

Abstract
Objectives
The epidemiology of invasive Haemophilus influenzae (Hi) has changed since the introduction of the Hi type b (Hib) vaccine. The aim of this study was to analyze the clinical and molecular epidemiology of Hi invasive disease in adults.
Methods
Clinical data of the 82 patients with Hi invasive infections were analyzed. Antimicrobial susceptibility, serotyping, and genotyping were studied (2008–2013).
Results
Men accounted for 63.4% of patients (whose mean age was 64.3 years). The most frequent comorbidities were immunosuppressive therapy (34.1%), malignancy (31.7%), diabetes, and COPD (both 22%). The 30-day mortality rate was 20.7%. The majority of the strains (84.3%) were nontypeable (NTHi) and serotype f was the most prevalent serotype in the capsulated strains. The highest antimicrobial resistance was for cotrimoxazole (27.1%) and ampicillin (14.3%). Twenty-three isolates (32.9%) had amino acid changes in the PBP3 involved in resistance. Capsulated strains were clonal and belonged to clonal complexes 6 (serotype b), 124 (serotype f), and 18 (serotype e), whereas NTHi were genetically diverse.
Conclusions
Invasive Hi disease occurred mainly in elderly and those with underlying conditions, and it was associated with a high mortality rate. NTHi were the most common cause of invasive disease and showed high genetic diversity.
Introduction
Haemophilus influenzae is a human-restricted pathogen that forms part of the normal nasopharyngeal microbiota. The presence or absence of a polysaccharide capsule divides this bacterium into two different groups [1], [2]. The variability of capsular polysaccharide means that encapsulated H. influenzae strains are classified into six serotypes, labeled a–f. Before the introduction of the conjugate vaccine against H. influenzae type b (Hib), Hib was the major serotype responsible for invasive infections in infants and young children, with meningoencephalitis as the most common clinical manifestation [1], [3], [4]. With the prevention provided by vaccination, colonization rates and invasive infections in children have been considerably reduced [1], [3]–[6]. Non-capsulated strains, also known as nontypeable H. influenzae (NTHi), frequently cause respiratory infections such as otitis media in children and exacerbations of chronic respiratory diseases and community-acquired pneumonia in fragile adult populations [1], [2], [7]–[9]. Notably, since introduction of the Hib vaccine a strain replacement has been observed in invasive infections and nontypeable strains have become predominant among cases of invasive H. influenzae disease in adults [10], [11].
The main aims of this study were to analyze the demographic and clinical characteristics of adult patients with invasive H. influenzae infections and to determine the antimicrobial resistance and molecular epidemiology of these invasive strains.
Materials and Methods
Ethical Statement
This study has been revised and approved for its publication by the Clinical Research Ethics Committee of Bellvitge University Hospital (PR223/14). Written informed consent was considered not necessary for the study, as it was a retrospective analysis of our usual everyday work. The data of the patients were anonymized for the purposes of this analysis. The confidential information of the patients was protected according to national normative.
Study design and clinical data
A six-year laboratory-based study (2008–2013) was conducted at the Bellvitge University Hospital, a tertiary care center for adult patients located in the south of Barcelona (Spain), the aim being to analyze the epidemiology of invasive H. influenzae. Invasive H. influenzae were defined as the isolation of Hi from blood, cerebrospinal fluid (CSF), or pleural fluid with clinical symptoms in the patient. The denominator used to estimate incidence was the number of persons by age group per year recorded in the public database hosted on the website of the Official Statistics Office of Catalonia (http://www.idescat.cat). Clinical and demographical data were retrieved from a prospective protocol of bacteremia cases recorded at our institution; in those patients without bacteremia their clinical records were reviewed.
Bacterial Strains
Invasive H. influenzae strains collected from sterile sites in our laboratory were stored at −80°C. Isolates were identified by mass spectrometry using a MALDI-TOF Biotyper version 3.0 (Bruker). Differentiation between H. influenzae and H. haemolyticus was performed by the detection of fucK, iga, and lgtC genes, using a previously described methodology [12]. Isolates with a positive detection for all three genes were considered H. influenzae.
Capsule Typing and Antimicrobial Susceptibility Testing
Capsular serotype was determined by PCR using primers and conditions described elsewhere [13]. Antimicrobial susceptibility was tested in all 82 isolates by disk diffusion as a part of the normal laboratory routine, following CLSI recommendations. In the 70 available isolates, minimal inhibitory concentration (MIC) was tested by the microdilution method using commercial panels (STRHAE2; Sensititre, West Sussex, England) and following Clinical Laboratory Standards Institute (CLSI) recommendations [14], [15]. β-lactamase activity was screened using the chromogenic cephalosporin method (nitrocefin disks, BD, Madrid, Spain). Identification of β-lactamase type was performed by PCR on all the positive β-lactamase isolates using previously described primers and conditions [16].
