

Abstract
The development of methodology to identify specific cell populations and circuits within the basal ganglia is rapidly transforming our ability to understand the function of this complex circuit. This mini-symposium highlights recent advances in delineating the organization and function of neural circuits in the external segment of the globus pallidus (GPe). Although long considered a homogeneous structure in the motor-suppressing “indirect-pathway,” the GPe consists of a number of distinct cell types and anatomical subdomains that contribute differentially to both motor and nonmotor features of behavior. Here, we integrate recent studies using techniques, such as viral tracing, transgenic mice, electrophysiology, and behavioral approaches, to create a revised framework for understanding how the GPe relates to behavior in both health and disease.
Introduction
The external segment of the globus pallidus (GPe) is centrally placed in the basal ganglia circuit and is classically considered a component of the motor-suppressing “indirect pathway” (Albin et al., 1989; DeLong, 1990; Smith et al., 1998; Obeso et al., 2008). The major inputs to the GPe are as follows: (1) GABAergic inputs from D2-expressing spiny projection neurons (D2 SPNs) in the striatum (Gerfen et al., 1990; Kita, 2007); and (2) glutamatergic inputs from the subthalamic nucleus (STN) (Smith et al., 1998; Kita, 2007).
The GPe is thought to contribute prominently to basal ganglia dysfunction in Parkinson's disease (PD). In animal models of PD, the firing of GPe neurons is typically reduced relative to control (Filion and Tremblay, 1991; Filion et al., 1991; Chan et al., 2011), consistent with the classical model of basal ganglia function that the indirect pathway is overactive in PD (Albin et al., 1989; DeLong, 1990). Additionally, the normally uncorrelated activity of GPe neurons becomes synchronized, a change thought to contribute to pathological oscillations that disrupt basal ganglia function and contribute to motor impairments (Bevan et al., 2002b; Brown, 2003, 2007; Mallet et al., 2008a, 2012).
Despite its typecasting as a homogeneous relay, connecting the striatum and STN in the indirect pathway, the GPe comprises a rich neural circuitry of diverse cell types that shape both motor and nonmotor features of behavior. Under normal conditions at rest, GPe neurons fire tonically, at rates of 10–80 Hz in vivo; they respond heterogeneously during movement, with complex temporal patterns and low correlation (DeLong et al., 1985; Jaeger et al., 1995; Nini et al., 1995; Turner and Anderson, 1997). Since the earliest investigations of GPe activity in behaving animals, it was observed that neurons fall into discrete categories, suggesting the existence of different neuronal populations. In his 1971 paper, DeLong (1971) described two subpopulations of neurons with strikingly different firing patterns: high-frequency pausers (∼80%) and low-frequency bursters (∼20%). This classification was also recently described in rats (Benhamou et al., 2012). Indeed, a growing body of anatomical, electrophysiological, and molecular literature demonstrates that the GPe is composed of a heterogeneous population of neurons that give rise to different circuits (Nambu and Llinaś, 1994; Hoover and Marshall, 1999; Bolam et al., 2000; Kita and Kita, 2001; Flandin et al., 2010; Nóbrega-Pereira et al., 2010; Benhamou et al., 2012; Mastro et al., 2014).
In this mini-symposium, we will first discuss anatomical strategies being used in primates to map the cortical origins of behaviorally distinct circuits in the GPe. Next, we will focus on one circuit in particular, the pallidostriatal pathway, to highlight how molecular, cellular, and behavioral work has come together to demonstrate a novel behavioral role for a molecularly defined cell type in the GPe, arkypallidal GPe neurons (Arky-GPe). Next, we will discuss the use of a recently established, genetic strategy to identify two additional classes of GPe neurons: Lhx6- and PV-GPe neurons. Finally, we will turn our discussion of the GPe to its role in disease, particularly PD. We will explore in-depth, cellular mechanisms of communication between the GPe and STN that may shed light on why circuitry within the GPe is so prone to disruption under conditions of low dopamine. We will also extend our discussion of cellular mechanisms of pathophysiology in the GPe beyond neurons to consider the role of glia in GPe dysfunction in PD.
Thus, by bringing together findings from cellular, anatomical, molecular, and behavioral neuroscience, this mini-symposium seeks to formulate a new, unified model of the GPe as a highly organized, central processing station within the basal ganglia that influences many aspects of motor and nonmotor behavior in health and disease.
Anatomical circuitry of distinct GPe functional territories
Early evidence for functionally diverse circuits in the GPe came from a study in primates, showing that focal, drug-induced activation of discrete regions of the nucleus produced distinct behavioral outputs (Grabli et al., 2004). In a companion anatomical study, François et al. (2004) identified separate striatal inputs, and pallidal and nigral outputs, associated with each behaviorally distinct territory of the GPe. Bicuculline injections into the posterior, ventrolateral “sensorimotor” territory of the GPe produced abnormal movements; into the middle “associative” territory produced hyperactivity and/or attention deficit; and into the anterior, ventral “limbic” territory produced stereotypies.
One hypothesis, based on a proposal put forth by Alexander et al. (1986), is that these functionally distinct zones in the GPe are part of larger, corticobasal ganglia loops, whose anatomical and functional segregation is conserved throughout the extent of the basal ganglia circuit. If this hypothesis were true, then behaviorally distinct territories of the GPe should be associated with different cortical areas. Specifically, motor territories of the GPe should be associated with different cortical areas than associative or limbic GPe territories.
To test this hypothesis, a retrograde, trans-synapticrabies virus was used to label the cortical areas associated with each of the three functionally defined territories of the GPe. Bicuculline injections in the three GPe territories yielded behavioral changes similar to those reported by Grabli et al. (2004). A rabies injection placed in limbic GPe, where bicuculline injections produced stereotypies, including excessive fur grooming, hair pulling, and nail biting, labeled cortical neurons in regions, including rostromedial prefrontal cortex and orbitofrontal cortex. A rabies injection placed in associative GPe, where bicuculline injections produced hyperactivity, labeled cortical neurons in regions, including rostral premotor and caudal prefrontal areas of cortex. A rabies injection into sensorimotor GPe, where bicuculline injections produced abnormal limb movements, labeled cortical neurons in regions, including primary motor cortex and premotor cortex. Interestingly, although abnormal movements, hyperactivity, and stereotypies were all expressed as observable changes in movement patterns, only the rabies injection into sensorimotor GPe produced dense cortical labeling in cortical motor regions. This finding may reflect the fact that, unlike the abnormal limb movements seen in bicuculline-induced disruption of sensorimotor GPe, hyperactivity and stereotypies are normal movements that are executed with abnormal frequency and intensity. These perturbations are not simply motor deficits but reflect disruption of motivational or cognitive aspects of behavior.
