

Abstract
Background
Shigella flexneri is an important human pathogen that has to adapt to the anaerobic environment in the gastrointestinal tract to cause dysentery. To define the influence of anaerobiosis on the virulence of Shigella, we performed deep RNA sequencing to identify transcriptomic differences that are induced by anaerobiosis and modulated by the anaerobic Fumarate and Nitrate Reduction regulator, FNR.
Results
We found that 528 chromosomal genes were differentially expressed in response to anaerobic conditions; of these, 228 genes were also influenced by FNR. Genes that were up-regulated in anaerobic conditions are involved in carbon transport and metabolism (e.g. ptsG, manX, murQ, cysP, cra), DNA topology and regulation (e.g. ygiP, stpA, hns), host interactions (e.g. yciD, nmpC, slyB, gapA, shf, msbB) and survival within the gastrointestinal tract (e.g. shiA, ospI, adiY, cysP). Interestingly, there was a marked effect of available oxygen on genes involved in Type III secretion system (T3SS), which is required for host cell invasion and pathogenesis. These genes, located on the large Shigella virulence plasmid, were down regulated in anaerobiosis in an FNR-dependent manner. We also confirmed anaerobic induction of csrB and csrC small RNAs in an FNR-independent manner.
Conclusions
Anaerobiosis promotes survival and adaption strategies of Shigella, while modulating virulence plasmid genes involved in T3SS-mediated host cell invasion. The influence of FNR on this process is more extensive than previously appreciated, although aside from the virulence plasmid, this transcriptional regulator does not govern expression of genes on other horizontally acquired sequences on the chromosome such as pathogenicity islands.
Electronic supplementary material
The online version of this article (doi:10.1186/1471-2164-15-438) contains supplementary material, which is available to authorized users.
Background
Shigella flexneri is a Gram-negative bacterium that causes dysentery, an acute human rectocolitis that usually results in destruction of the intestinal mucosa and bloody diarrhoea. The ability of this pathogen to invade epithelial cells at the colonic and rectal mucosal surface is a key determinant in the establishment of the disease. This is mediated by a Type III secretion system (T3SS) encoded on the large Shigella virulence plasmid [1, 2]. The T3SS acts like a molecular syringe that delivers molecules directly from the bacterial cytoplasm into host cells via a needle-like structure [1, 2]. However, before the bacterium reaches the large intestine and invades mucosal epithelial cells, Shigella must successfully survive the hostile conditions found in the gastrointestinal tract. Therefore the capacity of the bacterium to adapt to anaerobiosis, changes in pH, resist antimicrobial peptides, and acquire nutrients is essential for its pathogenesis [3, 4].
Anaerobiosis is known to influence the virulence of several enteric pathogens including Shigella, Escherichia coli, Salmonella spp., Vibrio cholerae and Yersinia enterocolitica[5–13]. In particular, S. flexneri has been shown to be primed for invasion in anaerobic conditions, in which it expresses longer T3SS needles while reducing Ipa (invasion plasmid antigen) effector secretion; this results from FNR-mediated repression of the virulence plasmid genes, spa32 and spa33[7]. FNR is a major regulator of anaerobic metabolism that is inactivated by the presence of oxygen. Its function depends on the integrity of its O2-sensitive [4Fe–4S] cluster, which is required for FNR dimerization and thence site-specific DNA binding and transcriptional regulation [14]. One RNA deep sequencing (RNA-seq) and several microarray studies have been performed to characterise the extent of the FNR regulon in E. coli and other Gram negative pathogens such as Salmonella enterica and Neisseria gonorrhoeae[15–20]. In E. coli, there were significant discrepancies between studies even when the same strain was examined. However some differences could be attributed to the use of media containing high levels of glucose, which represses expression from some FNR-activated promoters, and the delayed growth rate of mutants lacking FNR compared with wild-type strains under anaerobic conditions [16].
Here we define the regulatory role of oxygen and FNR in S. flexneri. We have applied two powerful whole-transcriptome approaches, RNA-seq complemented with Flow cell Reverse Transcription sequencing (FRT-seq), in which there is no amplification during library preparation, to quantify differences in gene expression induced by anaerobiosis and to define the contribution of FNR in this process. We found that Shigella grown anaerobically exhibits global transcriptional changes compared to when grown aerobically, with marked changes in metabolic and transport genes, as well as those involved in regulatory and virulence functions. Importantly, transcription from the Shigella virulence plasmid is extensively modified in anaerobiosis, with most of T3SS-related genes being down regulated in the absence of oxygen in an FNR-dependent manner, demonstrating that this highly conserved regulator of metabolism also controls the horizontally-acquired virulence genes on the plasmid, but not on the chromosome, in this important human pathogen.
Results
Growth conditions and RNA sequencing strategies
To determine the response of Shigella to anaerobiosis and the role of FNR in this process, we employed RNA-seq to compare the transcriptional profiles of wild type S. flexneri M90T and its Δfnr mutant grown in Luria-Bertani (LB) medium in the presence and absence of oxygen. Constantinidou et al. designed a supplemented, minimal salts medium (including LB) in which an E. coli fnr mutant exhibited similar growth as the parental strain in the absence of oxygen [16]. However, this medium did not support the growth of S. flexneri M90T. On the other hand, enriched-glucose media have been shown to repress some FNR-activated promoters [16]. Therefore, we chose LB with no added glucose for our experiments. Particular attention was paid to ensure that the culture volume, agitation, temperature and the growth stage of bacteria did not differ in aerobic and anaerobic conditions. Cultures were grown to an Optical Density at 600 nm (OD600) of 0.2 to avoid a reduction in the concentration of dissolved oxygen tension and total depletion of sugars that occurs during exponential growth [21, 22]. Furthermore until reach OD600 of 0.2 under anaerobiosis, there was no obvious delay in growth rate of the Δfnr mutant in relation to the wild-type strain (See Additional file 1: Figure S1). Three biological replicates were performed per strain in each condition, and differential expression between conditions was analysed with the DESeq R statistical package.
To assess the reproducibility of results obtained with RNA-seq data and to further characterise the role of FNR, the Shigella FNR regulon under anaerobiosis was also examined using FRT-seq, an alternative sequencing approach in which cDNA synthesis is performed on the sequencing flowcell thereby avoiding the possible PCR biases generated during library preparation using standard RNA-seq methods [23]. FRT-seq confirmed 77% of the genes found differentially expressed by RNA-seq, showing a robust concordance between the two techniques. Due to its higher sensitivity, FRT-seq detected more genes whose transcription was significantly influenced by the absence of FNR than RNA-seq (See Additional file 1: Table S2). A complete catalogue of significant differences is shown in Additional material (See Additional file 1: Tables S1 and S2) as well as a summary of the mapping statistics (See Additional file 1: Table S3). To confirm the results obtained by global analysis of the transcriptional profile, we performed strand-specific qRT-PCR to analyse mRNA levels of several genes found to be differentially expressed under anaerobic and aerobic growth conditions.
Identification of novel chromosomal genes influenced by the absence of oxygen in S. flexneri
Analysis of the RNA-seq data revealed that 528 chromosomal genes were differentially expressed by wild-type S. flexneri M90T grown under anaerobic conditions compared with aerobic conditions, with 363 genes being up-regulated, and 165 genes down-regulated. Additional file 1: Table S1 shows these genes classified into functional categories based on the database of Clusters of Orthologous Groups (COGs) [24]. As expected, most of the genes differentially expressed were related to energy production and metabolism (53%). The remaining genes were involved in cellular processes and signalling (15%), information storage and processing (8%) or were poorly characterized (24%). RNA-seq data also showed that from the above 528 differentially expressed genes, 228 genes (43%) were influenced by the absence of FNR under anaerobic conditions (See Additional file 1: Table S1).
Importantly the majority of genes that we found to be anaerobically induced/repressed have been identified in previous microarray studies with other enteric pathogens examining the effect of oxygen on the transcriptome and/or the two main anaerobic regulators, FNR and ArcA [6, 16–20, 25]. Consistent with previous work, we found increased expression of genes involved in anoxic carbon metabolism (focA-pfl, yfiD, fdnG, gldA, aspA, fumB, ansB), respiratory pathways (glpABC, nap, nir, ccm, nrfABC, frd), production of hydrogenases (hyb, hya, hyc, hyp), fermentation (adhE, ackA-pta, fdhF) and acid response (adiA, adiY, yjdE, gadA, hdeAB) under anaerobiosis (Additional file 1: Table S1) [6, 16–18, 20, 25, 26]. Our analysis also identified several anaerobically repressed genes that have been previously characterised [6, 16–20, 25]. These genes encode enzymes of the tricarboxylic acid cycle (ace, gltA, acn, icdA, sdh), aerobic dehydrogenases (glpD, betBA, gcd, aldA), transhydrogenases (udhA) and iron acquisition systems (exb, iuc, iutA, sit, suf, fep, fhu), and others (Additional file 1: Table S1) [6, 16–18, 20, 25, 27, 28].
The sensitivity of the direct sequencing approaches, RNA-seq and FRT-seq, compared with array-based methods enabled us to extend the repertoire of Shigella genes modulated by ambient oxygen. Table 1 shows all genes influenced by the presence of oxygen and not detected in previous microarray studies on E. coli and S. flexneri[6, 16–18, 20, 25]. The effect of FNR mutation on the transcription of previous genes under anaerobiosis (assessed by RNA-seq and FRT-seq) is also shown in Table 1. Several members of the phosphoenolpyruvate–carbohydrate phosphotransferase system (PTS), involved in the transport and phosphorylation of sugars, were up-regulated under anaerobic conditions. Examples include ptsHI, which encode the general PTS components phosphohistidine carrier protein (HPr) and Enzyme I (EI) respectively, and sugar-specific PTS components like ptsG and manXYZ (involved in glucose transport), treBC (trehalose transport and hydrolysis), mtlA (mannitol) and murQP that contribute to the uptake and catabolism of N-acetylmuramic acid [29–32]. Of note, the murQP operon, which is also involved in peptidoglycan recycling, showed an FNR-dependent expression pattern (Table 1, Figure 1A) [31].
Table 1.
Chromosomal genes differentially expressed in response to anaerobic conditions not previously published in E. coli and S. flexneri microarray analysis
| ORF ID ab | Gene | Description | RNA-seq c log2FC | RNA-seq c log2FC | FRT-seq c log2FC |
|---|---|---|---|---|---|
| WT no O 2 /O 2 | Δ fnr /WT no O 2 | Δ fnr /WT no O 2 | |||
| Metabolism | |||||
| Energy production and conversion | |||||
