

Abstract
There is a long‐standing controversy as to whether drought limits photosynthetic CO2 assimilation through stomatal closure or by metabolic impairment in C3 plants. Comparing results from different studies is difficult due to interspecific differences in the response of photosynthesis to leaf water potential and/or relative water content (RWC), the most commonly used parameters to assess the severity of drought. Therefore, we have used stomatal conductance (g) as a basis for comparison of metabolic processes in different studies. The logic is that, as there is a strong link between g and photosynthesis (perhaps co‐regulation between them), so different relationships between RWC or water potential and photosynthetic rate and changes in metabolism in different species and studies may be ‘normalized’ by relating them to g. Re‐analysing data from the literature using light‐saturated g as a parameter indicative of water deficits in plants shows that there is good correspondence between the onset of drought‐induced inhibition of different photosynthetic sub‐processes and g. Contents of ribulose bisphosphate (RuBP) and adenosine triphosphate (ATP) decrease early in drought development, at still relatively high g (higher than 150 mmol H2O m–2 s–1). This suggests that RuBP regeneration and ATP synthesis are impaired. Decreased photochemistry and Rubisco activity typically occur at lower g (<100 mmol H2O m–2 s–1), whereas permanent photoinhibition is only occasional, occurring at very low g (<50 mmol H2O m–2 s–1). Sub‐stomatal CO2 concentration decreases as g becomes smaller, but increases again at small g. The analysis suggests that stomatal closure is the earliest response to drought and the dominant limitation to photosynthesis at mild to moderate drought. However, in parallel, progressive down‐regulation or inhibition of metabolic processes leads to decreased RuBP content, which becomes the dominant limitation at severe drought, and thereby inhibits photosynthetic CO2 assimilation.
Key words: C3 plants, drought, water stress, photosynthesis, stomatal conductance, photochemistry, carboxylation, photophosphorylation, RuBP regeneration, Rubisco
INTRODUCTION
There is a long‐standing controversy as to whether drought mainly limits photosynthesis through stomatal closure (Sharkey, 1990; Chaves, 1991; Ortet al., 1994; Cornic and Massacci, 1996) or by metabolic impairment (Boyer, 1976; Lawlor, 1995). Evidence that impaired ATP synthesis is the main factor limiting photosynthesis even under mild drought (Boyer, 1976; Tezaraet al., 1999) has further stimulated debate (Cornic, 2000; Lawlor and Cornic, 2002).
Comparing results from different authors is difficult due to interspecific differences in the response of photosynthesis to leaf water potential and/or relative water content (RWC), the parameters most commonly used to assess the degree of drought (Tardieu and Simmoneau, 1998). To overcome this, we have exploited the relationship between stomatal conductance (g) and photosynthetic CO2 assimilation (Wonget al., 1979), since an early and progressive effect of drought is stomatal closure (Boyer, 1976; Sharkey, 1990; Chaves, 1991; Ortet al., 1994; Lawlor, 1995; Cornic and Massacci, 1996). We have recently demonstrated (Flexaset al., 2002; Medranoet al., 2002) that, by relating photosynthetic parameters to their corresponding light‐saturated g, a pattern of responses is revealed which is independent of acclimation processes and only slightly dependent on the cultivars and species. For instance, the relationships between different photosynthetic parameters and the absolute values of g in grapevines (Vitis vinifera) and several Mediterranean sclerophyll shrubs were very similar. This applied even when maximum g reached approx. 500 mmol H2O m–2 s–1 in grapevines, and only 200 mmol H2O m–2 s–1 in sclerophyll shrubs (Medranoet al., 2002). The relationship between different photosynthetic parameters and g was not observed with relative water content or leaf water potential, i.e. decreased photosynthesis caused by drought occurred at different leaf water status in different species, albeit at similar stomatal conductance. Based on these previous findings and using data from the literature, we have analysed at what values of g—and thus at different severity of drought—some photosynthetic metabolic processes are impaired.
MATERIALS AND METHODS
In order to see if g, relative water content or water potential provide a clearer basis or reference for the effects of drought on photosynthetic response to drought, we analysed the literature cited in Table 1.
Table 1.