Neuronal and functional studies of the pallidostriatal pathway
These studies in primates provide insights into the anatomical organization of functionally distinct circuits, preserved in the GPe. Like primate basal ganglia, rodent basal ganglia can also be grossly divided into sensorimotor, associative, and limbic territories based on patterns of cortical afferents. Sensorimotor striatum is located dorsolaterally within the caudoputamen, associative striatum is located anteriomedial to sensorimotor territory, and limbic striatum (better known as nucleus accumbens) is located ventral to sensorimotor and associative regions (Groenewegen et al., 1990; Voorn et al., 2004). Data on rodent striatopallidal projections are more limited but suggest a rough tripartite organization in which medial striatum projects to medial GPe, lateral striatum projects to lateral GPe, and ventral striatum projects to the ventral pallidum. (Mogenson et al., 1983; Haber et al., 1985; Groenewegen et al., 1990).
Studies in rodents are also providing insights into the organization of GPe circuits at the neuronal level. Developmental studies have established that most GPe neurons (>70%) are derived from the Nkx2-1 lineage of the medial ganglionic eminence, whereas almost all of the remainder (∼25%) originate from a distinct Pax6 lineage in the lateral ganglionic eminence (Flandin et al., 2010; Nóbrega-Pereira et al., 2010). Although the molecular phenotype of these developmentally distinct cell types remains to be established in older animals, Pax6-derived neurons might correspond with Arky-GPe neurons, a subset of GPe neurons that make strong projections, exclusively back to the striatum (Fig. 1). They form a remarkably dense (sometimes >10,000 boutons) GABAergic innervation of all striatal cell types (Mallet et al., 2012). This is in contrast to the pallidostriatal projection of a subset of prototypical GPe neurons (Bevan et al., 1998; Hoover and Marshall, 1999, 2002; Sato et al., 2000; Kita and Kita, 2001; Kita, 2007) that selectively innervate striatal GABAergic interneurons (Bevan et al., 1998).
Figure 1.
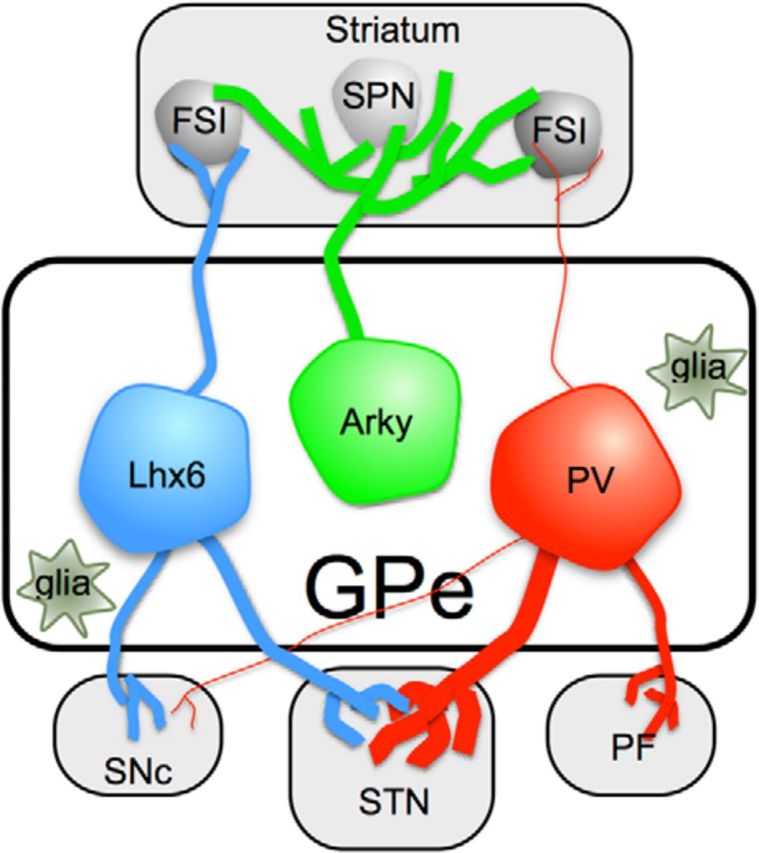
GPe cell types and their projection patterns. Arky-GPe neurons make up ∼25% of GPe neurons, project exclusively to striatum, and target both GABAergic interneurons, including fast-spiking interneurons (FSI) and SPNs (Mallet et al., 2012). Lhx6- and PV-GPe neurons, which make up 34% and 30%–40% of GPe neurons, respectively (Mastro et al., 2014), project to many of the same brain areas, but Lhx6-GPe neurons project more strongly to the striatum and substantia nigra pars compacta, and PV-GPe neurons project more strongly to the STN. PV-GPe neurons project uniquely to the parafascicular thalamic nucleus (PF), which is not innervated by Lhx6-GPe neurons. Striatal projections of both Lhx6- and PV-GPe neurons are selective for striatal GABAergic interneurons (Bevan et al., 1998; Mastro et al., 2014). Figure also illustrates the presence of glial cells in the GPe, which outnumber neurons.
Arky-GPe neurons make up ∼25% of GPe neurons in rats and specifically express preproenkephalin, whereas other GPe neurons do not (Mallet et al., 2012). In addition to being molecularly distinct from other GPe neurons, Arky-GPe neurons exhibit distinctive patterns of activity. In dopamine-depleted animals, most GPe neurons tend to fire out-of-phase with cortical cells during slow-wave and beta oscillations, but Arky-GPe neurons fire in-phase (Mallet et al., 2008a, 2012). Arky-GPe neurons may also map directly onto the firing pattern dichotomy seen in behaving animals, where ∼80% of GPe neurons are high-frequency pausers and ∼20% are low-frequency bursters (DeLong, 1971; Benhamou et al., 2012). During slow-wave sleep, most GPe neurons usually have high and regular firing rates that remain high during slow-wave sleep, whereas Arky-GPe neurons have lower and very irregular firing rates that are further suppressed during sleep.
