

Abstract
The discovery of the size and complexity of the human microbiome has resulted in an ongoing reevaluation of many concepts of health and disease, including diseases affecting the CNS. A growing body of preclinical literature has demonstrated bidirectional signaling between the brain and the gut microbiome, involving multiple neurocrine and endocrine signaling mechanisms. While psychological and physical stressors can affect the composition and metabolic activity of the gut microbiota, experimental changes to the gut microbiome can affect emotional behavior and related brain systems. These findings have resulted in speculation that alterations in the gut microbiome may play a pathophysiological role in human brain diseases, including autism spectrum disorder, anxiety, depression, and chronic pain. Ongoing large-scale population-based studies of the gut microbiome and brain imaging studies looking at the effect of gut microbiome modulation on brain responses to emotion-related stimuli are seeking to validate these speculations. This article is a summary of emerging topics covered in a symposium and is not meant to be a comprehensive review of the subject.
Introduction
Traditionally, microorganisms have not been considered of particular importance to the development and function of the CNS or in the pathophysiology of chronic brain diseases, such as disorders of mood and affect, Parkinson's disease, or Alzheimer's disease. The often quoted and remarkable ability of the parasite Toxoplasmosis gondii of hijacking the host's (e.g., rat) brain systems related to defensive behaviors and sexual attraction to manipulate the rat's behavior in a way that optimizes reproduction of the parasite (House et al., 2011) was considered an interesting outlier in the prevalent dogma of looking exclusively at the brain for causes of behavior and brain diseases. One exception to the traditional view has been autism spectrum disorder (ASD), a brain disease that has long been suspected to be related to altered gut microbiota (Mayer et al., 2014a), a concept that has recently been revisited both in rodent models and in human subjects. The “microbiome-free” worldview of neuroscience has dramatically changed with the discovery and characterization of the human microbiome and, in particular, with the gut microbiome (Human Microbiome Project Consortium, 2012). Although gut-brain interactions have been studied for decades, providing a wealth of information about the close interactions between the gut-associated immune system, enteric nervous system, and gut-based endocrine system (Mayer, 2011), these findings have largely been ignored by the psychiatric and neurological research community. The discovery of the gut microbiome has added a long overlooked component to the complex bidirectional signaling between mind, brain, gut, and its microbiome and surprisingly has triggered a tremendous interest by the professional and lay media, as well as by national funding agencies, including the National Institute of Mental Health. The initial skepticism about reports suggesting a profound role of an intact gut microbiota in shaping brain neurochemistry and emotional behavior has given way to an unprecedented paradigm shift in the conceptualization of many psychiatric and neurological diseases. Although many of the new concepts are primarily based on the intriguing experimental findings in rodents, initial studies in humans seem to support the notion that there is a relationship between the complex world of microbiota in our intestines and brain structure and function. Even though the majority of published studies of gut microbiome to brain signaling are based on microbiome analyses from stool samples, future studies will almost certainly expand the scope of investigations to mucosal samples taken from different regions of the gastrointestinal tract. Based on our current, still limited knowledge about these gut-microbiome-brain interactions, intriguing speculations have been proposed in a rapidly increasing number of review articles on the topic. They range from terms, such as “psychobiotic” or “melancholic” microbes (Cryan and Dinan, 2012), to concepts that humans are just the vehicle for the 100 trillion microorganisms living inside of us. The latter concept has been developed into the intriguing hypothesis that the gut microbiota have developed ways to “hack” into our reward system to make us crave certain foods and avoid others that are most beneficial to them (Alcock et al., 2014) Similarly, microbe-brain interactions have been recently proposed to be a key driver of the evolution of the social brain (Stilling et al., 2014b). The following review addresses some aspects of the rapidly evolving topic of gut-microbiome-brain interactions in health and disease (Fig. 1). Even though not a comprehensive review of the topic, it provides a glance into this emerging new view of neuroscience.
Figure 1.
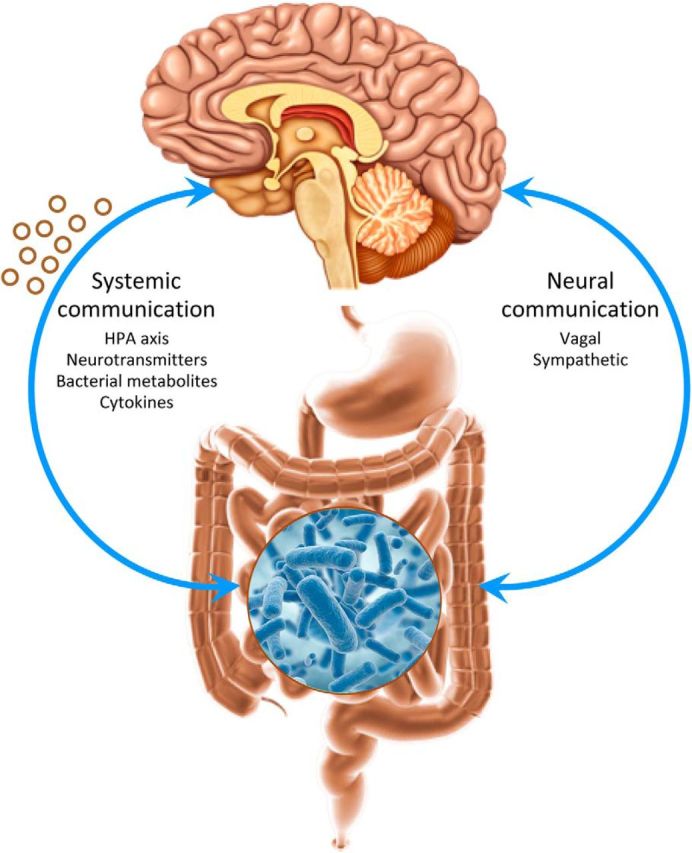
Bidirectional communication channels between the gut microbiome, the gut, and the brain. Endocrine-, neurocrine-, and inflammation-related signals generated by the gut microbiota and specialized cells within the gut can, in principal, affect the brain. In turn, the brain can influence microbial composition and function via endocrine and neural mechanisms.
Gut microbiota regulates stress, anxiety, and cognition: mechanisms and therapeutic potential
Accumulating evidence, largely from animal studies, suggests that different types of psychological stress can affect the composition of the gut microbiota. For example, maternal separation, restraint conditions, crowding, heat stress, and acoustic stress all alter the composition of the gut microbiota (Bailey et al., 2011; De Palma et al., 2014; Moloney et al., 2014). In addition, a growing body of data suggests that the microbiota may be involved in controlling behaviors relevant to stress-related disorders.
