

Abstract
The functions of the gastrointestinal (GI) tract include digestion, absorption, excretion, and protection. In this review, we focus on the electrical activity of the stomach and small intestine, which underlies the motility of these organs, and where the most detailed systems descriptions and computational models have been based to date. Much of this discussion is also applicable to the rest of the GI tract. This review covers four major spatial scales: cell, tissue, organ, and torso, and discusses the methods of investigation and the challenges associated with each. We begin by describing the origin of the electrical activity in the interstitial cells of Cajal, and its spread to smooth muscle cells. The spread of electrical activity through the stomach and small intestine is then described, followed by the resultant electrical and magnetic activity that may be recorded on the body surface. A number of common and highly symptomatic GI conditions involve abnormal electrical and/or motor activity, which are often termed functional disorders. In the last section of this review we address approaches being used to characterize and diagnose abnormalities in the electrical activity and how these might be applied in the clinical setting. The understanding of electrophysiology and motility of the GI system remains a challenging field, and the review discusses how biophysically based mathematical models can help to bridge gaps in our current knowledge, through integration of otherwise separate concepts.
The gastrointestinal (GI) system has a number of sophisticated and autonomous functions coordinated over a range of length and time scales. Recent years have seen major advances in determining the mechanisms and interactions responsible for these functions. A substantial challenge involves reintegrating this detailed knowledge into coherent descriptions of single cell, tissue, and organ function, and this review highlights significant early progress towards this goal.
The primary functions of the GI tract are digestion, absorption, excretion, and protection. These functions are achieved through a series of organs with distinct roles from mouth to anus. The stomach and small intestine are principally responsible for digestion and absorption, a process incorporating both physical (e.g., retropulsion in the stomach) and chemical (e.g., bile and enzymes in the small intestine) mechanisms. The large intestine is primarily concerned with desiccation and compaction of waste, with storage in the sigmoid colon and rectum prior to elimination.
Describing and predicting the behavior of integrated systems over multiple scales is a complex task, and physiologists now commonly approach this challenge with the aid of mathematical computational modeling. Recognizing the potential of this approach, the International Union of Physiological Sciences has fostered the Physiome Project to progress these modeling aims.1 The term comes from ‘physio-’ (life) and ‘-ome’ (as a whole). The Physiome Project is a worldwide public domain effort to provide a computational framework for understanding the physiology of an individual or species. It aims to develop integrative models at all levels of biological organization, from genes to the entire organism via gene regulatory networks, protein pathways, integrative cell function, tissue, and whole organ structure/function relations. Multiscale GI models are thus a key strategy in forming integrative systems descriptions of GI function, and models of gastric motility are emphasized in this review.
The review is organized into a scale of hierarchies: cellular and subcellular relationships are considered first, followed by tissue level, organ, and lastly, whole body relationships, before clinical directions are discussed (as illustrated in Figure 1). This organization mirrors our present ‘multiscale’ knowledge of physiological events such as peristalsis, which encompasses vast spatial and temporal scales from nanometers and microseconds (such as, voltage-gated ion channel behavior) to centimeters and hours (for example, meal transit time through the stomach).1
FIGURE 1.
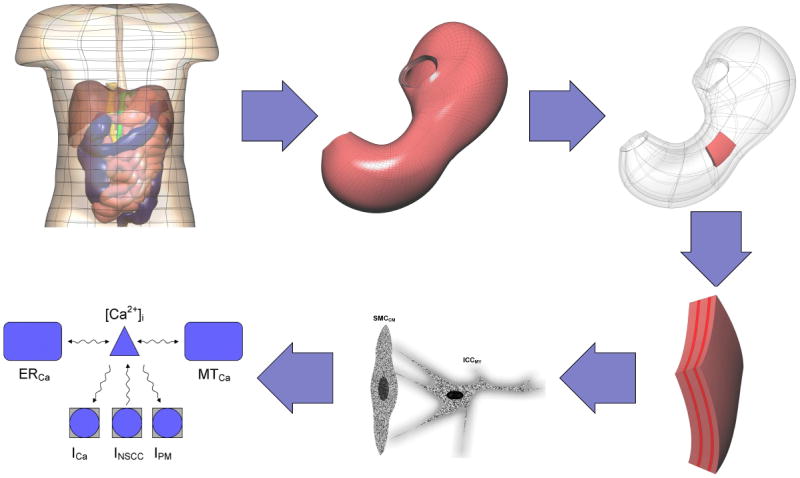
Illustration of the different spatial scales of models involved in the representation of the gastrointestinal system. An anatomical torso geometry is represented at the broadest scale. An anatomical model of the human stomach is an example of the models at organ level, while at the tissue level, the different muscle layers and their cell types must be represented. At the cellular level, smooth muscle cells in the circular direction (SMCCM) and interstitial cells of Cajal (ICCMY) are shown. The intracellular calcium [Ca2+]i signaling pathways in the Faville et al.2 ICC model are illustrated as an example of subcellular level models. The Faville et al2 ICC model states that [Ca2+]i is modulated by a number of transmembrane Ca2+ specific ion channels, which include an inward Ca2+ (ICa), non-selective cation channel (ISNCC), and Ca2+-ATPase (IPM). In addition, [Ca2+]i sequestration is also influenced at the subcellular level by the actions of the endoplasmic reticulum (ER) and mitochondria (MT).
This review will focus on the research of the electrical activity of the stomach and small intestine, and more specifically the underlying electrical activity that organizes motility. This is the area where the most detailed systems descriptions and mathematical models of GI function have been based to date,3-5 and from where efforts are beginning to yield clinically relevant outcomes (e.g., Ref 6). As other GI tract organs share a similar pattern of organization, this review will be of relevance to future research directions for the GI system as a whole.
CELLULAR LEVEL
GI motility is underpinned by omnipresent electrical activity. The fundamental event in the stomach and small intestine is the ‘slow wave’, a continuous, undulating change in membrane potential that propagates through the GI musculature in a coordinated fashion.7 Slow waves serve to induce and organize phasic contractions8 by moving the smooth muscle cell (SMC) membrane potential from a state of low open-probability for voltage-dependent Ca2+ channels (−80 to − 55 mV) to a potential with elevated probability of channel opening (−40 to − 25 mV).7 Sufficient Ca2+ influx in the wake of electrical slow waves may lead to SMC contraction, depending on additional regulatory input from neuronal, hormonal, paracrine, and inflammatory signals (see Ref 9 for more detail).
We now know that the interstitial cells of Cajal (ICCs) and not the SMCs, such as those illustrated in Figure 2, are responsible for initiating and propagating slow waves.7 Experiments with W/Wv mice which have a c-kit (a receptor necessary for the normal development of ICCs) mutation have revealed a decrease in particular ICC populations and a corresponding lack of intestinal slow wave activity.10 The mechanism by which ICCs are thought to generate slow waves is via the summation of many small amplitude membrane fluctuations termed ‘unitary potentials’ (UPs), which result from inward-directed ionic conductances through unique pacemaker channels in ICC cell membranes. When a certain depolarization threshold is reached within ICCs, UP coordination and summation leads to a rapid upstroke depolarization, followed by a regenerative potential reflected by the plateau phase of the slow wave.7 Because UP summation is achieved via a voltage-dependent mechanism, depolarization of adjacent interconnected ICCs leads to cell-to-cell entrainment and active propagation of slow wave events through ICC networks.2, 7
FIGURE 2.
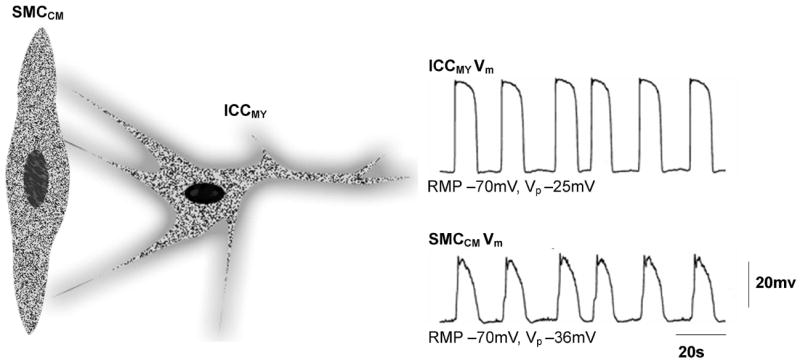
Schematic diagram of an Interstitial Cell of Cajal (ICC) and an associated smooth muscle cell (SMC) of the circular muscle (CM) along with membrane potential (Vm) traces from an ICC and SMC. Both the ICC and SMC membrane activities demonstrate a periodicity of 3 cpm. The amplitude of the ICC membrane potential (~45 mV) is higher than that of the SMC (~34 mV). The membrane potential traces are reproduced from Ref 11.