PPB3 Sequencing and Genotype Definition for Ampicillin Resistance
An internal region of the ftsI gene (796-1741 pb) was amplified by PCR and sequenced as previously described [17]. In accordance with previous descriptions [18], [19], H. influenzae were classified into four ampicillin-resistant genotypes: β-lactamase negative ampicillin susceptible (gBLNAS), strains without a detectable resistance mechanism; β-lactamase negative ampicillin resistant (gBLNAR), strains which presented mutations in the ftsI gene; β-lactamase positive ampicillin resistant (gBLPAR), strains producing β-lactamase; and β-lactamase positive amoxicillin/clavulanic acid resistant (gBLPACR), strains which presented both resistance mechanisms (β-lactamase production and mutations in the ftsI gene).
Molecular Typing
Genomic DNA was digested with SmaI and the fragments were separated by pulsed-field gel electrophoresis (PFGE), as reported previously [9]. PFGE band patterns were analyzed using the Fingerprinting II Software 3.0 (BioRad). The similarity of the PFGE banding patterns was estimated with the Dice coefficient, setting the optimization and tolerance at 1%. Isolates with ≥80% relatedness were considered highly genetically related [20]. Multilocus sequence typing (MLST) was performed by DNA sequencing of internal fragments of seven housekeeping genes (adk, atpG, frdB, fuck, mdh, pgi, and recA), as previously described [21]. Allele number and sequence types (ST) were assigned using the H. influenzae MLST website (http://haemophilus.mlst.net).
Statistical Analyses
Statistical analyses were performed using GraphPad Prism version 4, using Chi-square or Fisher’s exact tests, when appropriate, with P<.05 being considered significant.
Results
Clinical characteristics
During the period 2008 to 2013 a total of 3433 H. influenzae were isolated from adult patients in our hospital. Of these, 82 isolates (2.4%) caused invasive H. influenzae infection in 82 patients. The overall incidence for Hi invasive disease among adults in our area during the study period was 2.12 episodes per 100,000 population. By age group the incidence of invasive disease was higher among those aged 65 or older than among people ≤64 years (6.8/100,000 vs. 1.1/100,000; p<0.01). We observed no significant change in the incidence of invasive disease over the study period, neither overall nor by age group or serotype.
Table 1 shows the demographics, clinical characteristics, and underlying conditions for the 82 patients with invasive H. influenzae infection. Fifty-two (63.1%) cases occurred in men and the mean age of patients was 64.3 years. Most cases were community-acquired, and pneumonia was the most frequent type of infection (59.8%) The most common comorbidities were immunosuppressive conditions, malignancies, diabetes, chronic obstructive pulmonary disease (COPD), and heart disease (Table 1). In general, older patients had higher rates of underlying conditions than did those ≤64 years, especially for COPD (34.0% vs. 5.7%, p = 0.002) and heart disease (27.6% vs. 5.7%, p = 0.018). The 30-day mortality was 20.7% (n = 17), with no differences between younger and older adults (p = 0.58).
Table 1. Demographic data, clinical characteristics, and underlying conditions of 82 patients with an invasive H. influenzae episode during the period 2008–2013.
| Patients (n = 82) | |
| Characteristics [no. (%)] | |
| Age (mean ± SD); range | 64.3±16.1; 21–96 |
| Male sex | 52 (63.4) |
| Acquisition | |
| Community-acquired | 71 (86.6) |
| Hospital-acquired | 11 (13.4) |
| Source of infection | |
| Pneumonia/empyema | 49 (59.8) |
| Meningitis | 9 (11.0) |
| Biliary tract infection | 9 (11.0) |
| Primary bacteremia | 7 (8.5) |
| Epiglottitis | 2 (2.4) |
| Othersa | 6 (7.3) |
| Underlying conditions [no. (%)] | |
| Immunosuppressive therapy | 28 (34.1) |
| Solid organ malignancy | 26 (31.7) |
| Diabetes | 18 (22.0) |
| COPD | 18 (22.0) |
| Heart disease | 15 (18.3) |
| Chronic liver disease | 11 (13.4) |
| Hematologic malignancyb | 9 (11.0) |
| Cerebrovascular disease | 5 (6.1) |
| Organ transplantc | 4 (4.9) |
| HIV | 2 (2.4) |
| Othersd | 10 (12.2) |
| Shock | 18 (22.0) |
| Neutropenia | 8 (9.8) |
| Mortality | |
| <30 days | 17 (20.7) |
Facial cellulites, endometritis, liver abscess, and urinary-tract infection (n = 1, 1.2% each), and peritonitis (n = 2, 2.4%).
