

Abstract
Alzheimer’s disease (AD) is a progressive neurodegenerative disease that causes substantial public health care burdens. Intensive efforts have been made to find effective and safe disease-modifying treatment and symptomatic intervention alternatives against AD. Smart Soup (SS), a Chinese medicine formula composed of Rhizoma Acori Tatarinowii (AT), Poria cum Radix Pini (PRP) and Radix Polygalae (RP), is a typical prescription against memory deficits. Here, we assessed the efficacy of SS against AD. Oral administration of SS ameliorated the cognitive impairment of AD transgenic mice, with reduced Aβ levels, retarded Aβ amyloidosis and reduced Aβ-induced gliosis and neuronal loss in the brains of AD mice. Consistently, SS treatment reduced amyloid-related locomotor dysfunctions and premature death of AD transgenic Drosophila. Mechanistic studies showed that RP reduced Aβ generation, whereas AT and PRP exerted neuroprotective effects against Aβ. Taken together, our study indicates that SS could be effective against AD, providing a practical therapeutic strategy against the disease.
Introduction
Alzheimer’s disease (AD) is a fatal and progressive neurodegenerative disease which is characterized by the persistent worsening of cognitive function and daily living performance [1], [2], and it has been considered one of the most significant health, social and economic crises of this century. AD is a complex disease, and its pathological hallmarks include senile plaques formed with amyloid peptides, tangles containing twisted tau proteins, destroyed synapses, abnormal brain inflammation, eventual neuronal death and severe brain shrinkage [3]. Current FDA-approved Alzheimer’s drugs are only symptomatic interventions, such as acetylcholinesterase inhibitors and N-methyl-D-aspartate (NMDA) receptor antagonists, which are effective only for about half of the patients for approximately 6–12 months [4]. Up till now, no disease-modifying medications have been approved in the last 10 years. Considering that, as with many other complex diseases, the pathogenesis of AD has a multifactorial basis that includes both genetic and environmental risk factors, a successful therapeutic strategy against this disease might be a set of concerted pharmacological interventions that interact with multiple targets including Aβ/tau pathology, abnormal inflammation and neuronal loss.
The formulae of traditional Chinese medicine (TCM) have been well documented in various Chinese ancient literatures as prescriptions for specific ailments. According to the major precepts of TCM, these TCM formulae emphasize not only the symptoms but on restoring and maintaining the body homeostasis, which is very similar to the rationale of modern multi-targeted therapeutics [5], [6]. At present, TCM, as China Food and Drug Administration (CFDA)-approved and controlled medications, is widely practiced side by side with modern western medicine in almost all of China’s hospitals and clinics. TCM herbal formulae, including Smart Soup (SS), which have been applied for many centuries, are still prescribed by Chinese medical physicians to patients with aging-related cognitive impairment. SS, a three-herb formula officially documented in Gu Jin Yi Jian, a book published in 1576, is composed of Rhizoma Acori Tatarinowii (AT), Poria cum Radix Pini (PRP) and Radix Polygalae (RP). Each component is frequently used in different TCM formulae for their pharmacological efficacies against dysfunctions of the central nervous system (CNS, Table S1). AT has been shown to exhibit a neuroprotective action and attenuates learning and memory deficits [7]. PRP has been reported to possess sedative activity [8]. RP shows repairing effects on the memory and behavioral deficits in rats [9], exhibits neuroprotective effects [10], [11], enhances cognition and memory in elderly adults [12], [13]. This study was to assess the efficacy of SS against AD.
Materials and Methods
Ethics Statement
All animal experiments were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The animal protocols were approved by the Biological Research Ethics Committee, Shanghai Institutes for biological Sciences, Chinese Academy of Sciences. Animal pain and discomfort were minimized with efforts.
Preparation and the quality analysis of SS
The drug materials were purchased and identified according to the rigid specifications set by Chinese Pharmacopeia (2010 Edition). The CFDA-approved single-herb granules of Rhizoma Acori Tatarinowii (AT), Poria cum Radix Pini (PRP) and Radix Polygalae (RP) were obtained from Tianjiang Pharmaceutical, Jiangyin, China. The granule-mixed Smart Soup (SS-G) were prepared by mixing 10 g of AT, 10 g of PRP and 10 g of RP granules to a concentration of 1 g/ml in water.
The chemical constituent identification of each batch of SS was performed using HPLC-TOF/MS. In detail, an aliquot of 1 ml of SS-G was centrifuged at 12,000 rpm. The supernatant was filtered and used for analysis. HPLC-TOF/MS was performed on a 1200 Series HPLC instrument (Agilent, Waldbronn, Germany) coupled with an Agilent 6224 Accurate-Mass TOF LC/MS. The chromatographic separations were performed at 25°C on an Apollo-C18 reversed-phase column (4.6×250 mm i.d., 5 µm, Grace) connected to an EasyGuard Kit C18 guard column (4×2 mm, Grace). The separation was conducted with an acetonitrile/water gradient with 0.5% formic acid. The injection volume was 20 µl for MS analyses.
Quality analysis of SS using HPLC fingerprints
To assure the quality and thereby warrant the safety and effectiveness of the SS, the chromatographic fingerprints of SS were established and characterized using HPLC. Detection was performed at a wavelength of 320 nm at room temperature. Similarity analysis was performed using similarity evaluation system for TCM chromatographic fingerprints (Version 2004A, Chinese Pharmacopeia Commission) as recommended by CFDA.
APP/PS1 transgenic mice and drug treatment
The APPswe/PS1dE9 (APP/PS1) double-transgenic mice (The Jackson Laboratory, stock number 004462) were used in our investigation [14]–[16]. The mice were maintained and genotyped according to the guidance of Jackson Laboratory. The transgene-negative wild type (WT) littermates were used as age-matched controls.
APP/PS1 and WT mice were chronically administered 200 µl of SS (1 g/ml) or vehicle only (water) per 20 g mouse body weight by gavage once per day from 7 to 9 months old (n = 8–12 mice per group).
Morris water maze test
The Morris water maze (MWM) was performed as described [17]–[19]. The apparatus was a circular pool of 120 cm diameter filled with water with small white plastic balls maintained at 23.0±0.5°C. A transparent platform of 11 cm diameter 1 cm below the water surface was placed at a fixed point of one quadrant. Animals were brought to the behavior room, acclimatized and trained.
The training consisted of 10 consecutive days, with four trials per day. On day 4 and 7, a probe trial was performed, followed by four training trials. On the 11th day, a single probe trial was conducted. Swim paths were monitored using an automated tracking system (Ethovision XT software).
Objective recognition test
Tests were performed as previously described [20], [21] with modifications. The detailed procedures are schematically represented in Fig. S1. The apparatus consisted of an evenly illuminated soundproof box with a Plexiglas box (25 cm×25 cm×25 cm) inside. The procedure included four phases: pre-habituation, habituation, training and testing. The animals were familiarized with the environment for at least one day. On the 1st day of the experiment, the mice were randomly ordered and habituated to the empty box for 5 min. On the 2nd and 3rd day, each mouse was allowed to freely explore two identical objects, which were located at points with same distance from the nearest corner. On the 4th day, during the training phase, each mouse was allowed to explore the identical objects for 10 min first. After a one-hour interval, during the 10-min testing phase, the mouse was returned to the same box with one familiar object switched to a novel one.
To preclude the existence of olfactory cues, each mouse had its own packing paper in the box that wiped thoroughly before each testing.
