

Abstract
Alpha-melanocyte stimulating hormone (α-MSH) is a neuropeptide exhibiting anti-inflammatory activity in experimental models of autoimmune diseases. However, no studies thus far have examined the effects of α-MSH on systemic lupus erythematosus (SLE). This study aimed to determine the effects of an α-MSH agonist in induced murine lupus. Here we employed female Balb/cAn mice in which lupus was induced by pristane. Groups of lupus animals were treated daily with the α-MSH analogue [Nle4, DPhe7]-α-MSH (NDP–MSH) (1·25 mg/kg) injected intraperitoneally or saline for 180 days. Normal animals comprised the control group. Arthritis incidence, plasma immunoglobulin (Ig)G isotypes, anti-nuclear antibodies (ANA) and plasma cytokines were evaluated. Renal function was assessed by proteinuria and histopathological lesion. Glomerular levels of IgG, α-smooth muscle actin (α-SMA), inducible nitric oxide synthase (iNOS), C3, CD3, melanocortin receptors (MCR)1, corticotrophin-releasing factor (CRF) and α-MSH was estimated by immunohistochemistry. When compared with normal controls, lupus animals exhibited increased arthritis, IgG levels, ANA, interleukin (IL)-6, IL-10, proteinuria and mesangial cell proliferation together with glomerular expression of α-SMA and iNOS. Glomerular expression of MCR1 was reduced in lupus animals. NDP-MSH treatment reduced arthritis scores by 70% and also diminished IgG1 and IgG2a levels and ANA incidence. In the glomerulus, NDP–MSH treatment reduced cellularity by 50% together with reducing IgG deposits, and expression levels of α-SMA, iNOS and CRF were also all decreased. Taken together, our results suggest for the first time that α-MSH treatment improves several parameters of SLE disease activity in mice, and indicate that this hormone is an interesting potential future treatment option.
Keywords: α-MSH, arthritis, cytokines, experimental lupus, nephritis
Introduction
Systemic lupus erythematosus (SLE) is an autoimmune syndrome exhibiting heterogeneous outcomes that can affect multiple organs. The disease is characterized by tissue damage induced by chronic inflammation, the presence of autoantibodies and autoreactive lymphocytes and cytokine imbalances [1]. The kidneys are particularly susceptible to injury, as almost 60% of SLE patients will present renal involvement at some point. Immune complex deposition and inflammatory milieu are the key factors that amplify kidney lesions [2,3].
The neuro-immuno-endocrine system is known to participate in autoimmune diseases regulation. A disturbed hypothalamic–pituitary–adrenal (HPA) axis response has been associated with SLE in humans and in animal models [4,5]. Accordingly, pituitary polypeptide adrenocorticotrophic hormone (ACTH), a melanocortin family component, was widely used during the 1950s to treat both lupus and nephrotic syndrome. However, ACTH was quickly replaced by oral synthetic corticoid treatment due to administration advantages [6].
Today, interest in melanocortin neuropeptides has re-emerged through α-melanocyte-stimulating hormone (α-MSH). This peptide, derived from ACTH, has arisen as a new tool to control the inflammatory process [7]. Multiple experimental studies have described the anti-inflammatory properties of α-MSH during ischaemia–reperfusion injury [8] and in autoimmune diseases including uveitis [9], encephalomyelitis [10], contact hypersensitivity [11] and arthritis [12]. Although anti-inflammatory effects have also been reported in models of kidney disease, improving haemodynamic failure and reducing the severity of renal lesions [13–15], limited effects in restoring impaired renal function and proteinuria have been described [14,16,17].
Melanocortin receptors (MCR) are present in multiple tissues and cell types, including the glomerulus, renal tubules, neutrophils, monocytes, dendritic cells and lymphocytes [18]. MCR1 activation prevents nuclear factor kappa B (NF-κB) translocation and reduces the production of several inflammatory mediators [19]. In experimental models of nephritis, α-MSH increases animal survival and improves glomerular filtration. Simultaneously, it decreases apoptosis, fibrosis, neutrophil infiltration, cytokine expression and reactive oxygen species production [13–15].
Induced lupus models that resemble the SLE human disease are of pivotal relevance because they allow the study of mechanisms involved in the disease onset [20]. Among these models, intraperitoneal injection of mice with the hydrocarbon oil pristane (2,6,10,14-tetramethylpentadecane) is remarkable in that it faithfully exhibits the clinical manifestation and laboratorial abnormalities that are characteristic of SLE [21]. In addition, the pristane lupus-like model can be induced in the Balb/c strain, which is a more reliable colony of animals compared with spontaneous lupus mice [20].
Considering that SLE patients present antigen presentation defects, cytokine release, arthritis and nephritis, and suffer alterations in the HPA axis, a plausible connection exists between melanocortins and lupus pathophysiology. However, no study discusses the effect of α-MSH on lupus. Therefore, the present study evaluated the effects of an α-MSH analogue treatment on pristane-induced murine lupus.
Materials and methods
Animals
This protocol was approved by the local Research Ethic Committee of Hospital Clinics/School of Medicine, University of São Paulo, under process number 0612/09·09. All animal care and experimental procedures were developed in strict conformity with Universities Federation for Animals Welfare and the Committee of Brazilian College of Experimental Animals (COBEA).
Because the pristane-induced model presents female preponderance as human disease [22], only adult female mice were employed. Twenty adult BALB/cAnUnib mice, 8·5 weeks old and weighing 20–25 g, were employed. The animals were kept in conventional colony rooms at the Rheumatology Division facility with controlled temperature (23 ± 1°C) and a 12-h light/dark cycle with food in pellets and water ad libitum. To avoid stress interference, no experiments were initiated until 15 days after animal arrival.
Experimental design and α-MSH treatment
Lupus was induced in 15 mice with a single intraperitoneal (i.p.) injection of 0·5 ml pristane (2,6,10,14 tetramethylpentadecane; Sigma Chemical Co., St Louis, MO, USA) that had been previously filtered through a 0·22 μm membrane (Millipore, Billerica, MA, USA) [23]. Five control mice received the same volume of 0·9% saline. Peripheral blood (250 μl) was collected from the submandibular vein at baseline and 180 days after treatment.
Alpha-melanocyte-stimulating hormone analogue [Nle4, DPhe7]-α-MSH (NDP-MSH; Melanotan-I) was purchased from Peptides International (Louisville, KY, USA). The α-MSH analogue [Nle4, DPhe7]-α-MSH was chosen due to its higher stability, biodisponibility and resistance to proteolytic degradation compared with the native peptide. In addition, it does not interfere with endogenous glucocorticoid production [24,25].