This distinctive electrophysiological signature of Arky-GPe neurons facilitates their identification in freely moving rats performing behavioral tasks (Schmidt et al., 2014). One role of basal ganglia in motor control is the selection and initiation of actions, but another is the suppression or cancellation of inappropriate behaviors. Single-unit electrophysiology from multiple basal ganglia locations during cued choice tasks allows these various mechanisms to be dissected with high temporal precision (Gage et al., 2010; Leventhal et al., 2012). In particular, the stop-signal (action cancellation) task has been used to show that behavioral responses to “Go” and “Stop” cues involve a race between information processing in distinct BG pathways (Schmidt et al., 2013). The reaction time to respond to a Go cue reflects the relatively slow evolution of neural processing within the striatum (Leventhal et al., 2014). The striatum provides direct-pathway GABAergic inputs to the subtantia nigra pars reticulata (SNr), and the resulting reduction in SNr activity helps release actions (Hikosaka and Wurtz, 1983). Stop cue processing initially involves much faster signaling through STN (∼15 ms latencies), providing glutamatergic inputs to the same SNr neurons; if this occurs early enough, it can oppose action initiation (Schmidt et al., 2013). However, the Stop cue only causes a transient increase in STN-SNr firing, which is likely sufficient to pause, but not completely cancel, an action-in-preparation. Instead, successful, complete cancellation seems to additionally involve suppression of the Go process within striatum.
The GPe appears to provide this second, complementary mechanism for action cancellation. GPe neurons respond more slowly to the Stop cue (∼60–100 ms), but selectively when stopping is successful. This stop response is much more prominent in presumed Arky-GPe than other GPe neurons and matches the time course of striatal suppression on correct Stop trials. These exciting new findings directly implicate the Arky-GPe pallidostriatal pathway in behavioral inhibition, a critical set of mechanisms for ensuring that behavior is both flexible and well controlled.
Genetically defined cell types in GPe
If Arky-GPe neurons make up ∼25% of GPe neurons, what about the remaining 75%? Do the remaining GPe neurons correspond to a single population, or can they be further broken down with the aid of additional classification schemes? A recent study suggests that non–Arky-GPe neurons consist of at least two genetically distinct cell types, labeled in Lhx6 and PV transgenic mice, respectively (Mastro et al., 2014) (Fig. 1). The striatal projections of Lhx6- and PV-GPe neurons are selective for GABAergic interneurons (Mastro et al., 2014), suggesting that they are distinct from Arky-GPe neurons.
Lhx6 and PV-GPe neurons represent largely nonoverlapping populations in the GPe and exhibit opposite, graded distribution throughout the nucleus. Cell counts estimate that one-third of all GPe neurons are labeled in Lhx6 transgenic mice and one-third are labeled in PV transgenic mice. Lhx6-GPe neurons are more concentrated in the medial region of the GPe, and PV-GPe neurons are more concentrated in the lateral region. Based on patterns of calbinin distribution throughout rodent basal ganglia (Hontanilla et al., 1994, 1998; Kita, 2007) and other anatomical studies (Mogenson et al., 1983; Haber et al., 1985; Groenewegen et al., 1990), it has been hypothesized that the medial GPe corresponds to an associative territory, whereas the lateral GPe corresponds to a sensorimotor territory.
Both Lhx6- and PV-GPe neurons project to a number of brain regions, both within and outside the basal ganglia (Mastro et al., 2014). The main anatomical projections distinguishing these cell populations are as follows: stronger Lhx6-GPe projections to the substantia nigra pars compacta and striatum; and stronger PV-GPe projections to the STN and parafascicular nucleus of the thalamus (Fig. 1). Intriguingly, PV-GPe projections to the STN targeted the entirety of the nucleus, but projections of Lhx6-GPe neurons avoided the central territory, confirming topographical organization of GPe circuits in rodents (Mastro et al., 2014). It will be important to test whether this additional classification schemes also apply to the non–Arky-GPe neurons of rats, where the proportion of PV-expressing neurons has been reported to be closer to 50%–60% (Kita, 2007).
In addition to their anatomical distinctions, Lhx6- and PV-GPe neurons may be functionally distinct as well. First, recordings of their intrinsic firing properties in acute slices revealed that PV-GPe neurons have narrower action potentials, higher maximum firing rates, and higher baseline firing rates than Lhx6-GPe neurons (Mastro et al., 2014). Second, optogenetic activation of Lhx6- and PV-GPe neurons produces different effects on behavior. Interestingly, the effects of activating these cell types resemble the behavioral effects seen in primates following disruption in the associative and limbic territories of the GPe, respectively, suggesting conservation of circuit function across species.
Synaptic mechanisms of GPe dysfunction in PD
The GPe through its connections with the STN has been consistently implicated in the onset and maintenance of motor dysfunction in PD (Plenz and Kital, 1999; Bevan et al., 2002b; Burkhardt et al., 2007; Kita, 2007; Vitek et al., 2012). The GPe is the principal source of GABAergic inhibition in the STN (Smith et al., 1998) and acts primarily through postsynaptic GABAA receptors (Hallworth and Bevan, 2005). The equilibrium potential of GABAA receptor current in STN neurons is ∼−80 mV (Bevan et al., 2002a), which reflects active extrusion of Cl−, presumably by the K+/Cl− cotransporter KCC2, which STN neurons express in abundance (Kanaka et al., 2001). The axonal arborization of each GPe neuron traverses a large extent of the STN but is so sparse that it terminates on just a small fraction of neurons (Baufreton et al., 2009). That said, unitary GPe-STN synaptic connections are powerful (∼10–15 nS) because they are mediated by multiple synaptic contacts (Baufreton et al., 2009). Because each STN neuron receives input from ∼50–60 GPe neurons, the maximum inhibitory conductance generated by the GPe is therefore huge (Baufreton et al., 2009). Together, these properties suggest that the GPe exerts a powerful inhibitory influence on the STN. Indeed, GPe-STN transmission limits both the intrinsic activity of STN neurons (Hallworth and Bevan, 2005; Atherton et al., 2013) and shunts synaptic excitation ex vivo (Atherton et al., 2010). Furthermore, in vivo cortical excitation of the STN is curtailed by feedback inhibition from the GPe and inhibition of GPe neurons by D2 SPNs mediates disinhibition of the STN (Fujimoto and Kita, 1993; Maurice et al., 1998). So why do the GPe and STN rarely show correlated activity under normal conditions (Urbain et al., 2000; Magill et al., 2001; Mallet et al., 2008b)? This paradox is likely due to several factors. First, the selective nature of GPe-STN inputs reduces the probability of detecting connected neurons (Baufreton et al., 2009). Second, GPe-STN connections exhibit strong activity-dependent depression, which reduces the impact and thus detectability of unitary connections (Atherton et al., 2010). Third, STN neurons express ion channels, such as HCN, Nav1, and Cav1 and Cav3 channels, which complicate the nature of GPe-STN patterning (Otsuka et al., 2001; Hallworth et al., 2003; Baufreton et al., 2009; Atherton et al., 2010).