| SF5M90T_1519 | putative oxidoreductase, major subunit | 3.80 | -4.97 | -3.30 | |
| SF5M90T_3856 | yiaY | putative oxidoreductase | 3.18 | -2.73 | -2.67 |
| SF5M90T_1560 | putative oxidoreductase, major subunit | 3.06 | -3.87 | ||
| SF5M90T_3333 | pckA | phosphoenolpyruvate carboxykinase | 1.99 | 0.62 | |
| SF5M90T_3877 | yiaK | putative dehydrogenase | 1.93 | 1.22 | 0.93 |
| SF5M90T_3374 | ugpQ | glycerophosphodiester phosphodiesterase, cytosolic | 1.89 | ||
| SF5M90T_2534 | hmpA | dihydropteridine reductase, ferrisiderophore reductase activity | 1.47 | 5.38 | 5.85 |
| SF5M90T_33 | caiB | l-carnitine dehydratase | 1.32 | ||
| SF5M90T_3679 | atpF | membrane-bound ATP synthase, F0 sector, subunit b | 1.04 | ||
| SF5M90T_3680 | atpE | membrane-bound ATP synthase, F0 sector, subunit c | 1.04 | ||
| SF5M90T_3937 | ppc | phosphoenolpyruvate carboxylase | 0.91 | 0.64 | |
| SF5M90T_579 | galT | galactose-1-phosphate uridylyltransferase | 0.77 | ||
| SF5M90T_1419 | ydjA | predicted oxidoreductase | -1.17 | 1.69 | 1.52 |
| SF5M90T_2771 | ygaF | hydroxyglutarate oxidase | -1.31 | 4.12 | 2.99 |
| SF5M90T_4044 | gltP | glutamate-aspartate symport protein | -1.32 | ||
| SF5M90T_1603 | rnfB | electron transport complex protein | -1.46 | ||
| SF5M90T_2869 | fldB | flavodoxin 2 | -1.56 | ||
| SF5M90T_1602 | rnfA | Na + -translocating NADH-quinone reductase subunit E | -1.75 | ||
| SF5M90T_1011 | rutA | pyrimidine monooxygenase | -3.07 | ||
| Carbohydrate transport and metabolism | |||||
| SF4250 | treB | PTS system trehalose(maltose)-specific transporter subunits IIBC | 3.66 | ||
| SF5M90T_4160 | treC | trehalase 6-P hydrolase | 3.56 | ||
| SF5M90T_1379 | manX | PTS enzyme IIAB, mannose-specific | 3.36 | ||
| SF5M90T_1378 | manY | PTS enzyme IIC, mannose-specific | 3.11 | ||
| SF5M90T_1377 | manZ | PTS enzyme IID, mannose-specific | 2.89 | ||
| SF5M90T_3670 | rbsD | high affinity ribose transport protein | 2.71 | ||
| SF5M90T_1101 | ptsG | PTS system, glucose-specific IIBC component | 2.27 | ||
| SF5M90T_3491 | treF | cytoplasmic trehalase | 2.12 | ||
| SF5M90T_2419 | murP | PTS system N-acetylmuramic acid transporter subunits EIIBC | 2.09 | -0.71 | |
| SF5M90T_3499 | pfkA | 6-phosphofructokinase I | 2.08 | ||
| SF5M90T_2096 | fructose-bisphosphate aldolase | 2.02 | 2.08 | 1.86 | |
| SF5M90T_1001 | agp | periplasmic glucose-1-phosphatase | 2.00 | 1.40 | 1.57 |
| SF5M90T_2887 | rpiA | ribosephosphate isomerase, constitutive | 1.84 | ||
| SF5M90T_2898 | pgk | phosphoglycerate kinase | 1.84 | ||
| SF5M90T_1403 | gapA | glyceraldehyde-3-phosphate dehydrogenase A | 1.83 | ||
| SF5M90T_2097 | yegT | putative nucleoside permease protein | 1.74 | ||
| SF5M90T_2897 | fba | fructose-bisphosphate aldolase, class II | 1.56 | ||
| SF5M90T_2404 | ptsH | PTS system protein HPr | 1.56 | ||
| SF5M90T_3850 | mtlA | PTS system, mannitol-specific enzyme IIABC components | 1.52 | ||
| SF5M90T_1640 | ydhC | putative transport protein | 1.51 | 0.69 | |
| SF5M90T_2359 | beta-fructosidase | 1.49 | 0.72 | ||
| SF5M90T_3496 | tpiA | triosephosphate isomerase | 1.45 | ||
| SF5M90T_2405 | ptsI | PEP-protein phosphotransferase system enzyme I | 1.42 | ||
| SF5M90T_2808 | fucI | L-fucose isomerase | 1.41 | ||
| SF5M90T_2875 | bglA | 6-phospho-beta-glucosidase A | 1.27 | ||
| SF5M90T_3348 | malP | maltodextrin phosphorylase | 1.16 | 0.78 | |
| SF5M90T_1107 | ycfO | beta-hexosaminidase | 1.13 | ||
| SF5M90T_8 | talB | transaldolase B | 1.10 | ||
| SF5M90T_2033 | gnd | gluconate-6-phosphate dehydrogenase | 1.01 | ||
| SF5M90T_581 | galM | galactose-1-epimerase | 1.01 | 1.41 | 1.00 |
| SF5M90T_1805 | eda | keto-hydroxyglutarate-aldolase/keto-deoxy-phosphogluconate aldolase | 0.97 | ||
| SF5M90T_580 | galK | galactokinase | 0.95 | 0.62 | |
| SF5M90T_2913 | tktA | transketolase 1 isozyme | 0.80 | ||
| SF5M90T_2187 | fruB | PTS system fructose-specific transporter subunit IIA/HPr protein | -1.33 | ||
| SF5M90T_2186 | fruK | fructose-1-phosphate kinase | -1.75 | ||
| SF5M90T_3161 | ptsO | phosphocarrier protein NPr | -1.76 | ||
| SF5M90T_1637 | putative transport protein | -1.93 | 1.58 | ||
| SF5M90T_2185 | fruA | PTS system, fructose-specific transport protein | -1.99 | ||
| Aminoacid transport and metabolism | |||||
| SF5M90T_2823 | argA | N-acetylglutamate synthase | 1.94 | ||
| SF5M90T_1910 | fliY | putative periplasmic binding transport protein | 1.80 | ||
| SF5M90T_625 | ybgH | peptide transporter | 1.64 | -1.60 | -1.21 |
| SF5M90T_292 | pepD | aminoacyl-histidine dipeptidase (peptidase D) | 1.54 | 1.78 | 1.75 |
| SF5M90T_2879 | gcvT | aminomethyltransferase | 1.53 | 1.44 | |
| SF5M90T_284 | proA | gamma-glutamylphosphate reductase | 1.48 | ||
| SF5M90T_1121 | potD | spermidine/putrescine periplasmic transport protein | 1.44 | -0.69 | |
| SF5M90T_2674 | cysD | ATP:sulfurylase (ATP:sulfate adenylyltransferase), subunit 2 | 1.39 | -2.30 | -2.21 |
| SF5M90T_1514 | dcp | dipeptidyl carboxypeptidase II | 1.35 | 0.63 | |
| SF5M90T_285 | proB | gamma-glutamate kinase | 1.26 | -0.58 | |
| SF5M90T_2533 | glyA | serine hydroxymethyltransferase | 1.16 | ||
| SF5M90T_2967 | gsp | glutathionylspermidine synthetase/amidase | 1.16 | 0.70 | |
| SF5M90T_1806 | edd | 6-phosphogluconate dehydratase | 1.15 | -0.72 | |
| SF5M90T_807 | glutathione transporter ATP-binding protein | 1.08 | |||
| SF5M90T_2317 | hisJ | histidine-binding periplasmic protein of high-affinity histidine transport system | 1.05 | ||
| SF5M90T_806 | ybiK | putative asparaginase | 1.02 | ||
| SF5M90T_1122 | potC | spermidine/putrescine transport system permease | 0.97 | -1.09 | -0.92 |
| SF5M90T_2882 | pepP | proline aminopeptidase P II | 0.94 | ||
| SF5M90T_2877 | gcvP | glycine decarboxylase | 0.90 | 2.13 | 1.69 |
| SF5M90T_3687 | asnA | asparagine synthetase A | -1.23 | -1.61 | |
| SF5M90T_4099 | lysC | aspartokinase III, lysine sensitive | -1.33 | 1.37 | 1.14 |
| SF5M90T_1253 | trpE | anthranilate synthase component I | -1.57 | ||
| SF5M90T_4187 | cycA | transport of D-alanine, D-serine, and glycine | -1.69 | ||
| SF5M90T_1946 | yedA | putative transmembrane subunit | -1.79 | ||
| SF5M90T_3626 | yifK | putative amino acid/amine transport protein | -1.94 | ||
| SF5M90T_3385 | livJ | Leu/Ile/Val-binding protein precursor | -1.95 | ||
| SF5M90T_2843 | lysA | diaminopimelate decarboxylase | -2.11 | 2.44 | |
| SF5M90T_4029 | proP | low-affinity transport system; proline permease II | -2.65 | 1.05 | 0.75 |
| SF5M90T_4185 | ytfF | putative transmembrane subunit | -3.70 | 2.71 | |
| Nucleotide transport and metabolism | |||||
| SF5M90T_3587 | udp | uridine phosphorylase | 3.12 | 0.59 | |
| SF5M90T_2387 | nupC | permease of transport system for 3 nucleosides | 2.81 | ||
| SF5M90T_2949 | nupG | nucleoside permease | 2.47 | ||
| SF5M90T_674 | ybeK | putative tRNA synthetase | 1.60 | ||
| SF5M90T_444 | adk | adenylate kinase | 1.51 | -0.67 | |
| SF5M90T_2456 | purC | phosphoribosylaminoimidazole-succinocarboxamidesynthetase | 1.35 | ||
| SF5M90T_4182 | cpdB | 2′:3′-cyclic-nucleotide 2′-phosphodiesterase | 1.16 | ||
| SF5M90T_291 | gpt | guanine-hypoxanthine phosphoribosyltransferase | 1.10 | -0.89 | |
| SF5M90T_1598 | add | adenosine deaminase | 0.95 | ||
| SF5M90T_478 | purE | phosphoribosylaminoimidazole carboxylase | -1.41 | ||
| Coenzyme transport and metabolism | |||||
| SF5M90T_2274 | menB | dihydroxynaphtoic acid synthetase | 2.63 | -1.75 | -1.65 |
| SF5M90T_2687 | phenylacrylic acid decarboxylase-like protein | 1.83 | |||
| SF5M90T_2276 | menD | 2-oxoglutarate decarboxylase | 1.76 | -1.63 | -1.75 |
| SF5M90T_2273 | menC | O-succinylbenzoate synthase | 1.76 | -1.72 | -1.57 |
| SF5M90T_3142 | ispB | octaprenyl diphosphate synthase | 1.06 | ||
| SF5M90T_1613 | pdxH | pyridoxinephosphate oxidase | 1.06 | ||
| SF5M90T_2880 | visC | putative FAD-dependent oxidoreductase | 0.89 | ||
| SF5M90T_3011 | ribB | 3,4 dihydroxy-2-butanone-4-phosphate synthase | -1.10 | ||
| SF5M90T_3577 | yigC | putative oxidoreductase | -1.31 | ||
| SF5M90T_2885 | ygfA | putative ligase | -1.59 | ||
| SF5M90T_3957 | birA | biotin--protein ligase | -1.62 | -0.51 | |
| SF5M90T_2103 | thiM | hydoxyethylthiazole kinase | -1.95 | ||
| Lipid transport and metabolism | |||||
| SF5M90T_1094 | acpP | acyl carrier protein | 1.70 | -3.04 | |
| SF5M90T_2272 | menE | o-succinylbenzoate-CoA ligase | 1.60 | -1.54 | -1.80 |
| SF5M90T_2416 | ucpA | putative oxidoreductase | 1.45 | -0.56 | |
| SF5M90T_339 | sbmA | sensitivity to microcin B17, possibly envelope protein | -1.64 | ||
| Inorganic ion transport and metabolism | |||||
| SF5M90T_2903 | hypothetical lipoprotein | 3.41 | |||
| SF5M90T_929 | ycbO | alkanesulfonate transporter substrate-binding subunit | 3.04 | ||
| SF5M90T_2415 | cysP | thiosulfate binding protein | 2.81 | -1.51 | -1.88 |
| SF5M90T_1636 | sodB | superoxide dismutase | 2.52 | 2.13 | 1.67 |
| SF5M90T_1187 | putative ATP-binding protein of ABC transporter | 2.14 | |||
| SF5M90T_454 | copA | copper exporting ATPase | 1.95 | ||
| SF5M90T_1186 | putative iron compound ABC transporter permease | 1.69 | |||
| SF5M90T_1185 | iron ABC transporter ATP-binding protein | 1.52 | |||
| SF5M90T_4057 | yjcE | predicted cation/proton antiporter | 1.49 | ||
| SF5M90T_2675 | cysN | ATP-sulfurylase (ATP:sulfate adenylyltransferase), subunit 1 | 1.08 | -2.08 | -2.05 |
| SF5M90T_448 | ybaL | putative transport protein | -1.01 | -0.50 | |
| SF5M90T_2386 | mntH | divalent metal cation transporter | -1.93 | 1.61 | 1.68 |
| SF5M90T_330 | tauC | taurine transport system permease protein | -2.09 | ||
| SF5M90T_3769 | shiF | putative membrane transport protein | -2.16 | ||
| SF5M90T_3054 | ygjT | putative transport protein | -2.41 | ||
| SF5M90T_1102 | fhuE | outer membrane receptor for ferric iron uptake | -2.46 | ||
| SF5M90T_1483 | ydiE | hemin uptake protein | -2.93 | -2.20 | -1.55 |
| SF5M90T_1572 | mdtI | spermidine export protein | -3.61 | ||
| Secondary metabolites biosynthesis, transport and catabolism | |||||
| SF5M90T_1184 | putative SAM-dependent methyltransferase | 2.12 | |||
| SF5M90T_331 | tauD | taurine dioxygenase, 2-oxoglutarate-dependent | -2.97 | 1.94 | |
| Cellular processes and signalling | |||||
| Cell cycle control, cell division, chromosome partitioning | |||||
| SF5M90T_1243 | yciB | probable intracellular septation protein A | 0.93 | -0.80 | |
| Defense mechanisms | |||||
| SF5M90T_4215 | ampC | beta-lactamase; penicillin resistance | 1.55 | -1.69 | |
| SF5M90T_3751 | emrD | multidrug resistance protein D | 1.41 | ||
| SF5M90T_4273 | putative restriction modification enzyme R subunit | 1.41 | -0.94 | -0.99 | |
| SF5M90T_3781 | shiA | virulence factor | 1.30 | ||
| SF5M90T_101 | ampD | N-acetyl-anhydromuranmyl-L-alanine amidase | 1.18 | ||
| SF5M90T_772 | ybhF | putative ABC-type multidrug transport system component | 1.16 | 0.54 | |
| SF5M90T_771 | ybhS | putative ABC-type multidrug transport system component | 1.15 | ||
| SF5M90T_770 | ybhR | putative ABC-type multidrug transport system component | 0.90 | ||
| SF5M90T_418 | mdlA | ATP-binding component of a transport system | -1.29 | ||
| Signal transduction mechanisms | |||||
| SF5M90T_2126 | yehU | putative 2-component sensor protein | 1.36 | ||
| SF5M90T_3428 | uspA | universal stress protein | 0.86 | ||
| SF5M90T_2388 | yfeA | predicted diguanylate cyclase | -1.20 | -0.81 | |
| SF5M90T_4339 | creC | sensory histidine kinase | -1.63 | 1.43 | |
| Cell wall/membrane/envelope biogenesis | |||||
| SF5M90T_1923 | nmpC | outer membrane porin protein | 2.04 | -1.34 | -1.13 |
| SF5M90T_1618 | slyB | putative outer membrane protein | 1.82 | -1.43 | -0.92 |
| SF5M90T_952 | ompA | outer membrane protein 3a | 1.59 | ||
| SF5M90T_374 | tsx | outer membrane protein | 1.53 | ||