The NDP-MSH treatment was prepared according to the manufacturer's instructions. NDP-MSH was first dissolved in acetic acid solution and then added to 0·9% saline (final acid concentration 0·06%). Daily treatment with the hormone analogue started just after pristane injection and was always performed between 8:00 and 10:00 h [26,27]. Mice with pristane-induced lupus (LM) were allocated randomly in two groups of five or 10 animals: LM treated with saline (0·9%, pH = 5, LM) or LM treated with the NDP-MSH 1·25 mg/kg day i.p. (LM-MSH). The control group (control) consisted of normal animals treated with saline i.p. Before killing, the animals were anaesthetized with xylazine (5 mg/kg) and ketamine (50 mg/kg) administered i.p.
Total leucocyte counts
Total peripheral blood leucocytes were counted in a Neubauer chamber diluted 1:20 with Turk.
Autoantibody detection
Immunoglobulin (Ig)G1, IgG2a, IgG2b and IgG3 levels in diluted plasma (1:15 000) were measured by enzyme-linked immunosorbent assay (ELISA). IgG isotype standards from Southern Biotechnology (Birmingham, AL, USA) were used for standard curve fitting at concentrations of 15·62–1000 ng/ml.
Anti-nuclear and cytoplasmic autoantibodies (ANA) were detected by indirect immunofluorescence using a home-made HEp-2 cells assay. Slides were incubated for 40 min with 1:200 murine plasma in phosphate-buffered saline (PBS), washed and then incubated for 30 min with 1:50 fluorescein isothiocyanate-conjugated goat anti-mouse IgG H + L (Southern Biotechnology). After another wash, slides containing cells were analysed with a fluorescence microscope.
Quantification of plasma cytokines
Cytokine levels [interleukin (IL)-6, IL-10, tumour necrosis factor (TNF)-α and interferon (IFN)-γ] were analysed using a fluorokine mouse multi-analyte profile kit (R&D Systems, Minneapolis, MN, USA). According to the manufacture's protocol, plasma samples were diluted 1:2 and incubated with anti-cytokine beads overnight at 4°C. Events were counted on a luminex 200 system (Luminex, Austin, TX, USA). Data analyses were performed using the Milliplex Analyst version 3·5.5·0 (Vigenetech, Carlisle, MA, USA). A five-parameter regression formula was used to calculate the sample concentration. Data were expressed in pg/ml.
Arthritis evaluation
The arthritis severity was assessed 180 days after pristane inoculation, using a scoring system that considers oedema, erythema and number of affected joints as: 1 = small, 2 = moderate, 3 = marked and 4 = severe. Thus, the maximum total score per animal was 16 [28].
Renal function evaluation
Before killing, the animals were individually contained to collect spot urine. All samples were analysed using proteinuria measurements kits (Labtest Diagnosis, Minas Gerais, Brazil). To estimate urinary albumin concentration, 10 μl of urine was run on 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) and was Coomassie-stained. A pattern of bovine serum albumin (BSA) (Sigma Chemical Co.) was used to adjust the standard curve with concentrations from 0·0625 to 0·5 mg/ml. The bands were measured by densitometric analyses with ImageJ software.
Histopathological evaluation
After killing, the kidneys were perfused in situ with saline, the right kidney was removed and fixed in 10% buffered formalin, embedded in paraffin and sectioned at 3 μm thickness in the transversal plane containing the renal long axis. Slides were stained using haematoxylin and eosin (H&E) and periodic acid-Schiff (PAS) stains to highlight the glomerulus and pricrosirius red to stain collagen fibres. Cellularity was quantified on H&E-stained slides by counting the total glomerular cell nuclei. At least 30 glomeruli/slides were assessed, and the results were expressed as number of nuclei per glomerulus. Sections stained with PAS were graded as: 1+ = mild focal mesangial hypercellularity; 2+ = moderate mesangial hypercellularity; 3+ = complex endocapillary hypercellularity sometimes with mild sclerosis or necrosis; 4+ = severe endocapillary proliferative glomerulonephritis with necrosis or crescent formation. Scores ≥ 2+ were considered to be positive [29]. Pricrosirius red-stained slides were analysed under polarized light using an Olympus camera attached to an Olympus BX-51 microscope (Center Valley, PA, USA), and the collagen area was determined based on positive staining in the image analyses system.
Immunohistochemistry
Glomerular expression of α-smooth muscle actin (α-SMA), T cell marker CD3, complement component 3 (C3), corticotrophin-releasing factor (CRF), inducible nitric oxide synthase (iNOS), α-MSH, MCR1 and IgG were determined by immunohistochemical analyses in deparaffinized kidney sections. After rehydration, the endogenous peroxidase activity was ablated by incubation in 3% hydrogen peroxide for 10 min. Next, the sections were incubated with Tris/ethylenediamine tetraacetic acid (EDTA) (10 mM/1 mM buffer, pH 9·0) for 25 min and incubated with a biotin/avidin blocking solution (Dako, Glostrup, Denmark). Primary antibodies anti-α-SMA (ab5694; 1:100; Abcam, Cambridge, UK), anti-CD3 (ab5690; 1:100; Abcam), anti-C3 (sc-28294; 1:50; Santa Cruz Biotech, Santa Cruz, CA, USA), anti-CRF (h-019-06; 1:100; Phoenix Pharmaceuticals, Burlingame, CA, USA), anti-α-MSH (h-043-01; 1:100; Phoenix Pharmaceuticals), anti-MCR1 (ABIN686287, 1:100; Antibodies-online, Aachen, Germany), anti-iNOS (PA5-16855; 1:200; Thermo Scientific, Rockford, IL, USA) and anti-IgG (LS-C59195; 1:400; LifeSpan, Bellevue, WA, USA) were added to each section and incubated overnight at 4°C. After PBS washing, the slides were incubated for 30 min with EnVision-horseradish peroxidase (HRP), labelled streptavidin–biotin (LSAB)-HRP or Advance-HRP (Dako). Enzyme sandwich reactions were developed using 3,3′-diaminobenzidine (Sigma Chemical Co.), and then the slides were washed, counter-coloured with haematoxylin and mounted with Permount.
Image analyses
The slides were digitally archived using a Pannoramic Scan instrument (software version 1·11·25·0; 3DHistech, Budapest, Hungary) with a ×20 objective and expanded focus. Total levels of collagen fibres, α-SMA, CD3, C3, CRF, α-MSH, MCR1, iNOS and IgG in the glomerulus were quantified using Image-ProPlus version 4·1 software for Windows (Media Cybernetics, Silver Spring, MD, USA).
The positively stained areas were determined by colour threshold. These procedures generated a file containing a colour selection data, which was applied afterwards to the kidney sections. The results of each marker were expressed as the ratio of positively stained area per total glomerular area (μm2).