In PD and its experimental models, the GPe and STN are abnormally hypoactive and hyperactive, respectively (Galvan and Wichmann, 2008), consistent with an inhibitory action of the GPe on the STN. Furthermore, GPe-STN and STN neurons exhibit anticorrelated firing both during cortical slow-wave activity and activated cortical states in which abnormally persistent and widespread beta band activity is manifest (Hammond et al., 2007; Mallet et al., 2008b; Shimamoto et al., 2013). The altered firing rates of GPe and STN neurons are likely due to hyperactivity of D2 SPNs, which leads to excessive inhibition of GPe-STN neurons and disinhibition of the STN (Gerfen and Surmeier, 2011). The cause of pathologically correlated GPe-STN activity is less clear. Loss of dopamine is associated with profound alterations in the strength of connections in the indirect pathway. Thus, GABAergic connections between fast spiking interneurons and D2 SPNs (Gittis et al., 2011) and between GPe and STN neurons (Fan et al., 2012) strengthen profoundly. Furthermore, the intrinsic activity of GPe and STN neurons, which decorrelates GPe and STN activity by rendering synaptic integration phase-dependent (Wilson, 2013), is diminished following loss of dopamine (Zhu et al., 2002; Chan et al., 2011). Together, these changes may promote synchronous activity in the indirect pathway (Moran et al., 2011; Tachibana et al., 2011; Wilson, 2013). Indeed, in acute lesion models of PD, in which dopamine neurons rapidly degenerate, it takes several further days to weeks for pathological activity to develop (Mallet et al., 2008a; Degos et al., 2009), implying that synaptic, cellular, and network-level plasticity triggered by the loss of dopamine all participate in circuit dysfunction.
Role of glia in GPe dysfunction in PD
Neurons are not the only cell type in the GPe that may undergo alterations in disease. The GPe is known to harbor a rich number of glia, which indeed are estimated to vastly outnumber neurons (Lange et al., 1976) (Fig. 1). There are three main classes of glia in the brain: oligodendrocytes, microglia, and astrocytes. Although no published work available so far gives estimates of the density of different glia classes within the GPe, an enrichment of astrocytes in the GPe is demonstrated by the high density of nominal astrocytic molecules compared with neighboring brain regions (Dervan et al., 2004). Astrocytes are the most numerous cell class in the mammalian brain (Halassa et al., 2007; Giaume et al., 2010), and their relative abundance in GPe implies an important role in regulating GPe function.
Although we have begun to appreciate the role of astrocytes in a few brain areas (Halassa et al., 2007; Araque et al., 2014), the biological importance and disease relevance of astrocytes in the GPe and other basal ganglia nuclei are largely unexplored (Maragakis and Rothstein, 2006; Sofroniew and Vinters, 2010; Tong et al., 2014). Ultrastructural studies suggest that GPe astrocytic processes are strategically positioned to control striatopallidal transmission (Galvan et al., 2010). As hyperactive striatopallidal signaling is observed in PD and is relevant to disease symptomatology (Kravitz et al., 2010), it is intriguing to speculate that GPe astrocytes play a crucial role in gating striatopallidal GABA release and that this regulatory process is disrupted in the absence of dopamine, as seen in PD. Indeed, astrocytes express a myriad of surface receptors (e.g., dopamine receptors), thus allowing them to be regulated by local and long-range modulatory signals (Theodosis et al., 2008; Perea et al., 2009; Nedergaard and Verkhratsky, 2012; Araque et al., 2014). Astrocytes in turn interface with neurons through the transport and release of neuroactive substrates that regulate synaptic activity (Halassa et al., 2007; Perea et al., 2009). The idea that astrocytes negatively regulate striatopallidal transmission via a presynaptic inhibitory mechanism, and that dysregulation of this mechanism may lead to overactive striatopallidal transmission in PD, is being tested in a novel genetic model of the disease. Additionally, efforts are underway to identify the neuroactive substrates involved in the astrocytic regulation of striatopallidal transmission.
Given the importance of glia in proper neuronal function, it is perhaps not surprising that impaired astrocytic function is a common feature of neurodegenerative diseases (Maragakis and Rothstein, 2006; Halassa et al., 2007; Sofroniew and Vinters, 2010). Investigating the alterations of glia in the GPe and other basal ganglia nuclei may therefore identify new targets and strategies for treating otherwise, incurable neurodegenerative diseases, such as Huntington's disease and PD (Chan and Surmeier, 2014).
In conclusion, this mini-symposium is designed to highlight recent discoveries about the rich diversity of neural circuits in the GPe and their role in behavior and disease. In contrast to its usual depiction as a homogeneous nucleus that simply relays information to downstream basal ganglia structures, the GPe contains a rich diversity of cell types and projections. Insights into the richness of GPe organization are leading us toward a revised framework for understanding how neural activity in the GPe relates to behavior. Studies at the cellular level reveal important features of the organization and synaptic dynamics of GPe circuits that make them susceptible to dysfunction in PD, and suggest a previously underappreciated role for glia in disease pathophysiology. Finally, data from this mini-symposium demonstrate functionally conserved circuits in the GPe of rodents and primates that produce behavioral patterns seen in neuropsychiatric disorders, such as obsessive-compulsive disorder and attention-deficit hyperactivity disorder, demonstrating that the GPe is far more than a simple motor-control nucleus.