Object exploration time was the length of time when a mouse was sniffing, directing its nose to and pawing the object. The exploration time was recorded in a double-blinded manner. The location preference in the training phase and recognition index in the testing phase were calculated as following: Location preference means the time exploring one object relative to the time exploring two objects, and Recognition index means the time exploring the novel object relative to the time exploring two objects.
Immunohistochemistry and image analysis
The mice were anesthetized and transcardiac perfused with phosphate-buffered saline (PBS) buffer and then with 4% paraformaldehyde (PFA) in PBS. 30-µm thick brain sections were prepared and immunostained using a mouse antibody against Aβ (6E10, Covance) for amyloid plaques, a polyclonal rabbit antibody against GFAP (DAKO) for astrocytes, a mouse antibody against CD11b (BD) to assess microglia or anti-NeuN antibodies (Millipore) to assess neurons. Images were captured using a microscope (Carl Zeiss). Quantification was performed using Image-Pro Plus 5.1 software (Media Cybernetic), and the percentage of antibody-immunoreactive area was calculated. The plaques were identified as area >1,500 µm2, and the number of plaques were also calculated as three groups: 1,500 µm2< plaque size <3,000 µm2; 3,000 µm2<plaque size <6,000 µm2; plaque size >6,000 µm2. Four to five coronal sections were analyzed per mouse.
ELISA for Aβ
Hippocampal and cortical extracts were prepared as previously reported [22]. The accumulation of human Aβ40 and Aβ42 in these extracts was quantified using ELISA kits (ExCell Bio). Fly head extracts were prepared and the Aβ levels were assessed as reported [23]. Aβ40 and Aβ42 in SK-N-SH-APPsw cell or HEK293-APPsw cell culture medium were also measured with ELISA.
Drosophila culture and drug treatments
We used the pan-neuronal elav-GAL4 to express transgenes as described [24]. The upstream activating sequence (UAS) transgenic lines of Aβ42 and APP/BACE were provided by Dr. FD Huang and have been described in detail [25]. Canton S (CS) flies were used as WT controls. These flies were kept in a 25°C incubator with 65% humidity and 12 h light/12 h dark circle. The drugs were mixed with liquid food, 0.5 g of instant food (Q/SCQC0005S, Nestle) and 0.05% methyl p-hydroxybenzoate (Sinopharm) in a total of 1.4 ml of sterile distilled water. Every 3 days, the fly food was changed.
Survival assay
Sixty flies of each group were cultured at 25°C. The number of dead flies was recorded every day. Survival rates were calculated using the Kaplan–Meier estimation.
Locomotor assay
The locomotor assay of flies was performed as described previously with minor modifications [26]. Briefly, treated ten male flies (n = 30 for each group) were placed in a plastic 25-ml tube. After a 30-min recovering phase, flies were gently tapped to the bottom of the tube. The fly behavior was recorded with a video camera. After 10 s of climbing, the number of flies between the 0, 5, 10, 15, 20 and 25 ml scale marks were recorded (Fig. S2). The results for each group of flies are calculated by the formula below:
 Climbing Index = (flies above 20 ml scale mark) × 1 + (flies between 15 and 20 ml scale marks) × 0.8 + (flies between 10 and 15 ml scale marks) × 0.6 + (flies between 5 and 10 ml scale marks) × 0.4 + (flies below 5 ml scale mark) × 0.2.
Climbing Index = (flies above 20 ml scale mark) × 1 + (flies between 15 and 20 ml scale marks) × 0.8 + (flies between 10 and 15 ml scale marks) × 0.6 + (flies between 5 and 10 ml scale marks) × 0.4 + (flies below 5 ml scale mark) × 0.2.
Western blot analyses
Fly head lysate preparation and western blot analyses were performed as described previously [23], [27]. Briefly, thirty fly heads from each group were collected and lysed in RIPA buffer containing protease inhibitors. HEK293-APPsw cell lysates were also prepared. Proteins were separated on 16% tricine gel and probed with antibodies against APP C-terminal and β-actin (Sigma-Aldrich).
Preparation of Aβ peptides
The peptides were prepared according to the protocols described by Stine [28]. Briefly, hexafluoroisopropanol (HFIP)-treated Aβ42 peptides (Anaspec) were resuspended in dimethyl sulfoxide (DMSO). For the oligomeric conditions, the peptide was then diluted to a concentration of 100 µM with Ham’s F12 and incubated at 4°C for 24 hours. After centrifugation 10 min at 14,000 g, the supernatant with soluble Aβ42 oligomers was added to cultures. The soluble oligomeric Aβ peptides were confirmed using atomic force microscopy (AFM) and western blot analysis.
Thioflavin-T fluorescence assay
The Thioflavin-T (Th-T, Sigma-Aldrich) fluorescence assay was performed to measure amyloid fibril formation [29]. Th-T was prepared as a stock at 2 mM (avoid exposure to light) and filtered through a 0.22 µm filter. The Th-T stock solution was diluted into PBS on the day of analysis. HFIP-treated Aβ peptides were dissolved in DMSO at 5 mM, and then diluted in Th-T/PBS with a final concentration of 15 µM (Aβ) and 20 µM (Th-T). The Aβ/Th-T solutions were incubated at 37°C with or without TCM for six days in a black 96-well-plate (PerkinElmer). Fluorescence was measured using an EnVision multilabel plate reader (PerkinElmer), at excitation and emission wavelengths of 440 nm and 490 nm, respectively. Fluorescence of 20 µM Th-T/PBS solution was measured and used as blank. The fluorescence intensity reflects the degree of Aβ aggregation.
Primary culture of neuron
Primary neurons were cultured as described previously [30], [31]. The cortices and hippocampi obtained from newborn C57BL/6 mice were used. 5×104 cells per well in DMEM/Ham’s F12 containing 10% FBS (Invitrogen) were plated onto poly-D-lysine (Sigma-Aldrich) coated 96-well plates and maintained at 37°C for four hours, and the culture medium was then switched to B27/Neurobasal medium (Invitrogen). On day 6, the cells pre-treated with TCMs for two hours were incubated with 5 µM Aβ42 oligomers and TCMs for another 48 hours. Staining of primary neurons with a Tuj1 antibody (Covance) was imaged and analyzed using Operetta (PerkinElmer).
CellTiter-Glo assay
The primary neurons, SK-N-SH-APPsw cells and HEK293-APPsw cells were assessed for viability using the CellTiter-Glo luminescent cell viability assay (Promega) following the manufacturer’s instructions.
TUNEL assay
The assay was performed using the Kit from Roche following the manufacturer’s protocol. Primary neurons were pre-treated with TCMs for two hours followed by incubation with 5 µM Aβ42 oligomers and TCMs for another 24 hours. The total numbers of DAPI-stained or TUNEL-positive cells were counted.
Atomic Force Microscope (AFM)
Amyloid peptides for AFM analysis were prepared according to the protocols described by Stine [28]. First, HFIP-treated Aβ42 peptides (Anaspec) were dissolved in DMSO. 5 mM Aβ42 in DMSO were diluted in H2O to 100 µM and used immediately as Aβ42 monomers. 5 mM Aβ42 in DMSO were diluted in Ham’s F12 to 100 µM and incubated at 4°C for 24 hours and used as Aβ42 oligomers. Aβ42 fibrils were prepared by diluting 5 mM Aβ42 in DMSO with 10 mM HCl to 100 µM and incubated at 37°C for 24 hours. For AFM analysis, Aβ42 monomers, oligomers and fibrils samples were further diluted to a final concentration of 10 µM with distilled H2O and spotted on freshly cleaved mica disks for 3 minutes. After being rinsed three times with deionized water, the mica disks were dried overnight at room temperature. AFM was performed in tapping mode using a Molecular Force Prove 3D (MFP-3D, Asylum Research) with NSC11 cantilevers (MikroMasch, 48 Newton/meter spring constant). The data were analyzed with Igor-Pro (Wavemetrix). The resonance frequency was applied in the 330 kHz range, the scan rate was 1 Hz, and the resolution was 512×512 pixels.