| SF5M90T_256 | gtrB | bactoprenol glucosyl transferase | 1.36 | -2.59 | |
| SF5M90T_2039 | rfbC | dTDP-4-dehydrorhamnose 3,5-epimerase | 1.20 | -2.64 | |
| SF5M90T_4332 | slt | soluble lytic murein transglycosylase | 0.96 | 1.54 | 1.46 |
| SF5M90T_3951 | murI | glutamate racemase | 0.93 | -0.48 | |
| SF5M90T_3821 | rfaD | ADP-L-glycero-D-mannoheptose-6-epimerase | 0.82 | ||
| SF5M90T_1241 | tonB | transport protein | -1.72 | ||
| SF5M90T_3956 | murB | UDP-N-acetylenolpyruvoylglucosamine reductase | -2.23 | -0.60 | |
| Cell motility | |||||
| SF5M90T_1938 | fliQ | flagellar biosynthetic protein | 3.55 | ||
| Intracellular trafficking, secretion and vesicular transport | |||||
| SF5M90T_3964 | secE | preprotein translocase | 0.87 | -1.06 | -0.76 |
| SF5M90T_3580 | tatC | Sec-independent protein translocase | 0.84 | -0.57 | |
| SF5M90T_3501 | yiiO | uncharacterized periplasmic protein | -4.72 | 2.64 | |
| Posttranslational modification, protein turnover, chaperones | |||||
| SF5M90T_4204 | mopB | co-chaperonin GroES | 1.41 | ||
| SF5M90T_3279 | slyD | FKBP-type peptidyl-prolyl cis-trans isomerase | 1.31 | ||
| SF5M90T_462 | ybbN | putative thioredoxin-like protein | 0.90 | ||
| SF5M90T_407 | clpP | ATP-dependent proteolytic subunit of clpA-clpP serine protease | 0.84 | ||
| SF5M90T_3738 | ibpA | heat shock protein | -1.17 | ||
| SF5M90T_2074 | yegD | putative heat shock protein | -2.80 | -1.11 | |
| Information storage and processing | |||||
| Translation, ribosomal structure and biogenesis | |||||
| SF5M90T_2801 | yfiA | translation inhibitor protein RaiA | 2.70 | ||
| SF5M90T_2392 | gltX | glutamate tRNA synthetase, catalytic subunit | 1.65 | ||
| SF5M90T_155 | frr | ribosome releasing factor | 1.21 | ||
| SF5M90T_650 | glnS | glutamine tRNA synthetase | 1.08 | ||
| SF5M90T_3893 | glyQ | glycine tRNA synthetase, alpha subunit | 1.06 | -0.50 | |
| SF5M90T_4220 | yjeA | putative lysyl-tRNA synthetase | 0.95 | -1.75 | -1.72 |
| SF5M90T_3894 | glyS | glycine tRNA synthetase, beta subunit | 0.81 | ||
| Transcription | |||||
| SF5M90T_3025 | ygiP | putative transcriptional regulator/nucleoid-associated protein | 3.04 | -4.57 | -2.45 |
| SF5M90T_2417 | murR | HTH-type transcriptional regulator | 2.35 | ||
| SF5M90T_3510 | rhaR | positive regulator for rhaRS operon | 2.33 | ||
| SF5M90T_1595 | malI | repressor of malX and Y genes | 1.98 | ||
| SF5M90T_2125 | yehT | putative two-component response regulator | 1.80 | ||
| SF5M90T_1373 | cspC | cold shock protein | 1.59 | -1.38 | |
| SF5M90T_3349 | malT | positive regulator of mal regulon | 1.58 | 0.58 | |
| SF5M90T_3335 | ompR | osmolarity response regulator | 1.42 | ||
| SF5M90T_3453 | yiaG | putative transcriptional regulator | 1.38 | 3.65 | |
| SF5M90T_2089 | gatR | galactitol utilization operon repressor | 1.33 | ||
| SF5M90T_71 | cra | transcriptional repressor of fru operon and others | 1.16 | ||
| SF5M90T_4197 | yjdC | putative transcriptional regulator | 1.15 | 1.37 | |
| SF5M90T_1370 | putative regulator | 1.09 | |||
| SF5M90T_3578 | rfaH | transcriptional activator | -1.57 | -0.96 | |
| SF5M90T_4242 | yjeB | HTH-type transcriptional repressor | -1.95 | ||
| SF5M90T_984 | cspH | cold shock-like protein | -3.35 | ||
| Replication, recombination and repair | |||||
| SF5M90T_2925 | endA | DNA-specific endonuclease I | 1.42 | -3.26 | -2.74 |
| SF5M90T_3034 | ygjF | G/U mismatch-specific DNA glycosylase | 1.23 | 1.10 | |
| SF5M90T_410 | hupB | DNA-binding protein HU-beta | 1.08 | ||
| SF5M90T_775 | rhlE | putative ATP-dependent RNA helicase | -1.16 | ||
| SF5M90T_3117 | deaD | inducible ATP-independent RNA helicase | -1.20 | ||
| SF5M90T_1769 | dbpA | ATP-dependent RNA helicase | -1.86 | ||
| Poorly characterized | |||||
| General function prediction only | |||||
| SF5M90T_2762 | stpA | DNA-binding protein | 3.51 | -3.00 | -2.60 |
| SF5M90T_275 | putative crossover junction endodeoxyribonuclease | 2.84 | |||
| SF5M90T_2418 | muQ | N-acetylmuramic acid 6-phosphate etherase | 2.77 | -0.93 | |
| SF5M90T_1724 | putative acetyltransferase | 2.06 | -2.41 | ||
| SF5M90T_2435 | putative amino acid antiporter | 2.03 | 1.90 | 1.58 | |
| SF5M90T_2301 | yfbT | putative phosphatase | 2.00 | ||
| SF5M90T_773 | ybhG | putative membrane protein | 1.78 | 1.02 | |
| SF5M90T_1227 | hns | DNA-binding protein | 1.69 | -1.24 | |
| SF5M90T_2275 | yfbB | putative enzyme | 1.62 | -1.90 | -1.85 |
| SF5M90T_3225 | yrdA | putative transferase | 1.51 | ||
| SF5M90T_4236 | hfq | RNA-binding protein | 1.45 | -2.10 | |
| SF5M90T_3315 | gph | phosphoglycolate phosphatase | 1.25 | ||
| SF5M90T_2192 | yeiR | putative GTPases | 1.03 | ||
| SF5M90T_1919 | yedE | putative transport system permease protein | 1.03 | -1.83 | -1.96 |
| SF5M90T_3295 | yhfC | putative transport | 0.97 | -1.30 | -1.25 |
| SF5M90T_2205 | yejK | nucleoid-associated protein | 0.95 | ||
| SF5M90T_2066 | yegH | putative transport protein | 0.84 | ||
| SF5M90T_3344 | yhgH | putative gluconate periplasmic binding protein | 0.81 | ||
| SF5M90T_3102 | yraM | putative glycosylase | 0.73 | ||
| SF5M90T_794 | ybiP | putative enzyme | -1.14 | ||
| SF5M90T_2207 | yejM | putative sulfatase | -1.36 | ||
| SF5M90T_3139 | yhbE | putative permeases of drug/metabolite transporter superfamily | -1.36 | -0.90 | |
| SF5M90T_966 | yccA | putative carrier/transport protein | -1.38 | -1.18 | |
| SF5M90T_2742 | yqaB | putative phosphatase | -1.51 | ||
| SF5M90T_3882 | bax | putative ATP-binding protein | -1.70 | -0.77 | |
| SF5M90T_3370 | yhhX | putative regulator | -1.97 | 1.30 | |
| SF5M90T_3621 | aslB | putative arylsulfatase regulator | -2.73 | ||
| SF5M90T_2516 | putative enzyme | -3.91 | |||
| Function unknown | |||||
| SFxv_3833 | conserved hypothetical protein | 3.59 | -4.23 | ||
| SF5M90T_2431 | conserved hypothetical protein | 2.89 | |||
| SF5M90T_2432 | conserved hypothetical protein | 2.69 | |||
| SF5M90T_11 | uncharacterized protein | 2.57 | |||
| SF5M90T_1402 | yeaD | conserved hypothetical protein | 2.45 | ||
| SF5M90T_828 | ybjO | conserved hypothetical protein | 2.23 | -2.52 | |
| SF5M90T_1941 | dsrB | conserved hypothetical protein | 2.03 | -2.43 | |
| SF5M90T_2302 | yfbU | conserved hypothetical protein | 1.87 | ||
| SF5M90T_451 | ybaK | conserved hypothetical protein | 1.86 | -1.21 | -1.10 |
| SSJG_00311 | conserved hypothetical protein | 1.75 | -1.60 | ||
| SF5M90T_5 | yaaA | conserved hypothetical protein | 1.54 | ||
| SF5M90T_1387 | conserved hypothetical protein | 1.51 | |||
| SF5M90T_3911 | yiiU | conserved hypothetical protein | 1.45 | -2.49 | |
| SF5M90T_957 | conserved hypothetical protein | 1.37 | |||
| SF5M90T_4146 | yjgD | conserved hypothetical protein | 1.28 | -2.30 | |
| SF5M90T_2622 | conserved hypothetical protein | 1.24 | -1.46 | ||
| SF5M90T_3155 | yhbN | conserved hypothetical protein | 0.81 | ||
| SF5M90T_479 | ybbF | conserved hypothetical protein | -1.19 | ||
| SF5M90T_2195 | rtn | conserved hypothetical protein | -1.50 | ||
| SF5M90T_438 | ybaN | conserved hypothetical protein | -1.73 | ||
| SF5M90T_1853 | conserved hypothetical protein | -2.03 | -2.58 | ||
| SF5M90T_1647 | conserved hypothetical protein | -2.11 | |||
| SF5M90T_983 | ymcD | conserved hypothetical protein | -2.34 | ||
| SF5M90T_4094 | yjbA | P-starvation inducible protein PsiE | -2.51 | 1.69 | |
| SF5M90T_1110 | ycfJ | conserved hypothetical protein | -2.52 | 0.99 | |
| SF2861 | hypothetical protein remnant | -2.64 | |||
| SF5M90T_2146 | yohO | membrane protein | -2.96 | ||
| SF5M90T_1952 | putative outer membrane pore protein | -2.98 | |||
| SF5M90T_4307 | putative inner membrane protein | -3.40 | |||
| SF1231 | conserved hypothetical protein | -3.71 | -1.60 | ||
| SF5M90T_427 | ybaA | conserved hypothetical protein | -3.88 | ||
| Phage related | |||||
| S1668 | relF | prophage maintenance protein | 1.75 | ||
| SF5M90T_1793 | putative phage integrase protein | 1.45 | -1.60 | ||
| SF5M90T_1056 | hypothetical bacteriophage protein | 1.14 | |||
| SF5M90T_740 | putative bacteriophage protein | -1.93 | |||
aGenomes used as reference are: S. flexneri 5a str. M90T, S. flexneri 2a str. 301, S. flexneri 2002017, Shigella sp. D9 and S. flexneri 2457 T with GenBank accession numbers AGNM00000000, NC_004337, NC_017328, NZ_GG657384 and NC_004741 respectively.
bGenes are classified in functional categories based on the database of Clusters of Orthologous Groups (COGs). http://www.ncbi.nlm.nih.gov/COG/. Inside each subgroup, genes are arranged in descending order in relation to Log2 of Fold Change values of WT no O2/WT O2 comparison.
cLog2 of Fold Change values of WT no O2/WT O2 and Δfnr no O2/WT no O2 comparisons are presented. Only values considered differentially expressed are shown (p adjust <0.05).
Figure 1.
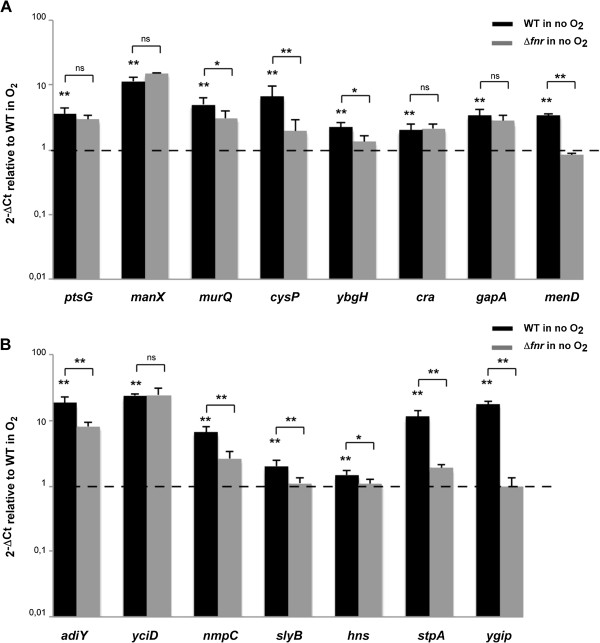
qRT-PCR verification of S. flexneri chromosomal genes induced under anaerobic growth conditions and the role of FNR in the process. Strand specific qRT-PCR analysis of mRNA levels of S. flexneri M90T chromosomal genes shown to be induced under anaerobiosis in RNA-seq analysis. Panel A shows transport and metabolic genes, and panel B acid resistance, OMP and regulatory genes. Data were calculated as the n-fold difference relative to polA (2-ΔCt, where ΔCt represents the difference in threshold cycle between the target and control genes). Results are shown in relation to the wild-type strain 2-ΔCt levels under aerobic conditions, here referred to as 1. Thus, values greater than 1 indicate increased transcription under anaerobiosis, and lower than 1 indicate the opposite. Significant differences were detected when wild-type 2-ΔCt levels under aerobic and anaerobic conditions, or wild-type vs. Δfnr 2-ΔCt levels under anaerobiosis were compared. ns = non-significant, P < 0.05, *; P <0.01, **; n = 4; Mann–Whitney test. Error bars show Standard Deviation (SD).
The expression of other genes involved in transport displayed altered expression in anaerobiosis. For instance, emrD, coding for a drug transporter, cysP, involved in the binding and uptake of sulfate and thiosulfate, yjcE, coding for a Na+/H+ exchanger, ybgH, which encodes a peptide transporter and genes involved in nucleoside transport and catabolism (tsx, nupC, nupG and udp) are induced in anaerobiosis (Table 1, Figure 1A) [33–40].
We found several metabolic genes induced under anaerobic growth such as cra, coding for the catabolite repressor/activator protein, Cra, tpiA, encoding a key enzyme of the gluconeogenic and glycolytic pathways, gapA, involved in glycolysis, yehU/yehT, coding for a two component system involved in responses to carbon starvation, malT, the transcriptional activator of the genes responsible for uptake and metabolism of maltodextrins and proA, which encodes an enzyme in proline biosynthesis [41–47]. The expression of these genes was not FNR-dependent (Table 1, Figure 1A).
In addition to metabolism, we observed anaerobic up-regulation of: genes involved in stress response such as cspC; genes coding for outer membrane proteins (OMPs) such as NmpC, OmpA and SlyB; genes with global regulatory functions such as yjgD that codes for RraB, which interacts with the endonuclease RNase E; yfiA, encoding a ribosome-associated protein that inhibits protein translation; and yejK, hns and its paralogue stpA coding for nucleoid-associated proteins responsible for chromosomal DNA compaction and global gene regulation [48–56]. Interestingly, anaerobic induction of cspC, nmpC, slyB, yjgD, hns and stpA was dependent, at least in part, on FNR (Table 1, Figure 1B). Anaerobiosis can also down-regulate transcription. This is the case for fruBKA, encoding the fructose PTS [29] (Table 1, Figure 2).
Figure 2.