Several experimental conditions have been used to study the role of the gut microbiota in preclinical models, including perturbation of the gut microbiome by ingestion of probiotics and antibiotics, fecal microbial transplant, and comparison of behaviors and biological readouts between germ-free animals (raised in a sterile environment from the time of birth) and those with a pathogen-free microbiome.
Germ-free animals
It is now a decade since Sudo et al. (2004) discovered that germ-free mice have an exaggerated hypothalamic-pituitary-adrenal axis response to restraint stress, an effect that was reversed by monocolonization with a particular Bifidobacterium species. This seminal observation motivated a number of research groups to investigate the role of the host gut microbiota on CNS function, with converging and intriguing results. Despite exaggerated neuroendocrine responses to stress, consistent reductions in anxiety-like behavior were observed in germ-free mice exposed to more ecologically relevant stressors, such as novel and aversive environments (elevated plus maze, light/dark box, open field) (Diaz Heijtz et al., 2011; Neufeld et al., 2011; Clarke et al., 2013). This phenotype was susceptible to reversal when animals were colonized early in life (Clarke et al., 2013). Interestingly, recent studies in germ-free animals in the stress-sensitive F344 rat strain showed similar exaggerated neuroendocrine responses but also revealed an increase in anxiety-like behavior (Crumeyrolle-Arias et al., 2014). Moreover, it has recently been shown that short-term colonization of germ-free mice in adulthood reduced anxiety-like behaviors (Nishino et al., 2013). Together, it is clear that studies in germ-free animals clearly show a relationship between gut microbiota and stress and anxiety-related behaviors, the nature of this relationship being influenced by temporal, sex, strain, and species factors that are not yet fully understood.
A growing number of studies are also investigating gene expression changes in different brain regions in germ-free mice. Most commonly, decreases in the hippocampal expression of BDNF, a key protein involved in neuronal plasticity and cognition, have been observed in germ-free mice relative to conventionally raised or conventionalized (i.e., initially germ-free mice colonized with the normal mouse gut microbiota) controls. Similar changes in BDNF expression have also been reported following antibiotic administration (Bercik et al., 2011b). Alterations in neurotransmitter signaling, including neurotransmitters and associated metabolites and neurotransmitter receptors, have also been described in specific brain regions of germ-free mice. Diaz Heijtz et al. (2011) took a genome-wide transcriptomic approach showing that genes associated with the citrate cycle (synaptic long-term potentiation, steroid hormone metabolism, and cyclic adenosine 5-phosphate-mediated signaling) were upregulated in germ-free mice. Interestingly, in these studies, the cerebellum and hippocampus have robust changes in gene expression, but the hypothalamus, the brain region involved in the stress response, showed almost no differential gene expression.
Some behavioral and biochemical parameters (including anxiety, sociability, hypothalamic-pituitary-adrenal axis, and tryptophan metabolism) could be reversed in germ-free mice by recolonization with a conventional microbiota or probiotic treatment, but others were unaffected by restoration of a normal microbiota (including 5-HT concentration and social cognition) (Stilling et al., 2014a). Indeed, it has been thought that reversibility of the anxiolytic phenotype in germ-free mice is only guaranteed if recolonization happens during a critical time window during early-life/adolescence (Neufeld et al., 2011; Clarke et al., 2013). This has recently been challenged with evidence that adult colonization affects anxiety-like behaviors (Collins et al., 2013; Nishino et al., 2013).
It is important to note that the clinical translatability of germ-free studies is limited. Early life antibiotic administration in rodents does not recapitulate many of the behavioral germ-free phenotypes (O'Mahony et al., 2014). One factor contributing to this limited translational relevance is the fact that germ-free animals exhibit major alterations in gastrointestinal function, including gross dilation of the proximal colon and alterations in motility, and in the immune system, which presumably also has important effects on the brain. Nonetheless, germ-free studies are powerful in that they test definitively whether the microbiota is involved in a specific aspect of brain function. Germ-free mice also enable the study of the impact of a particular bacterial or dietary intervention on the microbiota-gut-brain axis in isolation. Studies in germ-free mice can also be expanded to enable research on the “humanization” of the gut microbiota (i.e., transplanting fecal microbiota from specific human conditions or from animal models of disease). In this regard, intriguing studies have shown that the transplantation of microbiota from a high-anxiety mouse strain to a germ-free low-anxiety recipient in adulthood was sufficient to increase anxiety in the recipient, and the converse was also true (Collins et al., 2013). These studies also support the concept that the behavior of germ-free animals is susceptible to alteration even into adulthood. Further evidence of the microbial-based transferability of behavior comes from recent study whereby mice, whose baseline microbiota were ablated with antibiotics, were given the microbiota from donor animals that had been fed a high-fat diet. These mice had selective disruptions in exploratory, cognitive, and stereotypical behavior compared with mice transplanted with control microbiota in the absence of significant differences in body weight (Bruce-Keller et al., 2014).
Probiotics
A growing body of evidence from rodent studies further supports a role of the gut microbiome in modulating emotional behavior. In animal models, a range of probiotics have been investigated. Bifidobacterium and Lactobacillus are the main genera showing beneficial effects on anxiety- and depression-like behavior. However, even within bacterial genera, marked strain differences occur, and only a few strains have any positive effects (Dinan et al., 2013). Chronic treatment with Bifidobacterium infantis attenuated early-life stress-induced immune changes and depressive-like behaviors in adulthood (Desbonnet et al., 2010). Lactobacillus helveticus ROO52 has also been shown to reduce anxiety-like behavior and alleviate memory dysfunction (Ohland et al., 2013). Lactobacillus rhamnosus JB-1 reduced anxiety- and depression-related behaviors in the elevated plus maze and forced swim test, respectively (Bravo et al., 2011). Recent work by Matthews and Jenks (2013) demonstrated reduced anxiety and improved performance on a complex maze task after treatment with live Mycobacterium vaccae. Bifidobacterium longum normalizes anxiety-like behavior in a colitis model (Bercik et al., 2011a). Furthermore, a B. longum, but not L. rhamnosus strain, normalized infection-induced anxiety-like behavior (Bercik et al., 2010). A combination of L. rhamnosus and L. helveticus reversed stress-induced memory dysfunction in Citrobacter rodentium-infected mice (Gareau et al., 2011). More recently, it has been shown that VSL#3 (a mixture of eight different probiotics) was able to reverse age- associated deficits in long-term potentiation, the electrophysiological correlate of memory formation (Distrutti et al., 2014). Probiotic treatment has also proved efficacious in alleviating visceral pain responses in animal models (Rousseaux et al., 2007; McKernan et al., 2010). Another potential strategy for modulating the microbiome-gut-brain axis is the use of prebiotics, nondigestible food ingredients that promote the growth of beneficial gut microorganisms (probiotics). Surprisingly, there has been a paucity of studies in animals or humans with prebiotics, although specific prebiotics have been shown to increase brain BDNF levels (Savignac et al., 2013).