Statistical analyses
GraphPad Prism 5.0 was used. The data are shown as the mean ± S.E.M. The Kaplan-Meier test was employed to compare the differences between the survival curves using SPSS 16.0. Group differences were analyzed with one-way ordinary or repeated-measures analysis of variance (ANOVA) followed by Tukey multiple comparisons test. Student’s independent t test was applied for comparisons of two groups. Differences were significant when P<0.05.
Results
SS treatment ameliorates learning and memory deficits in APP/PS1 mice
Traditionally, TCM formulae are prepared as decoctions, i.e., the herbs are boiled for a fixed time, and the liquid is used. Recently, CFDA-approved single-herb granules have been widely accepted as alternatives for decoctions. These CFDA-approved single-herb granules are clearly documented for their chemical fingerprints and pharmacokinetic parameters along with standard manufacturing protocols, which are not only suitable for large-scale industry production but are also more appropriate for evaluation of their efficacies and exploration of the underlying molecular mechanisms. Thus, as described in the Methods section, we prepared SS using the CFDA-approved single-herb granules according to the recipe (granule-mixed SS, SS-G). We first analyzed the SS-G by HPLC-TOF/MS. The total ion current chromatograms corresponding to positive and negative signals of SS were obtained, as shown in Fig. S3 and Table S2. We found that the major chemical constituents identified in the SS-G were similar to the traditionally prepared SS decoctions (SS-D) [32]. Therefore, we used the SS-G for the following assays. Then, we compared the HPLC fingerprints of four different batches of SS-G. The similarity indices of these four batches of SS were between 0.943 and 0.982 (Fig. S4), demonstrating that SS was produced consistently with good quality.
The age-dependent Aβ accumulation was found in the brains of APP/PS1 mice, and memory deficits were began to show since the age of 6 months [33]. To explore the potential therapeutic effects of SS, we began the oral administration of SS to the 7-month-old APP/PS1 mice for two months. There were no obvious adverse effects or body weight loss (Table S3). We compared the mice behavioral phenotypes to assess the cognitive function of these transgenic AD mice after drug administration. We assessed the spatial memory of these mice in the MWM. The MWM is a common behavioral task used to determine spatial learning and reference memory deficits. SS treatment did not influence the swimming velocity and distance (Fig. 1, F and G). However, the APP/PS1 mice showed defects in learning compared with the WT mice (Fig. 1A). On day 5, vehicle-treated APP/PS1 mice spent more time than WT littermates to locate the hidden platform (P<0.05, APP/PS1 mice vs. WT mice, Fig. 1A). This difference constantly appeared on day 6, 7, 8 and 10, reflecting deficits in spatial memory of APP/PS1 mice, as reported previously [34]. Interestingly, since day 6, SS-treated APP/PS1 mice showed an improved performance compared with vehicle-treated APP/PS1 mice (P<0.01, Fig. 1A), indicating that the SS treatment alleviated the impairment of spatial learning in APP/PS1 mice. To assess the memory strength of spatial learning, we administered the probe trials on day 4, 7 and 11. The mice in all group showed a random swimming pattern on day 4 (Fig. S5). However, on the day 7 probe trial, compared with the vehicle-treated APP/PS1 mice, the SS-treated group spent more time searching for the platform in the target quadrant (Fig. 1, B and C), took less time to reach the position of the missing platform (P<0.05, Fig. 1D), and increased frequency of crossing within the position of the platform (P<0.05, Fig. 1E). These results suggest that administration of SS alleviates the deficient spatial reference memory of APP/PS1 mice.
Figure 1. SS treatment ameliorates learning and memory impairment in Morris Water Maze and Object recognition test.

(A) MWM test for SS and vehicle-treated APP/PS1 and WT mice. The mean escape latency was given for different test days. (B) The mean percent time in probe trial of MWM on day 7. TQ: Target quadrant; AL: Adjacent left; AR: Adjacent right; OP: Opposite. (C) Representative mice search paths from different groups. (D and E) The latency to target quadrant (D) and the frequency to pass the target position (E) in probe trial are shown. (F and G) The swimming velocity (F) and distance (G) in probe trial are shown. (H and I) Novel object recognition analysis. Preference scores of training phase (H) and Recognition Index of testing phase (I) during a 10-min testing phase are shown, respectively. n = 9–12 for each group. *P<0.05, **P<0.01, ***P<0.001, #P<0.05, ##P<0.01, ###P<0.001.
To further assess the learning and recognition memory processes of these AD mice, we applied the object recognition test (Fig. S1). There were no obvious differences among mice in all groups regarding which object they preferred or the location of the objects as the preference scores were all approximately 50% (Fig. 1H). In the testing phase, the WT mice significantly increased the time exploring the novel object (P<0.001, Fig. 1I). In contrast, the time exploring the novel object of vehicle-treated APP/PS1 mice did not increased obviously. Meanwhile, the SS-treated APP/PS1 mice explored the novel object longer than vehicle-treated APP/PS1 mice (P<0.001) indicating improved memory retention by SS treatment.
SS treatment reduced Aβ levels, Aβ amyloidosis, gliosis and neuron loss in the brains of APP/PS1 mice
APP/PS1 mice begin to develop cerebral amyloidosis at 2 months of age, and the Aβ levels or Aβ deposits in the brain can be detected since 6 months of age. Histology was performed on fixed brain tissues. APP/PS1 mice showed 6E10-positive amyloid plaques (Fig. 2A), which were significantly reduced in the brains of SS-treated APP/PS1 mice. Detailed plaque size distribution analysis revealed a reduction of different size plaques in SS-treated APP/PS1 mice compared with that of control APP/PS1 mice (Fig. 2B). The overall amyloid plaques of SS-treated APP/PS1 mice was reduced by 18.1% compared with that of control APP/PS1 mice (P<0.01, Fig. 2C), indicating that SS treatment prevented the formation of plaques and/or enhanced the clearance of amyloid plaques.
Figure 2. SS treatment alleviates Aβ levels and amyloid plaque burden, reduces gliosis and neuron loss in APP/PS1 mice.

(A–C) Representative half brain sections of WT mice, vehicle or SS-treated APP/PS1 mice stained with antibody against Aβ (6E10) and double staining of GFAP and 6E10 are shown. Scale bar, 1 mm. (B and C) Quantitative analysis of the number of 6E10-positive amyloid plaques (B) and Aβ covered area (C). n = 5 animals per group. (D and E) ELISA of soluble and insoluble Aβ40 and Aβ42 levels in cortical and hippocampal tissues of APP/PS1 mice. n = 6 for each group. (F, I and J) Representative images of WT mice, vehicle- and SS- treated APP/PS1 mice hippocampus and cortex double immunostaining of GFAP and 6E10 (F), CD11b (I) and NeuN (J). Arrows indicate astrocytes surrounding the amyloid plaques. Scale bar, 200 µm. (H) Coincidence of GFAP and Aβ burden in the brains of SS-treated APP/PS1 mice (red; n = 17) and vehicle-treated APP/PS1 mice (black; n = 17; P<0.0001). (G, K and L) The histograms depict the mean GFAP (G), CD11b (K), and NeuN (L) positive area ± S.E.M. in three groups. *P<0.05, **P<0.01, ***P<0.001.