Leukemia (n = 3, 3.6%), lymphoma (n = 1, 1.2%), and myeloma (n = 5, 6.1%).
Bone marrow transplant (n = 1, 1.2%), kidney transplant (n = 1, 1.2%), and liver transplantation (n = 2, 2.4%).
Cerebrospinal fluid fistula (n = 2, 2.4%), renal failure, autoimmune disease, and head trauma (n = 1, 1.2% each).
Invasive H. influenzae: sample origin and serotypes
The source of strains in the 82 invasive Hi cases was: blood (n = 70), CSF (n = 3), pleural fluid (n = 2), blood plus CSF (n = 5), and blood plus pleural fluid (n = 2). Serotypes of isolates causing meningitis were NTHi (n = 6) and Hif (n = 1), with one isolate being unavailable (n = 1).
Unfortunately, only isolates from 70 cases (70/82, 85.37%) were viable and available for microbiological studies. The majority of these isolates were NTHi (n = 59, 84.3%), and the frequency of capsulated isolates was low (n = 11, 15.7%). Among 11 capsulated strains, 9 were serotype f (Hif), 1 serotype b (Hib), and 1 serotype e (Hie) (Figure 1). Encapsulated H. influenzae were mainly isolated from blood (n = 9, 81.8%). Both Hib and Hie strains were isolated from patients with pneumonia. The foci of infection of Hif cases were pneumonia (n = 3), epiglottitis (n = 1), meningitis (n = 1), peritonitis (n = 1), facial cellulites (n = 1), and biliary tract infection (n = 1).
Figure 1. Distribution of 82 invasive H. influenzae isolated from adult patients (2008–2013).
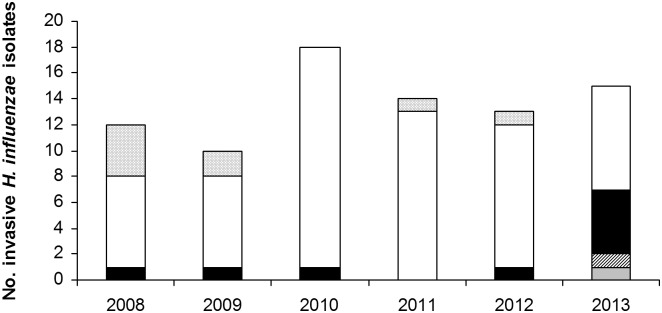
White bar: nontypeable H. influenzae. Black bar: H. influenzae serotype f. Dotted bar: no available isolated for serotyping. Grey bar: H. influenzae serotype b. Lined bar: H. influenzae serotype e.
Antimicrobial susceptibility
All 82 isolates tested by disk diffusion presented fully susceptibility to amoxicillin/clavulanic acid, cefotaxime, ceftriaxone, chloramphenicol, tetracycline, and ciprofloxacin. On the other hand, 23% of the isolates (n = 19) were resistant to cotrimoxazole and 8.5% (n = 7) were resistant to ampicillin due to β-lactamase production. Antimicrobial susceptibility of the 70 available isolates was tested by microdilution. All tested isolates were fully susceptible to amoxicillin/clavulanic acid, cefepime, cefotaxime, ceftriaxone, imipenem, chloramphenicol, tetracycline, and ciprofloxacin (Table 2). Capsulated isolates were fully susceptible to all tested antibiotics, with the exception of one strain that was resistant to rifampin. By contrast, NTHi isolates presented higher resistance rates to cotrimoxazole (27.1%), azithromycin (1.4%), and cefuroxime (1.4%) (Table 2). Seven NTHi isolates (10%) were ampicillin resistant: six (6/70, 8.6%) due to β-lactamase production (MIC>4 mg/L) and one due to a modified PBP3 (MIC = 4 mg/L). In addition, three strains (3/70, 4.3%) presented reduced ampicillin susceptibility (MIC = 2 mg/L). All six β-lactamase producers presented the bla TEM-1 gene.