Simultaneous intracellular recordings have confirmed that slow waves begin in ICCs before conducting passively to adjacent SMCs to which they are weakly coupled through sparse gap junctions forming a functional syncytium (behaving with cytoplasmic continuity).12 SMCs are unable to actively propagate slow waves, such that, without ICCs, slow waves rapidly decay in GI tissue in accordance with the conduction properties of the smooth muscle syncytium.13 During the plateau phase of slow waves, high-frequency (>30 cpm) oscillations of membrane potentials often referred to as ‘spikes’ may occur in SMCs.14 These threshold events represent true smooth muscle action potentials. Spikes are not essential for excitation–contraction coupling in GI smooth muscle, but their occurrence is associated with more forceful contractions.9
Computer-based mathematical models have proved to be useful tools for characterizing integrated GI cellular electrophysiology. The approaches that have been used to model the cellular events in ICCs and SMCs can be generalized into two categories: phenomenological models and biophysically based models. Phenomenological models reproduce the electrical events using relatively simple mathematical equations without a direct relationship with the underlying electrophysiology. Since the 1960s, GI electrical activity (GIEA) has been modeled as a chain of coupled Van der Pol relaxation oscillators.15 Sarna et al. expanded on the relaxation-oscillator concept to simulate gastric and intestinal electrical activity using a network of coupled oscillators.16, 17 These early relaxation oscillator models were acknowledged as a sufficient way to represent the GIEA for the subsequent two decades, until discovery of the role of ICCs raised questions about the validity of using only one type of coupled network to represent the GIEA. In a more recent review by Daniel et al.,18 the question was raised as to what the intrinsic periodic activity of the oscillator unit actually represented. As the pacesetting role of ICCs was becoming evident, an updated oscillator-based model was proposed by Aliev et al.19 Most importantly, this model acknowledged for the first time the differentiated functions of ICCs and SMCs as two interconnected electrical domains (see the ICC response in Figure 3). However, the limitations intrinsic to the relaxation oscillator-based models were still evident. In particular, Publicover and Sanders20 argued that the plasticity of the relaxation oscillator waveform limited it from modeling the experimentally recorded membrane potentials, and the effects of pharmaceutical agents cannot be investigated via phenomenological models.
FIGURE 3.
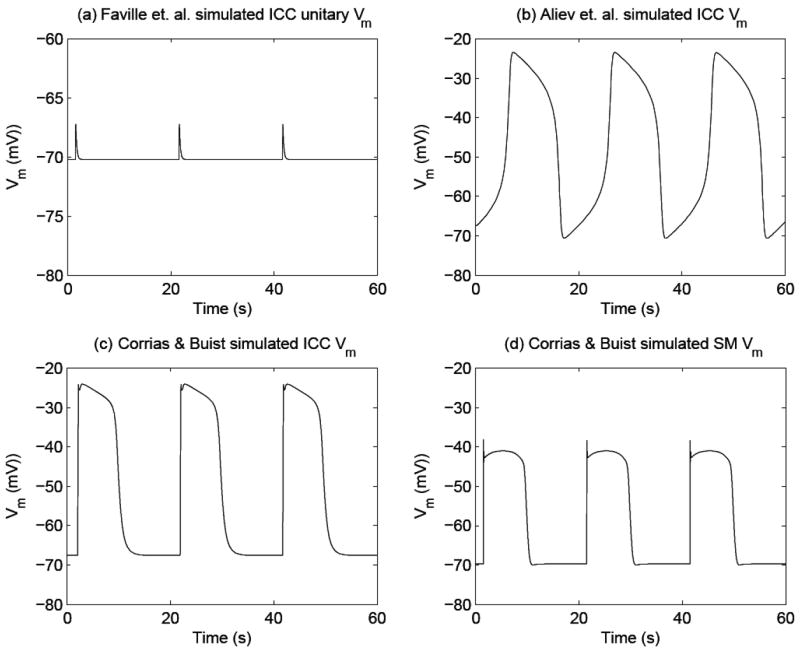
Simulated gastric electrical activity (GEA) using computer-based mathematical models. Shown in (a) is simulated unitary potentials (UPs) which are believed to generate pacemaker potentials in summation, using the Faville et al.2 ICC model. The simulated UPs contain an autonomous frequency of 3 cpm, amplitude of 3 mV, and resting Vm of 70 mV. Shown in (b) is the simulated membrane potential (Vm) of an ICC, which is known as the pacemaker potentials, using the Aliev et al.19 model. The simulated pacemaker potentials contain an autonomous frequency of 3 cpm. The peak and resting membrane potential (Vm) have to be scaled in order to match experimental data, as the Aliev model is a phenomenological model. Shown in (c) are simulated pacemaker potentials using the Corrias and Buist21 ICC model. The simulated pacemaker potential has an autonomous frequency of 3 cpm, an amplitude of 45 mV, and a resting Vm of -70 mV. Shown in (d) is the simulated Vm of canine gastric smooth muscle cells (SMCs), which is also known as slow waves, using the Corrias and Buist22 SMC model. This cell model requires a pacemaker potential as an input to depolarize the Vm (an output of the SMC model). The simulated slow wave has a frequency of 3 cpm, amplitude of 35 mV, and resting Vm of -70 mV.
Biophysically based models take into consideration the roles of ion gating variables in cellular action potentials. Much progress has been made in recent decades on simulations of nerve and cardiac electrical activities.23-25 As details of ion channels and intracellular activities of ICCs and SMCs have begun to be understood over recent years, a small number of biophysically based models of GIEA have been proposed. This reflects a trend towards modeling the GIEA from the cellular level using the core conduction (biophysically based) models proposed initially by Hodgkin and Huxley in 1952.23 In general, the biophysically based models allow individual ion currents to be inspected quantitatively under effects of parameters which carry physical quantities, such as temperature, ion concentration, and voltage, which would not be possible with the relaxation oscillator-based models.
Edwards and Hirst26 first proposed an ICC model based on unitary potential theory. The ion channels of the Edwards and Hirst ICC model are biophysically based. However, the unitary potentials in this model had a phenomenological representation, and therefore, the intracellular Ca2+ dynamics could not be captured with this model. Currently, there are three whole-cell biophysically based models specifically for simulating ICC pacemaker activity. The Youm et al.27 ICC model outlined seven ion conductances categorized into three ionic components (Ca2+, K+, and Na+) and a series of intracellular Ca2+ dynamics based on the Luo and Rudy25 (cardiac) approach to simulate intestinal pacemaker potentials. The Corrias and Buist21 ICC model described 10 ion conductances and the intracellular Ca2+ dynamics based on the mitochondria–ER relationship proposed by Fall and Keizer.28 The Faville et al.2 model is the first ICC model that contains a biophysically based representation of unitary potentials (see Figure 3). This model contains a series of Ca2+ dynamics derived from experimental data specifically derived from ICCs, and therefore, has more authenticity in terms of the intracellular Ca2+ dynamics relative to the other two biophysically based ICC models. In contrast, there is only one biophysically based GI SMC model to date, which consists of eight ion conductances.22
Integration of different types of mathematical models is essential for building an appropriate systems-based understanding, for example, the SMC model requires an ICC cell model to provide the pacemaker potential. The CellML language provides a medium to standardize the scripting of models through an open-source XML markup language database.29 CellML aims to simplify the exchange of computer-based mathematical models between different software platforms. It also enables them to reuse components from one model in another, thus accelerating model building. At present, there are a large number of different cellular descriptions available in the CellML model repository, with the majority relating to electrophysiology, calcium dynamics, and signal transduction, although, as noted above, there are only a handful of models that are related to the GI system. Currently the models in the CellML repository include the Faville et al.,2 Youm et al.,27 Corrias and Buist22 ICC models and the Corrias and Buist21 SM cell models.
TISSUE LEVEL
The above GI cellular elements described above, ICCs and SMCs, are anatomically united at the tissue level in a myenteric network capable of sophisticated autonomous behavior, as shown in Figure 4. The mucosa and submucosa do not directly contribute to motility, and have not been considered in tissue level models of motility. SMCs in the GI muscularis propria are organized in bundles of >1000 parallel fibers, orientated in circular (CM), longitudinal (LM) and oblique (OM; stomach only) directions.30
FIGURE 4.
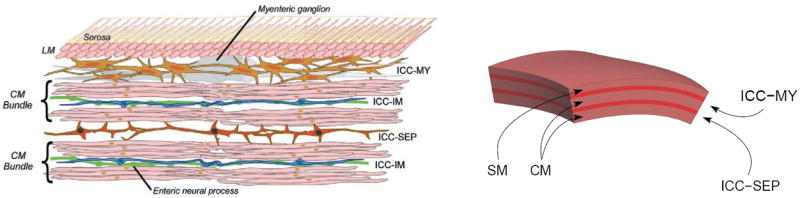
Schematic diagram illustrating the organization of Interstitial cells of Cajal (ICCs) and smooth muscle (SM) cells in the canine gastric antrum.31 SM are arranged in the longitudinal direction (LM) and the circular direction (CM) of the stomach. ICCs are distributed between the intermittent spaces of SM, which include myenteric ICCs (ICCMY), intramuscular ICCs (ICCIM), and septal ICCs (ICCSEP). Also shown on the right is the representation of the ICC and SM cells in an anatomically based computational model.