References used for the analysis of each photosynthetic sub‐process and in the construction of Fig. 2)
| Photosynthetic sub‐process | References | Species |
| RuBP availability | Flexas, 2000 | Vitis vinifera |
| Giménez et al., 1992 | Helianthus annuus | |
| Gunasekera and Berkowitz, 1993 | Nicotiana tabacum | |
| Santakumary and Berkowitz, 1991 | Spinacia oleracea | |
| Sharkey and Badger, 1982 | Xanthium strumarium | |
| Sharkey and Seeman, 1989 | Phaseolus vulgaris | |
| Stuhlfaulth et al., 1990 | Digitalis lanata | |
| Tezara et al., 1999 | Helianthus annuus | |
| Vu et al., 1987 | Glycine max | |
| Wingler et al., 1999 | Hordeum vulgare | |
| ATP synthesis | Havaux et al., 1987 | Nicotiana tabacum |
| Lawlor, 1983 | Triticum aestivum | |
| Meyer and de Kouchkovsky, 1992 | Lupinus albus | |
| Sharkey and Badger, 1982 | Xanthium strumarium | |
| Tezara et al., 1999 | Helianthus annuus | |
| Younis et al., 1979 | Spinacia oleracea | |
| Photochemistry | Björkman and Powles, 1984 | Nerium oleander |
| Brestic et al., 1995 | Phaseolus vulgaris | |
| Damesin and Rambal, 1995 | Quercus pubescens | |
| Demmig et al., 1988 | Nerium oleander | |
| Faria et al., 1998 | Quercus ilex, Q. suber, Olea europaea, Eucalyptus globulus | |
| Flexas, 2000; Flexas et al., 1998, 1999a, 1999b | Vitis vinifera | |
| Lal et al., 1996 | Vicia faba, Hordeum vulgare | |
| Meyer and Genty, 1999 | Rosa rubiginosa | |
| Munné‐Bosch and Alegre, 2000 | Melissa officinalis | |
| Munné‐Bosch et al., 1999 | Lavandula stoechas, Rosmarinus officinalis | |
| Pankovic et al., 1999 | Helianthus annuus | |
| Wingler et al., 1999 | Hordeum vulgare | |
| Rubisco activity | Antolín and Sánchez‐Díaz, 1993 | Medicago sativa |
| Castrillo and Calcagno, 1989 | Lycopersicon esculentum | |
| Holaday et al., 1992 | Triticum aestivum | |
| Lal et al., 1996 | Vicia faba, Hordeum vulgare | |
| Medrano et al., 1997 | Trifolium subterraneum | |
| Pankovic et al., 1999 | Helianthus annuus | |
| Plaut and Federman, 1991 | Gossypum hirsutum | |
| Tezara et al., 1999 | Helianthus annuus | |
| Vu and Yelenosky, 1988 | Citrus sinensis | |
| Vu et al., 1987 | Glycine max | |
| Wingler et al., 1999 | Hordeum vulgare | |
| Permanent photoinhibition | Angelopoulos et al., 1996 | Olea europaea |
| Brodribb, 1996 | Acacia melanoxylon, Eucalyptus tenuiramis, Podocarpus lawrencii | |
| Faria et al., 1998 | Quercus ilex, Q. suber, Olea europaea, Eucalyptus globulus | |
| Flexas et al., 1998; Flexas, 2000 | Vitis vinifera | |
| Méthy et al., 1996 | Quercus pubescens | |
| Ramanjulu et al., 1998 | Morus alba | |
| Valladares and Pearcy, 1997 | Heteromeles arbutifolia | |
| Ci inflexion point | Brodribb, 1996 | Acacia melanoxylon, Eucalyptus tenuiramis, Podocarpus lawrencii |
| Epron and Dreyer, 1993 | Quercus robur, Q. petraea | |
| Faver et al., 1996 | Gossypum hirsutum | |
| Flexas, 2000 | Vitis vinifera | |
| Giménez et al., 1992 | Helianthus annuus | |
| Jensen et al., 1996 | Brassica napus | |
| Johnson et al., 1987 | Triticum ssp. | |
| Lal et al., 1996 | Vicia faba, Hordeum vulgare | |
| Luo, 1991 | Abutilon theophrasti | |
| Martin and Ruiz‐Torres, 1992 | Triticum aestivum | |
| Nicolodi et al., 1988 | Medicago sativa | |
| Ramanjulu et al., 1998 | Morus alba | |
| Shangguan et al., 1999 | Triticum aestivum |
The species analysed in every reference are indicated in the right‐hand column.