Table 2. Antimicrobial susceptibility of 70 invasive H. influenzae.
| MIC50 | MIC90 | Range | % S | % I | % R | |
| Antimicrobial | (mg/L) | (mg/L) | (mg/L) | |||
| Ampicillina | 0.25 | 2 | ≤0.12–>4 | 85.7 | 4.3 | 10.0 |
| Amoxicillin/clavulanicacidb | ≤0.5 | 2 | ≤0.5–4 | 100 | ||
| Cefuroxime | 1 | 2 | ≤0.5–8 | 98.6 | 1.4 | |
| Cefepime | ≤0.25 | ≤0.25 | ≤0.25–0.25 | 100 | ||
| Cefotaxime | ≤0.06 | ≤0.06 | ≤0.06–0.06 | 100 | ||
| Ceftriaxone | ≤0.12 | ≤0.12 | ≤0.12 | 100 | ||
| Imipenem | 0.5 | 1 | ≤0.12–2 | 100 | ||
| Chloramphenicol | ≤1 | ≤1 | ≤1 | 100 | ||
| Tetracycline | ≤1 | 2 | ≤1–2 | 100 | ||
| Ciprofloxacin | ≤0.03 | ≤0.03 | ≤0.03 | 100 | ||
| Cotrimoxazolec | ≤0.5 | >2 | ≤0.5–>2 | 72.9 | 1.4 | 25.7 |
| Azithromycin | 1 | 2 | ≤0.12–>4 | 98.6 | 1.4 |
β-lactamase production: 8.6% (n = 6).
The ratio of amoxicillin/clavulanic acid was 2∶1.
The ratio of cotrimoxazole was 1∶19.
In order to determine the mutations in PBP3 the transpeptidase domain of the ftsI gene was sequenced in all the isolates. Thirty-four isolates (48.6%) presented mutations in the ftsI gene (Table 3). The observed mutations allow us to classify the strains into groups I and II, in accordance with Dabernat et al. [17]. The most frequent substitutions were those which were classified as group II (22/34, 64.7%). No isolates were observed in subgroup IId or in groups III and III-like. Additionally, eight isolates (23.5%) presented mutations in the ftsI gene, none of which were at the positions which defined the groups. For this reason, they were considered gBLNAS and classified into the miscellaneous group. All these eight isolates presented similar ampicillin MIC to susceptible strains (≤0.5 mg/L), suggesting that these mutations were not involved in decreased β-lactam susceptibility (Table 3).
Table 3. Amino acid substitutions in PBP3 among 70 invasive H. influenzae strains.
| BLNAR/BLPACR Genotype | Noisolates | Amino acid substitutions | MIC (mg/L) | BLa | |||||||||||||
| Ile 348 | Asp350 | Ala368 | Met377 | Met391 | Ala437 | Gly490 | Ala502 | Val509 | Arg517 | Asn526 | Ala530 | Phe531 | AMP | AMC | |||
| No changesb | 33 | ≤0.12 | ≤0.5 | – | |||||||||||||
| 3 | >4 | ≤0.5 | + | ||||||||||||||
| I | 3 | His | 0.5 | 1 | – | ||||||||||||
| 1 | Thr | His | – | ||||||||||||||
| IIa | 1 | Lys | 1 | 2 | – | ||||||||||||
| 2 | Asn | Glu | Lys | Ser | 1 | 1–2 | – | ||||||||||
| IIb | 1 | Val | Lys | 0.5 | 1 | – | |||||||||||
| 6 | Asn | Ile | Val | Lys | 0.5–4 | 1–4 | – | ||||||||||
| 1 | Asn | Ile | Val | Lys | >4 | 4 | + | ||||||||||
| 2 | Asn | Ile | Glu | Val | Lys | 0.5–1 | 1–2 | – | |||||||||
| 1 | Asn | Ile | Glu | Val | Lys | >4 | 4 | + | |||||||||
| IIc | 2 | Thr | Lys | 1–2 | 2–4 | – | |||||||||||
| 1 | Thr | Lys | >4 | 4 | + | ||||||||||||
| 5 | Asn | Thr | Lys | 1–2 | 2–4 | – | |||||||||||
| Miscellaneousc | 1 | Val | 0.5 | 1 | – | ||||||||||||
| 1 | Thr | ≤0.12 | ≤0.5 | – | |||||||||||||
| 1 | Asn | 0.12 | ≤0.5 | – | |||||||||||||
| 2 | Ile | 0.25 | ≤0.5 | – | |||||||||||||
| 1 | Leu | 0.25 | ≤0.5 | – | |||||||||||||
| 2 | Ser | 0.25 | ≤0.5 | – | |||||||||||||
BL: Beta-lactamase production: + (positive); – (negative).
Isolates without amino acid changes in PBP3 (gBLNAS).
Isolates grouped in the miscellaneous group were not gBLNAR.
Molecular epidemiology
Molecular typing by PFGE revealed 50 different patterns. Fourteen clusters grouped between two and seven related isolates, and 36 patterns were genotypically unique. Hif isolates were grouped into two related clusters, one with seven isolates and the other with two. The other 12 clusters grouped two NTHi isolates each.