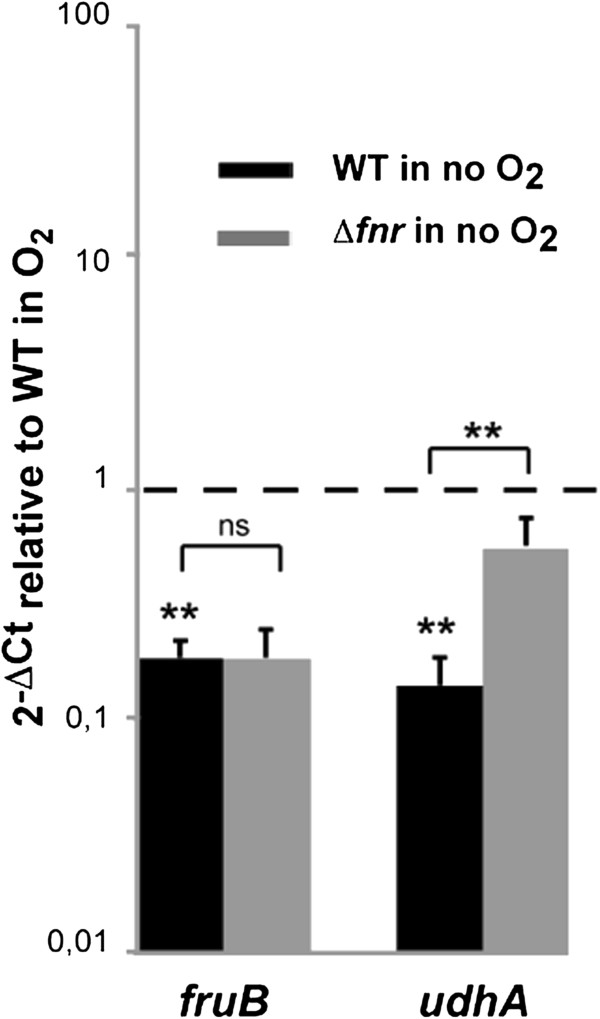
qRT-PCR verification of S. flexneri chromosomal genes repressed under anaerobic growth conditions and the role of FNR in the repression. Strand specific qRT-PCR analysis of mRNA levels of S. flexneri M90T chromosomal genes shown to be repressed in RNA-seq analysis. Data were calculated as the n-fold difference relative to polA (2-ΔCt, where ΔCt represents the difference in threshold cycle between the target and control genes). Results are shown in relation to wild-type 2-ΔCt levels under aerobic conditions, here referred to as 1. Thus, values greater than 1 indicate increased transcription under anaerobiosis and lower than 1 indicates the opposite. Significant differences were detected when wild-type 2-ΔCt levels under aerobic and anaerobic conditions or wild-type vs. Δfnr 2-ΔCt levels under anaerobiosis were compared. P <0.01, **; n = 4; Mann–Whitney test. Error bars show Standard Deviation (SD).
The analysis of genes known to be influenced by anaerobiosis revealed further functions of FNR. This is the case for ygiP, encoding a nucleoid-associated protein induced under anaerobic growth conditions, which we found is FNR-dependent [57]. Furthermore, we observed that menDBCE, genes required for the biosynthesis of quinones with essential roles in anaerobic electron transport systems, are affected by the presence of FNR in contrast to E. coli (Table 1, Figure 1A and B) [58–60].
Our study revealed extended regulatory roles for FNR, such as in the biosynthesis of L-cysteine. Previous work has demonstrated that cysK, which encodes an enzyme in L-cysteine biosynthesis, is subject to FNR regulation and identified an FNR-like domain in cysJ, which encodes a component of the sulfite reductase [16, 61]. Here, we found that loss of FNR affects the entire L-cysteine biosynthetic pathway including genes involved in the uptake and transport of sulfate (i.e. cysPUWAM), sulfate activation (cysDN), reduction to sulfide (cysJIH) and transformation into L-cysteine (cysK) (Table 1, Figure 1A, see Additional file 1: Table S2) [62–64].
Reprogramming of T3SS related genes under anaerobic conditions
Analysis of genes involved in Shigella virulence revealed that multiple genes on the Shigella virulence plasmid, including ipa-mxi-spa genes, were repressed under anaerobic growth in an FNR-dependent manner (Table 2). In contrast, only seven genes on the plasmid (yigB, ospI, shf, rfbU, virK, msbB and parA) were up-regulated in the absence of oxygen; all of these are regulated by FNR except parA and yigB (Table 2). Figure 3 shows effect of oxygen on expression of genes on the virulence plasmid genes. These findings were confirmed by strand specific qRT-PCR for several genes (Figures 4 and 5). Since excess ParA levels compared with ParB can affect plasmid partitioning, we also examined the transcription profile of parB[65]. Similar to parA, mRNA levels of parB are elevated during anaerobic growth (Figure 5). Consistent with this finding, there was no significant difference in loss of the virulence plasmid from bacteria grown in aerobic and anaerobic conditions (not shown).
Table 2.
Virulence plasmid genes differentially expressed in response to anaerobic conditions
| ORF ID ab | Gene | Description | RNA-seq c log2FC | RNA-seq c log2FC | FRT-seq c log2FC |
|---|---|---|---|---|---|
| WT no O 2 /O 2 | Δ fnr /WT no O 2 | Δ fnr /WT no O 2 | |||
| pWR501_0265 | yigB | hypothetical protein | 2.56 | ||
| pWR501_0225 | ospI | T3SS effector | 2.00 | -2.21 | |
| pWR501_0250 | shf | peptidoglycan deacetylase | 1.24 | -3.42 | -1.30 |
| pWR501_0251 | rfbU | glycosiltransferase | 1.21 | -1.65 | -1.16 |
| pWR501_0252 | virK | virulence protein | 1.15 | -2.16 | -1.03 |
| pWR501_0039 | parA | plasmid segregation protein | 1.13 | ||
| pWR501_0253 | msbB | acyltransferase | 1.07 | -3.03 | -1.08 |
| pWR501_0074 | sepA | secreted protease | -1.12 | 1.34 | 1.52 |
| pWR501_0177 | hypothetical protein | -1.55 | 1.04 | ||
| pWR501_0283 | ipaH1.4 | T3SS effector | -1.56 | ||
| pWR501_0175 | hypothetical protein | -1.58 | 2.29 | ||
| pWR501_0176 | hypothetical protein | -1.76 | 2.02 | ||
| pWR501_0015 | hypothetical protein | -2.00 | 1.52 | 1.05 | |
| pWR501_0002 | putative resolvase | -2.04 | 1.97 | ||
| pWR501_0007 | hypothetical protein | -2.25 | 1.63 | ||
| pWR501_0014 | hypothetical protein | -2.25 | 1.60 | 1.27 | |
| pWR501_0192 | virG | invasion protein | -2.25 | 2.36 | |
| pWR501_0144 | ipgF | unknown function | -2.38 | 3.20 | |
| pWR501_0051 | virF | transcriptional activator of virulence | -2.47 | ||
| pWR501_0006 | hypothetical protein | -2.54 | |||
| pWR501_0143 | ipgE | chaperon | -2.55 | 3.39 | |
| pWR501_0146 | mxiH | T3SS component | -2.56 | 1.55 | 4.10 |
| pWR501_0122 | hypothetical protein | -2.58 | 1.53 | 1.95 | |
| pWR501_0121 | hypothetical protein | -2.65 | |||
| pWR501_0191 | virA | T3SS effector | -2.66 | 2.42 | |
| pWR501_0147 | mxiI | T3SS component | -2.80 | 2.00 | 4.01 |
| pWR501_0013 | mkaD | mouse killing factor | -2.81 | 1.97 | 3.33 |
| pWR501_0145 | mxiG | T3SS component | -3.02 | 3.45 | |
| pWR501_0148 | mxiJ | T3SS component | -3.06 | 2.17 | 4.33 |
| pWR501_0031 | hypothetical protein | -3.14 | |||
| pWR501_0005 | hypothetical protein | -3.14 | 2.49 | ||
| pWR501_0292 | sopA | VirG-specific protease | -3.25 | 2.49 | |
| pWR501_0291 | hypothetical protein | -3.31 | |||
| pWR501_0138 | ipgB | invasion protein | -3.34 | 3.66 | |
| pWR501_0157 | spa15 | chaperon | -3.34 | 3.52 | 4.43 |
| pWR501_0012 | shET2-2 | enterotoxin | -3.38 | 3.41 | |
| pWR501_0156 | mxiA | T3SS component | -3.44 | 4.67 | |
| pWR501_0160 | spa32 | invasion protein | -3.45 | 1.34 | 4.63 |
| pWR501_0141 | hypothetical protein | -3.50 | 3.71 | ||
| pWR501_0004 | phoN2 | apyrase | -3.52 | 2.38 | 3.25 |
| pWR501_0150 | mxiL | hypothetical protein | -3.59 | 2.07 | 4.64 |
| pWR501_0132 | acp | hypothetical protein | -3.62 | 5.19 | |
| pWR501_0166 | spa-orf10 | hypothetical protein | -3.64 | 1.97 | 4.52 |
| pWR501_0158 | spa47 | T3SS component | -3.65 | 3.19 | 4.69 |
| pWR501_0140 | icsB | T3SS effector | -3.65 | 2.16 | 4.43 |
| pWR501_0162 | spa24 | T3SS component | -3.70 | 2.28 | 3.71 |
| pWR501_0159 | spa13 | T3SS component | -3.75 | 5.13 | |
| pWR501_0161 | spa33 | T3SS component | -3.75 | 1.99 | 4.03 |
| pWR501_0151 | mxiM | T3SS component | -3.81 | 2.40 | 4.25 |
| pWR501_0155 | mxiC | T3SS component | -3.86 | 2.43 | 4.95 |
| pWR501_0290 | hypothetical protein | -3.86 | 2.00 | ||
| pWR501_0030 | putative enterotoxin fragment | -3.90 | 2.20 | 4.54 | |
| pWR501_0163 | spa9 | T3SS component | -3.92 | 1.93 | 3.56 |
| pWR501_0137 | ipgC | chaperon | -3.93 | 1.86 | 4.04 |
| pWR501_0135 | ipaC | T3SS effector | -3.93 | 2.65 | 5.01 |
| pWR501_0165 | spa40 | T3SS component | -3.94 | 4.02 | |
| pWR501_0139 | ipgA | chaperon | -3.95 | 2.45 | 4.25 |
| pWR501_0153 | mxiD | T3SS component | -3.98 | 2.33 | 4.81 |
| pWR501_0152 | mxiE | transcriptional activator | -4.06 | 2.51 | 4.48 |
| pWR501_0154 | mxiD | T3SS component | -4.08 | 2.66 | 4.54 |
| pWR501_0134 | ipaD | T3SS effector | -4.16 | 2.99 | 4.92 |
| pWR501_0167 | spa-orf11 | hypothetical protein | -4.19 | 2.85 | 4.07 |
| pWR501_0136 | ipaB | T3SS effector | -4.24 | 2.97 | 4.59 |
| pWR501_0133 | ipaA | T3SS effector | -4.24 | 3.09 | 4.98 |
| pWR501_0003 | hypothetical protein | -4.57 | 3.29 | ||
| pWR501_0164 | spa29 | T3SS component | -5.06 | 2.36 | 3.32 |
| pWR501_0131 | virB | transcriptional activator | -5.17 | 2.40 | 4.26 |
a S. flexneri 5a str. M90T pWR501 virulence plasmid sequence was used as reference GenBank accession numbers AF348706.
bGenes are arranged in descending order in relation to Log2 of Fold Change values of WT no O2/WT O2 comparison.
cLog2 of Fold Change values of WT no O2/WT O2 and Δfnr no O2/WT no O2 comparisons are presented. Only values considered differentially expressed are shown (p adjust <0.05).
Figure 3.
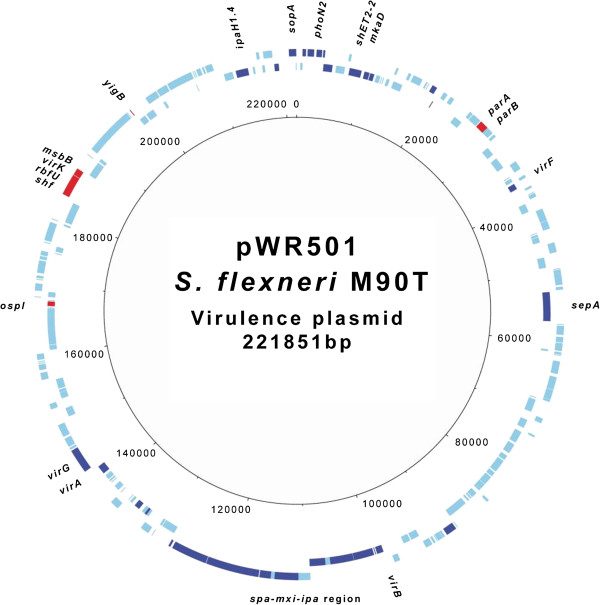
Circular map of genes differentially expressed in the virulence plasmid under anaerobiosis. Outer ring shows ORFs and their orientations. Genes differentially repressed and induced in the wild type M90T strain under anaerobiosis in relation to aerobic conditions were marked in deep blue and red respectively. Scale is in base pairs. The figure was generated with DNAPlotter.
Figure 4.
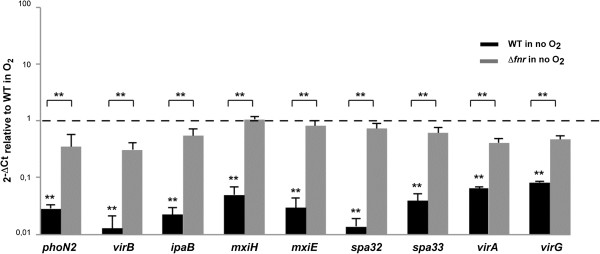
qRT-PCR verification of S. flexneri virulence plasmid genes repressed under anaerobic growth conditions and the role of FNR in the repression. Strand specific qRT-PCR analysis of S. flexneri M90T virulence plasmid genes mRNA levels shown to be repressed in RNA-seq analysis. Data were calculated as the n-fold difference relative to polA (2-ΔCt, where ΔCt represents the difference in threshold cycle between the target and control genes). Results are shown in relation to the wild-type 2-ΔCt levels under aerobic conditions (referred to as 1). Values greater than 1 indicate increased transcription under anaerobiosis, while lower than 1 indicate the opposite. Significant differences were detected with the wild-type strain 2-ΔCt levels under aerobic and anaerobic conditions, or wild-type vs. Δfnr 2-ΔCt levels under anaerobiosis were compared. P <0.01, **; n = 4; Mann–Whitney test. Error bars show Standard Deviation (SD).
Figure 5.
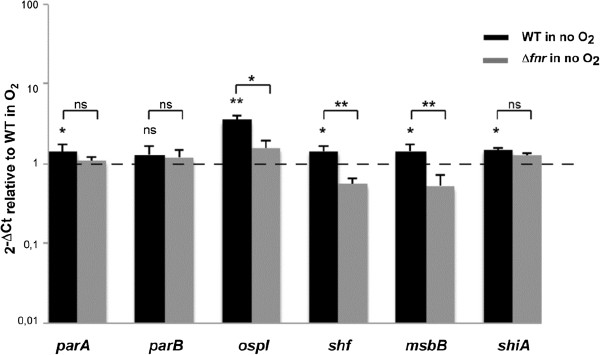
qRT-PCR verification of S. flexneri virulence plasmid genes induced under anaerobic growth conditions and the role of FNR in the induction. Strand specific qRT-PCR analysis of S. flexneri M90T virulence genes mRNA levels shown to be induced in RNA-seq analysis. Data were calculated as the n-fold difference relative to polA (2-ΔCt, where ΔCt represents the difference in threshold cycle between the target and control genes). Results are shown in relation to the wild-type 2-ΔCt levels under aerobic conditions (referred to as 1). Values greater than 1 indicate increased transcription under anaerobiosis, while lower than 1 indicate the opposite. Significant differences were detected with the wild-type strain 2-ΔCt levels under aerobic and anaerobic conditions, or wild-type vs. Δfnr 2-ΔCt levels under anaerobiosis were compared. P < 0.05, *; P <0.01, **; n = 4; Mann–Whitney test. Error bars show standard deviation (SD).