Different classes of ICCs have been identified by morphology, location, and function. The most relevant class to this review is ICC-MY, which are arranged in a network between the SM layers. The ICC-MY are normally responsible for slow wave generation and propagation. Another class of ICCs intermixed with SM fibers (ICC-IM) modulate and augment the response of SMCs to the ICC-MY-generated slow waves. ICC-IM also play an important role in the integration of enteric nervous signals, and may transduce neural or stretch signals to mediate changes in slow wave frequency (Refs 7,32 for more detail). In addition, ICC-IM are thought to have the capability to act as the pacemakers cells, instead of ICC-MY, under certain circumstances, such as vagal stimulation.33
In an intact ICC network, slow waves entrain in accordance with the site of highest intrinsic ICC pacemaker frequency found in the network.7 Slow waves, thereafter, spread to adjacent ICC and SM layers in accordance with the three-dimensional structure of the cellular tissue networks. Slow waves propagate simultaneously along SM fibers in the circumferential and longitudinal directions. However measurements in isolated tissue samples demonstrate that circumferential slow wave velocity is approximately 23 mm/s compared to approximately 11 mm/s in the longitudinal direction34 (refer to Figure 5). The net effect of these different propagation velocities is to create a transverse dominant contraction that spreads distally through tissue as a coordinated peristaltic wave front.
FIGURE 5.
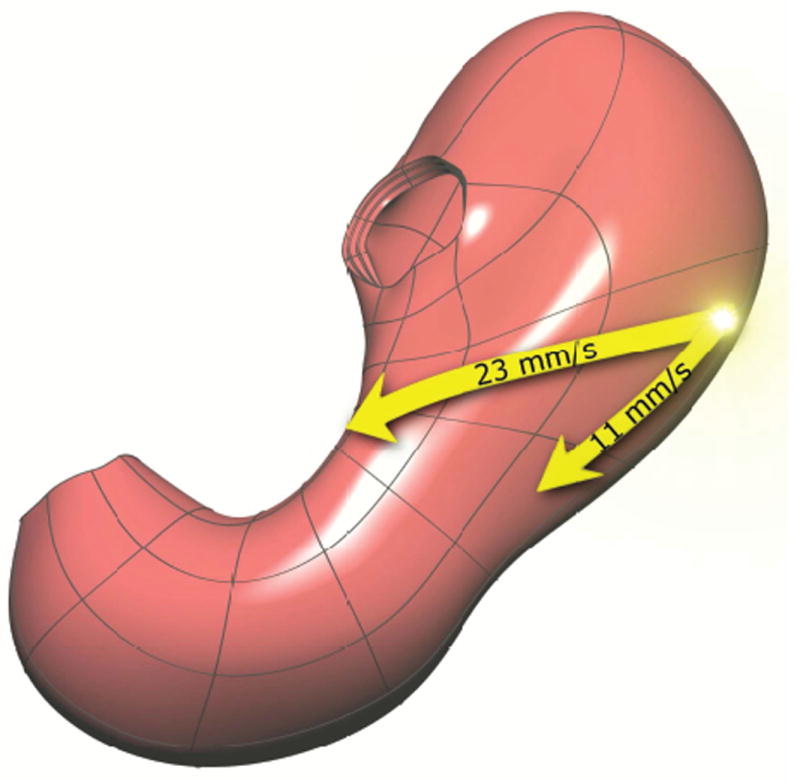
Illustration of the origin and relative propagation of the gastric slow waves. The slow wave originates in the mid-corpus region on the greater curvature and rapidly spreads in a circumferential dominant manner around the stomach, as well as propagating at a slower rate down the length of the stomach.34.
Experimental slow wave measurements at the cellular level have involved fine intracellular recordings describing membrane potentials, intrinsic frequencies, and ion conductances.11 At the tissue level, experimental recordings offer a macroscopic view (up to many cm) of the profile and patterns of slow wave propagation. The common serosal recording technique has involved sparse placement of typically four to eight electrodes along electrically active regions of the GI tract.35 More recently, Lammers et al.36 have significantly expanded GI electrical tissue mapping capability using high-resolution electrode arrays containing up to 256 electrodes recording simultaneously (discussed further below), allowing detailed spatiotemporal mapping of slow wave activity.
Continuum modeling may be employed to represent the electrical activity within the musculature of the tissue. The bidomain equations have been used to govern the spread of electrical activity within excitable tissue since the late 1960s.37-40 These equations describe the flow of ions within and between two domains (the volume within the cells and the remainder of the volume external to the cells). The bidomain framework has been widely used within the cardiac field for many years,41-46 however, it has only recently been applied to the GI system.5, 6 In the computational framework presented by Pullan et al.,3 the different muscle layers have been subdivided into layers corresponding to the anatomical ICC and SMC layers and shown in Figure 4.
ORGAN AND TORSO LEVEL
The stomach is the most dilated portion of the GI tract, and communicates proximally via the cardiac orifice with the esophagus, and distally with the duodenum via the pylorus. Three other gastric regions are typically described: the proximal distensible fundus, the distal funnel-like antrum, and the corpus of the stomach between the fundus and antrum (see Ref 47).
The pathway of normal and abnormal gastric slow wave propagation has not been extensively characterized between gastric regions, but a general description is known. There is no gastric ‘pacemaker node’ akin to the cardiac pacemaker nodes, but rather, slow waves are thought to arise in a diffuse pacemaker zone corresponding to that part of the ICC network with the highest intrinsic rate of pacemaking activity.7 This zone (shown in Figure 5) is located near the greater curvature half of the orad corpus.48 The fundus has been termed electrically ‘silent’, being free of slow wave activity, however at the cellular level, slow fluctuations of a relatively depolarized resting membrane potential are found that give rise to tonic muscular functions.7 From the greater curvature, slow waves track toward the pylorus at approximately 3 cycles/minute49 with increasing velocity and amplitude.50 A quiescent zone has recently been identified near the pylorus, where no slow waves can be recorded,51 and in which ICC-MY are found in significantly reduced quantities.52 This zone represents an effective electrical barrier, where gastric slow waves are decoupled from small intestinal slow waves. Spike activity may, however, traverse this area, presenting a possible means of gastroduodenal coordination.53
The small intestine begins distal to the pylorus. It is comprised of three sections (due denum, jejunum and ileum) with an average length of 4–6 m. Digesting contents (chyme) are propelled distally at 5–20 mm/s by weak peristaltic contractions, thereby taking 3–5 h to progress from the pylorus to the ileocaecal valve.54 The small intestine has intrinsic pacemaker activity along its length, decreasing from an intrinsic rate of approximately 12 cpm in the duodenum to 8–9 cpm in the terminal ileum.55, 56 It was previously thought that slow wave activity in the small intestine propagates only over a few centimeters at a time, naturally organizing small intestinal motility into segmental contractions, serving to slowly mix and spread chyme for digestion.7 However, a recent multichannel (240 electrode) study57 performed on the feline small intestine reported that the majority (73%) of slow waves did propagate continuously from the proximal duodenum to the ileocaecal junction, with conduction of the remaining waves being blocked, mostly at certain specific sites along the tract. The segmentation patterns that characterize small intestinal activity, therefore, likely result from the limited propagation of individual ‘spikes’, which occur in the wake of slow waves.53
In recent years, anatomically realistic models of the stomach have been developed with the aim of building an integrated physiological description of the spatial orientation of slow wave activity.3 These models have been integrated with tissue and cell-level models, and are beginning to demonstrate predictive capability with application to the diagnosis and treatment of gastric diseases.58 A gradient of intrinsic pacemaker potential frequency is assigned to the ICC layer of these mathematical models, to reproduce the propagation of entrained slow waves in the smooth muscle layer. It can be shown that the pacemaker potentials with lower frequency ‘phase-lock’ to those of higher frequency. Active propagation of slow waves is achieved once the ‘phase-locking’ becomes constant (entrainment). Future mathematical models of small bowel activity must faithfully represent the new multielectrode understanding of slow wave propagation and block, as described above.
Anatomical models of GI organs have been reported since the 1970s. The earlier methods of quantitatively capturing the anatomy of the intestines involved photographing of the in vitro duodenum during longitudinal smooth muscle contractions.59 As more advanced medical imaging modalities became available, the in vivo stomach configurations were captured using plain radiography, ultrasound, and in recent years, magnetic resonance (MR) imaging.60, 61 However, registration of the individual images remains a challenge to the reconstruction of a three-dimensional anatomical model. Many computational techniques have been proposed to circumvent the problem of image registration. In particular, Kita62 used an elastic stomach model to project the distorted contours of individual images onto the model, thereby identifying the same parts of the stomach in x-ray images. Liao et al.63 fitted a three-dimensional linear mesh of a rat stomach and then used an algorithm to smooth the surface of the stomach model.