Molecular typing by MLST showed high genetic variability among 59 NTHi isolates, which had 51 different sequence types (STs). After eBURST analysis, NTHi STs were distributed into three clonal groups, along with 45 singletons. Clonal group 1 grouped ST103 and ST134, with one isolate each. Group 2 was formed by three isolates, with ST3 (n = 1) and ST367 (n = 2). Finally, clonal group 3 comprised ST14 (n = 1) and a single locus variant (SLV, n = 1). Among the 45 singletons the most frequent ST was ST57, with three isolates. eBURST analysis of all the STs from NTHi isolates published in the MLST database revealed that 75.7% (n = 53) of our invasive isolates belonged to different clonal complexes (CC) defined by the analysis. Twenty-three isolates belonged to the five most prevalent CC among NTHi: CC1, CC3 (n = 5 each), CC41, CC57, and CC472 (n = 3 each).
Capsulated strains, by contrast, were genetically related. All type f isolates belonged to CC124, formed by ST124 (n = 7) and two single locus variants (recA5 and recA43). The type b strain belonged to CC6 (SLV ST6; recA15), while the Hie strain belonged to ST18.
Discussion
The epidemiology of invasive H. influenzae has changed since the introduction of the Hib conjugate vaccine for children, with nontypeable strains being the most frequent etiological agent in most cases of invasive Hi disease in adults. The incidence rate observed among our adults (2.12/100,000) is similar to that reported in the USA and in Europe [6], [22], [23]. As documented in other studies the incidence of invasive Hi disease increased with age (6.8/100,000 in patients ≥65 years old) [6], [23].
In the pre-vaccine era, Hib was the most important cause of invasive disease (mainly meningitis) in healthy children under 5 years of age [1], [4], [22]. In the United States the pre-vaccine incidence of invasive disease in adults was 1.7 cases per 100,000 persons, with Hib being responsible for 50% of invasive diseases due to H. influenzae [11]. In the United Kingdom the incidence of Hib infection in adults was low (9%) and it was assumed to be a consequence of transmission from children [24]. Since the widespread childhood immunization program in the 1990s, Hib infection has decreased considerably worldwide [4]. The percentage of Hib infection cases in adults varies depending on the region. In Illinois, for example, 17.7% of cases were Hib, whereas in Utah the figure was 9% [6], [11]. Only one of our isolates was identified as Hib (1.4%), a lower percentage than previously reported (4.92%) in another Spanish study [25]. Concerning non-b capsulated strains, Hif is currently the most frequent serotype causing invasive disease [1], [5], [11], [25]–[27]. However, despite the fact that in our hospital the number of invasive H. influenzae isolated from 2008–2013 remained stable, an increment in non-b capsulated strains was observed in 2013, when the number of capsulated isolates doubled due to an increase in serotype f.
During the pre-vaccination era, NTHi were not a frequent cause of invasive disease, even though they had been considered an important respiratory pathogen in adults [8]. However, a strain shift has been observed since vaccination, with NTHi being the strains most frequently responsible for invasive infections in adults [1]. NTHi were the most common cause of invasive Hi infection in adults in Illinois (34%), Utah (43%), Manitoba (57%), and Sweden [6], [11], [26], [27]. A recent publication in Spain reported 62% of NTHi among invasive isolates from adults (2004–2009) [25]. In our study, which analyzed invasive isolates from 2008 to 2013, 85.7% of strains were identified as NTHi, following the trend observed in other studies and adding to previously published data on more recently isolated strains.
As reported in other studies [3], [6], [11], most invasive H. influenzae disease infections occurred in older adults (n = 47 patients were ≥65 years old) and in those with underlying conditions. Increased life expectancy and the growing number of patients with underlying conditions may account for the high proportion of invasive Hi disease found in the present study. Pneumonia was the most common type of infection caused by invasive H. influenzae in adults, as reported elsewhere [10], [22], [24],[28]. In line with other studies [3], [6], [11], [28], [29], our patients with invasive H. influenzae infection showed a high mortality rate, although again this could be associated with age and underlying conditions. Nevertheless, we have to acknowledge the small number of cases identified as a limitation of our study.