Photosynthetic metabolism was divided into five sub‐processes implicated as important sites of inhibition of photosynthetic metabolism under drought. The sub‐processes were: (1) ribulose 1,5‐bisphosphate (RuBP) regeneration capacity (Giménezet al., 1992; Gunasekera and Berkowitz, 1993) as indicated by the RuBP content in leaves; (2) ATP synthesis (Youniset al., 1979; Meyer and de Kouchkovsky, 1992; Tezaraet al., 1999) as indicated by the ATP content of leaves or ATP synthase activity (photophosphorylation) or the amount of ATP synthase; (3) leaf photochemistry (Cornic and Massacci, 1996; Flexaset al., 1999a, b) as indicated by chlorophyll a fluorescence; (4) ribulose 1,5‐bisphosphate carboxylase/oxidase (Rubisco) activity (Castrillo and Calcagno, 1989; Medranoet al., 1997; Tezaraet al, 1999); and (5) permanent photoinhibition (Björkman and Powles, 1984; Valladares and Pearcy, 1997). In addition, the change in sub‐stomatal CO2 concentration (Ci) with progressive drought was also analysed as an indicator of the predominance of stomatal or non‐stomatal limitations to photosynthesis (Ortet al., 1994; Cornic and Massacci, 1996). We related the Ci inflexion point between decreasing and increasing Ci to the value of g.
The data were grouped according to the change in each of these five sub‐processes (Table 1), irrespective of the methods used to assess the effects of drought in each experiment (usually gas exchange or photoacoustic measurements, coupled with determinations of chlorophyll a fluorescence, on leaves, followed by destructive sampling and biochemical analyses). Changes in Rubisco activity and RuBP regeneration derived from CO2‐response curves of photosynthesis (A/Ci curves) were not considered, since they assume that regulation under non‐stressed conditions is applicable to stressed. In addition, they are difficult to compare with biochemical assessments (Medranoet al., 2002).
For each study and sub‐process, the threshold of g below which the sub‐processes was impaired by the drought treatment (i.e. the value of g at which each process started to become inhibited) was estimated. When g was not given, it was derived from the relationship between g and leaf water potential obtained for the same species under similar conditions either by the same or other authors. When there were uncertainties about the values of g, these studies were not included in the analysis.
Finally, for simplicity and because only approximate g values were usually available (or impossible, for example, to determine accurately from the figures given), the occurrence of inhibition of each sub‐process (expressed as a percentage of the total number of cases analysed) was related to four discrete intervals of g. These were: g > 150 mmol H2O m–2 s–1 (i.e. control plants to mild drought); 150 mmol H2O m–2 s–1 > g > 100 mmol H2O m–2 s–1 (i.e. moderate drought); 100 mmol H2O m–2 s–1 > g > 50 mmol H2O m–2 s–1 (i.e. severe drought); g < 50 mmol H2O m–2 s–1 (i.e. very severe drought). When the data were available, results were also related to discrete intervals of relative water content and leaf water potential.
This method determines the onset of changes in metabolism with progressive drought, by comparison with unstressed plants (the control). If the changes in a particular process occur with only small increase in stress, they appear in the range of g > 150 mmol H2O m–2 s–1 (i.e. control plants to mild drought). This is because the g values of the control plants are not distinguished from mildly stressed plants. It means that the onset of metabolic changes occurs with very limited drought as g starts to decrease.
RESULTS AND DISCUSSION
Using different values of stomatal conductance, g, as a reference to analyse the effects of drought on photosynthetic metabolism provides a clearer pattern of the changes in different parts of metabolism in response to drought than using relative water content or leaf water potential. This is illustrated in Fig. 1 for Rubisco activity. When plotted as a function of g intervals, Rubisco activity starts to decrease when g drops below 100 mmol H2O m–2 s–1 (Fig. 1A). However, when plotted as a function of the RWC intervals proposed by Lawlor (1995) to reflect different stages of drought effects on photosynthesis, no clear pattern was observed (Fig. 1B). Rubisco activity decreased in 65 % of studies at RWC between 80 and 50 %, but in a substantial proportion (35 %) of cases, Rubisco activity was lost at very high RWC (90–100 %). With leaf water potential as a reference (Fig. 1C), the pattern of response was even less clear, with Rubisco activity inhibited over a range of water potentials. Other photosynthetic processes showed similar responses to g, RWC and leaf water potential (not shown). Using RWC or water potential as references, only photochemistry and permanent photoinhibition showed a degree of correspondence similar to that observed when using g. Photochemistry decreased mainly between 80 and 50 % RWC with leaf water potentials below –1 MPa. However, permanent photoinhibition occurred at RWC between 80 and 50 % as well, but at leaf water potentials only below –1·5 MPa. Following this primary evaluation, we used g as a reference parameter to analyse the literature.