Footnotes
This work was supported by National Institutes of Health Grants R37NS041280, P50NS047085, R01MH101697, R01NS078435, R21DA032259, R21MH093888, R00NS076524, R01MH56661, P30NS076405, P40RR018604/P40ODO10996, F32MH080569, TS32NS07391, R01NS069777, R01NS069777-S1, and NS047085, Parkinson's Disease Foundation Grant PDF-IRG-1101, CHDI Grant A-5071, Centre National de la Recherche Scientifique and the Minister de l'Education Nationale de la Recherche et Technologique, BrainLinks-BrainTools Cluster of Excellence funded by the German Research Foundation (Deutsche Forschungsgemeinschaft Grant EXC 1086), National Alliance for Research on Schizophrenia and Depression, and Veterans Affairs Medical Research Service. Work reported by M.M.M. was performed in the laboratory of Dr. Peter L. Strick.
The authors declare no competing financial interests.
References
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci. 1989;12:366–375. doi: 10.1016/0166-2236(89)90074-X. [DOI] [PubMed] [Google Scholar]
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. doi: 10.1146/annurev.ne.09.030186.002041. [DOI] [PubMed] [Google Scholar]
- Araque A, Carmignoto G, Haydon PG, Oliet SH, Robitaille R, Volterra A. Gliotransmitters travel in time and space. Neuron. 2014;81:728–739. doi: 10.1016/j.neuron.2014.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atherton JF, Kitano K, Baufreton J, Fan K, Wokosin D, Tkatch T, Shigemoto R, Surmeier DJ, Bevan MD. Selective participation of somatodendritic HCN channels in inhibitory but not excitatory synaptic integration in neurons of the subthalamic nucleus. J Neurosci. 2010;30:16025–16040. doi: 10.1523/JNEUROSCI.3898-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atherton JF, Menard A, Urbain N, Bevan MD. Short-term depression of external globus pallidus-subthalamic nucleus synaptic transmission and implications for patterning subthalamic activity. J Neurosci. 2013;33:7130–7144. doi: 10.1523/JNEUROSCI.3576-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baufreton J, Kirkham E, Atherton JF, Menard A, Magill PJ, Bolam JP, Bevan MD. Sparse but selective and potent synaptic transmission from the globus pallidus to the subthalamic nucleus. J Neurophysiol. 2009;102:532–545. doi: 10.1152/jn.00305.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benhamou L, Bronfeld M, Bar-Gad I, Cohen D. Globus Pallidus external segment neuron classification in freely moving rats: a comparison to primates. PLoS One. 2012;7:e45421. doi: 10.1371/journal.pone.0045421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevan MD, Booth PA, Eaton SA, Bolam JP. Selective innervation of neostriatal interneurons by a subclass of neuron in the globus pallidus of the rat. J Neurosci. 1998;18:9438–9452. doi: 10.1523/JNEUROSCI.18-22-09438.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevan MD, Magill PJ, Hallworth NE, Bolam JP, Wilson CJ. Regulation of the timing and pattern of action potential generation in rat subthalamic neurons in vitro by GABA-A IPSPs. J Neurophysiol. 2002a;87:1348–1362. doi: 10.1152/jn.00582.2001. [DOI] [PubMed] [Google Scholar]
- Bevan MD, Magill PJ, Terman D, Bolam JP, Wilson CJ. Move to the rhythm: oscillations in the subthalamic nucleus-external globus pallidus network. Trends Neurosci. 2002b;25:525–531. doi: 10.1016/S0166-2236(02)02235-X. [DOI] [PubMed] [Google Scholar]
- Bolam JP, Hanley JJ, Booth PA, Bevan MD. Synaptic organisation of the basal ganglia. J Anat. 2000;196:527–542. doi: 10.1046/j.1469-7580.2000.19640527.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown P. Oscillatory nature of human basal ganglia activity: relationship to the pathophysiology of Parkinson's disease. Mov Disord. 2003;18:357–363. doi: 10.1002/mds.10358. [DOI] [PubMed] [Google Scholar]
- Brown P. Abnormal oscillatory synchronisation in the motor system leads to impaired movement. Curr Opin Neurobiol. 2007;17:656–664. doi: 10.1016/j.conb.2007.12.001. [DOI] [PubMed] [Google Scholar]
- Burkhardt JM, Constantinidis C, Anstrom KK, Roberts DC, Woodward DJ. Synchronous oscillations and phase reorganization in the basal ganglia during akinesia induced by high-dose haloperidol. Eur J Neurosci. 2007;26:1912–1924. doi: 10.1111/j.1460-9568.2007.05813.x. [DOI] [PubMed] [Google Scholar]
- Chan CS, Surmeier DJ. Astrocytes go awry in Huntington's disease. Nat Neurosci. 2014;17:641–642. doi: 10.1038/nn.3705. [DOI] [PubMed] [Google Scholar]
- Chan CS, Glajch KE, Gertler TS, Guzman JN, Mercer JN, Lewis AS, Goldberg AB, Tkatch T, Shigemoto R, Fleming SM, Chetkovich DM, Osten P, Kita H, Surmeier DJ. HCN channelopathy in external globus pallidus neurons in models of Parkinson's disease. Nat Neurosci. 2011;14:85–92. doi: 10.1038/nn.2692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Degos B, Deniau JM, Chavez M, Maurice N. Chronic but not acute dopaminergic transmission interruption promotes a progressive increase in cortical beta frequency synchronization: relationships to vigilance state and akinesia. Cereb Cortex. 2009;19:1616–1630. doi: 10.1093/cercor/bhn199. [DOI] [PubMed] [Google Scholar]
- DeLong MR. Activity of pallidal neurons during movement. J Neurophysiol. 1971;34:414–427. doi: 10.1152/jn.1971.34.3.414. [DOI] [PubMed] [Google Scholar]
- DeLong MR. Primate models of movement disorders of basal ganglia origin. Trends Neurosci. 1990;13:281–285. doi: 10.1016/0166-2236(90)90110-V. [DOI] [PubMed] [Google Scholar]
- DeLong MR, Crutcher MD, Georgopoulos AP. Primate globus pallidus and subthalamic nucleus: functional organization. J Neurophysiol. 1985;53:530–543. doi: 10.1152/jn.1985.53.2.530. [DOI] [PubMed] [Google Scholar]