Overall, accumulating evidence in rodent studies suggests that there are links among the microbiota composition, brain biochemistry, and behavior and that these interactions may be particularly important at critical neurodevelopmental windows (Borre et al., 2014). The underlying molecular mechanisms leading to these behavioral and biochemical alterations are not well understood. Bacterial metabolites include many neuroactive agents (Lyte, 2013; Wall et al., 2014), and understanding the specific components of the microbial metabolome will be important for understanding the role of the microbiome in brain health and disease (Holmes et al., 2012). Interestingly, there is now a growing appreciation of the role of epigenetic mechanisms in shaping brain and behavior, and it is worth noting that many bacterial metabolites can act as epigenetic modifiers (Stilling et al., 2014a).
Evidence for alterations in gut-microbiome-brain interactions in a rodent model of ASD
Alterations in the communication between the gut microbiome and the brain, including alterations in the composition and metabolic products of the gut microbiome, have been implicated in the complex pathophysiology of ASD. Gastrointestinal symptoms are a common comorbidity in ASD patients, even though the underlying mechanisms are largely unknown. Several types of rodent models for human ASD have been proposed: (1) naturally occurring rodent strains that demonstrate ASD-relevant behavioral traits; (2) models expressing a human genetic mutation associated with ASD; and (3) models with acquired behaviors resulting from various environmental insults either affecting the developing animal directly (Desbonnet et al., 2014; de Theije et al., 2014) or affecting the mother of ASD offspring. In one such prenatally induced model, exposure of pregnant mice treated with valproic acid resulted in ASD-like behavior in the offspring, which were associated with alterations in the gut microbiome associated with inflammatory and endocrine changes in both intestinal tract and in the nervous system.
Further linking the microbiota to ASD, germ-free mice showed reduced sociability and have social cognition deficits in the three-chamber test and exhibited increased repetitive grooming behavior compared with their conventional counterparts (Desbonnet et al., 2014). Interestingly, the deficits in sociability and repetitive behaviors, but not social cognition, were reversed by postweaning colonization. Recent provocative studies point to the ability of a Bacteroides fragilis strain given in early postweaning life to reverse gastrointestinal, microbiota, and selective behavioral changes induced in a prenatal infection model of neurodevelopmental disorders, such as ASD and schizophrenia (Hsiao et al., 2013).
Overall, there is intriguing preclinical and some clinical evidence to implicate alterations of the gut microbiome in the pathophysiology of ASD. However, it remains to be determined whether the observed microbiota changes are secondary to altered neural (CNS, enteric nervous system) regulation of key gut functions (motility, secretion), or if they represent primary peripheral alterations that affect brain development and function (Mayer et al., 2014a). As ASD is a heterogeneous group of disorders, it is unlikely that one disease mechanism (such as altered gut microbiota to brain signaling) applies to all disease phenotypes.
Gut-microbiome signaling in humans
Although our ability to study gut-microbiome-brain interactions is more limited in humans than it is in preclinical models, it is possible to obtain a broad view of the gut microbiota composition and metabolites by analysis of fecal samples and aim to correlate such findings with brain activity and structure using neuroimaging (Tillisch and Labus, 2014). Further, modulation of the gastrointestinal microbiota in humans via foods, supplements (including probiotics and prebiotics), or medications (including antibiotics) can be used as experimental probes for more mechanistic studies of brain-gut microbiome interactions (Mayer et al., 2014b).
Modulation of the gut-brain axis with antibiotics
Manipulation of gut bacteria by antibiotics is used clinically to improve brain function in hepatic encephalopathy, a complication of chronic liver disease (Butterworth, 2013). Patients with hepatic encephalopathy have variable alterations in cognitive function, presumably due to gut microbial metabolites that are not efficiently cleared by the diseased liver. In patients with mild hepatic encephalopathy, 8 weeks of oral treatment with a nonabsorbable antibiotic was associated with improvements in cognitive function based on a battery of standardized tests (Bajaj et al., 2013). These cognitive changes occurred in the absence of major changes in the overall microbiome composition by principal component analysis. However, metabolomic changes were detected in the serum. Short-chain fatty acids, a major metabolic product of the gut microbiota with well-known effects on the nervous system, were increased (Haast and Kiliaan, 2014). The authors hypothesized that the changes in profiles of fatty acids in the periphery may correspond to more favorable profiles of brain fatty acids, as a mechanism for improved cognition. Treatment of hepatic encephalopathy with the same antibiotic in an open label study was associated with altered functional connectivity during a cognitive task and changes in white matter integrity (as measured by fractional anisotropy) (Ahluwalia et al., 2014). These changes support previous work in animal models, in which antibiotic treatment has been associated with both behavioral changes and changes in neurochemistry (Bercik et al., 2011b). Surprisingly, despite the widespread use of antibiotics and clinical descriptions of cognitive and psychiatric side effects, little evaluation of the role of antibiotics in the human microbiome-brain axis has taken place (Sternbach and State, 1997; Tomé and Filipe, 2011).