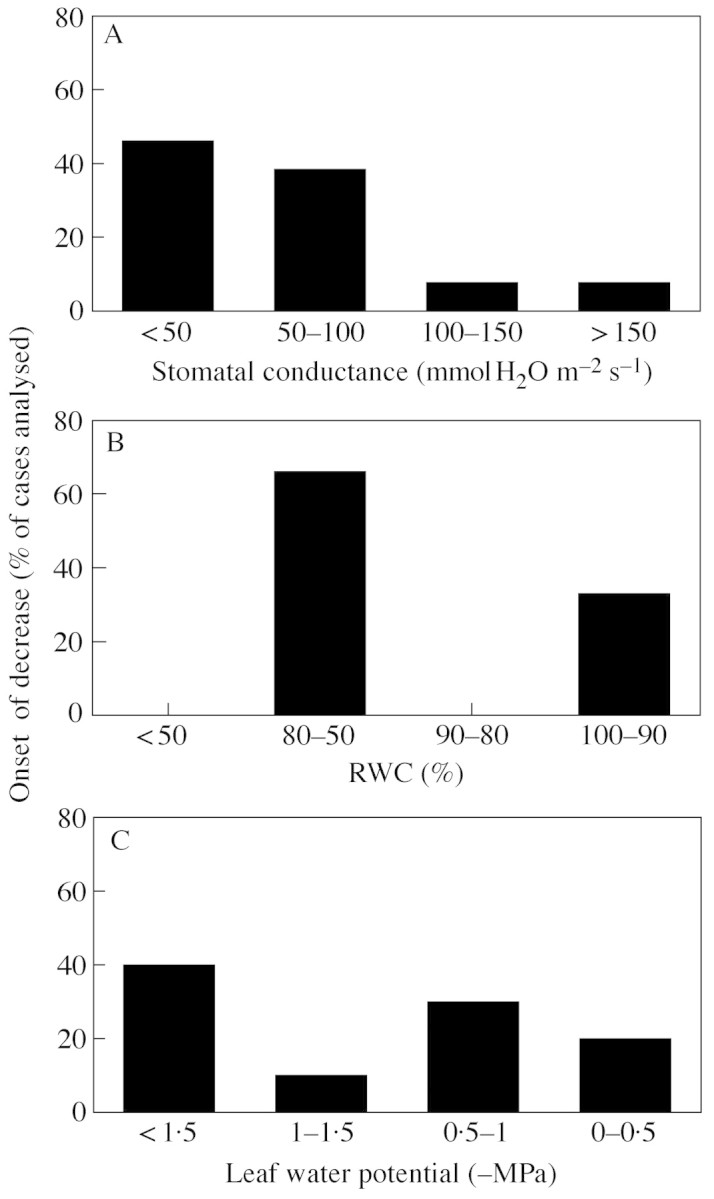
Fig. 1. Analysis of Rubisco activity under drought. The y‐axis shows the percentage (%) of the studies from the literature in which the activity of Rubisco first decreased in relation to intervals of (A) light‐saturated stomatal conductance, (B) of relative water content (RWC) and (C) of leaf water potential.
The results of this analysis are given in Fig. 2. Clearly, decreased RuBP (Fig. 2A) and impaired ATP synthesis (Fig. 2B) have been most frequently reported to occur in early phases of drought, when g is still relatively large (>150 mmol H2O m–2 s–1). Important exceptions are the studies of Sharkey and Seeman (1989), in which RuBP content of Phaseolus vulgaris was unaffected at g around 100 mmol H2O m–2 s–1, and of Ortiz‐Lópezet al. (1991), in which inhibition of ATPase in Helianthus annuus did not occur even at very low g (approx. 50 mmol H2O m–2 s–1).

Fig. 2. Occurrence of the onset of drought‐induced decrease of metabolic processes as a function of the corresponding light saturated stomatal conductance (g), from the literature (Table 1). The y‐axis shows the percentage (%) of the studies (the number is shown as n) in which the decrease occurred at different intervals of g. For simplicity, those studies in which no effect of drought on metabolism occurred are not included but are mentioned in the text. The effects on metabolism are represented by: A, RuBP content (RuBP regeneration, n = 10); B, ATP content (ATP synthesis, n = 6); C, Photochemistry (n = 14); D, Rubisco activity (n = 13); E, Permanent photoinhibition (n = 10); F, Appearance of the Ci inflexion point (n = 17).
Decreased photochemistry (Fig. 2C) and Rubisco activity (Fig. 2D) are commonly reported to occur at severe stress, and in our analysis this corresponded to g < 100 mmol H2O m–2 s–1. Only in the study of Munné‐Boschet al. (1999) in Rosmarinus officinalis, were electron transport rates unaffected even when g dropped to 75 mmol H2O m–2 s–1. We found only two reports, both using Phaseolus vulgaris, showing unaltered Rubisco activity at g < 100 mmol H2O m–2 s–1 (Sharkey and Seeman, 1989; Bresticet al., 1995).