Soluble Aβ oligomers are deleterious, and related to cognitive deficits in AD [35]–[40]. To quantify Aβ levels, we used an ELISA assay. ELISA analysis of SDS-soluble fractions and SDS-insoluble fractions demonstrated that there were high Aβ levels in both fractions from the hippocampi and cortices of APP/PS1 transgenic mice. SS treatment reduced cortical SDS-soluble Aβ40 and Aβ42 levels (30.4% reduction for Aβ42, P<0.05, and 33.7% reduction for Aβ40, P<0.05, Fig. 2D). We also observed a similar reduction of hippocampal SDS-soluble Aβ42 level after SS treatment (17.0% reduction, P<0.01, Fig. 2E). There were no significant differences between the SDS-insoluble formic acid-extractable fractions from vehicle-treated and SS-treated mice (Fig. 2, D and E) suggesting that SS reduces the level of toxic soluble Aβ. These data indicate that SS treatment reduces Aβ levels and the correlated amyloidosis in APP/PS1 mice.
Abnormal neuroinflammation, including accumulation of activated microglia and astrocytes, is a pathological characteristic of the neurodegenerative disease [41]. In the brains of AD patients and APP/PS1 transgenic mice, amyloid plaques are surrounded by activated microglia and reactive astrocytes [41]. Consistent with previous reports [42], we noted that GFAP-positive astrocytes surrounded the amyloid plaques in APP/PS1 mice (Fig. 2, A and F, arrows), and the GFAP-positive staining intensity in APP/PS1 mice was higher than that of WT mice (7.80±0.25 vs. 1.89±0.22 percent area, P<0.001, Fig. 2, F and G). Furthermore, coincident GFAP and Aβ staining was obvious in the brains of vehicle-treated APP/PS1 mice (R2 = 0.72, Fig. 2H). CD11b-positive microglia were also observed (Fig. 2I). We found significant reductions of astrocytes (staining intensity: 6.30% vs. 7.80%, P<0.01; coincidence: 0.68 vs. 0.72, P<0.001, Fig. 2, G and H) and microglia (staining intensity: 1.37% vs. 2.34%, Fig. 2, I and K) in the brains of SS-treated mice, demonstrating that the AD-like pathological gliosis has been significantly moderated by SS treatment in APP/PS1 mice.
As one of the major in vivo neurotoxic properties of Aβ, severe neuronal loss is also observed in APP/PS1 transgenic mice since the age of 10 months [43]. Consistently, in addition to the accumulation of microglia and astrocytes, we also observed significant less neurons (NeuN-positive cells) in the brains of APP/PS1 mice compared with WT mice (Fig. 2, J and L). Interestingly, we found an increase in neurons (NeuN-positive cells) in SS-treated APP/PS1 mice compared with control APP/PS1 mice (P<0.05, Fig. 2L). Together, these results suggest that in APP/PS1 mice, SS treatment retards the Aβ-related pathological gliosis and neuronal loss.
SS treatment improves locomotor functions and prolongs lifespan of AD transgenic Drosophila
Transgenic Drosophila models expressing human Aβ, APP or secretases have been investigated to gain insight into disease mechanisms as well as to elucidate potential therapeutic approaches [44]. AD transgenic Drosophila expressing human Aβ42 show memory deficiencies and premature death [45]. Very recent reports have also shown that the generation of Aβ oligomers through the APP processing by secretases in APP/BACE flies display a very similar pathology as that in AD patients [46]. Because the AD-like pathologies are evident within a few days in these Drosophila models, we performed a rapid in vivo comparison of SS vs. Memantine, the clinically approved medication for treatment of Alzheimer's disease, using transgenic flies carrying Aβ42 or APP/BACE.
We applied the climbing assay to assess the CNS dysfunction of these AD transgenic flies. The climbing assay is a behavioral test based on the negative geotaxis response of Drosophila. Thus, assessing the climbing ability of flies in a fixed time period can reflex their locomotor function.
Consistent with previous reports, flies expressing human APP and BACE or flies expressing Aβ42 showed premature death compared with CS flies (Fig. 3, A, C and E). We also observed significant decreases in the climbing ability of these flies (Fig. 3, B, D and F). Interestingly, when the Aβ42 and APP/BACE transgenic flies were cultured on either SS or Memantine, Aβ-induced premature death was significantly reduced (Fig. 3, C and E). Furthermore, SS or Memantine treatment considerably rescued the climbing ability of these transgenic flies (Fig. 3, D and F). These results indicate that SS treatment improves locomotor functions and reduces premature death of AD transgenic flies, similar to that of Memantine.
Figure 3. SS treatment improves locomotor functions and prolongs lifespan of AD transgenic Drosophila.

(A, C and E) CS, APP/BACE and Aβ transgenic flies were cultured on food containing different concentrations of SS (the triangle symbol stands for concentrations from low to high: 0.2, 0.6 and 2 mg/ml) or Memantine (120 µM). Survival curves for flies treated with either SS or Memantine. The data are presented as mean ± S.E.M. The right panel shows the mean survival days calculated according to the survival curves. (B, D and F) The climbing ability of CS, APP/BACE and Aβ transgenic flies treated with SS or Memantine at day 30 (for CS and APP/BACE flies) and day 20 (for Aβ flies). Values are mean ± S.E.M. Each value represents the mean of three experiments. *P<0.05, **P<0.01, ***P<0.001 vs. Ctrl group. Mem = Memantine.
RP reduces Aβ generation
To explore which granule in SS might reduce Aβ levels, we treated SK-N-SH-APPsw cells with different concentrations of each granule in SS. SK-N-SH-APPsw cells stably express APP protein carrying a Swedish mutant (K595N/M596L) and show elevated Aβ40 and Aβ42 secretion. As shown in Fig. 4, Aβ40 and Aβ42 levels in the culture medium of cells treated with SS were reduced in a dose-dependent manner. Interestingly, the same dose of RP but not that of AT or PRP exhibited similar Aβ40 and Aβ42 reduction effects as SS (Fig. 4, A and B). The viability of cells was not altered by any of these treatments (Fig. 4C). Similar results were observed using HEK293-APPsw cells (Fig. S6).
Figure 4. RP reduces the Aβ generation in SK-N-SH-APPsw cells.