The Shigella pathogenicity island SHI-1 is not present in S. flexneri M90T. Therefore, we examined the transcriptional profile of the SHI-2 pathogenicity island that includes the aerobactin, iron-uptake system [66]. As previously reported, we found that genes encoding the aerobactin system (iucABCD and iutA) were down-regulated under anaerobic conditions, as was shiF, a gene which is also involved in iron acquisition [6, 67]. In contrast, shiA, a SHI-2 gene involved in attenuating host inflammatory responses, was over-expressed under anaerobic conditions when compared to aerobic conditions [68]. Of note, no SHI-2 gene is subject to FNR regulation (Table 1, Figure 5, see Additional file 1: Table S1).
csrB and csrC sRNAs are induced in the absence of oxygen in S. flexneriM90T
Little is known about the small RNAs (sRNAs) in Shigella or their expression under anaerobic conditions. We analysed the sRNAs already described in Shigella as well as potential sRNAs homologues to those described in S. enterica serovar Typhimurium and found that anaerobic growth conditions induce the expression of csrB and csrC in an FNR-independent manner (Table 3, Figure 6) [69–71].
Table 3.
sRNAs differentially expressed in response to anaerobic conditions
| sRNA a | Adjacent genes | Description/class | Length (nt) | RNA-seq b log2FC | RNA-seq b log2FC |
|---|---|---|---|---|---|
| WT no O 2 /O 2 | Δ fnr /WT no O 2 | ||||
| csrB | syd/SF5M90T_2595 | protein-binding sRNA | 360 | 4.97 | |
| csrC | yihi/yihA | protein-binding sRNA | 245 | 3.38 |
asRNAs are arranged in descending order in relation to Log2 of Fold Change values of WT no O2/WT O2 comparison.
bLog2 of Fold Change values of WT no O2/WT O2 and Δfnr no O2/WT no O2 comparisons are presented. Only values considered differentially expressed are shown (p adjust <0.05).
Figure 6.
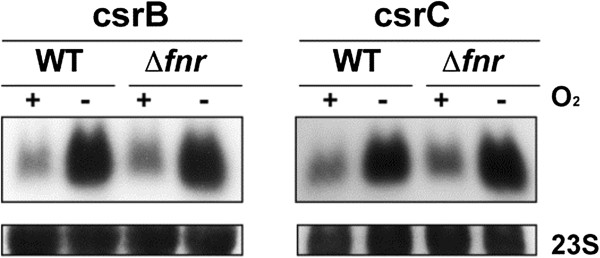
Verification of sRNAs results by Northern Blot. Northern blot analysis of csrB and crsC sRNAs expression under aerobic/anaerobic conditions. 10 μg of total RNA obtained from S. flexneri M90T wild‒type strain and its isogenic Δfnr mutant grown under aerobic and anaerobic conditions until OD600 = 0.2 were separated in 1,25% MOPS‒agarose gels, transferred to membranes and detected using probes specific for the sense strand.
Discussion
In vitro studies have several limitations in relation to in vivo studies; e.g., they cannot mimic the amount and type of carbon sources available for bacteria and lack the signals derived from the interaction with intestinal epithelium, human immune system or other bacteria present in the gut. However, if conducted accurately can provide valuable information.
In the current study we have, for the first time, employed RNA-sequencing to identify oxygen regulated genes in an enteric pathogen. Our findings confirm previous results, but as this method is more sensitive than array based approaches, we identified an extended repertoire of genes modulated by oxygen in an FNR-dependent or -independent manner. For instance, little is known about the role of Cra, a transcriptional regulator of carbon flux (that represses glycolysis and activates gluconeogenesis) here shown to be induced under anaerobic conditions [41]. Interestingly, mutation of cra increases both epithelial cell attachment and invasion by Shigella in aerobic conditions [72]. However, Cra has an entirely distinct role in the virulence of enterohemorrhagic E. coli (EHEC), a close relative of Shigella, when investigated under conditions mimicking the anaerobic environment of the intestinal tract. Under these circumstances, loss of Cra reduces attachment of bacteria to enterocytes [73]. Additionally, Salmonella cra mutants are avirulent when administered orally, indicating that Cra may have key roles in enteric pathogens in anaerobic conditions [74].
While there is an increasing recognition that carbon metabolism affects microbial virulence, it is still not clear whether distinct carbon energy sources are important or preferable for different members of the Enterobacteriaceae[72, 75–80]. For example, our results show that the expression pattern under anaerobic conditions of ptsG, manXYC and fruBKA involved in the transport of sugars is opposite in Shigella to that observed in E. coli[18, 20]. This could be simply due to the different growth medium used in the experiments or to distinct metabolic strategies between Shigella and other Enterobacteriaceae. In favour of the latter and its relationship with virulence it has been shown that mutation of ptsG induces the adherence and invasive capacity of enteroinvasive E. coli (EIEC) strains but not in Salmonella[81]. Further differences between Shigella and other Enterobacteriaceae include adiY, an AraC-like regulator, which activates expression of adiA and adiC, encoding the arginine-dependent acid resistance system (AR3). In Salmonella adiY expression is elevated under aerobic conditions, whereas in Shigella and in E. coli, increased expression of adiY occurs in anaerobiosis [20, 82]. These differences could be due to the strikingly different acid survival strategies that these bacteria seem to develop in spite of being close relatives [83]. Deletion of cad locus, a typical pathoadaptive mutation in Shigella spp., also induces the AR3 system suggesting that this system contributes to the survival of Shigella in its particular niche in the intestinal tract [84, 85].
Interestingly, we observed an FNR-dependent elevated expression under anaerobiosis of hns and overall of stpA and ygiP that encode nucleoid-associated proteins responsible for DNA compaction and global gene regulation, indicating that lack of oxygen profoundly modifies DNA topology in Shigella. Recently, it has been shown that FNR function is strongly inhibited by this kind of nucleoid-associated proteins, which block FNR access to many binding sites [20]. Our findings suggest that FNR is involved in this inhibition, probably indirectly, due to the absence of putative FNR binding-boxes in the promoter region of these genes [20].
To distinguish between direct and indirect effects of FNR, in vivo approaches based in chromatin immunoprecipitation followed by micro-array hybridization (ChIP-chip) or high-throughput sequencing (ChIP-seq) have been performed in E. coli[20, 86]. Correlation of FNR ChIP-seq peaks with transcriptomic data showed that less than half of the FNR-regulated operons could be attributed to direct FNR binding. Of note, FNR occupancy does not always correlate with the presence of a consensus FNR binding site or a change in expression [20, 86]. A total of 19 of E. coli ChIP-seq peaks are located in promoter regions of genes identified in Table 1 (i.e. ptsG, pfkA, gapA, yegT, ptsH, tpiA, lysC, menD, ribB, uspA, slyB, ompA, tonB, yjeA, cspH, deaD, dbpA, yccA and yhhX); only one of these, dbpA, has a canonical FNR binding sequence in its promoter region. Consistent with previous findings, only six of these 19 genes (lysC, menD, slyB, yjeA, yccA and yhhX) were influenced by FNR in our transcriptomic analysis. This result suggests that many FNR effects in Table 1 are likely to be indirect. However, we cannot rule out differences in regulation between E. coli and Shigella that could affect FNR function. Of note, this is the first time that menD, slyB, yjeA and yhhX have been identified as FNR regulated by transcriptome analysis, corroborating previous ChIP findings performed in E. coli.
sRNAs are widespread in bacteria and play critical roles in regulating physiological processes [87]. In Shigella, putative sRNAs have been identified by bioinformatics [69, 70]. However, the expression of these sRNAs has not been confirmed in all cases and little is known about their function or the physiological conditions that induce their expression. Here, we found that anaerobic growth induces expression of two sRNAs, csrB and csrC, independently of FNR. In E. coli csrB and csrC regulate the activity of CsrA, the carbon storage regulator although their function in Shigella has not been characterised so far [88, 89].
For genes directly involved in host:pathogen interactions, we found that oxygen influences the expression of almost all genes in the mxi-spa operon. These T3SS-related genes were down-regulated in the absence of oxygen in an FNR-dependent manner. This is likely to be mediated by VirB as this transcription factor controls many genes in this operon, is influenced by H-NS dependent DNA supercoiling and our findings demonstrate that virB gene is repressed in anaerobiosis [90]. The effect of oxygen on the Shigella T3SS is opposite to Salmonella in which FNR induces expression of invasion genes, and probably reflects the different sites occupied in the host by these two related intestinal pathogens [19]. The results further emphasise that the Shigella T3SS is inactive in anaerobic environments as we previously reported [7].
Inflammation at the site of invasive infection is a hallmark of intestinal shigellosis [91, 92]. Of note, expression of shiA is induced under anaerobiosis. This gene in the SHI-2 pathogenicity island encodes a factor that attenuates the intestinal inflammatory response in shigellosis by decreasing the recruitment of polymorphonuclear leukocytes and T-cells [68, 93]. Similarly OspI is the only T3SS-effector protein that was overexpressed in anaerobiosis; it also serves to dampen inflammatory responses by deaminating a glutamine in host ubiquitin-conjugating enzyme (UBC13) [94]. Thus, expression of both ShiA and OspI under low oxygen tension might dampen the extent of inflammatory responses to Shigella while it is in the anoxic environment of the intestinal lumen, impairing immune responses. Only one operon on the virulence plasmid, shf-rfbU-virK-msbB, was induced under anaerobiosis in an FNR-dependent manner. Interestingly, all these genes are implicated in modification of Shigella lipopolysaccharide (LPS), an important pro-inflammatory mediator [95–99].
The transcription of several genes encoding OMPs was induced under anaerobic growth. Both OmpA and OmpC have been implicated in Shigella virulence, while our results suggest that Tsx, Slp, NmpC, SlyB and YciD (OmpW) could also contribute to pathogenesis and be considered as potential vaccine targets [100, 101]. Indeed, Salmonella OmpW, Tsx and NmpC have already been demonstrated to be immunogenic [102, 103]. In addition to OMPs, transcription of gapA, which encodes glyceraldehyde-3-phosphate dehydrogenase, was induced under anaerobic conditions. Interestingly, this enzyme is exported by EHEC and enteropathogenic E. coli (EPEC) strains but not by non-pathogenic strains. Due to its ability to interact with plasminogen, fibrinogen and intestinal epithelial cells, it has been suggested that GapA might contribute in vivo to the interaction of EHEC and EPEC with the gut epithelium [104].
Conclusions
Overall, our RNA-seq based analysis revealed that in the anaerobic lumen of the intestine Shigella is predicted to prompt both survival and anti-host immune-modulatory activities of the bacterium. This occurs through a reprogramming of bacterial metabolism including altered transcription of genes encoding transport systems and metabolic pathways (Figure 7), likely reflecting the carbon energy sources available in the intestine. Modulation of LPS, along with ShiA and OspI may enable Shigella to subvert inflammatory responses prior to mucosal invasion. Our results highlight the central role of oxygen and FNR in these processes and how it governs bacterial interactions and entry into host cells [7, 68].
Figure 7.
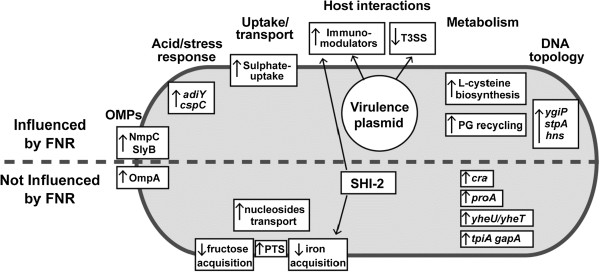
Summary of novel genes influenced by the absence of oxygen in Shigella identified by RNA-seq.
Methods
Bacterial strains and culture conditions
Bacterial strains and plasmids used in this study are shown in Additional file 1: Table S4. E. coli strains were grown in Luria-Bertani (LB; Invitrogen) broth or on LB agar plates while S. flexneri was propagated either in LB broth, tryptic soy broth (TCS; Sigma) or on TCS plates with Congo red (0.01%, Sigma). Experiments under anaerobiosis were performed in an anaerobic workstation (Whitley A35). When required, antibiotics were added at the following concentrations: chloramphenicol 20 μg/ml, ampicillin 100 μg/ml.
Deletion of fnrgene and complementation experiments
The fnr deletion mutant was generated by allelic exchange using pKO3blue plasmid as previously described [105]. Oligonucleotide primers used in this study are listed in Additional file 1: Table S5. Complementation of Δfnr mutant was performed with pBM2, a derivative of pBBR1MCS-4 plasmid that carries a copy of fnr gene under the control of its native promoter. The plasmid pBBR1MCS-4 was used as a control (See Additional file 1: Table S4). The absence of FNR in the Δfnr mutant and its presence in the complemented strain was confirmed by western blot using polyclonal antibodies against FNR as previously described [7] (See Additional file 1: Figure S1).
DNA and RNA extraction methods
S. flexneri M90T genomic DNA for sequencing was isolated as previously described [106]. For RNA extraction bacteria were grown in LB medium with and without oxygen. A 5 ml pre-inoculum was grown over night aerobically or anaerobically with shaking conditions. The pre-inoculums were diluted proportionally to their OD600nm to standardize the input of bacteria to a starting OD600nm of 0.005. Cultures (volume, 175 ml in 1 L flasks) were grown at 37°C, under shaking conditions (200 rpm) until the OD600nm reached 0.2. Three biological replicates were performed for each condition. Total RNA from bacterial pellets was extracted using TRIzol reagent method as previously described [107]. RNA qualities were determined using Agilent RNA Nano Chips (Agilent Technologies).
Genomic DNA was removed from RNA samples using TURBO DNase (Ambion) followed by a second DNase treatment with DNase I (Roche). DNase I treatment was repeated until DNA was not detected by genome-specific PCRs targeting four housekeeping genes (trpB, thrB, purN and mdh) (Additional file 1: Table S5). The RNA quality after DNase treatments was checked using Agilent RNA Nano Chips.