Permanent photoinhibition (Fig. 2E) was only occasional. Indeed, in about half the references analysed permanent photoinhibition did not occur; when it did, it was at very low g (<50 mmol H2O m–2 s–1) (see Epron and Dreyer, 1993; Fariaet al., 1998; Flexas and Medrano, 1998).
As stomata close, the CO2 inside the leaf, Ci, initially declines with increasing stress and then increases as drought becomes more severe (Lawlor, 1995). According to Cornic and Massacci (1996), Ci estimated from fluorescence decreases to compensation point under drought and can be estimated accurately. If Ci is high, this reflects the inaccuracies in the Ci calculation under drought (i.e. heterogeneous stomatal closure, cuticular conductance, etc.), which tend to overestimate Ci. The decrease in Ci indicates that stomatal limitations dominate, with moderate drought, irrespective of any metabolic impairment. However, at a certain stage of water stress, shown by a threshold value of g, Ci frequently increases, indicating the predominance of non‐stomatal limitations to photosynthesis. In most cases the point at which Ci starts to increase, which we call the Ci inflexion point, occurs at g around 50 mmol H2O m–2 s–1. Only Nicolodiet al. (1988) in Medicago sativa and Luo (1991) in Abutilon theophrasti observed the Ci inflexion point at higher g.
The results of this literature survey analysing the effects of drought on photosynthesis are consistent with a gradual pattern of response of photosynthesis to water stress similar to that proposed by Lawlor (1995). After an early partial closure of stomata, metabolic limitation, caused by either damage (i.e. permanent) or adjustment (i.e. reversible ‘down‐regulation’) occurs. The limitation at large g, when drought is mild, is often impaired ATP synthesis and thus ATP‐limited regeneration of RuBP. Further reduction of g as drought increases leads to reduced photochemical activity. The analysis shows that, as it is the Rubisco activity, this loss is more progressive with increasing drought than sometimes suggested (Lawlor, 1995; Lawlor and Cornic, 2002). Photoinhibition eventually occurs under conditions of very severe drought and almost complete stomata closure. The Ci inflexion point is also observed predominantly at low g.
This pattern of metabolic changes supports the assertion by Cornic (2000) that stomatal closure is the primary cause of the reduction in photosynthetic rate under mild drought, but shows that metabolic damage or down‐regulation—this analysis cannot distinguish between them—is progressive and commences with small changes in g under mild drought. In particular, decreased ATP content, implying impaired synthesis [and thus supporting the observations of Youniset al. (1979) and Tezaraet al. (1999) of impaired photophosphorylation and loss of ATP synthase, respectively] is important. To our knowledge, only one reference (Ortiz‐Lópezet al., 1991) reported no inhibition of ATPase under mild to moderate drought. A major consequence of loss of ATP would be limited RuBP regeneration under mild drought, shown clearly as an early effect of drought by our analysis. Nevertheless, despite the decreased capacity of these metabolic processes, decreased Ci confirms the predominance of stomatal limitation in restricting photosynthetic rate in the early phase of water loss. However, the metabolic changes are responsible for loss of photosynthetic potential during this phase (Lawlor and Cornic, 2002).
Our analysis does not include the effects of drought on nitrate reductase and sucrose phosphate synthase, enzymes shown in a number of studies to be inhibited under water stress. This is because too few analyses with information on g are available. The activities of these enzymes can be restored by placing the water‐stressed plant in high CO2 for a number of hours (Sharkey, 1990; Cornic and Massacci, 1996). This strongly suggests that CO2 availability in the chloroplast, mainly regulated by g, may serve as a signal to trigger metabolic adjustments in the leaf in response to water deficit. This would be consistent with the observed response of the different photosynthetic processes to g. ATP synthesis is probably not restored by elevated CO2 (Tanget al., 2002), suggesting that the enzyme is not impaired, directly or indirectly, by low CO2 concentration. Instead, increased magnesium concentration has been shown to inhibit ATP synthase (Tanget al., 2002). Alternatively, inhibition of ATP synthesis, and not lowered Ci, may be responsible for impairments to metabolism, which cannot be regulated by adjustments in metabolism. One of the major goals for future research on drought effects on photosynthesis should be to confirm how general are the responses that have been identified (Lawlor, 1995; Lawlor and Cornic, 2002). From an analysis of the literature over the widest range of drought and for a number of species with different responses to drought, we have shown that changes in metabolism occur despite stomatal closure. It is still uncertain if these are the consequences of damage to or adjustment (down‐regulation) in metabolism, and better understanding of the mechanisms is required.