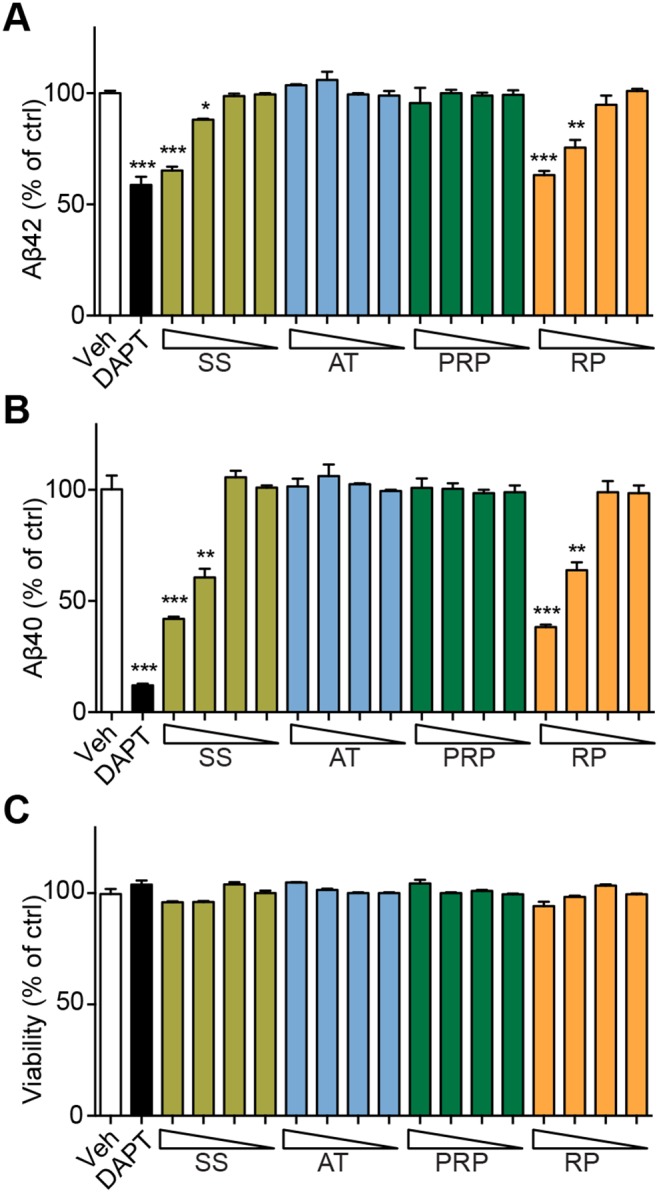
Aβ42 (A) and Aβ40 (B) in SK-N-SH-APPsw cell culture medium and cell viability (C) after treatment with SS, AT, PRP, RP for 24 hours, respectively (the triangle symbol stands for concentrations from high to low: 3000, 1000, 300 and 100 µg/ml for SS; 1000, 300, 100 and 30 µg/ml for AT, PRP and RP). *P<0.05, **P<0.01, ***P<0.001; DAPT: a γ-secretase inhibitor.
Then, we assessed whether RP ameliorates Aβ-related pathology by reducing Aβ generation in vivo. We found that RP showed no obvious beneficial effects in the Aβ transgenic flies expressing Aβ42 directly (Fig. 5A). Interestingly, APP/BACE transgenic flies that were cultured on food containing RP showed considerably reduced Aβ-induced premature death (Fig. 5B). Consistently, the climbing ability of these RP-treated APP/BACE transgenic flies was enhanced than that of vehicle-treated APP/BACE transgenic flies. Aβ levels in APP/BACE and Aβ transgenic flies were monitored by ELISA. As shown in Fig. 5C, APP/BACE flies cultured on food containing SS or RP produced significantly lower levels of Aβ40 and Aβ42 than control. Moreover, Aβ levels in Aβ transgenic flies cultured on food containing SS or RP were not different from that of control Aβ transgenic flies (Fig. 5D). This result suggests that RP may affect human APP proteolysis. To further assess the proteolysis process of APP in APP/BACE flies, full-length APP and APP C-terminal fragments (APP-CTF) in fly heads were monitored using western blots. There was no significant difference in full-length APP and APP-CTF levels in APP/BACE flies (Fig. S7). These results suggest that RP can reduce Aβ generation and thus moderate Aβ-related pathology in vivo.
Figure 5. RP treatment improves locomotor functions, prolongs lifespan and reduces Aβ levels of AD transgenic Drosophila.

CS, Aβ and APP/BACE transgenic flies were cultured on food containing different concentrations of RP (the triangle symbol indicates concentrations from low to high: 0.2, 0.6 and 2 mg/ml) or Memantine (120 µM). (A) Survival curves of flies treated with either RP or Memantine. The data are presented as the mean ± S.E.M. (B) The climbing ability of flies (right panels) was assessed at day 30 for CS and APP/BACE flies and at day 20 for Aβ flies. The values are the mean ± S.E.M. Each value represents the mean of three experiments. (C and D) Aβ and APP/BACE transgenic flies were cultured on SS, AT, PRP or RP (2 mg/ml). Aβ40 and Aβ42 levels in 500 fly heads were measured by ELISA assay. Mem = Memantine. *P<0.05, **P<0.01, ***P<0.001 vs. the control group.
AT and PRP moderate Aβ-induced neurotoxicity in vivo and in vitro
Aβ-induced neuronal apoptosis in brain is a typical feature of Alzheimer's disease [47]. Transgenic flies carrying the toxic human Aβ42 show neuronal dysfunction and premature death and thereby provide a unique system to evaluate potential protective effects against Aβ-induced neurotoxicity in vivo. We found that SS but not RP treatment improved locomotor functions and reduced premature death of Aβ transgenic flies (Fig. 3, E and F and Fig. 5, A and B), suggesting that either AT or PRP has a neuroprotective property. We treated Aβ42 transgenic flies with different concentrations of AT or PRP. As shown in Fig. 6, both AT and PRP prolonged lifespan and improved the climbing activity of Aβ42 flies (Fig. 6, A–D). Both AT and PRP also prolonged lifespan and improved climbing activity of APP/BACE flies but to a less significant extent (Fig. S8).
Figure 6. AT and PRP improves locomotor function and prolongs lifespan of AD transgenic Drosophila.

CS and Aβ transgenic flies were cultured on food containing different concentrations of AT (0.2, 0.6 and 2 mg/ml), PRP (0.2, 0.6 and 2 mg/ml) or Memantine (120 µM). (A, C) Survival curves for flies treated with either AT, PRP or Memantine. (B, D) The climbing ability of flies was assessed. The values are the mean ± S.E.M. Each value represents the mean of three experiments. Mem = Memantine. *P<0.05, **P<0.01, ***P<0.001 vs. the control group.
To investigate whether SS inhibits Aβ aggregation, we performed a Th-T fluorescence assay. Th-T fluorescence signals were gradual increased over time. Both SS and AT inhibited the fluorescence intensity almost completely (Fig. 7A), while PRP showed a less significant effect. SS, AT and PRP inhibited Aβ42 aggregation in a dose-dependent manner (Fig. 7B). To directly assess the neuroprotective effects of AT and PRP, we further monitored Aβ-induced cell death of primary mouse cortical neurons. Aβ42 oligomer preparations consistently provided small oligomers and expected structure as monitored by western blot and AFM (Fig. S9), respectively. Aβ42 oligomers effectively induced the death of cortical neurons within 48 hours as measured by the CellTiter-Glo assay (Fig. 7C) and Tuj1 staining (Fig. 7D). SS significantly prevented neurons from Aβ42 oligomers-induced cell death. Consistent with the in vivo results above, both AT and PRP improved primary neuron survival in the presence of toxic Aβ42 oligomers (Fig. 7, C and D). Aβ42 oligomers-induced cell apoptosis was further monitored using a TUNEL assay. As shown in Fig. 7E, approximately 40% of cells showed severe DNA fragmentation upon Aβ42 treatment. In the presence of SS, AT or PRP, the number of apoptotic cells was significantly reduced (Fig. 7, E and F). Together, these data indicate that both AT and PRP protect neurons from Aβ42-induced toxicity.
Figure 7. AT and PRP inhibit Aβ42 aggregation and exert neuroprotective effects against Aβ42 in primary neurons.

