the somatotopic auditory mirror system in humans. Current Biology. 2006;16:1824–9. doi: 10.1016/j.cub.2006.07.072. [DOI] [PubMed] [Google Scholar]
- Gosling SD, Rentfrow PJ, Swann WB., Jr A very brief measure of the Big-Five personality domains. Journal of Research in Psychology. 2003;37:504–28. [Google Scholar]
- Gross JJ, Sheppes G, Urry HL. Emotion generation and emotion regulation: a distinction we should make (carefully) Cognition & Emotion. 2010;25(5):765–81. doi: 10.1080/02699931.2011.555753. [DOI] [PubMed] [Google Scholar]
- Gutsell JN, Inzlicht M. Empathy constrained: prejudice predicts reduced mental simulation of actions during observation of out-groups. Journal of Experimental Social Psychology. 2010;46(5):841–5. [Google Scholar]
- Harris LT, Fiske ST. Dehumanizing the lowest of the low neuroimaging responses to extreme out-groups. Psychological Science. 2006;17(10):847–53. doi: 10.1111/j.1467-9280.2006.01793.x. [DOI] [PubMed] [Google Scholar]
- Herrmann CS, Knight RT. Mechanisms of human attention: event-related potentials and oscillations. Neuroscience and Biobehavioral Reviews. 2001;25(6):465–76. doi: 10.1016/s0149-7634(01)00027-6. [DOI] [PubMed] [Google Scholar]
- Huang JY, Bargh JA. The selfish goal: autonomously operating motivational structures as the proximate cause of human judgment and behavior. Behavioral and Brain Sciences. 2013 doi: 10.1017/S0140525X13000290. [DOI] [PubMed] [Google Scholar]
- Iacoboni M, Molnar-Szakacs I, Gallese V, Buccino G, Mazziotta JC, Rizzolatti G. Grasping the intentions of others with one’s own mirror neuron system. PLoS Biology. 2005;3:79. doi: 10.1371/journal.pbio.0030079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keysers C, Wicker B, Gazzola V, Anton JL, Fogassi L, Gallese V. A touching sight: SII/PV activation during the observation and experience of touch. Neuron. 2004;42:335–46. doi: 10.1016/s0896-6273(04)00156-4. [DOI] [PubMed] [Google Scholar]
- Lakin JL, Chartrand TL. Using nonconscious behavioural mimicry to create affiliation and rapport. Psychological Science. 2003;14(4):334–9. doi: 10.1111/1467-9280.14481. [DOI] [PubMed] [Google Scholar]
- Lakin JL, Jefferis VE, Cheng CM, Chartrand TL. The chameleon effect as social glue: evidence for the evolutionary significance of nonconscious mimicry. Journal of Nonverbal Behaviour. 2003;27(3):145–62. [Google Scholar]
- Masten CL, Morelli SA, Eisenberger NI. An fMRI investigation of empathy for ‘social pain’ and subsequent prosocial behavior. Neuroimage. 2011;55(1):381–8. doi: 10.1016/j.neuroimage.2010.11.060. [DOI] [PubMed] [Google Scholar]
- McIntosh DN. Spontaneous facial mimicry, liking and emotional contagion. Polish Psychological Bulletin. 2006;37:31–42. [Google Scholar]
- Meyer ML, Masten CL, Ma Y, et al. Empathy for the social suffering of friends and strangers recruits distinct patterns of brain activation. Social Cognitive and Affective Neuroscience. 2012;8:446–54. doi: 10.1093/scan/nss019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberman LM, Hubbard EM, McCleery JP, Altschuler EL, Ramachandran VS, Pineda JA. EEG evidence for mirror neuron dysfunction in autism spectrum disorders. Cognitive Brain Research. 2005;24:190–8. doi: 10.1016/j.cogbrainres.2005.01.014. [DOI] [PubMed] [Google Scholar]
- Oberman LM, Ramachandran VS, Pineda JA. Modulation of mu suppression in children with autism spectrum disorders in response to familiar or unfamiliar stimuli: the mirror neuron hypothesis. Neuropsychologia. 2008;46(5):1558–65. doi: 10.1016/j.neuropsychologia.2008.01.010. [DOI] [PubMed] [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Perry A, Bentin S. Mirror activity in the human brain while observing hand movements: a comparison between EEG desynchronization in the range and previous fMRI results. Brain Research. 2009;1282:126–32. doi: 10.1016/j.brainres.2009.05.059. [DOI] [PubMed] [Google Scholar]