To date, the Visible Human Project64 remains one of the best sources of high-resolution images of human anatomy, and it has been widely used for numerical simulations of human function. The outlines of the stomach in the Visible Human abdominal slices has been digitized and stacked to form a data cloud of the shape of the stomach, which has then been iteratively fitted using cubic Hermite basis functions and a nonlinear fitting procedure to create an anatomical stomach model.3, 65 The Visible Human data has also been used to model the anatomy of the gastroesophageal-junction,66 the intestines,67 and the pelvic floor.68 It is important to note, however, that the VH anatomy may not provide a true representation of in vivo human anatomy due to the loss of the natural tone of the muscles. For example, the average length of the human small intestine has been found to be upwards of 3 m shorter in life than in death.69 For this reason, digitizing CT or MR images obtained in vivo has more recently been viewed as the preferred method of obtaining anatomical stomach models for computational simulations of GEA.5, 70 An average model (in terms of stomach shape and volume) of many subject-specific models created in this way may be used in representative simulations, to overcome the problem of variability encountered between individual subjects.
In order to generate descriptive models for use in body-surface investigations such as electrogastrography (EGG) and magnetogastrography (MGG) (discussed in the following section), it is necessary to relate organ-level electrical activity to the body surface. The most common approach for projecting the electrical activity from the stomach to the body surface involves forward computation of current dipole sources. Smout71 presented one of the first models to use the dipole theory to represent GIEA within a cylinder. Others’ models used a single dipole to represent the electrical sources72, 73 followed by a larger band of moving dipoles to provide a more distributed representation of the slow wave.74-77 Each of these models, however, have used a simplified geometric representation of the stomach and torso, ranging from cylinders to cones to ellipses. More recently, the Aliev et al.19 model has been integrated with the bidomain equations, and solved using a structured finite element method to represent gastric electrical activity in an anatomical model of the human stomach.3 The gradient of the transmembrane potential over the stomach was then used to calculate equivalent dipole sources to compute electromagnetic fields external to the torso.78, 79
CLINICAL APPLICATIONS
The stomach and small intestine share important electrophysiology similarities with the heart, including pacemaker activity and electromechanical coupling. Just as cardiac electrical dysfunction underlies many important disorders of cardiac mechanical activity, it is widely assumed that abnormalities in electrical activity may underlie many GI motility disorders. For example, dysrhythmic GEA has been described in relation to common clinical problems such as gastroparesis35 and functional dyspepsia,80 and disorganized electrical activity is thought to play a role in postoperative ileus.81 However, whereas electrical activity is routinely measured and manipulated in cardiology practice with electrocardiograms (ECGs), pharmacology, and electrical stimulation, diagnostic and therapeutic electrophysiology has failed to enter routine gastroenterology practice up to the present time.
Improved methods of defining, diagnosing, and treating pathologies of GIEA are an important focus of ongoing research. Mathematical GI models of integrated cell, tissue, and organ function are increasingly being employed in this research challenge, where their predictive capability is being used to compliment standard biophysically based approaches. The current status of this research is reviewed below.
Serosal and Mucosal Recordings
Gastric electrograms can be made by placing electrodes either against the inner or outer surfaces of the stomach, to provide detailed information regarding slow wave activity of a particular stomach region.36, 82 Serosal recording necessitates an invasive procedure (either laparoscopy or laparotomy), which precludes it as an everyday clinical application. Endoscopic placement of electrodes on the mucosal surface is less invasive, but impractical for prolonged periods due to the discomfort of a nasogastric tube and potential for the electrodes to dislodge. Via serosal recordings in an animal model, Lammers et al.83 have recently described complex abnormal GI electrical events, like those known to occur pathologically in the heart, such as multiple ectopic pacemakers and re-entrant arrhythmias. However, further research is required to define the role such events may play in human disease. New methods that are appropriate for human research are now emerging, such as easily sterilized flexible printed circuit board (PCB) electrodes, that will facilitate future advances in this field.84
Du et al.58 have recently performed simulations of GEA across a virtual section of stomach musculature using a biophysically based computation model. Early results suggest good concordance with direct serosal recordings of GEA (refer to Figure 6). It is hoped such models may become a future platform to simulate GI disease states, for example, by generating predictions regarding ICC cell loss or network disruption on the propagation of slow wave activity.
FIGURE 6.
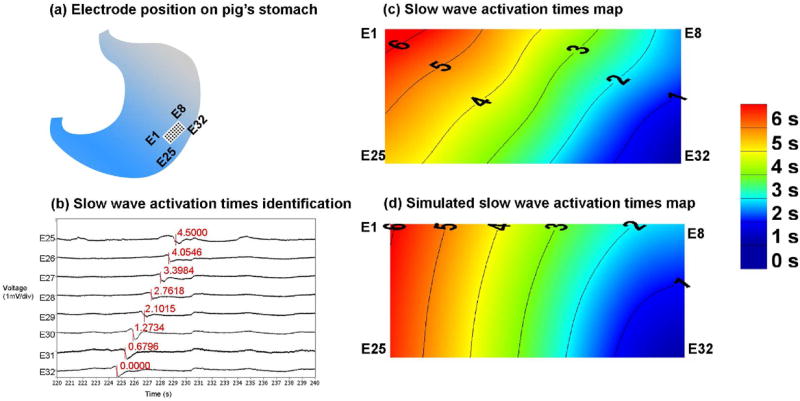
Illustration of gastric serosal recordings using a 32-channel electrode array (E1–E32), which was placed in the orientation on a porcine stomach as shown in (a). Shown in (b) are recorded extracellular traces corresponding to eight electrodes (out of 32) with slow wave activation times marked by the red vertical lines. The locations of the red lines were determined by the most negative deflection during a slow wave event. Also shown is (c) an activation times map corresponding to the signals in (b), the isochrones of activation times indicate that slow waves propagation was in the aboral direction (E32 towards E1). Shown in (d) is a simulated slow wave event with the activation times sampled over the same dimensions as the array of electrodes (a).The activation times and direction of the simulated slow waves (d) are in a reasonable agreement with the experimental recording (c).
Electrogastrography
EGG is a non invasive recording of GEA carried out by placement of cutaneous abdominal electrodes.85 EGG is convenient, and recordings correlate with measurements taken directly from the gastric serosa.86 However, because EGG is a summation of all gastric electrical activity occurring simultaneously, it lacks discriminative ability to demonstrate electrical differences among various stomach regions, and analysis, to date, have largely been limited to frequency dynamics.87 Gastric dysrhythmias such as bradygastria (<2 cpm) and tachygastrias (>5 cpm) have been described by EGG in association with clinical conditions including gastroparesis, functional dyspepsia, gastroesophageal reflux disease (GERD), and unexplained nausea and vomiting.67 However, EGG has not found a place in routine clinical practice because of the poor correlation with symptoms, gastric emptying, and manometry.88 Multichannel EGG has been proposed as a possible method to derive more detailed information, such as slow wave propagation and uncoupling,89 but this awaits more extensive validation.
Anatomically realistic multiscale models of GEA have been integrated within torso models to describe the conductance of EGG signals on to the body surface. Using this approach, it was shown that EGG is unlikely to be able to discriminate between normal and functionally uncoupled GEA in terms of the dominant frequency component caused by a retrograde pacemaker site.6 This finding awaits definitive physiological validation; although one study has suggested that multichannel EGG can discriminate severe uncoupling in a canine model by EGG and computer-assisted processing.90
Magnetogastrography and Magnetoenterogram
Most biological electrical fields have a corresponding magnetic field associated with them. The magnetic fields can be recorded via the use of a Superconducting QUantum Interference Device (SQUID) magnetometer. SQUID magnometers provide an attractive option for characterizing electrical activity in the GI system as the recordings can be made noninvasively and without direct contact of the skin. Electrical fields are a scalar quantity, while magnetic fields are a vector field with an orientation and a magnitude. Although the majority of SQUID sensors only measure one component of the magnetic field (typically the director orthogonal to the body) appropriate SQUID sensors are capable of measuring the three orthogonal magnetic fields at a given point. It has been shown in past studies that there is significant information present in the full vector field that may not be obtained by a single-channel measurement.91, 92 In addition, because of the spatial filtering effects, the spatial resolution of the SQUID magnetometer is considered to be greater than what can be achieved with a surface electrode array.
Figure 7 illustrates the magnetic and electrical field caused by a horizontally aligned dipole within the stomach. The magnetic field is represented by gold arrows located at a plane just above the body, and the electrical potential distribution on the body surface is indicated by the colored field.
FIGURE 7.