Modulation of the gut-brain axis with probiotics
Probiotics are used widely; and as consumer products, they represent a >20 billion dollar industry worldwide. However, despite many unsubstantiated claims, probiotic effects on the structure and function of the human gut microbiota have been studied in only a few specific strains, and our understanding of their effects on clinical symptoms is far from complete (Sanders et al., 2013). Several probiotics have been shown to have utility for specific gastrointestinal and global symptoms in irritable bowel syndrome, a chronic pain condition characterized by dysregulation of the brain-gut axis (Moayyedi et al., 2010; Mayer et al., 2014b). Whether these benefits are due primarily to peripheral actions in the gut or due to central effects is not known. In chronic fatigue syndrome, another disorder of brain–body interactions, a randomized, double-blind, placebo-controlled trial of a Lactobacillus-containing probiotic decreased anxiety, but not depression symptoms, in the active treatment group, and increased the relative abundance of Bifidobacterium and Lactobacillus in the stool (Rao et al., 2009). This study, published as a brief report, lacked detail in terms of the reported results, thus should be interpreted with caution. Two small studies of probiotics effects on mood and cognition in healthy individuals have been published, both of which suggest an effect on the microbiome brain axis. In the first, a Lactobacillius- and Bifidobacterium-containing probiotic was compared with placebo in healthy volunteers, measuring mood symptoms with the Hospital Anxiety and Depression Scale. The percentage decrease in the total Hospital Anxiety and Depression Scale score was greater in the probiotic group, but not in the subscales (Messaoudi et al., 2011). A reduction in urinary-free cortisol was also seen over the course of the treatment in the probiotic group, but not the placebo group, although the group difference was not significant. In this study, the authors ran an experimental arm of probiotic treatment in rodents, and consistent with the human results, saw improved behavioral performance in an anxiety-like task. In the second study, the effects of a Lactobacillus containing dairy drink were compared with placebo, with no significant group changes in mood, as measured by the Profile of Mood States (Benton et al., 2007). The authors suggest that this lack of effect may have been due to the overall positive mood of the sample, and did note a small effect when looking at a post hoc small subgroup of subjects at the lowest tertile of mood states. Surprisingly, this same study showed diminished memory scores in the probiotic groups versus the placebo group. Although these studies overall suggest a potential for positive effects of some probiotics on mood, clearly larger, well-designed clinical trials, ideally with biological as well as self-report outcomes in patient populations, are needed to make clear conclusions.
Imaging of the brain in gut-microbiome-brain interactions
Although they are important from a mechanistic standpoint, most preclinical studies have used nonphysiological experimental interventions (e.g., germ-free animals), simplified constructs of complex human emotions (anxiety and depression), and are often based on erroneous assumptions about the anatomical and functional homology of the rodent and human brains (in particular the prefrontal cortex and the anterior insula). Therefore, it is currently unclear what the translational value of the results obtained in these rodent models for understanding brain or brain-gut disorders in humans will be. Clearly, more studies in humans are needed to verify some of the intriguing animal findings. fMRI can be used in humans to observe changes in brain response after a probiotic or antibiotic interventions, just as it is commonly used to test pharmaceuticals or behavioral interventions (Mayer et al., 2002; Wise and Tracey, 2006; Tillisch et al., 2008). The effect of daily probiotic intake on brain responses to an emotion recognition task have been described in healthy women (Tillisch et al., 2013). In this study, women without any gastrointestinal symptoms, pain, or psychiatric disorder were randomized to treatment with a probiotic, a nonfermented dairy product or no treatment. The response to negative affect faces in an emotional recognition task was measured with fMRI before and after the treatment period. Compared with both control groups, the probiotic group showed reductions in response to the task in the insula and somatosensory cortices specifically, as well as across a widespread functional network, including emotional and sensory regions. Although alternative interpretations are possible, these findings may represent a reduction in vigilance to negative environmental stimuli in subjects who regular consume probiotics. The changes in brain activity were independent of self-reported gastrointestinal symptoms, indicating that the central effect was not likely due to an improved sense of digestive well-being. Confirming results from a previous study with detailed microbial and metabolomics analyses (McNulty et al., 2011), no group-specific changes in the overall architecture of the microbiota were observed after 4 weeks of probiotic ingestion, although stool samples did provide confirmation that the specific probiotic strains were present in the treatment group. This is consistent with the hypothesis that, at least in the short term, microbial metabolites rather than overall microbial configuration may be the salient result of probiotic ingestion (McNulty et al., 2011). Functional and structural neuroimaging, along with metabolomic and metagenomic measurements from stool, will be essential in providing a better understanding of how the gut-microbiome-brain axis functions in human health and disease.
Multilevel data integration of brain and microbiome-related data
Advances in computational approaches are urgently needed to better understand links between the gut microbiota and the brain. Of particular importance are tools to integrate large, highly multivariate datasets (Gonzalez and Knight, 2012; Navas-Molina et al., 2013). These datasets include taxonomic profiles obtained from 16S rRNA amplicon sequencing or shotgun metagenomics, gene catalogs from shotgun metagenomics, expression datasets from mRNA sequencing or proteomics, metabolite profiles from targeted or untargeted metabolomics, behavioral datasets, and imaging data, including structural and fMRI. Increasingly, these datasets also involve time series components, especially as the data acquisition methods decrease in cost (for example, 16S rRNA amplicon profiling has decreased in cost by almost a factor of a million over the past decade) (Kuczynski et al., 2012). As data analysis techniques in each of these areas individually are evolving rapidly, integrating them is even more of a moving target.
Many microbiome-brain links may have been overlooked because other causal pathways seemed initially more plausible. For example, TLR5 knock-out mice, in certain facilities, gain weight with respect to wild-type mice on the same diet and develop metabolic syndrome. This effect disappears when the mice are raised germ-free, and can be treated with antibiotics. Fascinatingly, the mechanism is behavioral: in obesogenic environments, the TLR5 knock-out mice have an altered microbial community that can be transmitted even to genetically normal wild-type mice that are raised germ-free. This altered microbial community induces a change in ingestive behavior (e.g., hyperphagy: obesity can be prevented and reversed by placing in the cage only the amount of food a wild-type mouse would eat) (Vijay-Kumar et al., 2010). Although the mechanism of this effect remains unknown, particular species of bacteria in the gut are known to affect the levels of appetite-regulating hormones, including leptin (Ravussin et al., 2012) and ghrelin (Queipo-Ortuño et al., 2013). Similarly, probiotic effects of L. rhamnosis on host GABA receptor expression require an intact vagus nerve (Bravo et al., 2011). The potential for many other microbially induced phenotypes to act via behavioral mechanisms, including neural signaling, is thus immense and at present underexplored.