- Pineda J. The functional significance of mu rhythms: translating ‘seeing’ and ‘hearing’ into ‘doing’. Brian Research Reviews. 2005;50:57–68. doi: 10.1016/j.brainresrev.2005.04.005. [DOI] [PubMed] [Google Scholar]
- Pineda J, Oberman LM. What goads cigarette smokers to smoke? Neural adaptation and the mirror neuron system. Brain Research. 2006;1121(1):128–35. doi: 10.1016/j.brainres.2006.08.128. [DOI] [PubMed] [Google Scholar]
- Singer T, Seymour B, O’Doherty J, Klaas ES, Dolan JD, Frith C. Empathic neural responses are modulated by the perceived fairness of others. Nature. 2006;439:466–9. doi: 10.1038/nature04271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sobhani M, Fox GR, Kaplan J, Aziz-Zadeh L. Interpersonal liking modulates motor-related neural regions. PLoS One. 2012;7(10):e46809. doi: 10.1371/journal.pone.0046809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnby-Borgstrom M. Automatic mimicry reactions as related to differences in emotional empathy. Scandinavian Journal of Psychology. 2002;43:433–43. doi: 10.1111/1467-9450.00312. [DOI] [PubMed] [Google Scholar]
- Sonnby-Borgstrom M, Johnsson P, Svensson O. Emotional empathy as related to mimicry reactions at different levels of information processing. Journal of Nonverbal Behaviour. 2003;27(1):3–23. [Google Scholar]
- Spielberger CD, Gorsuch RL, Lushene RE. Manual for the State-Trait Anxiety Inventory. Palo Alto, CA: Consulting Psychologists Press; 1970. [Google Scholar]
- Spunt RP, Lieberman MD. Dissociating modality-specific and supramodal neural systems for action understanding. The Journal of Neuroscience. 2012;32(10):3575–83. doi: 10.1523/JNEUROSCI.5715-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spunt RP, Satpute AB, Lieberman MD. Identifying the what, why, and how of an observed action: an fMRI study of mentalizing and mechanizing during action observation. Journal of Cognitive Neuroscience. 2011;23(1):63–74. doi: 10.1162/jocn.2010.21446. [DOI] [PubMed] [Google Scholar]
- Taylor SE, Klein LC, Lewis BP, Gruenewald TL, Gurung RA, Updegraff JA. Biobehavioral responses to stress in females: tend-and-befriend, not fight-or-flight. Psychological Review. 2000;107(3):411. doi: 10.1037/0033-295x.107.3.411. [DOI] [PubMed] [Google Scholar]
- Thioux M, Gazzola V, Keysers C. Action understanding: how, what and why. Current Biology. 2008;18(10):R431–4. doi: 10.1016/j.cub.2008.03.018. [DOI] [PubMed] [Google Scholar]
- Wakabayashi A, Baron-Cohen S, Wheelwright S, et al. Developing of short forms of the empathy quotient (EQ-Short) and the systemizing quotient (SQ-Short) Personality and Individual Differences. 2006;41:929–40. [Google Scholar]
- Watson D, Clark LA, Tellegen A. Development and validation of brief measures of positive and negative affect: the PANAS scales. Journal of Personality and Social Psychology. 1988;54(6):1063–70. doi: 10.1037//0022-3514.54.6.1063. [DOI] [PubMed] [Google Scholar]
- Williams J, Waiter G, Gilchrist A, Perrett D, Murray A, Whiten A. Neural mechanisms of imitation and ‘mirror neuron’ functioning in autism spectrum disorder. Neuropsychologia. 2006;44:610–21. doi: 10.1016/j.neuropsychologia.2005.06.010. [DOI] [PubMed] [Google Scholar]
- Yabar Y, Johnston L, Miles L, Peace V. Implicit behavioural mimicry: investigating the impact of group membership. Journal of Nonverbal Behaviour. 2006;30(3):97–113. [Google Scholar]