Statistical analyses
Significance tests were performed with spss version 17·0.1 software (spss, Inc., Chicago, IL, USA) or GraphPad prism version 6·0 Software (GraphPad software, San Diego, CA, USA). Data were initially compared with the Gaussian curve through the K–S test. Parametric data were analysed using one-way analysis of variance (anova) with pairwise post-test comparisons by the Neuman–Keuls method. Non-parametric data were compared using anova followed by modified Tukey's test and results are expressed as median and interquartile ranges. The incidence of ANA, arthritis and renal lesion were compared using Pearson's χ2 test. P < 0·05 was considered statistically significant.
Results
Inhibitory effect of NDP–MSH on arthritis development
Arthritis incidence and severity is shown in Fig. 1. LM mice exhibited a high incidence of arthritis (80%) and a mean score of 5·2. Treatment with NDP-MSH reduced both incidence (40%) and arthritis score (1·6, P < 0·05 versus LM).
Fig. 1.
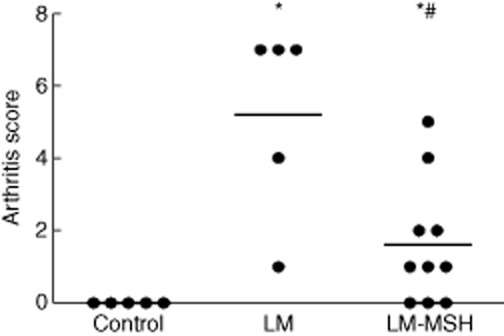
[Nle4, DPhe7]-α-MSH (NDP-MSH treatment attenuated the development of arthritis in lupus mice (LM) induced by pristane. LM treated with NDP-MSH (LM-MSH group) presented significant reduction of arthritis score. Severity of arthritis was assessed using a scoring system that considers oedema, erythema and number of affected joints as: 1 = small, 2 = moderate, 3 = marked and 4 = severe. Results are expressed as raw data with median; *P < 0·05 versus control and #P < 0·05 versus LM.
When compared with control animals, both the LM and LM-MSH groups exhibited greater weight gain by day 180 of the experiment (P < 0·01). LM animals presented with leucocytosis 180 days after pristane administration. NDP-MSH treatment had no impact on the white blood cell count. The relationships between right kidney, spleen and liver weights and body weights were not different between the groups.
NDP–MSH treatment reduces IgG1, IgG2a levels and ANA frequency
As shown in Fig. 2, plasma levels of IgG1 and IgG2a increased 180 days after pristane inoculation compared with control animals. NDP-MSH treatment reduced IgG1 and IgG2a levels (P < 0·05 and P < 0·001 versus LM, respectively). IgG2b and IgG3 levels did not differ between the groups.
Fig. 2.
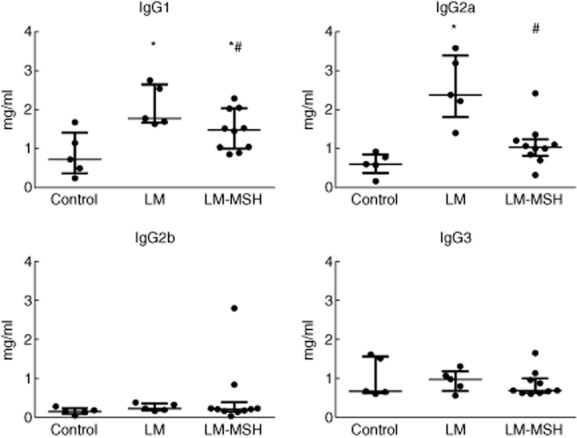
Plasma levels of immunoglobulin (Ig)G1, IgG2a, IgG2b and IgG3 180 days after pristane-induced lupus. [Nle4, DPhe7]-α-MSH (NDP-MSH treatment reduced IgG1 and IgG2a plasma levels 180 days after lupus induction. No significant change was observed in IgG2b and IgG3 plasma levels. Results are expressed as median with interquartile range. *P < 0·05 versus control and #P < 0·05 versus LM.
The plasma ANA assessment is presented in Fig. 3a. One hundred per cent (five of five) of LM exhibited ANA positivity at titres ≥1:200. None of the control animals (none of five) showed ANA positivity (P < 0·01) compared to 30% of LM-MSH animals (P < 0·01 versus LM). Figure 3b illustrates the absence of anti-nuclear antibodies registered in control animals and Fig. 3c shows a positive pattern verified in HEp-2 cells incubated with LM plasma.
Fig. 3.
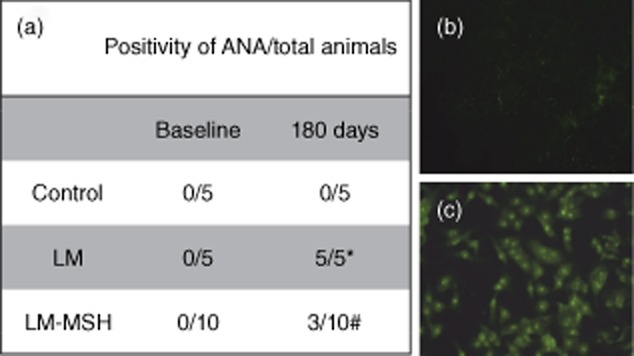
Effect of [Nle4, DPhe7]-α-MSH (NDP-MSH on anti-nuclear and anti-cytoplasmic plasma antibodies (ANA). ANA was detected by indirect immunofluorescence using human epithelial type 2 (HEp-2) cells home-made assay. (a) Negative pattern, (b) positive pattern of ANA (original magnification ×400). ANA was detected in all lupus mice (LM) 180 days after pristane lupus induction (c), whereas only 30% (three of 10) of LM treated with NDP-MSH (LM-MSH) had ANA. *P < 0·05 compared with control animals, #P < 0·05 compared with LM.
NDP–MSH effect on cytokine production
As shown in Table 1, both plasma IL-6 and IL-10 levels increased 180 days after pristane inoculation (P < 0·05 versus control). When compared with the LM group, treatment with NDP-MSH slightly reduced IL-6 and IL-10 levels. The LM-MSH group showed increased TNF-α levels when compared with the LM group (P < 0·01). IFN-γ levels were below the detection range in all groups.
Table 1.