Tools for tracking the dynamics of the microbiome are rapidly evolving (Caporaso et al., 2011; Vázquez-Baeza et al., 2013) and provide insights both into normal microbiome variation and into its changes in responses to therapy, including antibiotics (Lozupone et al., 2013) and fecal microbiota transplant (Khoruts A, Sadowsky M, University of Minnesota, unpublished data). These tools can easily be extended to other multivariate datasets, including features derived from imaging datasets, through use of the Biological Observation Matrix file format (McDonald et al., 2012). Essentially, the technique is to use a large population, such as the Human Microbiome Project Consortium (2012), as a data frame, then project the time series corresponding to one or more individuals as animations in that data frame, also recording derived features, such as the variability over time, the direction of change in the multivariate space, etc. (Carvalho et al., 2012).
The American Gut Project
An especially exciting opportunity is provided by the American Gut Project, which uses crowd sourcing (obtaining samples and/or assistance with data analysis from members of the general public) and crowd funding (obtaining financial support from members of the general public) to obtain thousands of fecal (and other) samples, and analyze the bacterial communities contained in those samples. Several studies have shown that storage of unfixed samples at room temperature do not alter the major microbiological conclusions (e.g., clustering of samples from the same subject together) (Lauber et al., 2010; Wu et al., 2010). Although there are differences between the stool microbiota and other sites in the distal large intestine in the same subject, these differences are smaller than the differences between subjects, and so, provide a good readout of the distal large intestine (although note that the small intestine differs substantially in its microbiology) (Eckburg et al., 2005; Hamady and Knight, 2009; Gevers et al., 2014). As of this writing, American Gut has publicly released sequence data from almost 4000 microbial samples from members of the general public. Excitingly, this dataset easily allows us to integrate information from clinical populations, and specialized studies are underway in depression, ASD, and irritable bowel syndrome and celiac disease (both of which have substantial comorbidity with depression), multiple sclerosis, and other conditions affecting the nervous system in which microbes are either known to be involved in humans, are involved in mouse models or where plausible, though speculative, pathways exist. Especially, the availability of methods to correlate different kinds of data, and to integrate detailed time-series, including cycles of remission and relapse or the effects of specific treatments, provide substantial hope for identifying new ways of stratifying patients and new treatment modalities.
In conclusion, the discovery and the explosive progress in the characterization of the gut microbiome have initiated a paradigm shift not only in medicine, but also in the basic and clinical domains of neuroscience. To understand the magnitude of this paradigm shift, the reader has to be reminded of the powerful grip of Descartes' separation of mind/brain on the one side (religion, psychiatry) and body on the other side (medicine) that has dominated Western science and medicine for hundreds of years. Not only is the concept of gut-microbiome-brain interactions in health and disease paradigm breaking, the emerging data-driven, analytical methodologies that are required to pursue the integration of massive amounts of data are equally revolutionary. It is difficult to predict the trajectory of exciting period of discovery: Will the gut microbiome add paradigm-transforming insights to our existing understanding of human brain function in health and disease, resulting in novel therapies, or will it represent an incremental step in understanding the inner workings of our brains? The next few years of research hold the potential of uncovering intriguing connections between gut bacteria and neurological conditions that may possibly impact human health.
Footnotes
This work was supported by National Institutes of Health/National Institute of Diabetes and Digestive and Kidney Diseases Grant R01 DK048351 to E.A.M., Grant P30 DK041301, National Institutes of Health/National Institute of Mental Health Grant R01 MH100556 to S.K.M., Autism Speaks to S.K.M., Simons Foundation SFARI Program to S.K.M., and Howard Hughes Medical Institute to R.K.
The authors declare no competing financial interests.
References
- Ahluwalia V, Wade JB, Heuman DM, Hammeke TA, Sanyal AJ, Sterling RK, Stravitz RT, Luketic V, Siddiqui MS, Puri P, Fuchs M, Lennon MJ, Kraft KA, Gilles H, White MB, Noble NA, Bajaj JS. Enhancement of functional connectivity, working memory and inhibitory control on multi-modal brain MR imaging with rifaximin in cirrhosis: implications for the gut-liver-brain axis. Metab Brain Dis. 2014 doi: 10.1007/s11011-014-9507-6. doi: 10.1007/s11011-014-9507-6. Advance online publication. Retrieved Mar. 4, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alcock J, Maley CC, Aktipis CA. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. Bioessays. 2014;36:940–949. doi: 10.1002/bies.201400071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey MT, Dowd SE, Galley JD, Hufnagle AR, Allen RG, Lyte M. Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain Behav Immun. 2011;25:397–407. doi: 10.1016/j.bbi.2010.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajaj JS, Heuman DM, Sanyal AJ, Hylemon PB, Sterling RK, Stravitz RT, Fuchs M, Ridlon JM, Daita K, Monteith P, Noble NA, White MB, Fisher A, Sikaroodi M, Rangwala H, Gillevet PM. Modulation of the metabiome by rifaximin in patients with cirrhosis and minimal hepatic encephalopathy. PLoS One. 2013;8:e60042. doi: 10.1371/journal.pone.0060042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benton D, Williams C, Brown A. Impact of consuming a milk drink containing a probiotic on mood and cognition. Eur J Clin Nutr. 2007;61:355–361. doi: 10.1038/sj.ejcn.1602546. [DOI] [PubMed] [Google Scholar]