- Dervan AG, Meshul CK, Beales M, McBean GJ, Moore C, Totterdell S, Snyder AK, Meredith GE. Astroglial plasticity and glutamate function in a chronic mouse model of Parkinson's disease. Exp Neurol. 2004;190:145–156. doi: 10.1016/j.expneurol.2004.07.004. [DOI] [PubMed] [Google Scholar]
- Fan KY, Baufreton J, Surmeier DJ, Chan CS, Bevan MD. Proliferation of external globus pallidus-subthalamic nucleus synapses following degeneration of midbrain dopamine neurons. J Neurosci. 2012;32:13718–13728. doi: 10.1523/JNEUROSCI.5750-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Filion M, Tremblay L. Abnormal spontaneous activity of globus pallidus neurons in monkeys with MPTP-induced parkinsonism. Brain Res. 1991;547:142–151. doi: 10.1016/0006-8993(91)90585-J. [DOI] [PubMed] [Google Scholar]
- Filion M, Tremblay L, Bédard PJ. Effects of dopamine agonists on the spontaneous activity of globus pallidus neurons in monkeys with MPTP-induced parkinsonism. Brain Res. 1991;547:152–161. doi: 10.1016/0006-8993(91)90586-K. [DOI] [PubMed] [Google Scholar]
- Flandin P, Kimura S, Rubenstein JL. The progenitor zone of the ventral medial ganglionic eminence requires Nkx2-1 to generate most of the globus pallidus but few neocortical interneurons. J Neurosci. 2010;30:2812–2823. doi: 10.1523/JNEUROSCI.4228-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- François C, Grabli D, McCairn K, Jan C, Karachi C, Hirsch EC, Féger J, Tremblay L. Behavioural disorders induced by external globus pallidus dysfunction in primates: II. Anatomical study. Brain. 2004;127:2055–2070. doi: 10.1093/brain/awh239. [DOI] [PubMed] [Google Scholar]
- Fujimoto K, Kita H. Response characteristics of subthalamic neurons to the stimulation of the sensorimotor cortex in the rat. Brain Res. 1993;609:185–192. doi: 10.1016/0006-8993(93)90872-K. [DOI] [PubMed] [Google Scholar]
- Gage GJ, Stoetzner CR, Wiltschko AB, Berke JD. Selective activation of striatal fast-spiking interneurons during choice execution. Neuron. 2010;67:466–479. doi: 10.1016/j.neuron.2010.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galvan A, Wichmann T. Pathophysiology of parkinsonism. Clin Neurophysiol. 2008;119:1459–1474. doi: 10.1016/j.clinph.2008.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galvan A, Hu X, Smith Y, Wichmann T. Localization and function of GABA transporters in the globus pallidus of parkinsonian monkeys. Exp Neurol. 2010;223:505–515. doi: 10.1016/j.expneurol.2010.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerfen CR, Surmeier DJ. Modulation of striatal projection systems by dopamine. Annu Rev Neurosci. 2011;34:441–466. doi: 10.1146/annurev-neuro-061010-113641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerfen CR, Engber TM, Mahan LC, Susel Z, Chase TN, Monsma FJ, Jr, Sibley DR. D1 and D2 dopamine receptor-regulated gene expression of striatonigral and striatopallidal neurons. Science. 1990;250:1429–1432. doi: 10.1126/science.2147780. [DOI] [PubMed] [Google Scholar]
- Giaume C, Koulakoff A, Roux L, Holcman D, Rouach N. Astroglial networks: a step further in neuroglial and gliovascular interactions. Nat Rev Neurosci. 2010;11:87–99. doi: 10.1038/nrn2757. [DOI] [PubMed] [Google Scholar]
- Gittis AH, Hang GB, LaDow ES, Shoenfeld LR, Atallah BV, Finkbeiner S, Kreitzer AC. Rapid target-specific remodeling of fast-spiking inhibitory circuits after loss of dopamine. Neuron. 2011;71:858–868. doi: 10.1016/j.neuron.2011.06.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabli D, McCairn K, Hirsch EC, Agid Y, Féger J, François C, Tremblay L. Behavioural disorders induced by external globus pallidus dysfunction in primates: I. Behavioural study. Brain. 2004;127:2039–2054. doi: 10.1093/brain/awh220. [DOI] [PubMed] [Google Scholar]
- Groenewegen HJ, Berendse HW, Wolters JG, Lohman AH. The anatomical relationship of the prefrontal cortex with the striatopallidal system, the thalamus and the amygdala: evidence for a parallel organization. Prog Brain Res. 1990;85:95–116. doi: 10.1016/s0079-6123(08)62677-1. discussion 116–118. [DOI] [PubMed] [Google Scholar]
- Haber SN, Groenewegen HJ, Grove EA, Nauta WJ. Efferent connections of the ventral pallidum: evidence of a dual striato pallidofugal pathway. J Comp Neurol. 1985;235:322–335. doi: 10.1002/cne.902350304. [DOI] [PubMed] [Google Scholar]
- Halassa MM, Fellin T, Haydon PG. The tripartite synapse: roles for gliotransmission in health and disease. Trends Mol Med. 2007;13:54–63. doi: 10.1016/j.molmed.2006.12.005. [DOI] [PubMed] [Google Scholar]
- Hallworth NE, Bevan MD. Globus pallidus neurons dynamically regulate the activity pattern of subthalamic nucleus neurons through the frequency-dependent activation of postsynaptic GABAA and GABAB receptors. J Neurosci. 2005;25:6304–6315. doi: 10.1523/JNEUROSCI.0450-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallworth NE, Wilson CJ, Bevan MD. Apamin-sensitive small conductance calcium-activated potassium channels, through their selective coupling to voltage-gated calcium channels, are critical determinants of the precision, pace, and pattern of action potential generation in rat subthalamic nucleus neurons in vitro. J Neurosci. 2003;23:7525–7542. doi: 10.1523/JNEUROSCI.23-20-07525.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond C, Bergman H, Brown P. Pathological synchronization in Parkinson's disease: networks, models and treatments. Trends Neurosci. 2007;30:357–364. doi: 10.1016/j.tins.2007.05.004. [DOI] [PubMed] [Google Scholar]