Effect of [Nle4, DPhe7]-α-MSH (NDP-MSH treatment on cytokine production of lupus mice (LM) and control mice
| Cytokine | Control (n = 5) | LM (n = 5) | LM-MSH (n = 10) |
|---|---|---|---|
| IL-6 (pg/ml) | 5·94 ± 2·36 | 84·31 ± 54·11* | 42·95 ± 13·85* |
| IL-10 (pg/ml) | 2·35 ± 0·45 | 5·34 ± 0·81* | 4·49 ± 0·68 |
| TNF-α (pg/ml) | 0·047 ± 0·016 | 2·12 ± 0·63* | 3·82 ± 0·52*# |
| IFN-γ (pg/ml) | – | – | – |
Results are expressed as mean ± standard error. *P < 0·05 versus control and #P < 0·05 versus LM. IL = interleukin; IFN = interferon; TNF = tumour necrosis factor.
NDP–MSH treatment prevented glomerular lesions
Figure 4 shows the results of the renal function evaluation in spot urine samples. At the end of the experimental period LM had significant proteinuria (P < 0·001), which was not affected by the NDP-MSH treatment (Fig. 4a). The same pattern was observed in albuminuria (Fig. 4b).
Fig. 4.
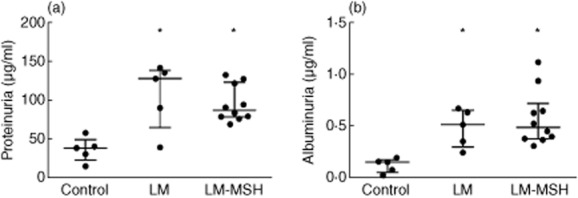
Effect of [Nle4, DPhe7]-α-MSH (NDP-MSH treatment on urinary parameters. Pristane-induced lupus mice (LM) developed proteinuria (a) and albuminuria (b). NDP-MSH treatment of LM had no effect on these urinary parameters. Results are expressed as median with interquartile range. *P < 0·05 versus control.
Histopathological analysis data are presented in Fig. 5. LM showed increased glomerular cellularity (Fig. 5a) compared with the control group (P < 0·01). The treatment of LM with NDP-MSH reduced this cellularity (P < 0·05 versus LM). Figure 5b shows the renal damage assessed 180 days after lupus induction. Lupus animals exhibited a profile characterized by moderate histopathological mesangial hypercellularity (score = 2) that was higher than the degree of injury observed in the control animals (P < 0·05). NDP-MSH treatment reduced this score (P < 0·05 versus LM). Notably, 80% of the animals treated with NDP-MSH showed a degree of damage less than 2. The fibrosis evaluation in pricrosirius red-stained slides revealed no difference between the control and LM groups (data not shown).
Fig. 5.
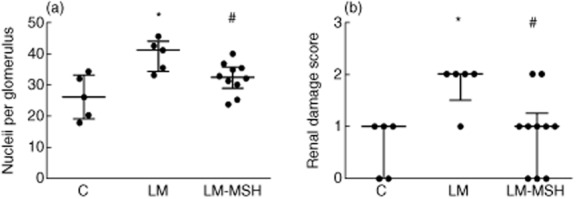
[Nle4, DPhe7]-α-MSH (NDP-MSH treatment limited renal lesion in pristane-induced lupus-like model. The analysis of glomerular cellularity and renal damage scores, assessed in periodic acid-Schiff (PAS)-stained sections, graded as: 1+ = mild focal mesangial hypercellularity; 2+ = moderate mesangial hypercellularity; 3+ = complex endocapillary hypercellularity sometimes with mild sclerosis or necrosis; 4+ = severe endocapillary proliferative glomerulonephritis, revealed an amelioration of these parameters in LM treated with NDP-MSH. The results are expressed as median with interquartile range, *P < 0·05 versus control and #P < 0·05 compared to the LM.
Effect of NDP–MSH treatment on glomerular IgG deposits and the expression of α-SMA, iNOS, CRF, α-MSH and MCR1
Glomerular deposition of IgG and the expression of α-SMA and iNOS are shown in Fig. 6. When compared with the control group, LM showed increased expression of all markers, which were decreased by NDP-MSH treatment. The figures depict the glomerular appearance of control animals (Fig. 6b), LM (Fig. 6c) and LM-MSH animals (Fig. 6d). The expression of CD3 and C3 did not differ between the groups (data not shown).
Fig. 6.

Effect of [Nle4, DPhe7]-α-MSH (NDP-MSH on glomerular immunoglobulin (Ig)G deposit, α-smooth muscle actin (α-SMA) and inducible nitric oxide synthase (iNOS) expression in pristane-induced lupus-like model. IgG deposits detected by immunohistochemistry in the glomeruli of lupus mice (LM) were abrogated by the treatment with NDP-MSH (a–d). Similarly, α-SMA expression (e–h) and iNOS expression (i–l) were also decreased significantly in LM submitted to NDP-MSH therapy. The results of each marker were expressed as the ratio of positively stained area per total glomerular area (μm2). Data are expressed as median with interquartile range. *P < 0·05 versus control, #P < 0·05 versus LM.
Glomerular expression of MCR1 was reduced in LM (P < 0·01 versus control) and in LM-MSH animals (P < 0·001 versus control). The expression of neuropeptides, CRF and α-MSH were similar in control and LM. Treatment with NDP-MSH reduced the expression of glomerular CRF (P < 0·01) but did not affect α-MSH expression (P = 0·12) (Fig. 7).
Fig. 7.

[Nle4, DPhe7]-α-MSH (NDP-MSH reduced the expression of corticotrophin-releasing factor (CRF) in the glomerulus of lupus mice (LM) (a–d). The expression of melanocortin receptor (MCR)1 diminished in LM, but was not influenced by NDP-MSH treatment (e–h). NDP–-MSH treatment reduced the α-MSH expression in the glomeruli of LM (i–l). The results of each marker were expressed as the ratio of positively stained area per total glomerular area (μm2). Data are expressed as median with interquartile range. *P < 0·05 versus control, #P < 0·05 versus LM.
Discussion
This study shows, for the first time to our knowledge, that the α-MSH super-analogue NDP-MSH improves several parameters of lupus disease progression in mice.
The chosen experimental model was an important component of our protocol, as pristane presence in human blood suggests a positive correlation with autoimmune disease incidence [30]. In this context, after pristane injection, mice developed a lupus-like syndrome characterized by the presence of hypergammaglobulinaemia, ANA, specific antibodies, arthritis, glomerulonephritis and abnormal cytokine production [21,22,29]. The incidence and severity of joint inflammation, one of the most common SLE manifestations, were suppressed by the treatment employed in this study. In accordance with our findings, neuropeptide therapy in other arthritis models was effective in controlling joint inflammation [12,31]. In human disease, α-MSH has been related to rheumatoid arthritis pathogenesis as a possible compensatory mechanism [32]. In the present study, the ability of NDP-MSH treatment reducing lupus activity was confirmed by the observed reductions in hypergammaglobulinaemia and ANA incidence. Accordingly, α-MSH treatment also reduced IgG1 and IgG2a levels in an airway inflammation model [33]. The importance of this impairment is corroborated by findings that treating pristane-induced lupus with other immunomodulatory compounds also changes these parameters [34,35].