- Bercik P, Verdu EF, Foster JA, Macri J, Potter M, Huang X, Malinowski P, Jackson W, Blennerhassett P, Neufeld KA, Lu J, Khan WI, Corthesy-Theulaz I, Cherbut C, Bergonzelli GE, Collins SM. Chronic gastrointestinal inflammation induces anxiety-like behavior and alters central nervous system biochemistry in mice. Gastroenterology. 2010;139:2102–2112. doi: 10.1053/j.gastro.2010.06.063. [DOI] [PubMed] [Google Scholar]
- Bercik P, Denou E, Collins J, Jackson W, Lu J, Jury J, Deng Y, Blennerhassett P, Macri J, McCoy KD, Verdu EF, Collins SM. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology. 2011a;141:599–609. doi: 10.1053/j.gastro.2011.04.052. [DOI] [PubMed] [Google Scholar]
- Bercik P, Park AJ, Sinclair D, Khoshdel A, Lu J, Huang X, Deng Y, Blennerhassett PA, Fahnestock M, Moine D, Berger B, Huizinga JD, Kunze W, McLean PG, Bergonzelli GE, Collins SM, Verdu EF. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol Motil. 2011b;23:1132–1139. doi: 10.1111/j.1365-2982.2011.01796.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borre YE, O'Keeffe GW, Clarke G, Stanton C, Dinan TG, Cryan JF. Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med. 2014;20:509–518. doi: 10.1016/j.molmed.2014.05.002. [DOI] [PubMed] [Google Scholar]
- Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, Bienenstock J, Cryan JF. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A. 2011;108:16050–16055. doi: 10.1073/pnas.1102999108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruce-Keller AJ, Salbaum JM, Luo M, Blanchard E, IV, Taylor CM, Welsh DA, Berthoud HR. Obese-type gut microbiota induce neurobehavioral changes in the absence of obesity. Biol Psychiatry. 2014 doi: 10.1016/j.biopsych.2014.07.012. doi: 10.1016/j.biopsych.2014.07.012. Advance online publication. Retrieved Jul. 18, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butterworth RF. The liver-brain axis in liver failure: neuroinflammation and encephalopathy. Nat Rev Gastroenterol Hepatol. 2013;10:522–528. doi: 10.1038/nrgastro.2013.99. [DOI] [PubMed] [Google Scholar]
- Caporaso JG, Lauber CL, Costello EK, Berg-Lyons D, Gonzalez A, Stombaugh J, Knights D, Gajer P, Ravel J, Fierer N, Gordon JI, Knight R. Moving pictures of the human microbiome. Genome Biol. 2011;12:R50. doi: 10.1186/gb-2011-12-5-r50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carvalho FA, Koren O, Goodrich JK, Johansson ME, Nalbantoglu I, Aitken JD, Su Y, Chassaing B, Walters WA, González A, Clemente JC, Cullender TC, Barnich N, Darfeuille-Michaud A, Vijay-Kumar M, Knight R, Ley RE, Gewirtz AT. Transient inability to manage proteobacteria promotes chronic gut inflammation in TLR5-deficient mice. Cell Host Microbe. 2012;12:139–152. doi: 10.1016/j.chom.2012.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke G, Grenham S, Scully P, Fitzgerald P, Moloney RD, Shanahan F, Dinan TG, Cryan JF. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry. 2013;18:666–673. doi: 10.1038/mp.2012.77. [DOI] [PubMed] [Google Scholar]
- Collins SM, Kassam Z, Bercik P. The adoptive transfer of behavioral phenotype via the intestinal microbiota: experimental evidence and clinical implications. Curr Opin Microbiol. 2013;16:240–245. doi: 10.1016/j.mib.2013.06.004. [DOI] [PubMed] [Google Scholar]
- Crumeyrolle-Arias M, Jaglin M, Bruneau A, Vancassel S, Cardona A, Daugé V, Naudon L, Rabot S. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology. 2014;42:207–217. doi: 10.1016/j.psyneuen.2014.01.014. [DOI] [PubMed] [Google Scholar]
- Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012;13:701–712. doi: 10.1038/nrn3346. [DOI] [PubMed] [Google Scholar]
- De Palma G, Collins SM, Bercik P, Verdu EF. The microbiota-gut-brain axis in gastrointestinal disorders: stressed bugs, stressed brain or both? J Physiol. 2014;592:2989–2997. doi: 10.1113/jphysiol.2014.273995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desbonnet L, Garrett L, Clarke G, Kiely B, Cryan JF, Dinan TG. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience. 2010;170:1179–1188. doi: 10.1016/j.neuroscience.2010.08.005. [DOI] [PubMed] [Google Scholar]
- Desbonnet L, Clarke G, Shanahan F, Dinan TG, Cryan JF. Microbiota is essential for social development in the mouse. Mol Psychiatry. 2014;19:146–148. doi: 10.1038/mp.2013.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Theije CG, Wu J, Koelink PJ, Korte-Bouws GA, Borre Y, Kas MJ, Lopes da Silva S, Korte SM, Olivier B, Garssen J, Kraneveld AD. Autistic-like behavioural and neurochemical changes in a mouse model of food allergy. Behav Brain Res. 2014;261:265–274. doi: 10.1016/j.bbr.2013.12.008. [DOI] [PubMed] [Google Scholar]
- Diaz Heijtz R, Wang S, Anuar F, Qian Y, Björkholm B, Samuelsson A, Hibberd ML, Forssberg H, Pettersson S. Normal gut microbiota modulates brain development and behavior. Proc Natl Acad Sci U S A. 2011;108:3047–3052. doi: 10.1073/pnas.1010529108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dinan TG, Stanton C, Cryan JF. Psychobiotics: a novel class of psychotropic. Biol Psychiatry. 2013;74:720–726. doi: 10.1016/j.biopsych.2013.05.001. [DOI] [PubMed] [Google Scholar]
- Distrutti E, O'Reilly JA, McDonald C, Cipriani S, Renga B, Lynch MA, Fiorucci S. Modulation of intestinal microbiota by the probiotic VSL#3 resets brain gene expression and ameliorates the age-related deficit in LTP. PLoS One. 2014;9:e106503. doi: 10.1371/journal.pone.0106503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science. 2005;308:1635–1638. doi: 10.1126/science.1110591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gareau MG, Wine E, Rodrigues DM, Cho JH, Whary MT, Philpott DJ, Macqueen G, Sherman PM. Bacterial infection causes stress-induced memory dysfunction in mice. Gut. 2011;60:307–317. doi: 10.1136/gut.2009.202515. [DOI] [PubMed] [Google Scholar]