ACKNOWLEDGEMENTS
We thank Drs D. W. Lawlor, M. A. J. Parry and W. Tezara for critical reading and useful comments on the manuscript.
Supplementary Material
Received: 27 April 2001; Returned for revision: 13 August 2001; Accepted: 22 October 2001.
References
- AngelopoulosK, Dichio B, Xiloyannis C.1996. Inhibition of photosynthesis in olive trees (Olea europaea L.) during water stress and rewatering. Journal of Experimental Botany 47: 1093–1100. [Google Scholar]
- AntolínMC, Sánchez‐Díaz M.1993. Effects of temporary droughts on photosynthesis of alfalfa plants. Journal of Experimental Botany 44: 1341–1349. [Google Scholar]
- BeadleCL, Jarvis PG.1977. Effects of shoot water status on some photosynthetic partial processes in Sitka spruce. Physiologia Plantarum 41: 7–13. [Google Scholar]
- BjörkmanO, Powles SB.1984. Inhibition of photosynthetic reactions under water stress: interaction with light level. Planta 161: 490–504. [DOI] [PubMed] [Google Scholar]
- BoyerJS.1976. Photosynthesis at low water potentials. Philosophical Transactions of the Royal Society B 273: 501–512. [Google Scholar]
- BresticM, Cornic G, Fryer MJ, Baker NR.1995. Does photorespiration protect the photosynthetic apparatus in French bean leaves from photoinhibition during drought stress? Planta 196: 450–457. [Google Scholar]
- BrodribbT.1996. Dynamics of changing intercellular CO2 concentration (Ci) during drought and determination of minimum functional Ci. Plant Physiology 111: 179–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CastrilloM, Calcagno AM.1989. Effects of water stress and rewatering on ribulose 1,5‐bisphosphate carboxylase activity, chlorophyll and protein contents in two cultivars of tomato. Journal of Horticultural Science 64: 717–724. [Google Scholar]
- CornicG.2000. Drought stress inhibits photosynthesis by decreasing stomatal aperture—not by affecting ATP synthesis. Trends in Plant Science 5: 187–188. [Google Scholar]
- CornicG, Massacci A.1996. Leaf photosynthesis under drought stress. In: Baker NR, ed. Photosynthesis and the Environment. The Netherlands: Kluwer Academic Publishers. [Google Scholar]
- ChavesMM.1991. Effects of water deficits on carbon assimilation. Journal of Experimental Botany 42: 1–16. [Google Scholar]
- DamesinC, Rambal S.1995. Field study of leaf photosynthetic performance by a Mediterranean deciduous oak tree (Quercus pubescens) during a summer drought. New Phytologist 131: 159–167. [Google Scholar]
- DemmigB, Winter K, Krüger A, Czygan FC.1988. Zeaxanthin and the heat dissipation of excess light energy in Nerium oleander exposed to a combination of high light and water stress. Plant Physiology 87: 17–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EpronD, Dreyer E.1993. Photosynthesis of oak leaves under water stress: maintenance of high photochemical efficiency of photosystem II and occurrence of non‐uniform CO2 assimilation. Tree Physiology 13: 107–117. [DOI] [PubMed] [Google Scholar]
- FariaT, Silvério D, Breia E, Cabral R, Abadia A, Abadia J, Pereira JS, Chaves, MM.1998. Differences in the response of carbon assimilation to summer stress (water deficits, high light and temperature) in four Mediterranean tree species. Physiologia Plantarum 102: 419–428. [Google Scholar]
- FaverKL, Gerik TJ, Thaxton PM, El‐Zik KM.1996. Late season water stress in cotton: II. Leaf gas exchange and assimilation capacity. Crop Science 36: 922–928. [Google Scholar]
- FlexasJ.2000. Regulation of the photosynthetic processes in response to drought in leaves of Vitis vinifera L. PhD Thesis, University of the Balearic Islands, Spain. (In Spanish). [Google Scholar]
- FlexasJ, Escalona JM, Medrano H.1998. Down‐regulation of photosynthesis by drought under field conditions in grapevine leaves. Australian Journal of Plant Physiology 25: 893–900. [Google Scholar]
- FlexasJ, Badger M, Chow WS, Medrano H, Osmond CB.1999a Analysis of the relative increase in photosynthetic O2 uptake when photosynthesis in grapevine leaves is inhibited following low night temperatures and/or water stress. Plant Physiology 121: 675–684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FlexasJ, Escalona JM, Medrano H.1999b Water stress induces different levels of photosynthesis and electron transport rate regulations in grapevines. Plant, Cell and Environment 22: 39–48. [Google Scholar]