Lupus immune dysfunction is characterized by uncontrolled cytokine production. Levels of IL-6 and IL-10 are tightly correlated with the activity and severity of the disease in humans [36] and in experimental models [37–39]. The importance of IL-6 in lupus is evidenced by the reductions in autoantibody production and renal injury observed in IL-6 knock-out mice injected with pristane [40]. Furthermore, clinical studies show that patients exhibit reduced SLE activity after treatment with anti-IL-6R [41]. Interestingly, patients with chronic renal disease present an inverse correlation between IL-6 and α-MSH levels [42]. Accordingly, our lupus animals exhibited increased IL-6 production, and NDP-MSH treatment reduced this production by 37·9%, although not statistically significantly. IL-10 levels are also elevated in mice with pristane-induced lupus, and attenuation of the disease is accompanied by IL-10 reduction [43]. Lupus patients treated with anti-IL10 therapy show impaired disease activity [44]. In our study, we did not observe a significant effect of α-MSH on IL-10 production. In this context, the effect of this neuropeptide on IL-10 remains controversial, as previous studies have reported both reduced IL-10 levels [13] or no effect [45,46]. As part of the mechanism of action of α-MSH is on NF-B preventing nucleus translocation, the anti-inflammatory activity certainly involves the reduction of proinflammatory markers [47,48]. Surprisingly, our treatment with NDP-MSH increases TNF-α levels. Although TNF-α is a proinflammatory cytokine, the role of TNF-α in lupus has not been definitively established. In this context, clinical studies have shown that anti-TNF therapy worsens disease activity and triggers a lupus-like disease [49]. In experimental models, the administration of high doses of TNF-α does not alter and sometimes delays nephritis development [50,51].
The most severe complication of SLE is renal involvement. In the present study, as well as in humans, not all mice developed nephritis. Previous studies of pristane-induced lupus showed similar incidence rates and the same patterns of glomerular lesion [29,38]. Regarding markers of renal injury, our schedule of treatment with NDP-MSH did not alter proteinuria/albuminuria, but it did improve other parameters of glomerular injury. Accordantly, similar findings were observed in other nephritis studies, where α-MSH did not affect plasma urea or creatinine but did improve histological markers [14,15]. The inefficacy of α-MSH on proteinuria was also reported on New Zealand black/white F1 mice [16]. Our data show that lupus is characterized by high glomerular α-SMA expression. In accordance with these results, mesangial cells of patients with LN class II also present increased α-SMA expression [52,53]. The administration of α-MSH also reduced glomerular expression α-SMA glomerular in models of liver damage [54] and pulmonary fibrosis [55]. In our view, the glomerular injury induced by pristane is not particularly severe. In our study, we detected no changes in CD3 and C3 expression or in glomerular fibrosis. In this context, only patients with proliferative NL (classes III and IV) present high infiltration of T lymphocytes in the kidney and necrosis [56].
One interesting point regarding the relationships between melanocortins is the negative feedback effect of α-MSH on CRF [57]. Our study corroborates this finding, as we observed minor expression of CRF in the NDP-MSH-treated group.
The increased expression of iNOS and increases in nitric oxide are associated with renal damage in SLE patients [58] and mice [59]. Pristane injection also increases iNOS expression in kidney [39]. To our knowledge, this is the first study showing the ability of α-MSH to reduce glomerular iNOS expression in a lupus-like disease model. Studies in vivo and in vitro supplied experimental data to support our results [60,61]. Indeed, iNOS expression is associated with inflammation and iNOS blockade ameliorates arthritis in animals [62].
Although the melanocortin receptor MCR1 is widely expressed in the kidney [18], no study has addressed its expression in human or experimental lupus. Our data show that NDP-MSH treatment reduces glomerular MCR1. In accordance with these results, down-regulation of the receptor has been documented previously following NDP-MSH treatment [63]. Nevertheless, animals with lupus but not receiving treatment also presented minor MCR1 expression. Attenuated MCR1 expression has been described previously during ischaemic kidney disease in rats and in human liver [64,65]. The reason for this effect remains unknown; however, the explanation could be receptor desensitization by high α-MSH levels during the development of the disease. The current study was unable to test this hypothesis, as we did not measure native α-MSH levels in our animals. In this regard, the effects of native α-MSH were completely overwhelmed by the high doses of its hormone analogue [66].
Altogether, our results show that α-MSH opposes several factors involved in lupus pathogenesis. The current treatment of lupus patients involves a large therapeutic arsenal that is often incapable of controlling multiple disease symptoms. We believe that the neuropeptide NDP-MSH shows potential to serve as an important new tool in treating this inflammatory condition.
Acknowledgments
The authors are grateful to Maria Aurora Gomes da Silva, Maria de Fátima de Almeida, Margarete Borges Galhardo Vendramini, Cleonice Bueno and Antonio dos Santos Filho for their skilful technical assistance and to Ana Maria de Lauro Castrucci, Anna Catania, Mauro Perretti, Minoru Satoh and Monique Kowalski Schimitid for helpful discussions. Isac de Castro performed the statistic analyses. This study was supported by grant 2009/54549-8 from the State of São Paulo Foundation for Research Support (FAPESP) and from the Brazilian Council of Scientific and Technologic Development (CNPq). The funders had no role in study design, data collection and analyses, decision to publish or preparation of the manuscript.
Author contributions
S. B. V. M., D. A. C. B. and I. L. N. conceived and designed the experiments. D. A. C. B., T. V. B. and D. M. A. C. M. performed the experiments. D. A. C. B. and S. B. V. M. analysed the data. S. B. V. M., I. L. N. and D. M. A. C. M. contributed reagents, materials and analysis tools. S. B. V. M. and D. A. C. B. wrote the paper.
Disclosures
The authors have declared that no conflicts of interest exist.