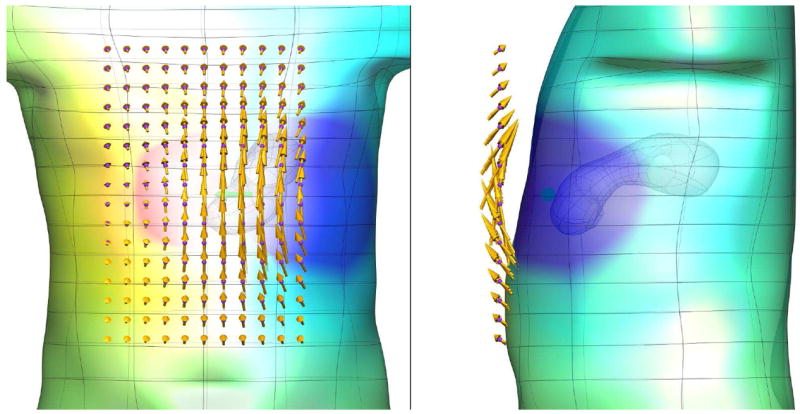
Anterior and sagittal views of simulated electric and magnetic fields due to a current dipole. A horizontal dipole (green arrow) in the stomach produces electric fields on the body surface (represented by the colored field; blue is negative and red is positive potential) and the magnetic field external to the body (represented by the gold arrows), with the length of the arrow indicating the strength of the magnetic field.
Recordings of magnetic fields resulting from GEA are known as magnetogastrograms (MGGs).93, 94 Recordings of magnetic fields resulting from GEA are known as magnetoenterograms (MGGs). Although small intestinal signals are much weaker in nature, they have also been recorded using a SQUID95, 96 and are referred to as magnetogastrograms (MENGs). This intestinal activity has never been reliably recorded using cutaneous electrodes. The difference in signal-to-noise ratio between the EENG and the MENG arises principally on account of the alternating tissue layers with high and low conductivities.97 The surface EENG is affected to a larger degree by this layered conductive structure, as well as the presence of local noise sources such as subdermal muscle activity. Although the use of SQUID technology has a number of advantages it has yet to gain widespread clinical acceptance, and is an active area of ongoing research. There is a need for additional computational simulations and modeling techniques to aid the interpretation of the magnetic field recordings.70
Gastric Electrical Stimulation
Gastric electrical stimulation (GES) has been attempted for over 40 years as a strategy to manipulate gastric electrical activity in order to treat several major diseases. Three main strategies have been employed: gastric pacing (low-frequency GES), high-frequency GES and neural GES (NGES).
Gastric pacing involves delivery of external impulses at frequencies near the intrinsic 3-cpm activity, with the aim of entraining slow wave activity.98 Most pacing studies have attempted to entrain slow waves in the normal (aboral) direction, aiming to promote gastric motility when it is deficient, as in postoperative ileus99 or gastroparesis.100, 101 One noncontrolled human gastroparesis trial to date has demonstrated the therapeutic feasibility of gastric pacing, by demonstrating significantly decreased gastric retention, reduced symptoms, and a decreased need for supplemental jejunostomy feeding in treated patients.101 Some recent studies have also attempted to entrain gastric slow waves in the reverse (retrograde) direction, as a means to restrict motility and reduce gastric emptying, as a treatment for obesity. Trials in obese Zucker rats,102 chronically instrumented dogs,103 and normal human volunteers,104 have demonstrated the potential of this strategy, by successfully showing reductions in solid food intake, without inducing intolerable symptoms.
The second stimulation strategy, high-frequency GES, involves delivery of stimuli far beyond the normal 3-cpm slow wave frequency (~14 Hz range). Commercial devices are available, including the Enterra device, (Medtronic, MI) for gastroparesis, and Transcend (Transneuronix, NJ) for obesity. The therapeutic mechanism of this strategy is presently unknown.105 The Enterra device has shown promise in noncontrolled trials to substantially improve symptoms of nausea and vomiting in severe gastroparetic, but awaits further validation in well controlled trials.106 The Transcend device has achieved modest success in clinical trials (20–30% excess weight loss).107
In the third stimulation strategy, NGES, ultra-high-frequency (>40 Hz range) stimuli are used to induce intramural cholinergic fibers to release acetylcholine, invoking gastric contractions.108 Retrograde NGES has also achieved significant reductions in food intake and weight in a canine pilot study (p < 0.05).109
One of the most pressing research problems in GES is to identify the optimum stimulation protocols to achieve the desired clinically relevant slow wave and motility outcomes. Technically, there are an infinite number of parameters to evaluate, including electrode placements, pulse frequency, pulse width, and pulse amplitude. Researchers are also attempting to find an effective means to reduce the power consumption of implantable pacing, or NGES devices.103 To date, GES research has proved a technically tedious exercise, requiring laborious trial and error experiments on animal models.110 More recently, biophysically based computational models of GIEA, such as those of Du et al.,58 have shown potential to act as hypothesis-testing tools for use in defining effective protocols for improved GI stimulation. This strategy may result in increased research efficiency, by transfer of developmental work to a computational rather than animal models.
CONCLUSION
Clinical applications of gastric electrophysiology remain in their relative infancy in comparison with the diagnostic and therapeutic cardiac applications of cardiac electrophysiology. However, there are a number of research approaches that have significant potential to aid with the diagnosis and treatment of GI disorders. Computational modeling using biophysical methodology which takes into account subcellular, cell, tissue, organ, and body elements has become an important adjunct in advancing our understanding of integrative physiology, and will help to develop predictions for clinical hypotheses testing.
Acknowledgments
This work was funded in part by a grant from the National Institutes of Health (R01 DK64775) and a NZ Society of Gastroenterology Research Fellowship. Peng Du is supported by a University of Auckland Doctoral Scholarship, and Greg O’Grady through a New Zealand Health Research Council Clinical Research Training Fellowship.
References
- 1.Hunter PJ, Borg TK. Integration from proteins to organs: the Physiome Project. Nat Rev Mol Cell Biol. 2003;4:237–243. doi: 10.1038/nrm1054. [DOI] [PubMed] [Google Scholar]
- 2.Faville RA, Pullan AJ, Sanders KM, Smith NP. A biophysically based mathematical model of unitary potential activity in interstitial cells of Cajal. Biophys J. 2008;95:88–104. doi: 10.1529/biophysj.107.122507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pullan AJ, Cheng LK, Yassi R, Buist ML. Modelling gastrointestinal bioelectric activity. Prog Biophys Mol Biol. 2004;85:523–550. doi: 10.1016/j.pbiomolbio.2004.02.003. [DOI] [PubMed] [Google Scholar]
- 4.Lin AS, Buist ML, Cheng LK, Smith NP, Pullan AJ. Computational simulations of the human magneto- and electroenterogram. Ann Biomed Eng. 2006;34(8):1322–1331. doi: 10.1007/s10439-006-9142-4. [DOI] [PubMed] [Google Scholar]
- 5.Cheng LK, Komuro R, Austin TM, Buist ML, Pullan AJ. Anatomically realistic multiscale models of normal and abnormal gastrointestinal electrical activity. World J Gastroenterol. 2007;13:1378–1383. doi: 10.3748/wjg.v13.i9.1378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Buist ML, Cheng LK, Sanders KM, Pullan AJ. Multiscale modelling of human gastric electric activity: can the electrogastrogram detect functional electrical uncoupling? Exp Physiol. 2006;91:383–390. doi: 10.1113/expphysiol.2005.031021. [DOI] [PubMed] [Google Scholar]
- 7.Sanders KM, Koh SD, Ward SM. Interstitial cells of cajal as pacemakers in the gastrointestinal tract. Annu Rev Physiol. 2006;68:307–343. doi: 10.1146/annurev.physiol.68.040504.094718. [DOI] [PubMed] [Google Scholar]
- 8.Kelly KA, La Force RC. Role of the gastric pacesetter defined by electrical pacing. Can J Physiol Pharmacol. 1972;50:1017–1019. doi: 10.1139/y72-147. [DOI] [PubMed] [Google Scholar]