- Gevers D, Kugathasan S, Denson LA, Vázquez-Baeza Y, Van Treuren W, Ren B, Schwager E, Knights D, Song SJ, Yassour M, Morgan XC, Kostic AD, Luo C, González A, McDonald D, Haberman Y, Walters T, Baker S, Rosh J, Stephens M, et al. The treatment-naive microbiome in new-onset Crohn's disease. Cell Host Microbe. 2014;15:382–392. doi: 10.1016/j.chom.2014.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez A, Knight R. Advancing analytical algorithms and pipelines for billions of microbial sequences. Curr Opin Biotechnol. 2012;23:64–71. doi: 10.1016/j.copbio.2011.11.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haast RA, Kiliaan AJ. Impact of fatty acids on brain circulation, structure and function. Prostaglandins Leukot Essent Fatty Acids. 2014 doi: 10.1016/j.plefa.2014.01.002. doi: 10.1016/j.plefa.2014.01.002. Advance online publication. Retrieved Jul. 18, 2014. [DOI] [PubMed] [Google Scholar]
- Hamady M, Knight R. Microbial community profiling for human microbiome projects: tools, techniques, and challenges. Genome Res. 2009;19:1141–1152. doi: 10.1101/gr.085464.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes E, Kinross J, Gibson GR, Burcelin R, Jia W, Pettersson S, Nicholson JK. Therapeutic modulation of microbiota-host metabolic interactions. Sci Transl Med. 2012;4:137rv136. doi: 10.1126/scitranslmed.3004244. [DOI] [PubMed] [Google Scholar]
- House PK, Vyas A, Sapolsky R. Predator cat odors activate sexual arousal pathways in brains of Toxoplasma gondii infected rats. PLoS One. 2011;6:e23277. doi: 10.1371/journal.pone.0023277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsiao EY, McBride SW, Hsien S, Sharon G, Hyde ER, McCue T, Codelli JA, Chow J, Reisman SE, Petrosino JF, Patterson PH, Mazmanian SK. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 2013;155:1451–1463. doi: 10.1016/j.cell.2013.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486:207–214. doi: 10.1038/nature11234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuczynski J, Lauber CL, Walters WA, Parfrey LW, Clemente JC, Gevers D, Knight R. Experimental and analytical tools for studying the human microbiome. Nat Rev Genet. 2012;13:47–58. doi: 10.1038/nrg3129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauber CL, Zhou N, Gordon JI, Knight R, Fierer N. Effect of storage conditions on the assessment of bacterial community structure in soil and human-associated samples. FEMS Microbiol Lett. 2010;307:80–86. doi: 10.1111/j.1574-6968.2010.01965.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lozupone CA, Stombaugh J, Gonzalez A, Ackermann G, Wendel D, Vázquez-Baeza Y, Jansson JK, Gordon JI, Knight R. Meta-analyses of studies of the human microbiota. Genome Res. 2013;23:1704–1714. doi: 10.1101/gr.151803.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyte M. Microbial endocrinology in the microbiome-gut-brain axis: how bacterial production and utilization of neurochemicals influence behavior. PLoS Pathog. 2013;9:e1003726. doi: 10.1371/journal.ppat.1003726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews DM, Jenks SM. Ingestion of Mycobacterium vaccae decreases anxiety-related behavior and improves learning in mice. Behav Processes. 2013;96:27–35. doi: 10.1016/j.beproc.2013.02.007. [DOI] [PubMed] [Google Scholar]
- Mayer EA. Gut feelings: the emerging biology of gut-brain communication. Nat Rev Neurosci. 2011;12:453–466. doi: 10.1038/nrn3071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayer EA, Berman S, Derbyshire SW, Suyenobu B, Chang L, Fitzgerald L, Mandelkern M, Hamm L, Vogt B, Naliboff BD. The effect of the 5-HT3 receptor antagonist, alosetron, on brain responses to visceral stimulation in irritable bowel syndrome patients. Aliment Pharmacol Ther. 2002;16:1357–1366. doi: 10.1046/j.1365-2036.2002.01287.x. [DOI] [PubMed] [Google Scholar]
- Mayer EA, Padua D, Tillisch K. Altered brain-gut axis in autism: comorbidity or causative mechanisms? Bioessays. 2014a;36:933–939. doi: 10.1002/bies.201400075. [DOI] [PubMed] [Google Scholar]
- Mayer EA, Savidge T, Shulman RJ. Brain-gut microbiome interactions and functional bowel disorders. Gastroenterology. 2014b;146:1500–1512. doi: 10.1053/j.gastro.2014.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald D, Clemente JC, Kuczynski J, Rideout JR, Stombaugh J, Wendel D, Wilke A, Huse S, Hufnagle J, Meyer F, Knight R, Caporaso JG. The Biological Observation Matrix (BIOM) format or: how I learned to stop worrying and love the ome-ome. Gigascience. 2012;1:7. doi: 10.1186/2047-217X-1-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKernan DP, Fitzgerald P, Dinan TG, Cryan JF. The probiotic Bifidobacterium infantis 35624 displays visceral antinociceptive effects in the rat. Neurogastroenterol Motil. 2010;22:1029–1035. doi: 10.1111/j.1365-2982.2010.01520.x. [DOI] [PubMed] [Google Scholar]
- McNulty NP, Yatsunenko T, Hsiao A, Faith JJ, Muegge BD, Goodman AL, Henrissat B, Oozeer R, Cools-Portier S, Gobert G, Chervaux C, Knights D, Lozupone CA, Knight R, Duncan AE, Bain JR, Muehlbauer MJ, Newgard CB, Heath AC, Gordon JI. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci Transl Med. 2011;3:106ra106. doi: 10.1126/scitranslmed.3002701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messaoudi M, Lalonde R, Violle N, Javelot H, Desor D, Nejdi A, Bisson JF, Rougeot C, Pichelin M, Cazaubiel M, Cazaubiel JM. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr. 2011;105:755–764. doi: 10.1017/S0007114510004319. [DOI] [PubMed] [Google Scholar]
- Moayyedi P, Ford AC, Talley NJ, Cremonini F, Foxx-Orenstein AE, Brandt LJ, Quigley EM. The efficacy of probiotics in the treatment of irritable bowel syndrome: a systematic review. Gut. 2010;59:325–332. doi: 10.1136/gut.2008.167270. [DOI] [PubMed] [Google Scholar]
- Moloney RD, Desbonnet L, Clarke G, Dinan TG, Cryan JF. The microbiome: stress, health and disease. Mamm Genome. 2014;25:49–74. doi: 10.1007/s00335-013-9488-5. [DOI] [PubMed] [Google Scholar]
- Navas-Molina JA, Peralta-Sánchez JM, González A, McMurdie PJ, Vázquez-Baeza Y, Xu Z, Ursell LK, Lauber C, Zhou H, Song SJ, Huntley J, Ackermann GL, Berg-Lyons D, Holmes S, Caporaso JG, Knight R. Advancing our understanding of the human microbiome using QIIME. Methods Enzymol. 2013;531:371–444. doi: 10.1016/B978-0-12-407863-5.00019-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neufeld KM, Kang N, Bienenstock J, Foster JA. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol Motil. 2011;23:255–264. doi: 10.1111/j.1365-2982.2010.01620.x. [DOI] [PubMed] [Google Scholar]