For RNA-seq, total RNA was reverse transcribed using SuperScript III reverse transcriptase (Invitrogen). Actinomycin D (6 μg/ml, Sigma) was added to the reaction to avoid spurious second-strand cDNA synthesis [108]. cDNA was purified using QIAquick PCR purification kit (Qiagen) and used for single stranded cDNA library construction as previously [109, 110]. FRT-seq Illumina libraries were constructed as previously described [111].
Reference genome, sequencing, read mapping and statistic analysis
The genome of S. flexneri M90T was sequenced at Wellcome Trust Sanger Institute using an Illumina HiSeq 2000 sequencer. A total of 0.7 Gb sequence data, in 75-bp paired reads, was obtained (acc. no. ERS033387) and assembled de novo using Velvet [112]. This assembled sequence, which is rich in IS1 elements and for which no attempt of gap closure was performed, is comprised of 501 contigs with a total size of 4.43 Mb. A M90T draft annotated genome was prepared and the annotation transferred from S. flexneri strain 8401 (acc. no. CP000266). Rfam searches were performed and the features identified were included in the annotation as well as Shigella published sRNAs [69, 70]. This draft genome was used as reference for the mapping of RNA-seq reads [113]. During the course of our study the S. flexneri M90T genome was published [114]. Therefore, final expression results are given using this latter locus tag systematic names for coding sequences.
RNA Sequencing was performed using an Illumina HiSeq 2000 sequencer. Raw data as well as mapped reads obtained per replicate were averaged per sample/condition and summarized, together with other interesting quality control parameters, in Additional file 1: Table S3. Processing of reads after mapping included the unmarking of duplicate reads followed by correction to allow for directional fidelity of the data [115]. Output files included per sample, a matrix of readcounts and RPKM values on both sense and antisense strands for genes as well as for automatic 50 bp+/- trimmed intergenic features created in the + strand. The R package DESeq, which implements negative binomial distribution statistics for RNA-seq data was used for statistical analysis [116]. A logarithmic transformed version of the count data (log(x + 1)) was used to avoid zero count values [117]. A p adjust value <0.05, which controls false discovery rate, was used for the cut-off calling of differential expression between conditions. Independent runs of analysis were carried out for sense and antisense directions. Ribosomal genes and repeated sequences, such as transposases or insertion sequences, were filtered out from final tables.
Strand-specific quantitative RT-PCR and Northern blot
A StepOnePlus Real Time PCR system (Applied Biosystems) was used to monitor real-time quantitative PCR. First-strand cDNA was synthesized as previously described but using genome specific primers carrying a tag sequence in the 5′-end instead of random primers. This tag sequence was unique and not found in the genome of S. flexneri M90T. Subsequent PCRs were performed using Power SYBR Green PCR Master Mix (Applied Biosystems) and the tag sequence as one of the paired primers (See Additional file 1: Table S5). As a result, only cDNAs synthesized with a 5′-end tagged primer were amplified. Results are the average of triplicate experiments performed, on at least four independent occasions. Data were expressed relative to polA mRNA levels. To monitor the specificity, final PCR products were analyzed by melting curves. Only samples with no amplification in the control aliquots (not subjected to reverse transcription) were included in the study. The amount of transcripts was expressed as the n-fold difference relative to the control gene (2-ΔCt where ΔCt represents the difference in threshold cycles between the target and control genes). Results were shown in relation to wild type 2-ΔCt levels under aerobic conditions, which were referred as 1. Thus, values greater than 1 indicate increased transcription in relation to the wild-type under aerobic conditions, and lower than 1 indicate the opposite. Significant differences were detected with Mann–Whitney test; values with P <0.05 were considered as significant.
Northern blots were performed as previously described [118]. Radiolabeled RNA probes synthesized with the MAXIscript kit (Ambion) were used to detect specifically the sense of the RNA-targets. The primers used for probes synthesis are listed in Additional file 1: Table S5.
Availability of supporting data
RNA-seq data has been submitted to the European Nucleotide Archive with accession code ERP003817 and the experiment has an ArrayExpress acc. no. E-ERAD-204.
Electronic supplementary material
Additional file 1: Table S1: Chromosomal genes differentially expressed in response to anaerobic conditions and the role of FNR in the induction. This table shows the chromosomal genes differentially expressed in RNA-seq analysis in wild-type S. flexneri M90T grown under anaerobic conditions compared to aerobic conditions, and in Δfnr mutant in relation to wild-type S. flexneri M90T when grown under anaerobic conditions. Genes are classified into functional categories based on the database of Clusters of Orthologous Groups (COGs). Table S2. FNR regulon under anaerobic conditions. This table contains all genes differentially expressed in the Δfnr mutant in relation to the wild-type S. flexneri M90T when grown under anaerobic conditions. RNA-seq and FRT-seq results are presented. Table S3. Summary of mapping statistics. Table S4. Strains and plasmids used in this study. Table S5. Oligonucleotides used in this study. Figure S1. Characterization of M90T Δfnr mutant. This figure confirms the absence of FNR in the Δfnr mutant and shows the growth curve of the mutant in comparison to the wild-type strain M90T and the complemented mutant under anaerobic conditions. (DOCX 4 MB)
Acknowledgements
We thank Jeffrey A. Cole for his advice and suggestions, Iñigo Lasa for pKO3blue plasmid, David Harris and Nathalie Smerdon for data and ArrayExpress submissions; Lira Mamanova for the FRT-seq Illumina library construction; Lesley A. H. Bowman, Malene Cohen, Haifang Zhang and Nuria Vergara for their thoughtful reading of the manuscript and members of Tang group for their help.
M. Vergara-Irigaray was funded by FP7 Marie Curie EIMID-IAPP-217768 grant, and Stopenterics EU grant no. 261472. M. Fookes and N. Thomson are supported by Wellcome Trust grant 098051.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contribution
MV-I participated in the design of the study, performed the research, participated in the analysis of RNA-seq and FRT-seq data and drafted the manuscript. MCF prepared the genome assembly, performed the analysis of RNA-seq and FRT-seq data and helped on figures preparation. NRT participated in the design of the study and the analysis of RNA-seq and FRT-seq data. CMT conceived of the study and participated in its design and coordination and helped to draft the manuscript. All authors read and approved the final manuscript.
Contributor Information
Marta Vergara-Irigaray, Email: mvergara.irigaray@gmail.com.
Maria C Fookes, Email: mcf@sanger.ac.uk.
Nicholas R Thomson, Email: nrt@sanger.ac.uk.
Christoph M Tang, Email: christoph.tang@path.ox.ac.uk.
References
- 1.Sansonetti PJ, Kopecko DJ, Formal SB. Involvement of a plasmid in the invasive ability of Shigella flexneri. Infect Immun. 1982;35(3):852–860. doi: 10.1128/iai.35.3.852-860.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sansonetti PJ, Hale TL, Dammin GJ, Kapfer C, Collins HH, Jr, Formal SB. Alterations in the pathogenicity of Escherichia coli K-12 after transfer of plasmid and chromosomal genes from Shigella flexneri. Infect Immun. 1983;39(3):1392–1402. doi: 10.1128/iai.39.3.1392-1402.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Allen CA, Torres AG. Host-microbe communication within the GI tract. Adv Exp Med Biol. 2008;635:93–101. doi: 10.1007/978-0-387-09550-9_8. [DOI] [PubMed] [Google Scholar]
- 4.Smith H. What happens to bacterial pathogens in vivo? Trends Microbiol. 1998;6(6):239–243. doi: 10.1016/S0966-842X(98)01250-5. [DOI] [PubMed] [Google Scholar]
- 5.Pederson KJ, Pierson DE. Ail expression in Yersinia enterocolitica is affected by oxygen tension. Infect Immun. 1995;63(10):4199–4201. doi: 10.1128/iai.63.10.4199-4201.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Boulette ML, Payne SM. Anaerobic regulation of Shigella flexneri virulence: ArcA regulates Fur and iron acquisition genes. J Bacteriol. 2007;189(19):6957–6967. doi: 10.1128/JB.00621-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Marteyn B, West NP, Browning DF, Cole JA, Shaw JG, Palm F, Mounier J, Prevost MC, Sansonetti P, Tang CM. Modulation of Shigella virulence in response to available oxygen in vivo. Nature. 2010;465(7296):355–358. doi: 10.1038/nature08970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.James BW, Keevil CW. Influence of oxygen availability on physiology, verocytotoxin expression and adherence of Escherichia coli O157. J Appl Microbiol. 1999;86(1):117–124. doi: 10.1046/j.1365-2672.1999.00639.x. [DOI] [PubMed] [Google Scholar]
- 9.Diard S, Toribio AL, Boum Y, Vigier F, Kansau I, Bouvet O, Servin A. Environmental signals implicated in Dr fimbriae release by pathogenic Escherichia coli. Microbes Infect. 2006;8(7):1851–1858. doi: 10.1016/j.micinf.2006.02.023. [DOI] [PubMed] [Google Scholar]
- 10.Singh RD, Khullar M, Ganguly NK. Role of anaerobiosis in virulence of Salmonella typhimurium. Mol Cell Biochem. 2000;215(1–2):39–46. doi: 10.1023/A:1026545630773. [DOI] [PubMed] [Google Scholar]
- 11.Khullar M, Singh RD, Smriti M, Ganguly NK. Anaerobiosis-induced virulence of Salmonella enterica subsp. enterica serovar Typhimurium: role of phospholipase Cgamma signalling cascade. J Med Microbiol. 2003;52(Pt 9):741–745. doi: 10.1099/jmm.0.05186-0. [DOI] [PubMed] [Google Scholar]
- 12.Contreras I, Toro CS, Troncoso G, Mora GC. Salmonella typhi mutants defective in anaerobic respiration are impaired in their ability to replicate within epithelial cells. Microbiology. 1997;143(Pt 8):2665–2672. doi: 10.1099/00221287-143-8-2665. [DOI] [PubMed] [Google Scholar]
- 13.Krishnan HH, Ghosh A, Paul K, Chowdhury R. Effect of anaerobiosis on expression of virulence factors in Vibrio cholerae. Infect Immun. 2004;72(7):3961–3967. doi: 10.1128/IAI.72.7.3961-3967.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kiley PJ, Beinert H. The role of Fe-S proteins in sensing and regulation in bacteria. Curr Opin Microbiol. 2003;6(2):181–185. doi: 10.1016/S1369-5274(03)00039-0. [DOI] [PubMed] [Google Scholar]
- 15.Whitehead RN, Overton TW, Snyder LA, McGowan SJ, Smith H, Cole JA, Saunders NJ. The small FNR regulon of Neisseria gonorrhoeae: comparison with the larger Escherichia coli FNR regulon and interaction with the NarQ-NarP regulon. BMC Genomics. 2007;8:35. doi: 10.1186/1471-2164-8-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Constantinidou C, Hobman JL, Griffiths L, Patel MD, Penn CW, Cole JA, Overton TW. A reassessment of the FNR regulon and transcriptomic analysis of the effects of nitrate, nitrite, NarXL, and NarQP as Escherichia coli K12 adapts from aerobic to anaerobic growth. J Biol Chem. 2006;281(8):4802–4815. doi: 10.1074/jbc.M512312200. [DOI] [PubMed] [Google Scholar]
- 17.Kang Y, Weber KD, Qiu Y, Kiley PJ, Blattner FR. Genome-wide expression analysis indicates that FNR of Escherichia coli K-12 regulates a large number of genes of unknown function. J Bacteriol. 2005;187(3):1135–1160. doi: 10.1128/JB.187.3.1135-1160.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Salmon K, Hung SP, Mekjian K, Baldi P, Hatfield GW, Gunsalus RP. Global gene expression profiling in Escherichia coli K12. The effects of oxygen availability and FNR. J Biol Chem. 2003;278(32):29837–29855. doi: 10.1074/jbc.M213060200. [DOI] [PubMed] [Google Scholar]
- 19.Fink RC, Evans MR, Porwollik S, Vazquez-Torres A, Jones-Carson J, Troxell B, Libby SJ, McClelland M, Hassan HM. FNR is a global regulator of virulence and anaerobic metabolism in Salmonella enterica serovar Typhimurium (ATCC 14028 s) J Bacteriol. 2007;189(6):2262–2273. doi: 10.1128/JB.00726-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Myers KS, Yan H, Ong IM, Chung D, Liang K, Tran F, Keles S, Landick R, Kiley PJ. Genome-scale analysis of Escherichia coli FNR reveals complex features of transcription factor binding. PLoS Genet. 2013;9(6):e1003565. doi: 10.1371/journal.pgen.1003565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rolfe MD, Rice CJ, Lucchini S, Pin C, Thompson A, Cameron AD, Alston M, Stringer MF, Betts RP, Baranyi J, Peck MW, Hinton JC. Lag phase is a distinct growth phase that prepares bacteria for exponential growth and involves transient metal accumulation. J Bacteriol. 2012;194(3):686–701. doi: 10.1128/JB.06112-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sezonov G, Joseleau-Petit D, D’Ari R. Escherichia coli physiology in Luria-Bertani broth. J Bacteriol. 2007;189(23):8746–8749. doi: 10.1128/JB.01368-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mamanova L, Andrews RM, James KD, Sheridan EM, Ellis PD, Langford CF, Ost TW, Collins JE, Turner DJ. FRT-seq: amplification-free, strand-specific transcriptome sequencing. Nat Methods. 2010;7(2):130–132. doi: 10.1038/nmeth.1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tatusov RL, Galperin MY, Natale DA, Koonin EV. The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000;28(1):33–36. doi: 10.1093/nar/28.1.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Salmon KA, Hung SP, Steffen NR, Krupp R, Baldi P, Hatfield GW, Gunsalus RP. Global gene expression profiling in Escherichia coli K12: effects of oxygen availability and ArcA. J Biol Chem. 2005;280(15):15084–15096. doi: 10.1074/jbc.M414030200. [DOI] [PubMed] [Google Scholar]