Following the clinical and epidemiological evaluation of samples the study aimed to investigate the antimicrobial resistance of invasive H. influenzae. Traditionally, the most common mechanism of β-lactam resistance in H. influenzae has been β-lactamase production, although this production has decreased over time [30]–[32]. The percentage of β-lactamase in invasive H. influenzae varies depending on the study, ranging from 10–24% [3], [25], [26], [32]–[38]. In our study the percentage of β-lactamase production was 8.6%, in line with published data although lower than the figures (16.9% and 24.2%) reported in two previous studies about invasive H. influenzae performed in 1999–2000 and 2004–2009 in Spain [3], [25]. β-lactam resistance due to alterations in PBP3 has also been reported worldwide [18], [26], [30], [39], [40]. In the present study, 32.9% of isolates were considered gBLNAR, with ampicillin MIC of 0.5–4 mg/L, presenting relevant mutations in the transpeptidase domain of ftsI. Although these mutations conferred reduced susceptibility to ampicillin, the isolates in question were not considered resistant according to current CLSI and EUCAST breakpoints, the exception being one strain which presented an ampicillin MIC of 4 mg/L. In our experience, the patients infected by BLNAR strains were successfully treated with amoxicillin/clavulanic acid, cefepime, ceftriaxone, piperacillin/tazobacatam, and quinolones (data not shown). Currently, these strains with altered MICs to β-lactams can be successfully treated with these antibiotics; however, the detection of BLNAR strains in the laboratory could improve the knowledge about the epidemiology of H. influenzae. The most common mutations found in invasive isolates were those that classified the strains into group IIb (47.8%), this being consistent with previous data reported by Resman et al., Shuel et al., and Bajanca et al. [26], [33], [37]. By contrast, however, with Spanish data published by García-Cobos et al., who found that group IIc was the most common BLNAR genotype in invasive H. influenzae (42.4%) [25].
Genotyping by PFGE and MLST showed a high diversity among NTHi strains, which were distributed into three clonal groups and 45 singletons. Despite the high genetic variability observed in NTHi, the majority of isolates were grouped according to the most prevalent clonal complexes defined by eBURST, using all the NTHi published in the MLST database. Capsulated strains, by contrast, were clonally related. These results are consistent with other studies that also described this difference in diversity between NTHi and capsular H. influenzae [26], [33], [37]. Moreover, all capsulated strains found in our study belonged to international disseminated global clones. For instance, all nine Hif isolates were grouped in CC124 (ST124 and two single locus variants), which has been identified in the USA and other European countries [25], [33], [37], [38]. Hib and Hie isolates belonged, respectively, to the CC6 and CC18 clones, which have been detected worldwide (http://haemophilus.mlst.net/).
In conclusion, NTHi were the most frequent cause of invasive Hi disease in adults, who frequently presented underlying conditions, and they were associated with a high mortality rate. In our hospital, however, there was an increase in capsular strains, generally serotype f, during 2013. It should also be noted that reduced ampicillin susceptibility was observed in a high percentage of invasive H. influenzae due to mutations in PBP3. Despite the reduction in Hib, continuous monitoring of invasive H. influenzae infections should be performed, not only because of the recent increase in capsulated non-b strains but also in order to detect changes in the epidemiology of invasive H. influenzae.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.
Funding Statement
Funding was provided by Fondo de Investigaciones Sanitarias de la Seguridad Social (PI0901904), and by CIBER de Enfermedades Respiratorias (CIBERES - CB06/06/0037), run by the ISCIII (Instituto de Salud Carlos III, Madrid, Spain). CP was supported by an FPU Grant AP2010-3202 (Formación de Profesorado Universitario, Ministerio de Educación, Spain). SM was supported by “Sara Borrell postdoctoral contract CD10/00298” from the Instituto de Salud Carlos III (ISCIII), Madrid, Spain. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Agrawal A, Murphy TF (2011) Haemophilus influenzae infections in the H. influenzae type b conjugate vaccine era. J Clin Microbiol 49: 3728–3732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Erwin AL, Smith AL (2007) Nontypeable Haemophilus influenzae: understanding virulence and commensal behavior. Trends Microbiol 15: 355–362. [DOI] [PubMed] [Google Scholar]