- FlexasJ, Bota J, Escalona JM, Sampol B, Medrano, H.2002. Effects of drought on photosynthesis in grapevines under field conditions: an evaluation of stomatal and mesophyll limitations. Australian Journal of Plant Physiology (in press). [DOI] [PubMed] [Google Scholar]
- GiménezC, Mitchell VJ, Lawlor DW.1992. Regulation of photosynthesis rate of two sunflower hybrids under water stress. Plant Physiology 98: 516–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GunasekeraD, Berkowitz GA.1993. Use of transgenic plants with Rubisco antisense DNA to evaluate the rate limitation of photosynthesis under water stress. Plant Physiology 103: 629–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HavauxM, Canaani O, Malkin S.1987. Inhibition of photosynthetic activities under slow water stress measured in vivo by the photoacoustic method. Physiologia Plantarum 70: 503–510. [Google Scholar]
- HoladayAS, Ritchie SW, Nguyen HT.1992. Effects of water deficit on gas‐exchange parameters and ribulose 1,5‐bisphosphate carboxylase activation in wheat. Environmental and Experimental Botany 32: 403–410. [Google Scholar]
- JensenCR, Mogensen G, Mortensen VO, Andersen MN, Schjoerring JK, Thage JH, Koribidis J.1996. Leaf photosynthesis and drought adaptation in field‐grown oilseed rape (Brassica napus L.). Australian Journal of Plant Physiology 23: 631–644. [Google Scholar]
- JohnsonRC, Mornhinweg DW, Ferris DM, Heitholt JJ.1987. Leaf photosynthesis and conductance of selected Triticum species at different water potentials. Plant Physiology 83: 1014–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LalA, Ku MSB, Edwards GE.1996. Analysis of inhibition of photosynthesis due to water stress in the C3 species Hordeum vulgare and Vicia faba: electron transport, CO2 fixation and carboxylation capacity. Photosynthesis Research 49: 57–69. [DOI] [PubMed] [Google Scholar]
- LawlorDW.1983. Integration of biochemical processes in the physiology of water stressed plants. In: Marcelle R, Clijters H, von Puche M, eds. Effects of stress on photosynthesis. The Hague, Boston, London: Martinus Nijhoff/Dr W. Junk Publishers. [Google Scholar]
- LawlorDW.1995. The effects of water deficit on photosynthesis. In: Smirnoff N, ed. Environment and Plant Metabolism. Flexibility and Acclimation. Oxford: BIOS Scientific Publishers. [Google Scholar]
- LawlorDW, Cornic G.2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant, Cell and Environment (in press). [DOI] [PubMed] [Google Scholar]
- LuoY.1991. Changes of Ci/Ca in association with non‐stomatal limitation to photosynthesis in water stressed Abutilon theophrasti Photosynthetica 25: 273–279. [Google Scholar]
- MartinB, Ruiz‐Torres NA.1992. Effects of water‐deficit stress on photosynthesis, its components and component limitations, and on water use efficiency in wheat (Triticum aestivum L.). Plant Physiology 100: 733–739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MedranoH, Escalona JM, Bota J, Gulías J, Flexas J.2002. Regulation of photosynthesis of C3 plants in response to progressive drought: the stomatal conductance as a reference parameter. Annals of Botany (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- MedranoH, Parry MA, Socias X, Lawlor DW.1997. Long‐term water stress inactivates Rubisco in subterranean clover. Annals of Applied Biology 131: 491–501. [Google Scholar]
- MéthyM, Damesin C, Rambal S.1996. Drought and photosystem II activity in two Mediterranean oaks. Annales des Sciences Forestières 53: 255–262. [Google Scholar]
- MeyerS, de Kouchkovsky Y.1992. ATPase state and activity in thylakoids from normal and water‐stressed lupin. FEBS Letters 303: 233–236. [DOI] [PubMed] [Google Scholar]
- MeyerS, Genty B.1999. Heterogeneous inhibition of photosynthesis over the leaf surface of Rosa rubiginosa L. during water stress and abscisic acid treatment: induction of a metabolic component by limitation of CO2 diffusion. Planta 210: 126–131. [DOI] [PubMed] [Google Scholar]
- Munné‐BoschS, Alegre L.2000. The significance of β‐carotene, α‐tocopherol and the xanthophyll cycle in droughted Melissa officinalis L. plants. Australian Journal of Plant Physiology 27: 139–146. [Google Scholar]
- Munné‐BoschS, Nogués S, Alegre L.1999. Diurnal variations of photosynthesis and dew absorption by leaves in two evergreen shrubs growing in Mediterranean field conditions. New Phytologist 144: 109–119. [Google Scholar]