- Hikosaka O, Wurtz RH. Visual and oculomotor functions of monkey substantia nigra pars reticulate: IV. Relation of substantia nigra to superior colliculus. J Neurophysiol. 1983;49:1285–1301. doi: 10.1152/jn.1983.49.5.1285. [DOI] [PubMed] [Google Scholar]
- Hontanilla B, Parent A, Giménez-Amaya JM. Compartmental distribution of parvalbumin and calbindin D-28k in rat globus pallidus. Neuroreport. 1994;5:2269–2272. doi: 10.1097/00001756-199411000-00016. [DOI] [PubMed] [Google Scholar]
- Hontanilla B, Parent A, de las Heras S, Giménez-Amaya JM. Distribution of calbindin D-28k and parvalbumin neurons and fibers in the rat basal ganglia. Brain Res Bull. 1998;47:107–116. doi: 10.1016/S0361-9230(98)00035-5. [DOI] [PubMed] [Google Scholar]
- Hoover BR, Marshall JF. Population characteristics of preproenkephalin mRNA-containing neurons in the globus pallidus of the rat. Neurosci Lett. 1999;265:199–202. doi: 10.1016/S0304-3940(99)00251-7. [DOI] [PubMed] [Google Scholar]
- Hoover BR, Marshall JF. Further characterization of preproenkephalin mRNA-containing cells in the rodent globus pallidus. Neuroscience. 2002;111:111–125. doi: 10.1016/S0306-4522(01)00565-6. [DOI] [PubMed] [Google Scholar]
- Jaeger D, Gilman S, Aldridge JW. Neuronal activity in the striatum and pallidum of primates related to the execution of externally cued reaching movements. Brain Res. 1995;694:111–127. doi: 10.1016/0006-8993(95)00780-T. [DOI] [PubMed] [Google Scholar]
- Kanaka C, Ohno K, Okabe A, Kuriyama K, Itoh T, Fukuda A, Sato K. The differential expression patterns of messenger RNAs encoding K-Cl cotransporters (KCC1,2) and Na-K-2Cl cotransporter (NKCC1) in the rat nervous system. Neuroscience. 2001;104:933–946. doi: 10.1016/S0306-4522(01)00149-X. [DOI] [PubMed] [Google Scholar]
- Kita H. Globus pallidus external segment. Prog Brain Res. 2007;160:111–133. doi: 10.1016/S0079-6123(06)60007-1. [DOI] [PubMed] [Google Scholar]
- Kita H, Kita T. Number, origins, and chemical types of rat pallidostriatal projection neurons. J Comp Neurol. 2001;437:438–448. doi: 10.1002/cne.1294. [DOI] [PubMed] [Google Scholar]
- Kravitz AV, Freeze BS, Parker PR, Kay K, Thwin MT, Deisseroth K, Kreitzer AC. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature. 2010;466:622–626. doi: 10.1038/nature09159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lange H, Thörner G, Hopf A, Schröder KF. Morphometric studies of the neuropathological changes in choreatic diseases. J Neurol Sci. 1976;28:401–425. doi: 10.1016/0022-510X(76)90114-3. [DOI] [PubMed] [Google Scholar]
- Leventhal DK, Gage GJ, Schmidt R, Pettibone JR, Case AC, Berke JD. Basal ganglia beta oscillations accompany cue utilization. Neuron. 2012;73:523–536. doi: 10.1016/j.neuron.2011.11.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leventhal DK, Stoetzner C, Abraham R, Pettibone J, DeMarco K, Berke JD. Dissociable effects of dopamine on learning and performance within sensorimotor striatum. Basal Ganglia. 2014;4:43–54. doi: 10.1016/j.baga.2013.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magill PJ, Bolam JP, Bevan MD. Dopamine regulates the impact of the cerebral cortex on the subthalamic nucleus-globus pallidus network. Neuroscience. 2001;106:313–330. doi: 10.1016/S0306-4522(01)00281-0. [DOI] [PubMed] [Google Scholar]
- Mallet N, Pogosyan A, Márton LF, Bolam JP, Brown P, Magill PJ. Parkinsonian beta oscillations in the external globus pallidus and their relationship with subthalamic nucleus activity. J Neurosci. 2008a;28:14245–14258. doi: 10.1523/JNEUROSCI.4199-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallet N, Pogosyan A, Sharott A, Csicsvari J, Bolam JP, Brown P, Magill PJ. Disrupted dopamine transmission and the emergence of exaggerated beta oscillations in subthalamic nucleus and cerebral cortex. J Neurosci. 2008b;28:4795–4806. doi: 10.1523/JNEUROSCI.0123-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallet N, Micklem BR, Henny P, Brown MT, Williams C, Bolam JP, Nakamura KC, Magill PJ. Dichotomous organization of the external globus pallidus. Neuron. 2012;74:1075–1086. doi: 10.1016/j.neuron.2012.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maragakis NJ, Rothstein JD. Mechanisms of disease: astrocytes in neurodegenerative disease. Nat Clin Pract Neurol. 2006;2:679–689. doi: 10.1038/ncpneuro0355. [DOI] [PubMed] [Google Scholar]
- Mastro KJ, Bouchard RS, Holt HA, Gittis AH. Transgenic mouse lines subdivide distinct neuronal populations in the external segment of the globus pallidus. J Neurosci. 2014;34:2087–2099. doi: 10.1523/JNEUROSCI.4646-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurice N, Deniau JM, Glowinski J, Thierry AM. Relationships between the prefrontal cortex and the basal ganglia in the rat: physiology of the corticosubthalamic circuits. J Neurosci. 1998;18:9539–9546. doi: 10.1523/JNEUROSCI.18-22-09539.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mogenson GJ, Swanson LW, Wu M. Neural projections from nucleus accumbens to globus pallidus, substantia innominata, and lateral preoptic-lateral hypothalamic area: an anatomical and electrophysiological investigation in the rat. J Neurosci. 1983;3:189–202. doi: 10.1523/JNEUROSCI.03-01-00189.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran RJ, Mallet N, Litvak V, Dolan RJ, Magill PJ, Friston KJ, Brown P. Alterations in brain connectivity underlying beta oscillations in Parkinsonism. PLoS Comput Biol. 2011;7:e1002124. doi: 10.1371/journal.pcbi.1002124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nambu A, Llinaś R. Electrophysiology of globus pallidus neurons in vitro. J Neurophysiol. 1994;72:1127–1139. doi: 10.1152/jn.1994.72.3.1127. [DOI] [PubMed] [Google Scholar]