(A) The effects of SS (3000 µg/ml), AT (1000 µg/ml), PRP (1000 µg/ml) or RP (1000 µg/ml) on Aβ42 aggregation, as measured by Th-T fluorescence assay. (B) SS, AT, PRP and RP (3000, 1000, 300, 100 and 30 µg/ml for SS; 1000, 300, 100 and 30 µg/ml for AT, PRP and RP) inhibited Aβ42 aggregation in dose-dependent manner. (C and D) Cell viability of primary cultured neurons pre-treated with SS or AT, PRP for two hours followed by incubation with Aβ42 oligomers (5 µM) for another 48 hours (300, 100, 30 and 10 µg/ml for SS; 100, 30, 10 and 3 µg/ml for AT and PRP). Viable cells were quantified using a CellTiter-Glo assay (A). Tuj1-positive cells were counted and presented (D). *P<0.05 compared with the Aβ42-treated group. (E and F) TUNEL analysis of the primary neurons pre-treated with SS (100 µg/ml), AT (30 µg/ml), or PRP (30 µg/ml) for two hours followed by incubation with Aβ42 oligomers (5 µM) for another 24 hours. The green, TUNEL-positive cells are merged with blue DAPI-positive cells (E). The TUNEL-positive cells and DAPI-positive cells pre-treated with SS or AT, PRP (300, 100, 30 and 10 µg/ml for SS; 100, 30, 10 and 3 µg/ml for AT and PRP) were counted in three independent experiments (F). *P<0.05, **P<0.01, ***P<0.001 vs. the Aβ42-treated group, ##P<0.01 vs. the control group.
Discussion
Our findings in this study demonstrate that SS, the three-herb TCM formula, can ameliorate AD-related pathological symptoms. More interestingly, our results indicate that SS treatment not only ameliorated AD-related symptoms but also exhibited disease-modifying effects such as reduction of Aβ levels and Aβ amyloidosis, retardation of neuronal loss and decrease in astrocytes and microglia. Collectively, our study suggests that it is possible to alleviate symptoms and modify the disease simultaneously, and this ancient formula provides an example of such effects. Further extensive investigation is required to identify the active functional constituents as well as to elucidate the correlate molecular mechanisms, which should finally lead us to a systematic therapeutic strategy against this complicated disease.
It is characteristic of TCM that each formula comprises several herbs with different or distinct functions that work synergistically at multiple targets of a complicated disease. Our studies indicate that three herbs of SS have different therapeutic targets of AD: RT reduced Aβ generation, and AT and PRP protected neuron against Aβ-induced toxicity. RP reduced Aβ40 and Aβ42 generation in SK-N-SH-APPsw cells, showed beneficial effects in flies expressing APP/BACE but did not significantly affect locomotor function or survival in flies expressing Aβ directly. Consistently, RP treatment reduced Aβ40 and Aβ42 levels in APP/BACE transgenic fly heads, but had no significant effects in Aβ transgenic fly heads, indicating that RP reduced Aβ generation and affected the proteolysis of human APP. Furthermore, AT and PRP showed inhibitory effects on Aβ42 aggregation and protected primary neurons against Aβ42 oligomers-induced cell toxicity and apoptosis. In AD transgenic flies, AT and PRP treatment alleviated Aβ-induced premature death and motor neuron dysfunction in both Aβ transgenic flies and APP/BACE flies, suggesting that AT and PRP protect neurons through other mechanisms but may not affect APP cleavage or processing. Unfortunately, the functional active constituents of each herb are unclear in the present study. Obviously, further elucidation of the detailed molecular mechanisms should provide more efficient screening models to identify those functional constituents in each herb. Furthermore, it is of note that SS treatment improved locomotor activity and prolonged lifespan in AD Drosophila models, which makes it possible to quickly identify/verify functional constituents of SS in the near future. On the other hand, SS is one of many TCM formulae documented against memory loss and cognitive impairment. We must evaluate other formulae using current molecular, cellular and animal models as performed in this study, which may eventually lead to a more effective treatment against AD.
TCMs have been orally administered as decoctions, such as this formula for the so-called Smart Soup. Recently, CFDA-approved single-herb granules have been widely used due to their ease of large-scale industry production with stringent manufacturing protocols, better quality control with well-documented chemical fingerprints, and reproducible pharmacokinetic parameters. In our study, we found that the granule preparation (a mixture of three single-herb granules) was as effective as the traditional decoctions (the “soup” preparation), which simplifies future studies of individual herbs or their combination.
Although SS has been prescribed by Chinese medical physicians to patients with aging-related cognitive impairment, a multicenter, double-blind, randomized, placebo-controlled study should be carried out to evaluate the overall efficacies of SS against AD. Furthermore, whether SS should be prescribed alone or combined with other FDA-approved drugs should be assessed in future clinical trials. Moreover, it remains to be determined whether SS or other TCM formulae can be applied for MCI or for preventing neurodegenerative diseases.
Supporting Information
A schematic diagram of novel object recognition.
(TIF)
A sketch of the locomotor assay.
(TIF)
MS total ion current chromatograms of SS (SS-G). Negative-ion (A) and positive-ion (B) modes were selected for TOF/MS analysis. Forty-five compound peaks were tentatively identified on the basis of mass measurements and retention times. Of the 45 peaks of SS, AT accounts for 5 of 45 peaks, PRP accounts for 8 of 45 peaks, and RP accounts for 32 of 45 peaks.
(TIF)
HPLC fingerprints of four batches of SS (from bottom to top: 201102, 201205, 201301 and 201311). The peaks are the characteristic and representative chemical constituents detected in SS. The similarity indices of four batches of samples were between 0.943 and 0.982.
(TIF)
Representative raw data of the MWM search paths of SS- or vehicle-treated APP/PS1 mice or WT littermates.
(TIF)
RP reduces the Aβ generation in HEK293-APPsw cells. Aβ42 (A) and Aβ40 (B) in HEK293-APPsw cell culture medium and cell viability (C) after treatment with SS, AT, PRP, RP for 8 hours, respectively (the triangle symbol indicates concentrations from high to low: 3000, 1000, 300 and 100 µg/ml for SS; 1000, 300, 100 and 30 µg/ml for AT, PRP and RP). *P<0.05, **P<0.01, ***P<0.001; DAPT, a γ-secretase inhibitor.
(TIF)
SS and RP do not alter APP expression in APP/BACE transgenic Drosophila . Western blot of human APP in APP/BACE transgenic flies cultured on SS, AT, PRP or RP (2 mg/ml). Lane 1: HEK293-APPsw cell lysates; Lane 2–6: head lysates of APP/BACE flies. Full-length APP (∼110 kD), APP-CTFs (∼10–12 kD) and Appl-CTFs (∼15 kD) were detected.
(TIF)
AT and PRP improves locomotor functions and prolongs lifespan of APP/BACE transgenic Drosophila . APP/BACE transgenic flies were cultured on food containing different concentrations of AT, PRP (0.2, 0.6, or 2 mg/ml) or Memantine (120 µM). (A and C) Survival curves for flies treated with either AT, PRP or Memantine. (B and D) The climbing ability of flies was assayed. The values are the mean ± S.E.M. Each value represents the mean of three experiments. Mem = Memantine. *P<0.05, **P<0.01, ***P<0.001 vs. the control group.
(TIF)
Representative Western blots and Atomic Force Microscope (AFM) for Aβ42 oligomers. (A) Representative western blots of Aβ42 monomers and oligomers separated by SDS-PAGE using 16% tricine gel and probed with the antibody 6E10 are shown. (B) AFM images of Aβ42 monomers, oligomers and fibrils are shown. The sample was scanned with AFM analysis at 10 µM. Scale bar, 0.5 µm.
(TIF)
Herbal Ingredients in the SS.
(PDF)
Compounds identified in SS by HPLC-TOF/MS.
(PDF)
The detailed information of each mouse in each group.