- 9.Sanders KM. Regulation of smooth muscle excitation and contraction. Neurogastroenterol Motil. 2008;20:39–53. doi: 10.1111/j.1365-2982.2008.01108.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Huizinga JD, Thuneberg L, Kluppel M, Malysz J, Mikkelsen HB, et al. W/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature. 1995;373:347–349. doi: 10.1038/373347a0. [DOI] [PubMed] [Google Scholar]
- 11.Hirst GD, Edwards FR. Generation of slow waves in the antral region of guinea-pig stomach–a stochastic process. J Physiol. 2001;535:165–180. doi: 10.1111/j.1469-7793.2001.00165.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cousins HM, Edwards FR, Hickey H, Hill CE, Hirst GD. Electrical coupling between the myenteric interstitial cells of Cajal and adjacent muscle layers in the guinea-pig gastric antrum. J Physiol. 2003;550:829–844. doi: 10.1113/jphysiol.2003.042176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ordog T, Ward SM, Sanders KM. Interstitial cells of Cajal generate electrical slow waves in the murine stomach. J Physiol. 1999;518:257–269. doi: 10.1111/j.1469-7793.1999.0257r.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ver Donck L, Lammers WJ, Moreaux B, Smets D, Voeten J, Vekemans J, Schuurkes JA, Coulie B. Mapping slow waves and spikes in chronically instrumented conscious dogs: implantation techniques and recordings. Med Biol Eng Comput. 2006;44:170–178. doi: 10.1007/s11517-005-0018-9. [DOI] [PubMed] [Google Scholar]
- 15.Nelsen TS, Becker JC. Simulation of the electrical and mechanical gradient of the small intestine. Am J Physiol. 1968;214:749–757. doi: 10.1152/ajplegacy.1968.214.4.749. [DOI] [PubMed] [Google Scholar]
- 16.Sarna SK, Daniel EE, Kingma YJ. Simulation of slow-wave electrical activity of small intestine. Am J Physiol. 1971;221:166–175. doi: 10.1152/ajplegacy.1971.221.1.166. [DOI] [PubMed] [Google Scholar]
- 17.Sarna SK, Daniel EE, Kingma YJ. Effects of partial cuts on gastric electrical control activity and its computer model. Am J Physiol. 1972;223:332–340. doi: 10.1152/ajplegacy.1972.223.2.332. [DOI] [PubMed] [Google Scholar]
- 18.Daniel EE, Bardakjian BL, Huizinga JD, Diamant NE. Relaxation oscillator and core conductor models are needed for understanding of GI electrical activities. Am J Physiol. 1994;266:G339–G349. doi: 10.1152/ajpgi.1994.266.3.G339. [DOI] [PubMed] [Google Scholar]
- 19.Aliev RR, Richards W, Wikswo JP. A simple nonlinear model of electrical activity in the intestine. J Theor Biol. 2000;204:21–28. doi: 10.1006/jtbi.2000.1069. [DOI] [PubMed] [Google Scholar]
- 20.Publicover NG, Sanders KM. Are relaxation oscillators an appropriate model of gastrointestinal electrical activity? Am J Physiol. 1989;256:265–274. doi: 10.1152/ajpgi.1989.256.2.G265. [DOI] [PubMed] [Google Scholar]
- 21.Corrias A, Buist ML. Quantitative cellular description of gastric slow wave activity. Am J Physiol Gastrointest Liver Physiol. 2008;294:G989–G995. doi: 10.1152/ajpgi.00528.2007. [DOI] [PubMed] [Google Scholar]
- 22.Corrias A, Buist ML. A quantitative model of gastric smooth muscle cellular activation. Ann Biomed Eng. 2007;35:1595–1607. doi: 10.1007/s10439-007-9324-8. [DOI] [PubMed] [Google Scholar]
- 23.Hodgkin AL, Huxley AF. A quantitative description of membrane current and its application to conduction and excitation in nerve. J Physiol. 1952;117:500–544. doi: 10.1113/jphysiol.1952.sp004764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Luo CH, Rudy Y. A model of the ventricular cardiac action potential. Depolarization repolarization and their interaction. Circ Res. 1991;68:1501–1526. doi: 10.1161/01.res.68.6.1501. [DOI] [PubMed] [Google Scholar]
- 25.Luo CH, Rudy Y. A dynamic model of the cardiac ventricular action potential. I. Simulations of ionic currents and concentration changes. Circ Res. 1994;74:1071–1096. doi: 10.1161/01.res.74.6.1071. [DOI] [PubMed] [Google Scholar]
- 26.Edwards FR, Hirst GD. Mathematical description of regenerative potentials recorded from circular smooth muscle of guinea pig antrum. Am J Physiol Gastrointest Liver Physiol. 2003;285:G661–G670. doi: 10.1152/ajpgi.00145.2003. [DOI] [PubMed] [Google Scholar]
- 27.Youm JB, Kim N, Han J, Kim E, Joo H, et al. A mathematical model of pacemaker activity recorded from mouse small intestine. Philos Trans A Math Phys Eng Sci. 2006;364:1135–1154. doi: 10.1098/rsta.2006.1759. [DOI] [PubMed] [Google Scholar]
- 28.Fall CP, Keizer JE. Mitochondrial modulation of intracellular Ca(2 +) signaling. J Theor Biol. 2001;210:151–165. doi: 10.1006/jtbi.2000.2292. [DOI] [PubMed] [Google Scholar]
- 29.Cuellar AA, Lloyd CM, Nielsen PF, Bullivant DP, Nickerson DP, Hunter PJ. An overview of CellML 1.1, a biological model description language simulation. Simulation. 2003;79:740–747. [Google Scholar]
- 30.Johnson LR, editor. Physiology of the Gastrointestinal Tract. New York: Raven Press; 2006. pp. 533–576. [Google Scholar]
- 31.Hirst GD. An additional role for ICC in the control of gastrointestinal motility? J Physiol. 2001;537(Pt 1):1. doi: 10.1111/j.1469-7793.2001.0001k.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Farrugia G. Interstitial cells of Cajal in health and disease. Neurogastroenterol Motil. 2008;20:54–63. doi: 10.1111/j.1365-2982.2008.01109.x. [DOI] [PubMed] [Google Scholar]
- 33.Hirst GD, Dickens EJ, Edwards FR. Pacemaker shift in the gastric antrum of guinea-pigs produced by excitatory vagal stimulation involves intramuscular interstitial cells. J Physiol. 2002;541:917–928. doi: 10.1113/jphysiol.2002.018614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bauer AJ, Publicover NG, Sanders KM. Origin and spread of slow waves in canine gastric antral circular muscle. Am J Physiol. 1985;249(6Pt1):G800–G806. doi: 10.1152/ajpgi.1985.249.6.G800. [DOI] [PubMed] [Google Scholar]
- 35.Chen JD, Lin Z, Pan J, McCallum RW. Abnormal gastric myoelectrical activity and delayed gastric emptying in patients with symptoms suggestive of gastroparesis. Dig Dis Sci. 1996;41:1538–1545. doi: 10.1007/BF02087897. [DOI] [PubMed] [Google Scholar]
- 36.Lammers WJ, Stephen B, Arafat K, Manefield GW. High resolution electrical mapping in the gastrointestinal system: initial results. Neurogastroenterol Motil. 1996;8:207–216. doi: 10.1111/j.1365-2982.1996.tb00259.x. [DOI] [PubMed] [Google Scholar]
- 37.Schmitt OH. Information Processing in the Nervous System. New York, NY: Springer-Verlag; 1969. Biological information processing using the concept of interpenetrating domains. [Google Scholar]
- 38.Muler AL, Markin VS. Electrical properties of anisotropic nerve-muscle syncytia - I. Distribution of the electrotonic potential. Biofizika. 1977;22:307–312. [PubMed] [Google Scholar]
- 39.Miller WT, Geselowitz DB. Simulation studies of the electrocardiogram. I. The normal heart. Circ Res. 1978;43:301–315. doi: 10.1161/01.res.43.2.301. [DOI] [PubMed] [Google Scholar]
- 40.Tung L. PhD Thesis. M.I.T; Boston, MA: 1978. A Bidomain Model for Describing Ischemia Myocardial DC Potentials. [Google Scholar]
- 41.Geselowitz DB, Miller WT., III A bidomain model for anisotropic cardiac muscle. Ann Biomed Eng. 1983;11:191–206. doi: 10.1007/BF02363286. [DOI] [PubMed] [Google Scholar]
- 42.Plonsey R, Barr RC. Current flow patterns in two-dimensional anisotropic bisyncytia with normal and extreme conductivities. Biophys J. 1984;45:557–571. doi: 10.1016/S0006-3495(84)84193-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Barr RC, Plonsey R. Propagation of excitation in idealized anisotropic two-dimensional tissue. Biophys J. 1984;45:1191–1202. doi: 10.1016/S0006-3495(84)84268-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Roth BJ, Wikswo JP. A bidomain model for the extracellular potential and magnetic field of cardiac tissue. IEEE Trans Biomed Eng. 1986;33:467–469. doi: 10.1109/TBME.1986.325804. [DOI] [PubMed] [Google Scholar]