References
- 1.Munoz LE, Janko C, Schulze C, et al. Autoimmunity and chronic inflammation – two clearance-related steps in the etiopathogenesis of SLE. Autoimmun Rev. 2010;10:38–42. doi: 10.1016/j.autrev.2010.08.015. [DOI] [PubMed] [Google Scholar]
- 2.Davidson A, Aranow C. Lupus nephritis: lessons from murine models. Nat Rev Rheumatol. 2010;6:13–20. doi: 10.1038/nrrheum.2009.240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zubair A, Frieri M. Lupus nephritis: review of the literature. Curr Allergy Asthma Rep. 2013;13:580–586. doi: 10.1007/s11882-013-0394-4. [DOI] [PubMed] [Google Scholar]
- 4.Harle P, Straub RH, Wiest R, et al. Increase of sympathetic outflow measured by neuropeptide Y and decrease of the hypothalamic–pituitary–adrenal axis tone in patients with systemic lupus erythematosus and rheumatoid arthritis: another example of uncoupling of response systems. Ann Rheum Dis. 2006;65:51–56. doi: 10.1136/ard.2005.038059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shanks N, Moore PM, Perks P, Lightman SL. Alterations in hypothalamic–pituitary–adrenal function correlated with the onset of murine SLE in MRL+/+ and lpr/lpr mice. Brain Behav Immun. 1999;13:348–360. doi: 10.1006/brbi.1998.0535. [DOI] [PubMed] [Google Scholar]
- 6.Bomback AS, Radhakrishnan J. Treatment of nephrotic syndrome with adrenocorticotropic hormone (ACTH) Discov Med. 2011;12:91–96. [PubMed] [Google Scholar]
- 7.Patel HB, Leoni G, Melendez TM, Sampaio ALF, Perretti M. Melanocortin control of cell trafficking in vascular inflammation. Adv Exp Med Biol. 2010;681:88–106. doi: 10.1007/978-1-4419-6354-3_7. [DOI] [PubMed] [Google Scholar]
- 8.Giuliani D, Minutoli L, Ottani A, et al. Melanocortins as potential therapeutic agents in severe hypoxic conditions. Front Neuroendocrinol. 2012;33:179–193. doi: 10.1016/j.yfrne.2012.04.001. [DOI] [PubMed] [Google Scholar]
- 9.Taylor AW, Yee DG, Nishida T, Namba K. Neuropeptide regulation of immunity. The immunosuppressive activity of alpha-melanocyte-stimulating hormone (alpha-MSH) Ann NY Acad Sci. 2000;917:239–247. doi: 10.1111/j.1749-6632.2000.tb05389.x. [DOI] [PubMed] [Google Scholar]
- 10.Taylor AW, Kitaichi N. The diminishment of experimental autoimmune encephalomyelitis (EAE) by neuropeptide alpha-melanocyte stimulating hormone (alpha-MSH) therapy. Brain Behav Immun. 2008;22:639–646. doi: 10.1016/j.bbi.2007.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rheins LA, Cotleur AL, Kleier RS, Hoppenjans WB, Saunder DN, Nordlund JJ. Alpha-melanocyte stimulating hormone modulates contact hypersensitivity responsiveness in C57/BL6 mice. J Invest Dermatol. 1989;93:511–517. doi: 10.1111/1523-1747.ep12284064. [DOI] [PubMed] [Google Scholar]
- 12.Ceriani G, Diaz J, Murphree S, Catania A, Lipton JM. The neuropeptide alpha-melanocyte-stimulating hormone inhibits experimental arthritis in rats. Neuroimmunomodulation. 1994;1:28–32. doi: 10.1159/000097087. [DOI] [PubMed] [Google Scholar]
- 13.Doi K, Hu X, Yuen PS, et al. AP214, an analogue of alpha-melanocyte-stimulating hormone, ameliorates sepsis-induced acute kidney injury and mortality. Kidney Int. 2008;73:1266–1274. doi: 10.1038/ki.2008.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kolgazi M, Arbak S, Alican I. The effect of alpha-melanocyte stimulating hormone on gentamicin-induced acute nephrotoxicity in rats. J Appl Toxicol. 2007;27:183–188. doi: 10.1002/jat.1191. [DOI] [PubMed] [Google Scholar]
- 15.Lee SY, Jo SK, Cho WY, Kim HK, Won NH. The effect of alpha-melanocyte-stimulating hormone on renal tubular cell apoptosis and tubulointerstitial fibrosis in cyclosporine A nephrotoxicity. Transplantation. 2004;78:1756–1764. doi: 10.1097/01.tp.0000144332.44435.ab. [DOI] [PubMed] [Google Scholar]
- 16.Hart DA. Evidence that LiCl enhancement of the survival of NZB/W lupus mice is not mediated by alpha-melanocyte stimulating hormone. J Trace Microprobe Tech. 1998;16:111–118. [Google Scholar]
- 17.Miyaji T, Hu X, Star RA. Alpha-melanocyte-simulating hormone and interleukin-10 do not protect the kidney against mercuric chloride-induced injury. Am J Physiol Renal Physiol. 2002;282:F795–801. doi: 10.1152/ajprenal.00203.2001. [DOI] [PubMed] [Google Scholar]
- 18.Lindskog A, Ebefors K, Johansson ME, et al. Melanocortin 1 receptor agonists reduce proteinuria. J Am Soc Nephrol. 2010;21:1290–1298. doi: 10.1681/ASN.2009101025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Eves PC, Haycock JW. Melanocortin signalling mechanisms. Adv Exp Med Biol. 2010;681:19–28. doi: 10.1007/978-1-4419-6354-3_2. [DOI] [PubMed] [Google Scholar]
- 20.Perry D, Sang A, Yin Y, Zheng YY, Morel L. Murine models of systemic lupus erythematosus. J Biomed Biotechnol. 2011;2011:271694. doi: 10.1155/2011/271694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bigazzi PE. Murine lupus induced by tetramethylpentadecane: an animal model of systemic human autoimmunity. Clin Immunol. 2005;114:97–99. doi: 10.1016/j.clim.2004.09.009. [DOI] [PubMed] [Google Scholar]
- 22.Smith-Bouvier DL, Divekar AA, Sasidhar M, et al. A role for sex chromosome complement in the female bias in autoimmune disease. J Exp Med. 2008;205:1099–1108. doi: 10.1084/jem.20070850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Satoh M, Reeves WH. Induction of lupus-associated autoantibodies in BALB/c mice by intraperitoneal injection of pristane. J Exp Med. 1994;180:2341–2346. doi: 10.1084/jem.180.6.2341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sawyer TK, Sanfilippo PJ, Hruby VJ, et al. 4-Norleucine, 7-D-phenylalanine-alpha-melanocyte-stimulating hormone: a highly potent alpha-melanotropin with ultralong biological activity. Proc Natl Acad Sci USA. 1980;77:5754–5758. doi: 10.1073/pnas.77.10.5754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dorr RT, Dawson BV, al-Obeidi F, Hadley ME, Levine N, Hruby VJ. Toxicologic studies of a superpotent alpha-melanotropin, [Nle4, D-Phe7]alpha-MSH. Invest New Drugs. 1988;6:251–258. doi: 10.1007/BF00173642. [DOI] [PubMed] [Google Scholar]