- 26.Gong S, Richard H, Foster JW. YjdE (AdiC) is the arginine:agmatine antiporter essential for arginine-dependent acid resistance in Escherichia coli. J Bacteriol. 2003;185(15):4402–4409. doi: 10.1128/JB.185.15.4402-4409.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Schweizer H, Larson TJ. Cloning and characterization of the aerobic sn-glycerol-3-phosphate dehydrogenase structural gene glpD of Escherichia coli K-12. J Bacteriol. 1987;169(2):507–513. doi: 10.1128/jb.169.2.507-513.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Eshoo MW. lac fusion analysis of the bet genes of Escherichia coli: regulation by osmolarity, temperature, oxygen, choline, and glycine betaine. J Bacteriol. 1988;170(11):5208–5215. doi: 10.1128/jb.170.11.5208-5215.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tchieu JH, Norris V, Edwards JS, Saier MH., Jr The complete phosphotransferase system in Escherichia coli. J Mol Microbiol Biotechnol. 2001;3(3):329–346. [PubMed] [Google Scholar]
- 30.Dahl U, Jaeger T, Nguyen BT, Sattler JM, Mayer C. Identification of a phosphotransferase system of Escherichia coli required for growth on N-acetylmuramic acid. J Bacteriol. 2004;186(8):2385–2392. doi: 10.1128/JB.186.8.2385-2392.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hadi T, Dahl U, Mayer C, Tanner ME. Mechanistic studies on N-acetylmuramic acid 6-phosphate hydrolase (MurQ): an etherase involved in peptidoglycan recycling. Biochemistry. 2008;47(44):11547–11558. doi: 10.1021/bi8014532. [DOI] [PubMed] [Google Scholar]
- 32.Jaeger T, Arsic M, Mayer C. Scission of the lactyl ether bond of N-acetylmuramic acid by Escherichia coli “etherase”. J Biol Chem. 2005;280(34):30100–30106. doi: 10.1074/jbc.M502208200. [DOI] [PubMed] [Google Scholar]
- 33.Nishino K, Yamaguchi A. Analysis of a complete library of putative drug transporter genes in Escherichia coli. J Bacteriol. 2001;183(20):5803–5812. doi: 10.1128/JB.183.20.5803-5812.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sirko A, Zatyka M, Sadowy E, Hulanicka D. Sulfate and thiosulfate transport in Escherichia coli K-12: evidence for a functional overlapping of sulfate- and thiosulfate-binding proteins. J Bacteriol. 1995;177(14):4134–4136. doi: 10.1128/jb.177.14.4134-4136.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Verkhovskaya ML, Barquera B, Wikstrom M. Deletion of one of two Escherichia coli genes encoding putative Na+/H + exchangers (ycgO) perturbs cytoplasmic alkali cation balance at low osmolarity. Microbiology. 2001;147(Pt 11):3005–3013. doi: 10.1099/00221287-147-11-3005. [DOI] [PubMed] [Google Scholar]
- 36.Casagrande F, Harder D, Schenk A, Meury M, Ucurum Z, Engel A, Weitz D, Daniel H, Fotiadis D. Projection structure of DtpD (YbgH), a prokaryotic member of the peptide transporter family. J Mol Biol. 2009;394(4):708–717. doi: 10.1016/j.jmb.2009.09.048. [DOI] [PubMed] [Google Scholar]
- 37.Hantke K. Phage T6–colicin K receptor and nucleoside transport in Escherichia coli. FEBS Lett. 1976;70(1):109–112. doi: 10.1016/0014-5793(76)80737-5. [DOI] [PubMed] [Google Scholar]
- 38.Westh Hansen SE, Jensen N, Munch-Petersen A. Studies on the sequence and structure of the Escherichia coli K-12 nupG gene, encoding a nucleoside-transport system. Eur J Biochem. 1987;168(2):385–391. doi: 10.1111/j.1432-1033.1987.tb13431.x. [DOI] [PubMed] [Google Scholar]
- 39.Craig JE, Zhang Y, Gallagher MP. Cloning of the nupC gene of Escherichia coli encoding a nucleoside transport system, and identification of an adjacent insertion element, IS 186. Mol Microbiol. 1994;11(6):1159–1168. doi: 10.1111/j.1365-2958.1994.tb00392.x. [DOI] [PubMed] [Google Scholar]
- 40.Pritchard RH, Ahmad SI. Fluorouracil and the isolation of mutants lacking uridine phosphorylase in Escherichia coli: location of the gene. Mol Gen Genet. 1971;111(1):84–88. doi: 10.1007/BF00286557. [DOI] [PubMed] [Google Scholar]
- 41.Saier MH, Jr, Ramseier TM. The catabolite repressor/activator (Cra) protein of enteric bacteria. J Bacteriol. 1996;178(12):3411–3417. doi: 10.1128/jb.178.12.3411-3417.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Anderson A, Cooper RA. Gluconeogenesis in Escherichia coli The role of triose phosphate isomerase. FEBS Lett. 1969;4(1):19–20. doi: 10.1016/0014-5793(69)80184-5. [DOI] [PubMed] [Google Scholar]
- 43.Branlant G, Branlant C. Nucleotide sequence of the Escherichia coli gap gene. Different evolutionary behavior of the NAD + -binding domain and of the catalytic domain of D-glyceraldehyde-3-phosphate dehydrogenase. Eur J Biochem. 1985;150(1):61–66. doi: 10.1111/j.1432-1033.1985.tb08988.x. [DOI] [PubMed] [Google Scholar]
- 44.Kraxenberger T, Fried L, Behr S, Jung K. First insights into the unexplored two-component system YehU/YehT in Escherichia coli. J Bacteriol. 2012;194(16):4272–4284. doi: 10.1128/JB.00409-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Boos W, Shuman H. Maltose/maltodextrin system of Escherichia coli: transport, metabolism, and regulation. Microbiol Mol Biol Rev. 1998;62(1):204–229. doi: 10.1128/mmbr.62.1.204-229.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Baich A. Proline synthesis in Escherichia coli.A proline-inhibitable glutamic acid kinase. Biochim Biophys Acta. 1969;192(3):462–467. doi: 10.1016/0304-4165(69)90395-X. [DOI] [PubMed] [Google Scholar]
- 47.Baich A. The biosynthesis of proline in Escherichia coli: phosphate-dependent glutamate -semialdehyde dehydrogenase (NADP), the second enzyme in the pathway. Biochim Biophys Acta. 1971;244(1):129–134. doi: 10.1016/0304-4165(71)90129-2. [DOI] [PubMed] [Google Scholar]
- 48.Atlung T, Ingmer H. H-NS: a modulator of environmentally regulated gene expression. Mol Microbiol. 1997;24(1):7–17. doi: 10.1046/j.1365-2958.1997.3151679.x. [DOI] [PubMed] [Google Scholar]
- 49.Cohen-Or I, Shenhar Y, Biran D, Ron EZ. CspC regulates rpoS transcript levels and complements hfq deletions. Res Microbiol. 2010;161(8):694–700. doi: 10.1016/j.resmic.2010.06.009. [DOI] [PubMed] [Google Scholar]
- 50.Molloy MP, Herbert BR, Walsh BJ, Tyler MI, Traini M, Sanchez JC, Hochstrasser DF, Williams KL, Gooley AA. Extraction of membrane proteins by differential solubilization for separation using two-dimensional gel electrophoresis. Electrophoresis. 1998;19(5):837–844. doi: 10.1002/elps.1150190539. [DOI] [PubMed] [Google Scholar]
- 51.Pugsley AP, Schnaitman CA. Identification of three genes controlling production of new outer membrane pore proteins in Escherichia coli K-12. J Bacteriol. 1978;135(3):1118–1129. doi: 10.1128/jb.135.3.1118-1129.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chai TJ, Foulds J. Purification of protein A, an outer membrane component missing in Escherichia coli K-12 ompA mutants. Biochim Biophys Acta. 1977;493(1):210–215. doi: 10.1016/0005-2795(77)90274-4. [DOI] [PubMed] [Google Scholar]
- 53.Ludwig A, Tengel C, Bauer S, Bubert A, Benz R, Mollenkopf HJ, Goebel W. SlyA, a regulatory protein from Salmonella typhimurium, induces a haemolytic and pore-forming protein in Escherichia coli. Mol Gen Genet. 1995;249(5):474–486. doi: 10.1007/BF00290573. [DOI] [PubMed] [Google Scholar]
- 54.Gao J, Lee K, Zhao M, Qiu J, Zhan X, Saxena A, Moore CJ, Cohen SN, Georgiou G. Differential modulation of E. coli mRNA abundance by inhibitory proteins that alter the composition of the degradosome. Mol Microbiol. 2006;61(2):394–406. doi: 10.1111/j.1365-2958.2006.05246.x. [DOI] [PubMed] [Google Scholar]
- 55.Polikanov YS, Blaha GM, Steitz TA. How hibernation factors RMF, HPF, and YfiA turn off protein synthesis. Science. 2012;336(6083):915–918. doi: 10.1126/science.1218538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lee C, Marians KJ. Characterization of the nucleoid-associated protein YejK. J Biol Chem. 2013;288(44):31503–31516. doi: 10.1074/jbc.M113.494237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Teramoto J, Yoshimura SH, Takeyasu K, Ishihama A. A novel nucleoid protein of Escherichia coli induced under anaerobiotic growth conditions. Nucleic Acids Res. 2010;38(11):3605–3618. doi: 10.1093/nar/gkq077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Unden G. Differential roles for menaquinone and demethylmenaquinone in anaerobic electron transport of E. coli and their fnr-independent expression. Arch Microbiol. 1988;150(5):499–503. doi: 10.1007/BF00422294. [DOI] [PubMed] [Google Scholar]
- 59.Shestopalov AI, Bogachev AV, Murtazina RA, Viryasov MB, Skulachev VP. Aeration-dependent changes in composition of the quinone pool in Escherichia coli. Evidence of post-transcriptional regulation of the quinone biosynthesis. FEBS Lett. 1997;404(2–3):272–274. doi: 10.1016/S0014-5793(97)00143-9. [DOI] [PubMed] [Google Scholar]
- 60.Sharma V, Hudspeth ME, Meganathan R. Menaquinone (vitamin K2) biosynthesis: localization and characterization of the menE gene from Escherichia coli. Gene. 1996;168(1):43–48. doi: 10.1016/0378-1119(95)00721-0. [DOI] [PubMed] [Google Scholar]
- 61.Gruez A, Zeghouf M, Bertrand J, Eschenbrenner M, Coves J, Fontecave M, Pignol D, Fontecilla-Camps JC. The FNR-like domain of the Escherichia coli sulfite reductase flavoprotein component: crystallization and preliminary X-ray analysis. Acta Crystallogr D Biol Crystallogr. 1998;54(Pt 1):135–136. doi: 10.1107/S090744499701069X. [DOI] [PubMed] [Google Scholar]
- 62.Sirko A, Hryniewicz M, Hulanicka D, Bock A. Sulfate and thiosulfate transport in Escherichia coli K-12: nucleotide sequence and expression of the cysTWAM gene cluster. J Bacteriol. 1990;172(6):3351–3357. doi: 10.1128/jb.172.6.3351-3357.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Leyh TS, Taylor JC, Markham GD. The sulfate activation locus of Escherichia coli K12: cloning, genetic, and enzymatic characterization. J Biol Chem. 1988;263(5):2409–2416. [PubMed] [Google Scholar]
- 64.Loughlin RE. Polarity of the cysJIH operon of Salmonella typhimurium. J Gen Microbiol. 1975;86(2):275–282. doi: 10.1099/00221287-86-2-275. [DOI] [PubMed] [Google Scholar]
- 65.Abeles AL, Friedman SA, Austin SJ. Partition of unit-copy miniplasmids to daughter cells. III. The DNA sequence and functional organization of the P1 partition region. J Mol Biol. 1985;185(2):261–272. doi: 10.1016/0022-2836(85)90402-4. [DOI] [PubMed] [Google Scholar]
- 66.Moss JE, Cardozo TJ, Zychlinsky A, Groisman EA. The selC-associated SHI-2 pathogenicity island of Shigella flexneri. Mol Microbiol. 1999;33(1):74–83. doi: 10.1046/j.1365-2958.1999.01449.x. [DOI] [PubMed] [Google Scholar]
- 67.Lemaitre C, Bidet P, Bingen E, Bonacorsi S. Transcriptional analysis of the Escherichia coli ColV-Ia plasmid pS88 during growth in human serum and urine. BMC Microbiol. 2012;12:115. doi: 10.1186/1471-2180-12-115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ingersoll MA, Moss JE, Weinrauch Y, Fisher PE, Groisman EA, Zychlinsky A. The ShiA protein encoded by the Shigella flexneri SHI-2 pathogenicity island attenuates inflammation. Cell Microbiol. 2003;5(11):797–807. doi: 10.1046/j.1462-5822.2003.00320.x. [DOI] [PubMed] [Google Scholar]
- 69.Peng J, Yang J, Jin Q. An integrated approach for finding overlooked genes in Shigella. PLoS One. 2011;6(4):e18509. doi: 10.1371/journal.pone.0018509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Skippington E, Ragan MA. Evolutionary dynamics of small RNAs in 27 Escherichia coli and Shigella genomes. Genome Biol Evol. 2012;4(3):330–345. doi: 10.1093/gbe/evs001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kroger C, Dillon SC, Cameron AD, Papenfort K, Sivasankaran SK, Hokamp K, Chao Y, Sittka A, Hebrard M, Handler K, Colgan A, Leekitcharoenphon P, Langridge GC, Lohan AJ, Loftus B, Lucchini S, Ussery DW, Dorman CJ, Thomson NR, Vogel J, Hinton JC. The transcriptional landscape and small RNAs of Salmonella enterica serovar Typhimurium. Proc Natl Acad Sci USA. 2012;109(20):E1277–E1286. doi: 10.1073/pnas.1201061109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Gore AL, Payne SM. CsrA and Cra influence Shigella flexneri pathogenesis. Infect Immun. 2010;78(11):4674–4682. doi: 10.1128/IAI.00589-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Njoroge JW, Nguyen Y, Curtis MM, Moreira CG, Sperandio V. Virulence meets metabolism: Cra and KdpE gene regulation in enterohemorrhagic Escherichia coli. MBio. 2012;3(5):e00280–12. doi: 10.1128/mBio.00280-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Allen JH, Utley M, van Den Bosch H, Nuijten P, Witvliet M, McCormick BA, Krogfelt KA, Licht TR, Brown D, Mauel M, Leatham MP, Laux DC, Cohen PS. A functional cra gene is required for Salmonella enterica serovar typhimurium virulence in BALB/c mice. Infect Immun. 2000;68(6):3772–3775. doi: 10.1128/IAI.68.6.3772-3775.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Eisenreich W, Dandekar T, Heesemann J, Goebel W. Carbon metabolism of intracellular bacterial pathogens and possible links to virulence. Nat Rev Microbiol. 2010;8(6):401–412. doi: 10.1038/nrmicro2351. [DOI] [PubMed] [Google Scholar]