- 45.Henriquez CS. Simulating the electrical behavior of cardiac tissue using the bidomain model. Crit Rev Biomed Eng. 1993;21:1–77. [PubMed] [Google Scholar]
- 46.Pullan AJ, Buist ML, Cheng LK. Mathematically Modelling the Electrical Activity of the Heart: From Cell to Body Surface and Back Again. Singapore: World Scientific Publishing; 2007. [Google Scholar]
- 47.Soybel DI. Anatomy and physiology of the stomach. Surg Clin North Am. 2005;85:875–894. doi: 10.1016/j.suc.2005.05.009. [DOI] [PubMed] [Google Scholar]
- 48.Kelly KA, Code CF. Canine gastric pacemaker. Am J Physiol. 1971;220:112–118. doi: 10.1152/ajplegacy.1971.220.1.112. [DOI] [PubMed] [Google Scholar]
- 49.Kwong NK, Brown BH, Whittaker GE, Duthie HL. Electrical activity of the gastric antrum in man. Br J Surg. 1970;57:913–916. doi: 10.1002/bjs.1800571211. [DOI] [PubMed] [Google Scholar]
- 50.Kelly KA, Code CF, Elveback LR. Patterns of canine gastric electrical activity. Am J Physiol. 1969;217:461–470. doi: 10.1152/ajplegacy.1969.217.2.461. [DOI] [PubMed] [Google Scholar]
- 51.Lammers WJ, Stephen B, Adeghate E, Ponery S, Pozzan O. The slow wave does not propagate across the gastroduodenal junction in the isolated feline preparation. Neurogastroenterol Motil. 1998;10:339–349. doi: 10.1046/j.1365-2982.1998.00113.x. [DOI] [PubMed] [Google Scholar]
- 52.Wang XY, Lammers WJ, Bercik P, Huizinga JD. Lack of pyloric interstitial cells of Cajal explains distinct peristaltic motor patterns in stomach and small intestine. Am J Physiol Gastrointest Liver Physiol. 2005;289:G539–G549. doi: 10.1152/ajpgi.00046.2005. [DOI] [PubMed] [Google Scholar]
- 53.Lammers WJ, Slack JR, Stephen B, Pozzan O. The spatial behaviour of spike patches in the feline gastroduodenal junction in vitro. Neurogastroenterol Motil. 2000;12:467–473. doi: 10.1046/j.1365-2982.2000.00223.x. [DOI] [PubMed] [Google Scholar]
- 54.Guyton AC, Hall JE. Textbook of Medical Physiology. 10. Philadelphia, PA: W.B. Saunders Company; 2000. Chapter 63: Propulsion and mixing of food in the alimentary tract. [Google Scholar]
- 55.Christensen J, Schedl HP, Clifton JA. The small intestinal basic electrical rhythm (slow wave) frequency gradient in normal men and patients with variety of diseases. Gastroenterology. 1966;50:309–315. [PubMed] [Google Scholar]
- 56.Fleckenstein P. Migrating electrical spike activity in the fasting human small intestine. Am J Dig Dis. 1978;23:769–775. doi: 10.1007/BF01079784. [DOI] [PubMed] [Google Scholar]
- 57.Lammers WJ, Stephen B. Origin and propagation of individual slow waves along the intact feline small intestine. Exp Physiol. 2008;93:334–346. doi: 10.1113/expphysiol.2007.039180. [DOI] [PubMed] [Google Scholar]
- 58.Du P, O’Grady G, Windsor JA, Cheng LK, Pullan AJ. A tissue framework for simulating the effects of gastric electrical stimulation and in vivo validation. IEEE Trans Biomed Eng. 2009;56:2755–61. doi: 10.1109/TBME.2009.2027690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Melville J, Macagno E, Christensen J. Longitudinal contractions in the duodenum: their fluid-mechanical function. Am J Physiol. 1975;228:1887–1892. doi: 10.1152/ajplegacy.1975.228.6.1887. [DOI] [PubMed] [Google Scholar]
- 60.Pal A, Indireshkumar K, Schwizer W, Abrahamsson B, Fried M, et al. Gastric flow and mixing studied using computer simulation. Proc Biol Sci. 2004;271:2587–2594. doi: 10.1098/rspb.2004.2886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Pal A, Brasseur JG, Abrahamsson B. A stomach road or “Magenstrasse” for gastric emptying. J Biomech. 2007;40:1202–1210. doi: 10.1016/j.jbiomech.2006.06.006. [DOI] [PubMed] [Google Scholar]
- 62.Kita Y. Elastic-model driven analysis of several views of a deformable cylindrical object. IEEE Trans Pattern Anal Mach Intell. 1996;18:1150–1162. [Google Scholar]
- 63.Liao D, Zhao J, Gregersen H. Regional surface geometry of the rat stomach based on three-dimensional curvature analysis. Phys Med Biol. 2005;50:231–246. doi: 10.1088/0031-9155/50/2/004. [DOI] [PubMed] [Google Scholar]
- 64.Spitzer VM, Whitlock DG. The visible human dataset: the anatomical platform for human simulation. Anat Rec. 1998;253:49–57. doi: 10.1002/(SICI)1097-0185(199804)253:2<49::AID-AR8>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 65.Bradley CP, Pullan AJ, Hunter PJ. Geometric modeling of the human torso using cubic Hermite elements. Ann Biomed Eng. 1997;25:96–111. doi: 10.1007/BF02738542. [DOI] [PubMed] [Google Scholar]
- 66.Yassi R, Cheng LK, Al-Ali S, Smith NP, Pullan AJ, et al. An anatomically based mathematical model of the gastroesophageal junction. Conf Proc IEEE Eng Med Biol Soc. 2004;1:635–638. doi: 10.1109/IEMBS.2004.1403238. [DOI] [PubMed] [Google Scholar]
- 67.Lin Z, Chen JDZ. Electrogastrography (EGG) In: Akay M, editor. Wiley Encyclopedia of Biomedical Engineering. New York, NY: John Wiley & Sons; 2006. [Google Scholar]
- 68.Noakes KF, Bissett IP, Pullan AJ, Cheng LK. Anatomically realistic three-dimensional meshes of the pelvic floor and anal canal for finite element analysis. Ann Biomed Eng. 2008;36:1060–1071. doi: 10.1007/s10439-008-9471-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Fanucci A, Cerro P, Fraracci L, Ietto F. Small bowel length measured by radiography. Gastrointest Radiol. 1984;9:349–351. doi: 10.1007/BF01887866. [DOI] [PubMed] [Google Scholar]
- 70.Komuro R, Cheng LK, Pullan AJ. Comparison and analysis of inter-subject variability of simulated magnetic activity generated from gastric electrical activity. Ann Biomed Eng. 2008;36:1049–1059. doi: 10.1007/s10439-008-9480-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Smout AJ, van der Schee EJ, Grashuis JL. What is measured in electrogastrography? Dig Dis Sci. 1980;25:179–187. doi: 10.1007/BF01308136. [DOI] [PubMed] [Google Scholar]
- 72.Kothapalli B. Electrogastrogram simulation using a three-dimensional model. Med Biol Eng Comput. 1993;31:482–486. doi: 10.1007/BF02441983. [DOI] [PubMed] [Google Scholar]
- 73.Liang J, Chen JD. What can be measured from surface electrogastrophy? Computer simulations. 1997;42:1331–1343. doi: 10.1023/a:1018869300296. [DOI] [PubMed] [Google Scholar]
- 74.Mirizzi N, Stella R, Scafoglieri U. Model to simulate the gastric electrical control and response activity on the stomach wall and on the abdominal surface. Med Biol Eng Comput. 1986;24:157–163. doi: 10.1007/BF02443929. [DOI] [PubMed] [Google Scholar]
- 75.Mintchev MP, Bowes KL. Conoidal dipole model of electrical field produced by the human stomach. Med Biol Eng Comput. 1995;33:179–184. doi: 10.1007/BF02523038. [DOI] [PubMed] [Google Scholar]
- 76.Bradshaw LA, Myers A, Wikswo JP, Richards WO. A spatio-temporal dipole simulation of gastrointestinal magnetic fields. IEEE Trans Biomed Eng. 2003;50:836–847. doi: 10.1109/TBME.2003.813549. [DOI] [PubMed] [Google Scholar]
- 77.Irimia A, Bradshaw LA. Ellipsoidal electrogastrographic forward modelling. Phys Med Biol. 2005;50:4429–4444. doi: 10.1088/0031-9155/50/18/012. [DOI] [PubMed] [Google Scholar]
- 78.Buist ML, Cheng LK, Yassi R, Bradshaw LA, Richards WO, Pullan AJ. An anatomical model of the gastric system for producing bioelectric and biomagnetic fields. Physiol Meas. 2004;25:849–861. doi: 10.1088/0967-3334/25/4/006. [DOI] [PubMed] [Google Scholar]
- 79.Austin TM, Li L, Pullan AJ, Cheng LK. Effects of gastrointestinal tissue structure on computed dipole vectors. Biomed Eng Online. 2007;6:39. doi: 10.1186/1475-925X-6-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lin X, Chen JZ. Abnormal gastric slow waves in patients with functional dyspepsia assessed by multichannel electrogastrography. Am J Physiol Gastrointest Liver Physiol. 2001;280:G1370–G1375. doi: 10.1152/ajpgi.2001.280.6.G1370. [DOI] [PubMed] [Google Scholar]