- 26.Sinha PS, Schiöth HB, Tatro JB. Roles of the melanocortin-4 receptor in antipyretic and hyperthermic actions of centrally administered α-MSH. Brain Res. 2004;1001:150–158. doi: 10.1016/j.brainres.2003.12.007. [DOI] [PubMed] [Google Scholar]
- 27.Ribeiro RA, Vale ML, Thomazzi SM, et al. Involvement of resident macrophages and mast cells in the writhing nociceptive response induced by zymosan and acetic acid in mice. Eur J Pharmacol. 2000;387:111–118. doi: 10.1016/s0014-2999(99)00790-6. [DOI] [PubMed] [Google Scholar]
- 28.Patten C, Bush K, Rioja I, et al. Characterization of pristane-induced arthritis, a murine model of chronic disease: response to antirheumatic agents, expression of joint cytokines, and immunopathology. Arthritis Rheum. 2004;50:3334–3345. doi: 10.1002/art.20507. [DOI] [PubMed] [Google Scholar]
- 29.Richards HB, Satoh M, Jennette JC, Croker BP, Yoshida H, Reeves WH. Interferon-gamma is required for lupus nephritis in mice treated with the hydrocarbon oil pristane. Kidney Int. 2001;60:2173–2180. doi: 10.1046/j.1523-1755.2001.00045.x. [DOI] [PubMed] [Google Scholar]
- 30.Dahlgren J, Takhar H, Anderson-Mahoney P, Kotlerman J, Tarr J, Warshaw R. Cluster of systemic lupus erythematosus (SLE) associated with an oil field waste site: a cross sectional study. Environ Health. 2007;6:8. doi: 10.1186/1476-069X-6-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gomez-Sanmiguel AB, Martin AI, Nieto-Bona MP, et al. Systemic alpha melanocyte stimulating hormone administration decreases arthritis-induced anorexia and muscle wasting. Am J Physiol Regul Integr Comp Physiol. 2013;304:R877–886. doi: 10.1152/ajpregu.00447.2012. [DOI] [PubMed] [Google Scholar]
- 32.Catania A, Gerloni V, Procaccia S, et al. The anticytokine neuropeptide alpha-melanocyte-stimulating hormone in synovial fluid of patients with rheumatic diseases: comparisons with other anticytokine molecules. Neuroimmunomodulation. 1994;1:321–328. doi: 10.1159/000097183. [DOI] [PubMed] [Google Scholar]
- 33.Raap U, Brzoska T, Sohl S, et al. Alpha-melanocyte-stimulating hormone inhibits allergic airway inflammation. J Immunol. 2003;171:353–359. doi: 10.4049/jimmunol.171.1.353. [DOI] [PubMed] [Google Scholar]
- 34.Zhou LL, Wei W, Si JF, Yuan DP. Regulatory effect of melatonin on cytokine disturbances in the pristane-induced lupus mice. Mediators Inflamm. 2010;2010:951210. doi: 10.1155/2010/951210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ouyang S, Gong B, Li JZ, et al. Structural insights into a human anti-IFN antibody exerting therapeutic potential for systemic lupus erythematosus. J Mol Med (Berl) 2012;90:837–846. doi: 10.1007/s00109-012-0866-3. [DOI] [PubMed] [Google Scholar]
- 36.Chun HY, Chung JW, Kim HA, et al. Cytokine IL-6 and IL-10 as biomarkers in systemic lupus erythematosus. J Clin Immunol. 2007;27:461–466. doi: 10.1007/s10875-007-9104-0. [DOI] [PubMed] [Google Scholar]
- 37.Chae BS, Shin TY, Kim DK, Eun JS, Leem JY, Yang JH. Prostaglandin E-2-mediated dysregulation of proinflammatory cytokine production in pristane-induced lupus mice. Arch Pharm Res. 2008;31:503–510. doi: 10.1007/s12272-001-1185-6. [DOI] [PubMed] [Google Scholar]
- 38.Chowdhary VR, Grande JP, Luthra HS, David CS. Characterization of haemorrhagic pulmonary capillaritis: another manifestation of pristane-induced lupus. Rheumatology (Oxf) 2007;46:1405–1410. doi: 10.1093/rheumatology/kem117. [DOI] [PubMed] [Google Scholar]
- 39.Akaogi J, Yamada H, Kuroda Y, Nacionales DC, Reeves WH, Satoh M. Prostaglandin E2 receptors EP2 and EP4 are up-regulated in peritoneal macrophages and joints of pristane-treated mice and modulate TNF-alpha and IL-6 production. J Leukoc Biol. 2004;76:227–236. doi: 10.1189/jlb.1203627. [DOI] [PubMed] [Google Scholar]
- 40.Richards HB, Satoh M, Shaw M, Libert C, Poli V, Reeves WH. Interleukin 6 dependence of anti-DNA antibody production: evidence for two pathways of autoantibody formation in pristane-induced lupus. J Exp Med. 1998;188:985–990. doi: 10.1084/jem.188.5.985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shirota Y, Yarboro C, Fischer R, Pham TH, Lipsky P, Illei GG. Impact of anti-interleukin-6 receptor blockade on circulating T and B cell subsets in patients with systemic lupus erythematosus. Ann Rheum Dis. 2013;72:118–128. doi: 10.1136/annrheumdis-2012-201310. [DOI] [PubMed] [Google Scholar]
- 42.Airaghi L, Garofalo L, Cutuli MG, et al. Plasma concentrations of alpha-melanocyte-stimulating hormone are elevated in patients on chronic haemodialysis. Nephrol Dial Transplant. 2000;15:1212–1216. doi: 10.1093/ndt/15.8.1212. [DOI] [PubMed] [Google Scholar]
- 43.Urbonaviciute V, Starke C, Pirschel W, et al. Toll-like receptor 2 is required for autoantibody production and development of renal disease in pristane-induced lupus. Arthritis Rheum. 2013;65:1612–1623. doi: 10.1002/art.37914. [DOI] [PubMed] [Google Scholar]