- Nishino R, Mikami K, Takahashi H, Tomonaga S, Furuse M, Hiramoto T, Aiba Y, Koga Y, Sudo N. Commensal microbiota modulate murine behaviors in a strictly contamination-free environment confirmed by culture-based methods. Neurogastroenterol Motil. 2013;25:521–528. doi: 10.1111/nmo.12110. [DOI] [PubMed] [Google Scholar]
- Ohland CL, Kish L, Bell H, Thiesen A, Hotte N, Pankiv E, Madsen KL. Effects of Lactobacillus helveticus on murine behavior are dependent on diet and genotype and correlate with alterations in the gut microbiome. Psychoneuroendocrinology. 2013;38:1738–1747. doi: 10.1016/j.psyneuen.2013.02.008. [DOI] [PubMed] [Google Scholar]
- O'Mahony SM, Felice VD, Nally K, Savignac HM, Claesson MJ, Scully P, Woznicki J, Hyland NP, Shanahan F, Quigley EM, Marchesi JR, O'Toole PW, Dinan TG, Cryan JF. Disturbance of the gut microbiota in early-life selectively affects visceral pain in adulthood without impacting cognitive or anxiety-related behaviors in male rats. Neuroscience. 2014;277:885–901. doi: 10.1016/j.neuroscience.2014.07.054. [DOI] [PubMed] [Google Scholar]
- Queipo-Ortuño MI, Seoane LM, Murri M, Pardo M, Gomez-Zumaquero JM, Cardona F, Casanueva F, Tinahones FJ. Gut microbiota composition in male rat models under different nutritional status and physical activity and its association with serum leptin and ghrelin levels. PLoS One. 2013;8:e65465. doi: 10.1371/journal.pone.0065465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao AV, Bested AC, Beaulne TM, Katzman MA, Iorio C, Berardi JM, Logan AC. A randomized, double-blind, placebo-controlled pilot study of a probiotic in emotional symptoms of chronic fatigue syndrome. Gut Pathog. 2009;1:6. doi: 10.1186/1757-4749-1-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravussin Y, Koren O, Spor A, LeDuc C, Gutman R, Stombaugh J, Knight R, Ley RE, Leibel RL. Responses of gut microbiota to diet composition and weight loss in lean and obese mice. Obesity (Silver Spring) 2012;20:738–747. doi: 10.1038/oby.2011.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rousseaux C, Thuru X, Gelot A, Barnich N, Neut C, Dubuquoy L, Dubuquoy C, Merour E, Geboes K, Chamaillard M, Ouwehand A, Leyer G, Carcano D, Colombel JF, Ardid D, Desreumaux P. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat Med. 2007;13:35–37. doi: 10.1038/nm1521. [DOI] [PubMed] [Google Scholar]
- Sanders ME, Guarner F, Guerrant R, Holt PR, Quigley EM, Sartor RB, Sherman PM, Mayer EA. An update on the use and investigation of probiotics in health and disease. Gut. 2013;62:787–796. doi: 10.1136/gutjnl-2012-302504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savignac HM, Corona G, Mills H, Chen L, Spencer JP, Tzortzis G, Burnet PW. Prebiotic feeding elevates central brain derived neurotrophic factor, N- methyl-d-aspartate receptor subunits and d-serine. Neurochem Int. 2013;63:756–764. doi: 10.1016/j.neuint.2013.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sternbach H, State R. Antibiotics: neuropsychiatric effects and psychotropic interactions. Harv Rev Psychiatry. 1997;5:214–226. doi: 10.3109/10673229709000304. [DOI] [PubMed] [Google Scholar]
- Stilling RM, Dinan TG, Cryan JF. Microbial genes, brain and behavior: epigenetic regulation of the gut-brain axis. Genes Brain Behav. 2014a;13:69–86. doi: 10.1111/gbb.12109. [DOI] [PubMed] [Google Scholar]
- Stilling RM, Bordenstein SR, Dinan TG, Cryan JF. Friends with social benefits: host-microbe interactions as a driver of brain evolution and development? Front Cell Infect Microbiol. 2014b doi: 10.3389/fcimb.2014.00147. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu XN, Kubo C, Koga Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol. 2004;558:263–275. doi: 10.1113/jphysiol.2004.063388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tillisch K, Labus JS. Neuroimaging the microbiome-gut-brain axis. Adv Exp Med Biol. 2014;817:405–416. doi: 10.1007/978-1-4939-0897-4_18. [DOI] [PubMed] [Google Scholar]
- Tillisch K, Wang Z, Kilpatrick L, Holschneider DP, Mayer EA. Studying the brain-gut axis with pharmacological imaging. Ann N Y Acad Sci. 2008;1144:256–264. doi: 10.1196/annals.1418.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tillisch K, Labus J, Kilpatrick L, Jiang Z, Stains J, Ebrat B, Guyonnet D, Legrain-Raspaud S, Trotin B, Naliboff B, Mayer EA. Consumption of fermented milk product with probiotic modulates brain activity. Gastroenterology. 2013;144:1394–1401. doi: 10.1053/j.gastro.2013.02.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomé AM, Filipe A. Quinolones: review of psychiatric and neurological adverse reactions. Drug Saf. 2011;34:465–488. doi: 10.2165/11587280-000000000-00000. [DOI] [PubMed] [Google Scholar]
- Vázquez-Baeza Y, Pirrung M, Gonzalez A, Knight R. EMPeror: a tool for visualizing high-throughput microbial community data. Gigascience. 2013;2:16. doi: 10.1186/2047-217X-2-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S, Sitaraman SV, Knight R, Ley RE, Gewirtz AT. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science. 2010;328:228–231. doi: 10.1126/science.1179721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wall R, Cryan JF, Ross RP, Fitzgerald GF, Dinan TG, Stanton C. Bacterial neuroactive compounds produced by psychobiotics. Adv Exp Med Biol. 2014;817:221–239. doi: 10.1007/978-1-4939-0897-4_10. [DOI] [PubMed] [Google Scholar]
- Wise RG, Tracey I. The role of fMRI in drug discovery. J Magn Reson Imaging. 2006;23:862–876. doi: 10.1002/jmri.20584. [DOI] [PubMed] [Google Scholar]
- Wu GD, Lewis JD, Hoffmann C, Chen YY, Knight R, Bittinger K, Hwang J, Chen J, Berkowsky R, Nessel L, Li H, Bushman FD. Sampling and pyrosequencing methods for characterizing bacterial communities in the human gut using 16S sequence tags. BMC Microbiol. 2010;10:206. doi: 10.1186/1471-2180-10-206. [DOI] [PMC free article] [PubMed] [Google Scholar]