- 3. Campos J, Hernando M, Roman F, Perez-Vazquez M, Aracil B, et al. (2004) Analysis of invasive Haemophilus influenzae infections after extensive vaccination against H. influenzae type b. J Clin Microbiol 42: 524–529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Peltola H (2000) Worldwide Haemophilus influenzae type b disease at the beginning of the 21st century: global analysis of the disease burden 25 years after the use of the polysaccharide vaccine and a decade after the advent of conjugates. Clin Microbiol Rev 13: 302–317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Dominguez A, Bou R, Carmona G, Latorre C, Pineda V, et al. (2004) Invasive disease caused by Haemophilus influenzae: the sensitivity of statutory reporting. Ann Epidemiol 14: 31–35. [DOI] [PubMed] [Google Scholar]
- 6. Rubach MP, Bender JM, Mottice S, Hanson K, Weng HY, et al. (2011) Increasing incidence of invasive Haemophilus influenzae disease in adults, Utah, USA. Emerg Infect Dis 17: 1645–1650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Eldika N, Sethi S (2006) Role of nontypeable Haemophilus influenzae in exacerbations and progression of chronic obstructive pulmonary disease. Curr Opin Pulm Med 12: 118–124. [DOI] [PubMed] [Google Scholar]
- 8. Foxwell AR, Kyd JM, Cripps AW (1998) Nontypeable Haemophilus influenzae: pathogenesis and prevention. Microbiol Mol Biol Rev 62: 294–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Puig C, Calatayud L, Marti S, Tubau F, Garcia-Vidal C, et al. (2013) Molecular Epidemiology of Nontypeable Haemophilus influenzae Causing Community-Acquired Pneumonia in Adults. PLoS One 8: e82515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Adam HJ, Richardson SE, Jamieson FB, Rawte P, Low DE, et al. (2010) Changing epidemiology of invasive Haemophilus influenzae in Ontario, Canada: evidence for herd effects and strain replacement due to Hib vaccination. Vaccine 28: 4073–4078. [DOI] [PubMed] [Google Scholar]
- 11. Dworkin MS, Park L, Borchardt SM (2007) The changing epidemiology of invasive Haemophilus influenzae disease, especially in persons > or = 65 years old. Clin Infect Dis 44: 810–816. [DOI] [PubMed] [Google Scholar]
- 12. Binks MJ, Temple B, Kirkham LA, Wiertsema SP, Dunne EM, et al. (2012) Molecular surveillance of true nontypeable Haemophilus influenzae: an evaluation of PCR screening assays. PLoS One 7: e34083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Falla TJ, Crook DW, Brophy LN, Maskell D, Kroll JS, et al. (1994) PCR for capsular typing of Haemophilus influenzae . J Clin Microbiol 32: 2382–2386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clinical and Laboratory Standards Institute (2012) Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Growth Aerobically. Approved Standard 8th-ed. M7-A9. Clinical and Laboratory Standards Institute. Wayne, PA.
- 15.Clinical Laboratory Standard Institute (2013) Performance standards for antimicrobial susceptibility testing: 23rd informational supplement. CLSI document M100-S23. Clinical and Laboratory Standards Institute. Wayne, PA.
- 16. Tenover FC, Huang MB, Rasheed JK, Persing DH (1994) Development of PCR assays to detect ampicillin resistance genes in cerebrospinal fluid samples containing Haemophilus influenzae . J Clin Microbiol 32: 2729–2737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Dabernat H, Delmas C, Seguy M, Pelissier R, Faucon G, et al. (2002) Diversity of beta-lactam resistance-conferring amino acid substitutions in penicillin-binding protein 3 of Haemophilus influenzae . Antimicrob Agents Chemother 46: 2208–2218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Garcia-Cobos S, Campos J, Cercenado E, Roman F, Lazaro E, et al. (2008) Antibiotic resistance in Haemophilus influenzae decreased, except for beta-lactamase-negative amoxicillin-resistant isolates, in parallel with community antibiotic consumption in Spain from 1997 to 2007. Antimicrob Agents Chemother 52: 2760–2766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kim IS, Ki CS, Kim S, Oh WS, Peck KR, et al. (2007) Diversity of ampicillin resistance genes and antimicrobial susceptibility patterns in Haemophilus influenzae strains isolated in Korea. Antimicrob Agents Chemother 51: 453–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Hotomi M, Kono M, Togawa A, Arai J, Takei S, et al. (2010) Haemophilus influenzae and Haemophilus haemolyticus in tonsillar cultures of adults with acute pharyngotonsillitis. Auris Nasus Larynx 37: 594–600. [DOI] [PubMed] [Google Scholar]
- 21. Meats E, Feil EJ, Stringer S, Cody AJ, Goldstein R, et al. (2003) Characterization of encapsulated and noncapsulated Haemophilus influenzae and determination of phylogenetic relationships by multilocus sequence typing. J Clin Microbiol 41: 1623–1636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ladhani S, Slack MP, Heath PT, von GA, Chandra M, et al. (2010) Invasive Haemophilus influenzae Disease, Europe, 1996–2006. Emerg Infect Dis 16: 455–463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. MacNeil JR, Cohn AC, Farley M, Mair R, Baumbach J, et al. (2011) Current epidemiology and trends in invasive Haemophilus influenzae disease–United States, 1989–2008. Clin Infect Dis 53: 1230–1236. [DOI] [PubMed] [Google Scholar]