- 76.Joseph B, Przybilla K, Stuhler C, Schauer K, Slaghuis J, Fuchs TM, Goebel W. Identification of Listeria monocytogenes genes contributing to intracellular replication by expression profiling and mutant screening. J Bacteriol. 2006;188(2):556–568. doi: 10.1128/JB.188.2.556-568.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Marrero J, Rhee KY, Schnappinger D, Pethe K, Ehrt S. Gluconeogenic carbon flow of tricarboxylic acid cycle intermediates is critical for Mycobacterium tuberculosis to establish and maintain infection. Proc Natl Acad Sci USA. 2010;107(21):9819–9824. doi: 10.1073/pnas.1000715107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Tang Y, Wu W, Zhang X, Lu Z, Chen J, Fang W. Catabolite control protein a of Streptococcus suis type 2 contributes to sugar metabolism and virulence. J Microbiol. 2012;50(6):994–1002. doi: 10.1007/s12275-012-2035-3. [DOI] [PubMed] [Google Scholar]
- 79.Heroven AK, Sest M, Pisano F, Scheb-Wetzel M, Steinmann R, Bohme K, Klein J, Munch R, Schomburg D, Dersch P. Crp Induces Switching of the CsrB and CsrC RNAs in Yersinia pseudotuberculosis and Links Nutritional Status to Virulence. Front Cell Infect Microbiol. 2012;2:158. doi: 10.3389/fcimb.2012.00158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Altier C, Suyemoto M, Lawhon SD. Regulation of Salmonella enterica serovar typhimurium invasion genes by csrA. Infect Immun. 2000;68(12):6790–6797. doi: 10.1128/IAI.68.12.6790-6797.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gotz A, Goebel W. Glucose and glucose 6-phosphate as carbon sources in extra- and intracellular growth of enteroinvasive Escherichia coli and Salmonella enterica. Microbiology. 2010;156(Pt 4):1176–1187. doi: 10.1099/mic.0.034744-0. [DOI] [PubMed] [Google Scholar]
- 82.Kieboom J, Abee T. Arginine-dependent acid resistance in Salmonella enterica serovar Typhimurium. J Bacteriol. 2006;188(15):5650–5653. doi: 10.1128/JB.00323-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lin J, Lee IS, Frey J, Slonczewski JL, Foster JW. Comparative analysis of extreme acid survival in Salmonella typhimurium, Shigella flexneri, and Escherichia coli. J Bacteriol. 1995;177(14):4097–4104. doi: 10.1128/jb.177.14.4097-4104.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Casalino M, Prosseda G, Barbagallo M, Iacobino A, Ceccarini P, Latella MC, Nicoletti M, Colonna B. Interference of the CadC regulator in the arginine-dependent acid resistance system of Shigella and enteroinvasive E. coli. Int J Med Microbiol. 2010;300(5):289–295. doi: 10.1016/j.ijmm.2009.10.008. [DOI] [PubMed] [Google Scholar]
- 85.Maurelli AT, Fernandez RE, Bloch CA, Rode CK, Fasano A. “Black holes” and bacterial pathogenicity: a large genomic deletion that enhances the virulence of Shigella spp. and enteroinvasive Escherichia coli. Proc Natl Acad Sci USA. 1998;95(7):3943–3948. doi: 10.1073/pnas.95.7.3943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Grainger DC, Aiba H, Hurd D, Browning DF, Busby SJ. Transcription factor distribution in Escherichia coli: studies with FNR protein. Nucleic Acids Res. 2007;35(1):269–278. doi: 10.1093/nar/gkl1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Waters LS, Storz G. Regulatory RNAs in bacteria. Cell. 2009;136(4):615–628. doi: 10.1016/j.cell.2009.01.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Weilbacher T, Suzuki K, Dubey AK, Wang X, Gudapaty S, Morozov I, Baker CS, Georgellis D, Babitzke P, Romeo T. A novel sRNA component of the carbon storage regulatory system of Escherichia coli. Mol Microbiol. 2003;48(3):657–670. doi: 10.1046/j.1365-2958.2003.03459.x. [DOI] [PubMed] [Google Scholar]
- 89.Liu MY, Gui G, Wei B, Preston JF, 3rd, Oakford L, Yuksel U, Giedroc DP, Romeo T. The RNA molecule CsrB binds to the global regulatory protein CsrA and antagonizes its activity in Escherichia coli. J Biol Chem. 1997;272(28):17502–17510. doi: 10.1074/jbc.272.28.17502. [DOI] [PubMed] [Google Scholar]
- 90.Tobe T, Yoshikawa M, Mizuno T, Sasakawa C. Transcriptional control of the invasion regulatory gene virB of Shigella flexneri: activation by virF and repression by H-NS. J Bacteriol. 1993;175(19):6142–6149. doi: 10.1128/jb.175.19.6142-6149.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Perdomo OJ, Cavaillon JM, Huerre M, Ohayon H, Gounon P, Sansonetti PJ. Acute inflammation causes epithelial invasion and mucosal destruction in experimental shigellosis. J Exp Med. 1994;180(4):1307–1319. doi: 10.1084/jem.180.4.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Perdomo JJ, Gounon P, Sansonetti PJ. Polymorphonuclear leukocyte transmigration promotes invasion of colonic epithelial monolayer by Shigella flexneri. J Clin Invest. 1994;93(2):633–643. doi: 10.1172/JCI117015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ingersoll MA, Zychlinsky A. ShiA abrogates the innate T-cell response to Shigella flexneri infection. Infect Immun. 2006;74(4):2317–2327. doi: 10.1128/IAI.74.4.2317-2327.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Sanada T, Kim M, Mimuro H, Suzuki M, Ogawa M, Oyama A, Ashida H, Kobayashi T, Koyama T, Nagai S, Shibata Y, Gohda J, Inoue J, Mizushima T, Sasakawa C. The Shigella flexneri effector OspI deamidates UBC13 to dampen the inflammatory response. Nature. 2012;483(7391):623–626. doi: 10.1038/nature10894. [DOI] [PubMed] [Google Scholar]
- 95.Kaoukab-Raji A, Biskri L, Bernardini ML, Allaoui A. Characterization of SfPgdA, a Shigella flexneri peptidoglycan deacetylase required for bacterial persistence within polymorphonuclear neutrophils. Microbes Infect. 2012;14(7–8):619–627. doi: 10.1016/j.micinf.2012.01.009. [DOI] [PubMed] [Google Scholar]
- 96.Kaniuk NA, Vinogradov E, Li J, Monteiro MA, Whitfield C. Chromosomal and plasmid-encoded enzymes are required for assembly of the R3-type core oligosaccharide in the lipopolysaccharide of Escherichia coli O157:H7. J Biol Chem. 2004;279(30):31237–31250. doi: 10.1074/jbc.M401879200. [DOI] [PubMed] [Google Scholar]
- 97.Wing HJ, Goldman SR, Ally S, Goldberg MB. Modulation of an outer membrane protease contributes to the virulence defect of Shigella flexneri strains carrying a mutation in the virK locus. Infect Immun. 2005;73(2):1217–1220. doi: 10.1128/IAI.73.2.1217-1220.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.D’Hauteville H, Khan S, Maskell DJ, Kussak A, Weintraub A, Mathison J, Ulevitch RJ, Wuscher N, Parsot C, Sansonetti PJ. Two msbB genes encoding maximal acylation of lipid A are required for invasive Shigella flexneri to mediate inflammatory rupture and destruction of the intestinal epithelium. J Immunol. 2002;168(10):5240–5251. doi: 10.4049/jimmunol.168.10.5240. [DOI] [PubMed] [Google Scholar]
- 99.Zhong QP. Pathogenic effects of Opolysaccharide from Shigella flexneri strain. World J Gastroenterol. 1999;5(3):245–248. doi: 10.3748/wjg.v5.i3.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Bernardini ML, Sanna MG, Fontaine A, Sansonetti PJ. OmpC is involved in invasion of epithelial cells by Shigella flexneri. Infect Immun. 1993;61(9):3625–3635. doi: 10.1128/iai.61.9.3625-3635.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Ambrosi C, Pompili M, Scribano D, Zagaglia C, Ripa S, Nicoletti M. Outer membrane protein A (OmpA): a new player in shigella flexneri protrusion formation and inter-cellular spreading. PLoS One. 2012;7(11):e49625. doi: 10.1371/journal.pone.0049625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Singh R, Shasany AK, Aggarwal A, Sinha S, Sisodia BS, Khanuja SP, Misra R. Low molecular weight proteins of outer membrane of Salmonella typhimurium are immunogenic in Salmonella induced reactive arthritis revealed by proteomics. Clin Exp Immunol. 2007;148(3):486–493. doi: 10.1111/j.1365-2249.2007.03362.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Yang TC, Ma XC, Liu F, Lin LR, Liu LL, Liu GL, Tong ML, Fu ZG, Zhou L. Screening of the Salmonella paratyphi A CMCC 50973 strain outer membrane proteins for the identification of potential vaccine targets. Mol Med Rep. 2012;5(1):78–83. doi: 10.3892/mmr.2011.587. [DOI] [PubMed] [Google Scholar]
- 104.Egea L, Aguilera L, Gimenez R, Sorolla MA, Aguilar J, Badia J, Baldoma L. Role of secreted glyceraldehyde-3-phosphate dehydrogenase in the infection mechanism of enterohemorrhagic and enteropathogenic Escherichia coli: interaction of the extracellular enzyme with human plasminogen and fibrinogen. Int J Biochem Cell Biol. 2007;39(6):1190–1203. doi: 10.1016/j.biocel.2007.03.008. [DOI] [PubMed] [Google Scholar]
- 105.Solano C, Garcia B, Latasa C, Toledo-Arana A, Zorraquino V, Valle J, Casals J, Pedroso E, Lasa I. Genetic reductionist approach for dissecting individual roles of GGDEF proteins within the c-di-GMP signaling network in Salmonella. Proc Natl Acad Sci USA. 2009;106(19):7997–8002. doi: 10.1073/pnas.0812573106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Marmur J. A procedure for the isolation of Deoxyribonucleic acid from micro-organisms. J Mol Biol. 1961;3:208–218. doi: 10.1016/S0022-2836(61)80047-8. [DOI] [Google Scholar]
- 107.Toledo-Arana A, Dussurget O, Nikitas G, Sesto N, Guet-Revillet H, Balestrino D, Loh E, Gripenland J, Tiensuu T, Vaitkevicius K, Barthelemy M, Vergassola M, Nahori MA, Soubigou G, Regnault B, Coppee JY, Lecuit M, Johansson J, Cossart P. The Listeria transcriptional landscape from saprophytism to virulence. Nature. 2009;459(7249):950–956. doi: 10.1038/nature08080. [DOI] [PubMed] [Google Scholar]
- 108.Perocchi F, Xu Z, Clauder-Munster S, Steinmetz LM. Antisense artifacts in transcriptome microarray experiments are resolved by actinomycin D. Nucleic Acids Res. 2007;35(19):e128. doi: 10.1093/nar/gkm683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Croucher NJ, Fookes MC, Perkins TT, Turner DJ, Marguerat SB, Keane T, Quail MA, He M, Assefa S, Bahler J, Kingsley RA, Parkhill J, Bentley SD, Dougan G, Thomson NR. A simple method for directional transcriptome sequencing using Illumina technology. Nucleic Acids Res. 2009;37(22):e148. doi: 10.1093/nar/gkp811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Quail MA, Kozarewa I, Smith F, Scally A, Stephens PJ, Durbin R, Swerdlow H, Turner DJ. A large genome center’s improvements to the Illumina sequencing system. Nat Methods. 2008;5(12):1005–1010. doi: 10.1038/nmeth.1270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Mamanova L, Turner DJ. Low-bias, strand-specific transcriptome Illumina sequencing by on-flowcell reverse transcription (FRT-seq) Nat Protoc. 2011;6(11):1736–1747. doi: 10.1038/nprot.2011.399. [DOI] [PubMed] [Google Scholar]
- 112.Zerbino DR, Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008;18(5):821–829. doi: 10.1101/gr.074492.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R. The Sequence Alignment/Map format and SAMtools. Bioinformatics. 2009;25(16):2078–2079. doi: 10.1093/bioinformatics/btp352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Onodera NT, Ryu J, Durbic T, Nislow C, Archibald JM, Rohde JR. Genome sequence of Shigella flexneri serotype 5a strain M90T Sm. J Bacteriol. 2012;194(11):3022. doi: 10.1128/JB.00393-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Croucher NJ, Vernikos GS, Parkhill J, Bentley SD. Identification, variation and transcription of pneumococcal repeat sequences. BMC Genomics. 2011;12:120. doi: 10.1186/1471-2164-12-120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Anders S, Huber W. Differential expression analysis for sequence count data. Genome Biol. 2010;11(10):R106. doi: 10.1186/gb-2010-11-10-r106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Simon Anders WH. Diferential expression of RNA-Seq data at the gene level -the DESeq package. 2013. [Google Scholar]
- 118.Lasa I, Toledo-Arana A, Dobin A, Villanueva M, de los Mozos IR, Vergara-Irigaray M, Segura V, Fagegaltier D, Penades JR, Valle J, Solano C, Gingeras TR. Genome-wide antisense transcription drives mRNA processing in bacteria. Proc Natl Acad Sci USA. 2011;108(50):20172–20177. doi: 10.1073/pnas.1113521108. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1: Chromosomal genes differentially expressed in response to anaerobic conditions and the role of FNR in the induction. This table shows the chromosomal genes differentially expressed in RNA-seq analysis in wild-type S. flexneri M90T grown under anaerobic conditions compared to aerobic conditions, and in Δfnr mutant in relation to wild-type S. flexneri M90T when grown under anaerobic conditions. Genes are classified into functional categories based on the database of Clusters of Orthologous Groups (COGs). Table S2. FNR regulon under anaerobic conditions. This table contains all genes differentially expressed in the Δfnr mutant in relation to the wild-type S. flexneri M90T when grown under anaerobic conditions. RNA-seq and FRT-seq results are presented. Table S3. Summary of mapping statistics. Table S4. Strains and plasmids used in this study. Table S5. Oligonucleotides used in this study. Figure S1. Characterization of M90T Δfnr mutant. This figure confirms the absence of FNR in the Δfnr mutant and shows the growth curve of the mutant in comparison to the wild-type strain M90T and the complemented mutant under anaerobic conditions. (DOCX 4 MB)