

Abstract
Nitrification plays a central role in the nitrogen cycle by determining the oxidation state of nitrogen and its subsequent bioavailability and cycling. However, relatively little is known about the underlying ecology of the microbial communities that carry out nitrification in freshwater ecosystems—and particularly within high-altitude oligotrophic lakes, where nitrogen is frequently a limiting nutrient. We quantified ammonia-oxidizing archaea (AOA) and bacteria (AOB) in 9 high-altitude lakes (2289–3160 m) in the Sierra Nevada, California, USA, in relation to spatial and biogeochemical data. Based on their ammonia monooxygenase (amoA) genes, AOB and AOA were frequently detected. AOB were present in 88% of samples and were more abundant than AOA in all samples. Both groups showed >100 fold variation in abundance between different lakes, and were also variable through time within individual lakes. Nutrient concentrations (ammonium, nitrite, nitrate, and phosphate) were generally low but also varied across and within lakes, suggestive of active internal nutrient cycling; AOB abundance was significantly correlated with phosphate (r2 = 0.32, p<0.1), whereas AOA abundance was inversely correlated with lake elevation (r2 = 0.43, p<0.05). We also measured low rates of ammonia oxidation—indicating that AOB, AOA, or both, may be biogeochemically active in these oligotrophic ecosystems. Our data indicate that dynamic populations of AOB and AOA are found in oligotrophic, high-altitude, freshwater lakes.
Introduction
Nitrogen (N) is an essential nutrient for all life, and its availability serves as a critical factor for the growth of individual organisms, community composition, and ecosystem primary productivity in freshwater lakes [1], [2]. In many ecosystems, N availability—both quantity and chemical form—is largely dictated by microbial communities, which transform inorganic N into bioavailable forms, and actively cycle N through oxidation-reduction (redox) processes. Phosphorus (P) typically limits primary production in freshwater [3], but both absolute amounts, and relative ratios, of N and P are highly variable due to variations in lake nutrient sources, as well as internal cycling by phytoplankton, zooplankton, and microbes [1], [4], [5]. In oligotrophic aquatic systems, in particular, differences in size, growth rate, and chemical form of available nutrients may favor microorganisms in competition with phytoplankton for N [6]. Microbial control of both N quantity and chemical form has important implications for the degree of eutrophication in these ecosystems, and the degree to which allocthonous N inputs (i.e. atmospheric pollutants) may affect oligotrophic lakes [7].
Within the microbial N cycle, nitrification is a two-step process that involves the aerobic oxidation of reduced inorganic N compounds (i.e. NH3/NH4 +) to nitrite (NO2 -) and the subsequent oxidation of NO2 - to nitrate (NO3 -). Nitrification links the mineralization of N to its eventual removal as dinitrogen gas (N2) via either denitrification or anaerobic ammonium oxidation (anammox). The first step of nitrification is carried out by a few bacterial lineages within the Beta- and Gamma-proteobacteria and also by the archaeal phylum Thaumarchaeota (previously know as the group 1 Crenarchaeota) [8]. These ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA) use ammonia monoxygenase (AMO) to catalyze the oxidation of NH4 + to NO2 -. As AOA were confirmed to be capable of ammonia oxidation only recently [9], the physical and chemical factors that control the abundance and function of these organisms, and their relative influence on nitrification rates, are not entirely understood—particularly in freshwater environments [10]–[13].
AOA, AOB, and nitrification have been examined within few freshwater lakes, yet AOA appear to be important and dynamic components of lake plankton and biogeochemical cycles: Thaumarchaeota are abundant [14], most appear to be AOA [15], and their populations fluctuate over time [15] and with depth [16]. AOB are found in lakes ranging from temperate eutrophic, to high-altitude oligotrophic, but in contrast to AOA, how AOB abundance varies in lakes through space and time is not well known [17]–[19]. The abundances of AOA and AOB can be controlled by differential sensitivities to temperature [20], pH [21], [22], ammonium concentrations [23], and light [24]—all of which may be relevant in high elevation lakes, but have not been examined. Nitrification varies with depth and time, and is quantitatively important within lake water columns [10], [25]–[27]—for example, Finlay et al. [25] showed that within-lake production of NO3 - through nitrification is the predominant source of NO3 - in Lake Superior. However, a lone study has measured both AOA and ammonia oxidation rates in freshwater lakes [10], and AOB have rarely been quantified in lakes [28]. We therefore know little about variations in AOA and AOB abundance and activity over time and across different lakes—let alone how AOA, AOB, and ammonia oxidation rates respond to changes in temperature, N availability, and other environmental factors within freshwater systems.