(PDF)
Acknowledgments
We thank all members of the laboratory for sharing reagents and ideas.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by National Natural Science Foundation of China (31371419), Chinese Academy of Sciences (KSCX2-EW-Q-1-01, KSCX2-YW-R-252), National Science and Technology Support Program (2012BAI10B03), the Ministry of Science and Technology (2011CB910202) and Shanghai Municipal Commission for Science and Technology (13401900600). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hardy J, Selkoe DJ (2002) The amyloid hypothesis of Alzheimer's disease: progress and problems on the road to therapeutics. Science 297: 353–356. [DOI] [PubMed] [Google Scholar]
- 2. Selkoe DJ (2002) Deciphering the genesis and fate of amyloid beta-protein yields novel therapies for Alzheimer disease. The Journal of clinical investigation 110: 1375–1381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Selkoe DJ (1997) Alzheimer's disease: genotypes, phenotypes, and treatments. Science 275: 630–631. [DOI] [PubMed] [Google Scholar]
- 4. Winslow BT, Onysko MK, Stob CM, Hazlewood KA (2011) Treatment of Alzheimer disease. American family physician 83: 1403–1412. [PubMed] [Google Scholar]
- 5. Howes MJ, Houghton PJ (2003) Plants used in Chinese and Indian traditional medicine for improvement of memory and cognitive function. Pharmacology, biochemistry, and behavior 75: 513–527. [DOI] [PubMed] [Google Scholar]
- 6. Tian XY, Liu L (2012) Drug discovery enters a new era with multi-target intervention strategy. Chinese journal of integrative medicine 18: 539–542. [DOI] [PubMed] [Google Scholar]
- 7. Lee B, Choi Y, Kim H, Kim SY, Hahm DH, et al. (2003) Protective effects of methanol extract of Acori graminei rhizoma and Uncariae Ramulus et Uncus on ischemia-induced neuronal death and cognitive impairments in the rat. Life sciences 74: 435–450. [DOI] [PubMed] [Google Scholar]
- 8. Tong XG, Liu JL, Cheng YX (2010) A new pregnane steroid from Poria cum radix pini. Journal of Asian natural products research 12: 419–423. [DOI] [PubMed] [Google Scholar]
- 9. Chen YL, Hsieh CL, Wu PH, Lin JG (2004) Effect of Polygala tenuifolia root on behavioral disorders by lesioning nucleus basalis magnocellularis in rat. Journal of ethnopharmacology 95: 47–55. [DOI] [PubMed] [Google Scholar]
- 10. Park CH, Choi SH, Koo JW, Seo JH, Kim HS, et al. (2002) Novel cognitive improving and neuroprotective activities of Polygala tenuifolia Willdenow extract, BT-11. Journal of neuroscience research 70: 484–492. [DOI] [PubMed] [Google Scholar]
- 11. Lin Z, Gu J, Xiu J, Mi T, Dong J, et al. (2012) Traditional chinese medicine for senile dementia. Evidence-based complementary and alternative medicine: eCAM 2012: 692621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Lee JY, Kim KY, Shin KY, Won BY, Jung HY, et al. (2009) Effects of BT-11 on memory in healthy humans. Neuroscience letters 454: 111–114. [DOI] [PubMed] [Google Scholar]
- 13. Shin KY, Lee JY, Won BY, Jung HY, Chang KA, et al. (2009) BT-11 is effective for enhancing cognitive functions in the elderly humans. Neuroscience letters 465: 157–159. [DOI] [PubMed] [Google Scholar]
- 14. Jankowsky JL, Slunt HH, Ratovitski T, Jenkins NA, Copeland NG, et al. (2001) Co-expression of multiple transgenes in mouse CNS: a comparison of strategies. Biomolecular engineering 17: 157–165. [DOI] [PubMed] [Google Scholar]
- 15. Reiserer RS, Harrison FE, Syverud DC, McDonald MP (2007) Impaired spatial learning in the APPSwe + PSEN1DeltaE9 bigenic mouse model of Alzheimer's disease. Genes, brain, and behavior 6: 54–65. [DOI] [PubMed] [Google Scholar]
- 16. Bero AW, Yan P, Roh JH, Cirrito JR, Stewart FR, et al. (2011) Neuronal activity regulates the regional vulnerability to amyloid-beta deposition. Nature neuroscience 14: 750–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Morris R (1984) Developments of a water-maze procedure for studying spatial learning in the rat. Journal of neuroscience methods 11: 47–60. [DOI] [PubMed] [Google Scholar]
- 18. Alamed J, Wilcock DM, Diamond DM, Gordon MN, Morgan D (2006) Two-day radial-arm water maze learning and memory task; robust resolution of amyloid-related memory deficits in transgenic mice. Nature protocols 1: 1671–1679. [DOI] [PubMed] [Google Scholar]
- 19. Teng L, Zhao J, Wang F, Ma L, Pei G (2010) A GPCR/secretase complex regulates beta- and gamma-secretase specificity for Abeta production and contributes to AD pathogenesis. Cell research 20: 138–153. [DOI] [PubMed] [Google Scholar]
- 20. Bevins RA, Besheer J (2006) Object recognition in rats and mice: a one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nature protocols 1: 1306–1311. [DOI] [PubMed] [Google Scholar]
- 21. Liu X, Zhao X, Zeng X, Bossers K, Swaab DF, et al. (2013) beta-arrestin1 regulates gamma-secretase complex assembly and modulates amyloid-beta pathology. Cell research 23: 351–365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Lazarov O, Robinson J, Tang YP, Hairston IS, Korade-Mirnics Z, et al. (2005) Environmental enrichment reduces Abeta levels and amyloid deposition in transgenic mice. Cell 120: 701–713. [DOI] [PubMed] [Google Scholar]
- 23. Chakraborty R, Vepuri V, Mhatre SD, Paddock BE, Miller S, et al. (2011) Characterization of a Drosophila Alzheimer's disease model: pharmacological rescue of cognitive defects. PloS one 6: e20799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401–415. [DOI] [PubMed] [Google Scholar]
- 25. Zhao XL, Wang WA, Tan JX, Huang JK, Zhang X, et al. (2010) Expression of beta-amyloid induced age-dependent presynaptic and axonal changes in Drosophila. The Journal of neuroscience: the official journal of the Society for Neuroscience 30: 1512–1522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. White KE, Humphrey DM, Hirth F (2010) The dopaminergic system in the aging brain of Drosophila. Frontiers in neuroscience 4: 205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Mhatre SD, Satyasi V, Killen M, Paddock BE, Moir RD, et al. (2014) Synaptic abnormalities in a Drosophila model of Alzheimer's disease. Disease models & mechanisms 7: 373–385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Stine WB Jr, Dahlgren KN, Krafft GA, LaDu MJ (2003) In vitro characterization of conditions for amyloid-beta peptide oligomerization and fibrillogenesis. The Journal of biological chemistry 278: 11612–11622. [DOI] [PubMed] [Google Scholar]
- 29. Lu JH, Ardah MT, Durairajan SS, Liu LF, Xie LX, et al. (2011) Baicalein inhibits formation of alpha-synuclein oligomers within living cells and prevents Abeta peptide fibrillation and oligomerisation. Chembiochem: a European journal of chemical biology 12: 615–624. [DOI] [PubMed] [Google Scholar]