- Nedergaard M, Verkhratsky A. Artifact versus reality: how astrocytes contribute to synaptic events. Glia. 2012;60:1013–1023. doi: 10.1002/glia.22288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nini A, Feingold A, Slovin H, Bergman H. Neurons in the globus pallidus do not show correlated activity in the normal monkey, but phase-locked oscillations appear in the MPTP model of parkinsonism. J Neurophysiol. 1995;74:1800–1805. doi: 10.1152/jn.1995.74.4.1800. [DOI] [PubMed] [Google Scholar]
- Nóbrega-Pereira S, Gelman D, Bartolini G, Pla R, Pierani A, Marín O. Origin and molecular specification of globus pallidus neurons. J Neurosci. 2010;30:2824–2834. doi: 10.1523/JNEUROSCI.4023-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obeso JA, Rodriguez-Oroz MC, Benitez-Temino B, Blesa FJ, Guridi J, Marin C, Rodriguez M. Functional organization of the basal ganglia: therapeutic implications for Parkinson's disease. Mov Disord. 2008;23(Suppl 3):S548–S559. doi: 10.1002/mds.22062. [DOI] [PubMed] [Google Scholar]
- Otsuka T, Murakami F, Song WJ. Excitatory postsynaptic potentials trigger a plateau potential in rat subthalamic neurons at hyperpolarized states. J Neurophysiol. 2001;86:1816–1825. doi: 10.1152/jn.2001.86.4.1816. [DOI] [PubMed] [Google Scholar]
- Perea G, Navarrete M, Araque A. Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci. 2009;32:421–431. doi: 10.1016/j.tins.2009.05.001. [DOI] [PubMed] [Google Scholar]
- Plenz D, Kital ST. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus. Nature. 1999;400:677–682. doi: 10.1038/23281. [DOI] [PubMed] [Google Scholar]
- Sato F, Lavallée P, Lévesque M, Parent A. Single-axon tracing study of neurons of the external segment of the globus pallidus in primate. J Comp Neurol. 2000;417:17–31. doi: 10.1002/(SICI)1096-9861(20000131)417:1<17::AID-CNE2>3.0.CO%3B2-I. [DOI] [PubMed] [Google Scholar]
- Schmidt R, Leventhal DK, Mallet N, Chen F, Berke JD. Canceling actions involves a race between basal ganglia pathways. Nat Neurosci. 2013;16:1118–1124. doi: 10.1038/nn.3456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt R, Mallet N, Leventhal DK, Chen F, Berke JD. The globus pallidus cancels actions by suppressing striatal output. Soc Neurosci Abstr. 2014;40 442.17. [Google Scholar]
- Shimamoto SA, Ryapolova-Webb ES, Ostrem JL, Galifianakis NB, Miller KJ, Starr PA. Subthalamic nucleus neurons are synchronized to primary motor cortex local field potentials in Parkinson's disease. J Neurosci. 2013;33:7220–7233. doi: 10.1523/JNEUROSCI.4676-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith Y, Bevan MD, Shink E, Bolam JP. Microcircuitry of the direct and indirect pathways of the basal ganglia. Neuroscience. 1998;86:353–387. doi: 10.1016/S0306-4522(98)00004-9. [DOI] [PubMed] [Google Scholar]
- Sofroniew MV, Vinters HV. Astrocytes: biology and pathology. Acta Neuropathol. 2010;119:7–35. doi: 10.1007/s00401-009-0619-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tachibana Y, Iwamuro H, Kita H, Takada M, Nambu A. Subthalamo–pallidal interactions underlying parkinsonian neuronal oscillations in the primate basal ganglia. Eur J Neurosci. 2011;34:1470–1484. doi: 10.1111/j.1460-9568.2011.07865.x. [DOI] [PubMed] [Google Scholar]
- Theodosis DT, Poulain DA, Oliet SH. Activity-dependent structural and functional plasticity of astrocyte–neuron interactions. Physiol Rev. 2008;88:983–1008. doi: 10.1152/physrev.00036.2007. [DOI] [PubMed] [Google Scholar]
- Tong X, Ao Y, Faas GC, Nwaobi SE, Xu J, Haustein MD, Anderson MA, Mody I, Olsen ML, Sofroniew MV, Khakh BS. Astrocyte Kir4.1 ion channel deficits contribute to neuronal dysfunction in Huntington's disease model mice. Nat Neurosci. 2014;17:694–703. doi: 10.1038/nn.3691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner RS, Anderson ME. Pallidal discharge related to the kinematics of reaching movements in two dimensions. J Neurophysiol. 1997;77:1051–1074. doi: 10.1152/jn.1997.77.3.1051. [DOI] [PubMed] [Google Scholar]
- Urbain N, Gervasoni D, Soulière F, Lobo L, Rentéro N, Windels F, Astier B, Savasta M, Fort P, Renaud B, Luppi PH, Chouvet G. Unrelated course of subthalamic nucleus and globus pallidus neuronal activities across vigilance states in the rat. Eur J Neurosci. 2000;12:3361–3374. doi: 10.1046/j.1460-9568.2000.00199.x. [DOI] [PubMed] [Google Scholar]
- Vitek JL, Zhang J, Hashimoto T, Russo GS, Baker KB. External pallidal stimulation improves parkinsonian motor signs and modulates neuronal activity throughout the basal ganglia thalamic network. Exp Neurol. 2012;233:581–586. doi: 10.1016/j.expneurol.2011.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voorn P, Vanderschuren LJ, Groenewegen HJ, Robbins TW, Pennartz CM. Putting a spin on the dorsal-ventral divide of the striatum. Trends Neurosci. 2004;27:468–474. doi: 10.1016/j.tins.2004.06.006. [DOI] [PubMed] [Google Scholar]
- Wilson CJ. Active decorrelation in the basal ganglia. Neuroscience. 2013;250:467–482. doi: 10.1016/j.neuroscience.2013.07.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu Z, Bartol M, Shen K, Johnson SW. Excitatory effects of dopamine on subthalamic nucleus neurons: in vitro study of rats pretreated with 6-hydroxydopamine and levodopa. Brain Res. 2002;945:31–40. doi: 10.1016/S0006-8993(02)02543-X. [DOI] [PubMed] [Google Scholar]