- 24. Sarangi J, Cartwright K, Stuart J, Brookes S, Morris R, et al. (2000) Invasive Haemophilus influenzae disease in adults. Epidemiol Infect 124: 441–447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Garcia-Cobos S, Arroyo M, Perez-Vazquez M, Aracil B, Lara N, et al. (2014) Isolates of beta-lactamase-negative ampicillin-resistant Haemophilus influenzae causing invasive infections in Spain remain susceptible to cefotaxime and imipenem. J Antimicrob Chemother 69: 111–116. [DOI] [PubMed] [Google Scholar]
- 26. Resman F, Ristovski M, Forsgren A, Kaijser B, Kronvall G, et al. (2012) Increase of beta-lactam-resistant invasive Haemophilus influenzae in Sweden, 1997 to 2010. Antimicrob Agents Chemother 56: 4408–4415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Tsang RS, Sill ML, Skinner SJ, Law DK, Zhou J, et al. (2007) Characterization of invasive Haemophilus influenzae disease in Manitoba, Canada, 2000–2006: invasive disease due to non-type b strains. Clin Infect Dis 44: 1611–1614. [DOI] [PubMed] [Google Scholar]
- 28. Perdue DG, Bulkow LR, Gellin BG, Davidson M, Petersen KM, et al. (2000) Invasive Haemophilus influenzae disease in Alaskan residents aged 10 years and older before and after infant vaccination programs. JAMA 283: 3089–3094. [DOI] [PubMed] [Google Scholar]
- 29. Laupland KB, Schonheyder HC, Ostergaard C, Knudsen JD, Valiquette L, et al. (2011) Epidemiology of Haemophilus influenzae bacteremia: a multi-national population-based assessment. J Infect 62: 142–148. [DOI] [PubMed] [Google Scholar]
- 30. Heilmann KP, Rice CL, Miller AL, Miller NJ, Beekmann SE, et al. (2005) Decreasing prevalence of beta-lactamase production among respiratory tract isolates of Haemophilus influenzae in the United States. Antimicrob Agents Chemother 49: 2561–2564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Perez-Trallero E, Martin-Herrero JE, Mazon A, Garcia-Delafuente C, Robles P, et al. (2010) Antimicrobial resistance among respiratory pathogens in Spain: latest data and changes over 11 years (1996–1997 to 2006–2007). Antimicrob Agents Chemother 54: 2953–2959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Sill ML, Tsang RS (2008) Antibiotic susceptibility of invasive Haemophilus influenzae strains in Canada. Antimicrob Agents Chemother 52: 1551–1552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bajanca-Lavado MP, Simoes AS, Betencourt CR, Sa-Leao R (2013) Characteristics of Haemophilus influenzae invasive isolates from Portugal following routine childhood vaccination against H. influenzae serotype b (2002–2010). Eur J Clin Microbiol Infect Dis. 10.1007/s10096-013-1994-6. [DOI] [PubMed]
- 34. Giufre M, Cardines R, Caporali MG, Accogli M, D’Ancona F, et al. (2011) Ten years of Hib vaccination in Italy: prevalence of non-encapsulated Haemophilus influenzae among invasive isolates and the possible impact on antibiotic resistance. Vaccine 29: 3857–3862. [DOI] [PubMed] [Google Scholar]
- 35. Ladhani S, Heath PT, Ramsay ME, Slack MP (2008) Changes in antibiotic resistance rates of invasive Haemophilus influenzae isolates in England and Wales over the last 20 years. J Antimicrob Chemother 62: 776–779. [DOI] [PubMed] [Google Scholar]
- 36. Shuel M, Law D, Skinner S, Wylie J, Karlowsky J, et al. (2010) Characterization of nontypeable Haemophilus influenzae collected from respiratory infections and invasive disease cases in Manitoba, Canada. FEMS Immunol Med Microbiol 58: 277–284. [DOI] [PubMed] [Google Scholar]
- 37. Shuel M, Hoang L, Law DK, Tsang R (2011) Invasive Haemophilus influenzae in British Columbia: non-Hib and non-typeable strains causing disease in children and adults. Int J Infect Dis 15: e167–e173. [DOI] [PubMed] [Google Scholar]
- 38. Sill ML, Law DK, Zhou J, Skinner S, Wylie J, et al. (2007) Population genetics and antibiotic susceptibility of invasive Haemophilus influenzae in Manitoba, Canada, from 2000 to 2006. FEMS Immunol Med Microbiol 51: 270–276. [DOI] [PubMed] [Google Scholar]
- 39. Jansen WT, Verel A, Beitsma M, Verhoef J, Milatovic D (2006) Longitudinal European surveillance study of antibiotic resistance of Haemophilus influenzae . J Antimicrob Chemother 58: 873–877. [DOI] [PubMed] [Google Scholar]
- 40. Shuel ML, Tsang RS (2009) Canadian beta-lactamase-negative Haemophilus influenzae isolates showing decreased susceptibility toward ampicillin have significant penicillin binding protein 3 mutations. Diagn Microbiol Infect Dis 63: 379–383. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.