- 30. Cartier AE, Djakovic SN, Salehi A, Wilson SM, Masliah E, et al. (2009) Regulation of synaptic structure by ubiquitin C-terminal hydrolase L1. The Journal of neuroscience: the official journal of the Society for Neuroscience 29: 7857–7868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Cisse M, Halabisky B, Harris J, Devidze N, Dubal DB, et al. (2011) Reversing EphB2 depletion rescues cognitive functions in Alzheimer model. Nature 469: 47–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Ling Y, Li Z, Chen M, Sun Z, Fan M, et al. (2013) Analysis of multiple constituents in Cong-Ming-Tang, a Chinese herbal formula for the treatment of amnesia, by high-performance liquid chromatography with quadrupole time-of-flight mass spectrometry. Phytochemical analysis: PCA 24: 677–688. [DOI] [PubMed] [Google Scholar]
- 33. Jankowsky JL, Fadale DJ, Anderson J, Xu GM, Gonzales V, et al. (2004) Mutant presenilins specifically elevate the levels of the 42 residue beta-amyloid peptide in vivo: evidence for augmentation of a 42-specific gamma secretase. Human molecular genetics 13: 159–170. [DOI] [PubMed] [Google Scholar]
- 34. Cao D, Lu H, Lewis TL, Li L (2007) Intake of sucrose-sweetened water induces insulin resistance and exacerbates memory deficits and amyloidosis in a transgenic mouse model of Alzheimer disease. The Journal of biological chemistry 282: 36275–36282. [DOI] [PubMed] [Google Scholar]
- 35. Lesne S, Koh MT, Kotilinek L, Kayed R, Glabe CG, et al. (2006) A specific amyloid-beta protein assembly in the brain impairs memory. Nature 440: 352–357. [DOI] [PubMed] [Google Scholar]
- 36. Walsh DM, Selkoe DJ (2007) A beta oligomers - a decade of discovery. Journal of neurochemistry 101: 1172–1184. [DOI] [PubMed] [Google Scholar]
- 37. Walsh DM, Klyubin I, Fadeeva JV, Cullen WK, Anwyl R, et al. (2002) Naturally secreted oligomers of amyloid beta protein potently inhibit hippocampal long-term potentiation in vivo. Nature 416: 535–539. [DOI] [PubMed] [Google Scholar]
- 38. Shankar GM, Li S, Mehta TH, Garcia-Munoz A, Shepardson NE, et al. (2008) Amyloid-beta protein dimers isolated directly from Alzheimer's brains impair synaptic plasticity and memory. Nature medicine 14: 837–842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Cheng IH, Scearce-Levie K, Legleiter J, Palop JJ, Gerstein H, et al. (2007) Accelerating amyloid-beta fibrillization reduces oligomer levels and functional deficits in Alzheimer disease mouse models. The Journal of biological chemistry 282: 23818–23828. [DOI] [PubMed] [Google Scholar]
- 40. Lesne S, Kotilinek L, Ashe KH (2008) Plaque-bearing mice with reduced levels of oligomeric amyloid-beta assemblies have intact memory function. Neuroscience 151: 745–749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. McGeer EG, McGeer PL (2003) Inflammatory processes in Alzheimer's disease. Progress in neuro-psychopharmacology & biological psychiatry 27: 741–749. [DOI] [PubMed] [Google Scholar]
- 42.Kraft AW, Hu X, Yoon H, Yan P, Xiao Q, et al.. (2012) Attenuating astrocyte activation accelerates plaque pathogenesis in APP/PS1 mice. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. [DOI] [PMC free article] [PubMed]
- 43. Hamilton A, Holscher C (2012) The effect of ageing on neurogenesis and oxidative stress in the APP(swe)/PS1(deltaE9) mouse model of Alzheimer's disease. Brain research 1449: 83–93. [DOI] [PubMed] [Google Scholar]
- 44. Muqit MM, Feany MB (2002) Modelling neurodegenerative diseases in Drosophila: a fruitful approach? Nature reviews Neuroscience 3: 237–243. [DOI] [PubMed] [Google Scholar]
- 45. Bilen J, Bonini NM (2005) Drosophila as a model for human neurodegenerative disease. Annual review of genetics 39: 153–171. [DOI] [PubMed] [Google Scholar]
- 46. Iijima K, Iijima-Ando K (2008) Drosophila models of Alzheimer's amyloidosis: the challenge of dissecting the complex mechanisms of toxicity of amyloid-beta 42. Journal of Alzheimer's disease: JAD 15: 523–540. [DOI] [PubMed] [Google Scholar]
- 47. Dahlgren KN, Manelli AM, Stine WB Jr, Baker LK, Krafft GA, et al. (2002) Oligomeric and fibrillar species of amyloid-beta peptides differentially affect neuronal viability. The Journal of biological chemistry 277: 32046–32053. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A schematic diagram of novel object recognition.
(TIF)
A sketch of the locomotor assay.
(TIF)
MS total ion current chromatograms of SS (SS-G). Negative-ion (A) and positive-ion (B) modes were selected for TOF/MS analysis. Forty-five compound peaks were tentatively identified on the basis of mass measurements and retention times. Of the 45 peaks of SS, AT accounts for 5 of 45 peaks, PRP accounts for 8 of 45 peaks, and RP accounts for 32 of 45 peaks.
(TIF)
HPLC fingerprints of four batches of SS (from bottom to top: 201102, 201205, 201301 and 201311). The peaks are the characteristic and representative chemical constituents detected in SS. The similarity indices of four batches of samples were between 0.943 and 0.982.
(TIF)
Representative raw data of the MWM search paths of SS- or vehicle-treated APP/PS1 mice or WT littermates.
(TIF)
RP reduces the Aβ generation in HEK293-APPsw cells. Aβ42 (A) and Aβ40 (B) in HEK293-APPsw cell culture medium and cell viability (C) after treatment with SS, AT, PRP, RP for 8 hours, respectively (the triangle symbol indicates concentrations from high to low: 3000, 1000, 300 and 100 µg/ml for SS; 1000, 300, 100 and 30 µg/ml for AT, PRP and RP). *P<0.05, **P<0.01, ***P<0.001; DAPT, a γ-secretase inhibitor.
(TIF)
SS and RP do not alter APP expression in APP/BACE transgenic Drosophila . Western blot of human APP in APP/BACE transgenic flies cultured on SS, AT, PRP or RP (2 mg/ml). Lane 1: HEK293-APPsw cell lysates; Lane 2–6: head lysates of APP/BACE flies. Full-length APP (∼110 kD), APP-CTFs (∼10–12 kD) and Appl-CTFs (∼15 kD) were detected.
(TIF)
AT and PRP improves locomotor functions and prolongs lifespan of APP/BACE transgenic Drosophila . APP/BACE transgenic flies were cultured on food containing different concentrations of AT, PRP (0.2, 0.6, or 2 mg/ml) or Memantine (120 µM). (A and C) Survival curves for flies treated with either AT, PRP or Memantine. (B and D) The climbing ability of flies was assayed. The values are the mean ± S.E.M. Each value represents the mean of three experiments. Mem = Memantine. *P<0.05, **P<0.01, ***P<0.001 vs. the control group.
(TIF)
Representative Western blots and Atomic Force Microscope (AFM) for Aβ42 oligomers. (A) Representative western blots of Aβ42 monomers and oligomers separated by SDS-PAGE using 16% tricine gel and probed with the antibody 6E10 are shown. (B) AFM images of Aβ42 monomers, oligomers and fibrils are shown. The sample was scanned with AFM analysis at 10 µM. Scale bar, 0.5 µm.
(TIF)
Herbal Ingredients in the SS.
(PDF)
Compounds identified in SS by HPLC-TOF/MS.
(PDF)
The detailed information of each mouse in each group.
(PDF)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.