- 81.Behm B, Stollman N. Postoperative ileus: etiologies and interventions. Clin Gastroenterol Hepatol. 2003;1:71–80. doi: 10.1053/cgh.2003.50012. [DOI] [PubMed] [Google Scholar]
- 82.Coleski R, Hasler W. Regional gastric slow wave propagation and coupling measured by endoscopy-directed multichannel gastric mucosal recording in healthy humans: effects of acute hyperglycaemia. Neurogastroenterol Motil. 2006;18:740. [Google Scholar]
- 83.Lammers WJ, Ver Donck L, Stephen B, Smets D, Schuurkes JA. Focal activities and re-entrant propagations as mechanisms of gastric tachyrrhythmias. Gastroenterology. 2008;135:1601–1611. doi: 10.1053/j.gastro.2008.07.020. [DOI] [PubMed] [Google Scholar]
- 84.Du P, O’Grady G, Egbuji JU, Lammers WJ, Budgett D, Nielsen P, Windsor JA, Pullan AJ, Cheng LK. High resolution mapping of in-vivo gastrointestinal slow wave activity using flexible printed circuit board electrodes: methodology and validation. Ann Biomed Eng. 2009;37:839–846. doi: 10.1007/s10439-009-9654-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Parkman HP, Hasler WL, Barnett JL, Eaker EY American Motility Society Clinical GI Motility Testing Task Force. Electrogastrography: a document prepared by the gastric section of the American Motility Society Clinical GI Motility Testing Task Force. Neurogastroenterol Motil. 2003;15(2):89–102. doi: 10.1046/j.1365-2982.2003.00396.x. [DOI] [PubMed] [Google Scholar]
- 86.Chen JD, Schirmer BD, McCallum RW. Serosal and cutaneous recordings of gastric myoelectrical activity in patients with gastroparesis. Am J Physiol. 1994;266:G90–G98. doi: 10.1152/ajpgi.1994.266.1.G90. [DOI] [PubMed] [Google Scholar]
- 87.Verhagen MAM, Van Schelven LJ, Samsom M, Smout AJPM. Pitfalls in the analysis of electrogastrographic recordings. Gastroenterology. 1999;117:453–460. doi: 10.1053/gast.1999.0029900453. [DOI] [PubMed] [Google Scholar]
- 88.Abid S, Lindberg G. Electrogastrography: poor correlation with antro-duodenal manometry and doubtful clinical usefulness in adults. World J Gastroenterol. 2007;13:5101–5107. doi: 10.3748/wjg.v13.i38.5101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Simonian HP, Panganamamula K, Parkman HP, Xu X, Chen JZ, Lindberg G, Xu H, Shao C, Ke MY, Lykke M, Hansen P, Barner B, Buhl H. Multichannel electrogastrography (EGG) in normal subjects: a multicenter study. Dig Dis Sci. 2004;49:594–601. doi: 10.1023/b:ddas.0000026304.83214.50. [DOI] [PubMed] [Google Scholar]
- 90.Mintchev MP, Otto SJ, Bowes KL. Electrogastrography can recognise gastric electrical uncoupling in dogs. Gastroenterology. 1997;112:2006–2011. doi: 10.1053/gast.1997.v112.pm9178693. [DOI] [PubMed] [Google Scholar]
- 91.Bradshaw LA, Ladipo JK, Staton DJ, Wikswo JP, Jr, Richards WO. The human vector magnetogastrogram and magnetoenterogram. IEEE Trans Biomed Eng. 1999;46:959–970. doi: 10.1109/10.775406. [DOI] [PubMed] [Google Scholar]
- 92.Bradshaw LA, Myers A, Richards WO, Drake W, Wikswo JP. Vector projection of biomagnetic fields. Med Biol Eng Comput. 2005;43:85–93. doi: 10.1007/BF02345127. [DOI] [PubMed] [Google Scholar]
- 93.Comani S, Basile M, Casciardi S, Del Gratta C, Di Luzio S, et al. Biomagnetism: Clinical Aspects. Proceedings of the 8th International Conference on Biomagnetism; Munster. 1992. pp. 639–642. [Google Scholar]
- 94.Bradshaw LA, Irimia A, Sims JA, Gallucci MR, Palmer RL, Richards WO. Biomagnetic characterization of spatiotemporal parameters of the gastric slow wave. Neurogastroenterol Motil. 2006;18:619–631. doi: 10.1111/j.1365-2982.2006.00794.x. [DOI] [PubMed] [Google Scholar]
- 95.Richards WO, Bradshaw LA, Staton DJ, Garrard CL, Liu F, et al. Magnetoenterography (MENG): noninvasive measurement of bioelectric activity in human small intestine. Dig Dis Sci. 1996;41:2293–2301. doi: 10.1007/BF02100117. [DOI] [PubMed] [Google Scholar]
- 96.Bradshaw LA, Allos SH, Wikswo JP, Jr, Richards WO. Correlation and comparison of magnetic and electric detection of small intestinal electrical activity. Am J Physiol. 1997;272(5Pt1):G1159–G1167. doi: 10.1152/ajpgi.1997.272.5.G1159. [DOI] [PubMed] [Google Scholar]
- 97.Bradshaw LA, Richards WO, Wikswo JP., Jr Volume conductor effects on the spatial resolution of magnetic fields and electric potentials from gastrointestinal electrical activity. Med Biol Eng Comput. 2001;39:35–43. doi: 10.1007/BF02345264. [DOI] [PubMed] [Google Scholar]
- 98.Bortolotti M. The “electrical way” to cure gastroparesis. Am J Gastroenterol. 2002;97:1874–1883. doi: 10.1111/j.1572-0241.2002.05898.x. [DOI] [PubMed] [Google Scholar]
- 99.Bilgutay AM, Wingrove R, Griffen WO, Bonnabeau RC, Lillehei CW. Gastro-intestinal pacing: a new concept in the treatment of ileus. Ann Surg. 1963;158:338–348. doi: 10.1097/00000658-196315830-00003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Bellahsene BE, Lind CD, Schirmer BD, Updike OL, McCallum RW. Acceleration of gastric emptying with electrical stimulation in a canine model of gastroparesis. Am J Physiol. 1992;262:G826–G834. doi: 10.1152/ajpgi.1992.262.5.G826. [DOI] [PubMed] [Google Scholar]
- 101.McCallum RW, Chen JD, Lin Z, Schirmer BD, Williams RD, et al. Gastric pacing improves emptying and symptoms in patients with gastroparesis. Gastroenterology. 1998;114:456–461. doi: 10.1016/s0016-5085(98)70528-1. [DOI] [PubMed] [Google Scholar]
- 102.Yin J, Chen JD. Retrograde gastric electrical stimulation reduces food intake and weight in obese rats. Obes Res. 2005;13:1580–1587. doi: 10.1038/oby.2005.194. [DOI] [PubMed] [Google Scholar]
- 103.Zhang J, Xu X, Chen JD. Chronic tachygastrial electrical stimulation reduces food intake in dogs. Obesity. 2007;15:330–339. doi: 10.1038/oby.2007.557. [DOI] [PubMed] [Google Scholar]
- 104.Yao S, Ke M, Wang Z, Xu D, Zhang Y, et al. Retrograde gastric pacing reduces food intake and delays gastric emptying in humans: a potential therapy for obesity? Dig Dis Sci. 2005;50:1569–1575. doi: 10.1007/s10620-005-2899-8. [DOI] [PubMed] [Google Scholar]
- 105.Zhang J, Chen JD. Systematic review: applications and future of gastric electrical stimulation. Aliment Pharmacol Ther. 2006;24:991–1002. doi: 10.1111/j.1365-2036.2006.03087.x. [DOI] [PubMed] [Google Scholar]
- 106.O’Grady G, Egbuji J, Du P, Cheng LK, Pullan AJ, Windsor JA. High-frequency gastric electrical stimulation for the treatment of gastroparesis: a meta-analysis. World Journal of Surgery. doi: 10.1007/s00268-009-0096-1. Submitted. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Cigaina V. Long-term follow-up of gastric stimulation for obesity: the Mestre 8-year experience. Obes Surg. 2004;14(Suppl 1):S14–S22. doi: 10.1007/BF03342133. [DOI] [PubMed] [Google Scholar]
- 108.Mintchev MP, Sanmiguel CP, Amaris M, Bowes KL. Microprocessor-controlled movement of solid gastric content using sequential neural electrical stimulation. Gastroenterology. 2000;118:258–263. doi: 10.1016/s0016-5085(00)70207-1. [DOI] [PubMed] [Google Scholar]
- 109.Aelen P, Neshev E, Cholette M, Crisanti K, Mitchell P, Debru E, Church N, Mintchev MP. Manipulation of food intake and weight dynamics using retrograde neural gastric electrical stimulation in a chronic canine model. Neurogastroenterol Motil. 2008;20:358–368. doi: 10.1111/j.1365-2982.2007.01027.x. [DOI] [PubMed] [Google Scholar]
- 110.Familoni BO, Abell TL, Nemoto D, Voeller G, Johnson B. Efficacy of electrical stimulation at frequencies higher than basal rate in canine stomach. Dig Dis Sci. 1997;42:892–897. doi: 10.1023/a:1018804128695. [DOI] [PubMed] [Google Scholar]