- NicolodiC, Massacci A, Di Marco G.1988. Water status effects on net photosynthesis in field‐grown alfalfa. Crop Science 28: 944–948. [Google Scholar]
- OrtDR, Oxborough K, Wise RR.1994. Depressions of photosynthesis in crops with water deficits. In: Baker NR, Bowyer JR, eds. Photoinhibition of photosynthesis from molecular mechanisms to the field. Oxford: BIOS Scientific Publishers. [Google Scholar]
- Ortiz‐LópezA, Ort DR, Boyer JS.1991. Photophosphorylation in attached leaves of Helianthus annuus at low water potentials. Plant Physiology 96: 1018–1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PankovicD, Sakac, Z, Kevresan, S, Plesnicar, M.1999. Acclimation to long‐term water deficit in the leaves of two sunflower hybrids: photosynthesis, electron transport and carbon metabolism. Journal of Experimental Botany 50: 127–138. [Google Scholar]
- PlautZ, Federman E.1991. Acclimation of CO2 assimilation in cotton leaves to water stress and salinity. Plant Physiology 97: 515–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- RamanjuluS, Sreenivasalu N, Giridhara Kumar S, Sudhakar C.1998. Photosynthetic characteristics in mulberry during water stress and rewatering. Photosynthetica 35: 259–263. [Google Scholar]
- SantakumariM, Berkowitz GA.1991. Chloroplast volume: cell water potential relationships and acclimation of photosynthesis to leaf water deficits. Photosynthesis Research 28: 9–20. [DOI] [PubMed] [Google Scholar]
- ShangguanZ, Shao M, Dyckmans J.1999. Interaction of osmotic adjustment and photosynthesis in winter wheat under soil drought. Journal of Plant Physiology 154: 753–758. [Google Scholar]
- SharkeyTD.1990. Water stress effects on photosynthesis. Photosynthetica 24: 651. [Google Scholar]
- SharkeyTD, Badger MR.1982. Effects of water stress on photosynthetic electron transport, photophosphorylation and metabolite levels of Xanthium strumarum mesophyll cells. Planta 156: 199–206. [DOI] [PubMed] [Google Scholar]
- SharkeyTD, Seeman JR.1989. Mild water stress effects on carbon‐reduction cycle intermediates, ribulose bisphosphate carboxylase activity, and spatial homogeneity of photosynthesis in intact leaves. Plant Physiology 89: 1060–1065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- StuhlfaulthT, Scheuermann R, Fock HP.1990. Light energy dissipation under water stress conditions. Contribution of reassimilation and evidence for additional processes. Plant Physiology 92: 1053–1061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- TangAC, Kawamitsa Y, Kanechi M, & Boyer JS.2002. Photosynthesis at low water potentials in leaf discs lacking epidermis. Annals of Botany (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- TardieuF, Simmonneau T.1998. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. Journal of Experimental Botany 49: 419–432. [Google Scholar]
- TezaraW, Mitchell VJ, Driscoll SD, Lawlor DW.1999. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 401: 914–917. [Google Scholar]
- ValladaresF, Pearcy RW.1997. Interactions between water stress, sun‐shade acclimation, heat tolerance and photoinhibition in the sclerophyll Heteromeles arbutifolia Plant, Cell and Environment 20: 25–36. [Google Scholar]
- VuJCV, Yelenosky G.1988. Water deficit and associated changes in some photosynthetic parameters in leaves of ‘Valencia’ orange (Citrus sinensis [L.] Osbeck). Plant Physiology 88: 375–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- VuJCV, Allen Jr LH, Bowes G.1987. Drought stress and elevated CO2 effects on soybean ribulose bisphosphate carboxylase activity and canopy photosynthetic rates. Plant Physiology 83: 573–578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WinglerA, Quick WP, Bungard RA, Bailey KJ, Lea PJ, Leegood RC.1999. The role of photorespiration during drought stress: an analysis utilizing barley mutants with reduced activities of photorespiratory enzymes. Plant, Cell and Environment 22: 361–373. [Google Scholar]
- WongSC, Cowan IR, Farquhar GD.1979. Stomatal conductance correlates with photosynthetic capacity. Nature 282: 424–426. [Google Scholar]
- YounisHM, Boyer JS, Govindjee.1979. Conformation and activity of chloroplast coupling factor exposed to low chemical potential of water in cells. Biochimica et Biophysica Acta 548: 328–340. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}