- 44.Llorente L, Richaud-Patin Y, Garcia-Padilla C, et al. Clinical and biologic effects of anti-interleukin-10 monoclonal antibody administration in systemic lupus erythematosus. Arthritis Rheum. 2000;43:1790–1800. doi: 10.1002/1529-0131(200008)43:8<1790::AID-ANR15>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- 45.Figueiredo J, Ferreira AE, Silva RL, et al. NDP-MSH inhibits neutrophil migration through nicotinic and adrenergic receptors in experimental peritonitis. Naunyn Schmiedebergs Arch Pharmacol. 2013;386:311–318. doi: 10.1007/s00210-013-0834-7. [DOI] [PubMed] [Google Scholar]
- 46.Catania A, Lonati C, Sordi A, Leonardi P, Carlin A, Gatti S. The peptide NDP-MSH induces phenotype changes in the heart that resemble ischemic preconditioning. Peptides. 2010;31:116–122. doi: 10.1016/j.peptides.2009.09.030. [DOI] [PubMed] [Google Scholar]
- 47.Haycock JW, Wagner M, Morandini R, Ghanem G, Rennie IG, Mac Neil S. Alpha-melanocyte-stimulating hormone inhibits NF-kappaB activation in human melanocytes and melanoma cells. J Invest Dermatol. 1999;113:560–566. doi: 10.1046/j.1523-1747.1999.00739.x. [DOI] [PubMed] [Google Scholar]
- 48.Ichiyama T, Zhao H, Catania A, Furukawa S, Lipton JM. Alpha-melanocyte-stimulating hormone inhibits NF-kappaB activation and IkappaBalpha degradation in human glioma cells and in experimental brain inflammation. Exp Neurol. 1999;157:359–365. doi: 10.1006/exnr.1999.7064. [DOI] [PubMed] [Google Scholar]
- 49.Almoallim H, Al-Ghamdi Y, Almaghrabi H, Alyasi O. Anti-tumor necrosis factor-alpha induced systemic lupus erythematosus. Open Rheumatol J. 2012;6:315–319. doi: 10.2174/1874312901206010315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jacob CO, McDevitt HO. Tumour necrosis factor-alpha in murine autoimmune ‘lupus’ nephritis. Nature. 1988;331:356–358. doi: 10.1038/331356a0. [DOI] [PubMed] [Google Scholar]
- 51.Brennan DC, Yui MA, Wuthrich RP, Kelley VE. Tumor necrosis factor and IL-1 in New Zealand black/white mice. Enhanced gene expression and acceleration of renal injury. J Immunol. 1989;143:3470–3475. [PubMed] [Google Scholar]
- 52.Makni K, Jarraya F, Khabir A, et al. Renal alpha-smooth muscle actin: a new prognostic factor for lupus nephritis. Nephrology (Carlton) 2009;14:499–505. doi: 10.1111/j.1440-1797.2009.01140.x. [DOI] [PubMed] [Google Scholar]
- 53.Hurtado A, Asato C, Escudero E, et al. Clinicopathologic correlations in lupus nephritis in Lima, Peru. Nephron. 1999;83:323–330. doi: 10.1159/000045424. [DOI] [PubMed] [Google Scholar]
- 54.Lee TH, Jawan B, Chou WY, et al. Alpha-melanocyte-stimulating hormone gene therapy reverses carbon tetrachloride induced liver fibrosis in mice. J Gene Med. 2006;8:764–772. doi: 10.1002/jgm.899. [DOI] [PubMed] [Google Scholar]
- 55.Xu PB, Mao YF, Meng HB, Tian YP, Deng XM. Sty39, a novel alpha-melanocyte-stimulating hormone analogue, attenuates bleomycin-induced pulmonary inflammation and fibrosis in mice. Shock. 2010;35:308–314. doi: 10.1097/SHK.0b013e3181f8f15e. [DOI] [PubMed] [Google Scholar]
- 56.Yoshimoto S, Nakatani K, Iwano M, et al. Elevated levels of fractalkine expression and accumulation of CD16+ monocytes in glomeruli of active lupus nephritis. Am J Kidney Dis. 2007;50:47–58. doi: 10.1053/j.ajkd.2007.04.012. [DOI] [PubMed] [Google Scholar]
- 57.Tozawa F, Suda T, Dobashi I, Ohmori N, Kasagi Y, Demura H. Central administration of alpha-melanocyte-stimulating hormone inhibits corticotropin-releasing factor release in adrenalectomized rats. Neurosci Lett. 1994;174:117–119. doi: 10.1016/0304-3940(94)90133-3. [DOI] [PubMed] [Google Scholar]
- 58.Wang JS, Tseng HH, Shih DF, Jou HS, Ger LP. Expression of inducible nitric oxide synthase and apoptosis in human lupus nephritis. Nephron. 1997;77:404–411. doi: 10.1159/000190316. [DOI] [PubMed] [Google Scholar]
- 59.Njoku C, Self SE, Ruiz P, Hofbauer AF, Gilkeson GS, Oates JC. Inducible nitric oxide synthase inhibitor SD-3651 reduces proteinuria in MRL/lpr mice deficient in the NOS2 gene. J Investig Med. 2008;56:911–919. doi: 10.231/JIM.0b013e3181889e13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Star RA, Rajora N, Huang J, Stock RC, Catania A, Lipton JM. Evidence of autocrine modulation of macrophage nitric oxide synthase by alpha-melanocyte-stimulating hormone. Proc Natl Acad Sci USA. 1995;92:8016–8020. doi: 10.1073/pnas.92.17.8016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Chiao H, Kohda Y, McLeroy P, Craig L, Housini I, Star RA. Alpha-melanocyte-stimulating hormone protects against renal injury after ischemia in mice and rats. J Clin Invest. 1997;99:1165–1172. doi: 10.1172/JCI119272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.de Mello SB, Novaes GS, Laurindo IM, Muscara MN, Maciel FM, Cossermelli W. Nitric oxide synthase inhibitor influences prostaglandin and interleukin-1 production in experimental arthritic joints. Inflamm Res. 1997;46:72–77. doi: 10.1007/s000110050086. [DOI] [PubMed] [Google Scholar]
- 63.Siegrist W, Stutz S, Eberle AN. Homologous and heterologous regulation of alpha-melanocyte-stimulating hormone receptors in human and mouse melanoma cell lines. Cancer Res. 1994;54:2604–2610. [PubMed] [Google Scholar]
- 64.Lee YS, Park JJ, Chung KY. Change of melanocortin receptor expression in rat kidney ischemia-reperfusion injury. Transplant Proc. 2008;40:2142–2144. doi: 10.1016/j.transproceed.2008.07.101. [DOI] [PubMed] [Google Scholar]
- 65.Gatti S, Colombo G, Turcatti F, et al. Reduced expression of the melanocortin-1 receptor in human liver during brain death. Neuroimmunomodulation. 2006;13:51–55. doi: 10.1159/000094513. [DOI] [PubMed] [Google Scholar]
- 66.Nelson ME, Bryant SM, Aks SE. Melanotan II injection resulting in systemic toxicity and rhabdomyolysis. Clin Toxicol. 2012;50:1169–1173. doi: 10.3109/15563650.2012.740637. [DOI] [PubMed] [Google Scholar]