Of particular relevance are the potential inhibitory effects of light on nitrification: while these have been known for some time (reviewed by [29]), the relative effects of different wavelengths of light on AOA versus AOB, and in the field versus lab, are mixed. AOA appear highly sensitive to light in controlled experiments [24], [30]: the AOA Nitrosopumilus maritimus and Nitrosotalea devanaterra were inhibited by lower light levels than AOB, and showed little recovery of ammonia oxidation over 8/16 hour light/dark cycles [30]. French et al. [24] likewise found that ammonia oxidation by three freshwater AOA isolates was strongly inhibited by white and blue light, whereas an AOB isolate was inhibited only by blue light and recovered partial oxidation ability in the dark. Notably, all of the AOA isolates used in these studies have been recovered from sediments or soil, and it is possible that pelagic AOA are less light-sensitive—for example, Auguet and Casamayor [14] proposed that surface waters of mountain lakes are an archaeal ‘hotspot’ based on high crenarchaeal abundance in the neuston. AOA also actively express amoA, and nitrification is known to occur at least transiently, in the upper ocean [31]–[33].
We quantified the abundance of AOA and AOB across a high-elevation lake transect in Yosemite National Park, in the Sierra Nevada mountain range, California, USA (Figure 1). In Sierra Nevada lakes, the differing susceptibility of AOA and AOB to photoinhibition could be a crucial factor in N cycling, as there is a strong natural increase in ultraviolet (UV) radiation with increasing elevation [26]. Moreover, these high-altitude lakes have relatively low light attenuation due to high transparency typical of oligotrophic aquatic ecosystems found at high elevations [26], [34]. Freshwater lakes are traditionally limited by phosphorous availability [35], but the availability of N is also a critical factor for primary productivity in aquatic ecosystems of the Sierra Nevada, where biological activity in ∼22% of lakes is strictly limited by N availability [36]. Internal N cycling may therefore play an important role in the overall productivity and structure of these freshwater ecosystems. We used natural variations in temperature, radiation, and N deposition, based on elevational and temporal variability between sampling sites, to examine the prevalence and abundance of AOA and AOB in high-altitude lakes.
Figure 1. Sampling locations (white circles) in Yosemite National Park displayed on 10 m resolution elevation data from the United States Geological Survey National Elevation Dataset (http://nationalmap.gov/).

Inset: location of Yosemite shown as the green shaded area within the state of California.
Materials and Methods
Study Site
The Sierra Nevada (California) is a 400-mile long mountain range that gradually rises from the valley floor from west to east and reaches an apex of 3,000 to 4,200 meter peaks on its eastern edge (Figures 1 and 2). Vegetation along the mountain range is composed of grasslands and foothill woodlands at lower elevations, with a transition to mixed conifer forests, and then alpine meadows and lakes at higher elevations. Aquatic ecosystems in the Sierra Nevada are located downwind of urban and agricultural areas that emit high levels of N [37] and so experience elevated levels of N deposition [38]. This N deposition is known to increase N concentrations in lakes [7], and in the case of the Sierra Nevada, represents a large fraction of the N input to high-elevation lakes: Baron et al. [39] established a critical load threshold of 1.5 kg N ha−1 year−1 for high-elevation lakes located in Rocky Mountain National Park, yet current annual N loading in the Sierra Nevada (i.e. Emerald Lake Watershed, Sequoia National Park) ranges from 2.0 to 4.9 kg N ha−1 year−1 [40]. Moreover, the watersheds are high in granitic parent material and generally have thin soils; both characteristics cause aquatic ecosystems in the Sierra Nevada to have limited buffering capacity in terms of their ability to neutralize foreign chemical species [41]. Clow et al. [42] suggested that this property, coupled with high precipitation at high elevations, leads to high N loading at high elevations despite greater distances from emission sources. Our data are relevant to this as our selected sites range from 2300 m (Harden Lake) to 3160 m (Upper Gaylor Lake) (Table 1 and Figure 2) and our transect terminus is adjacent to the steep Sierra escarpment (Figure 1).
Figure 2. Lake elevation plotted against longitude for the nine lakes sampled in this study.
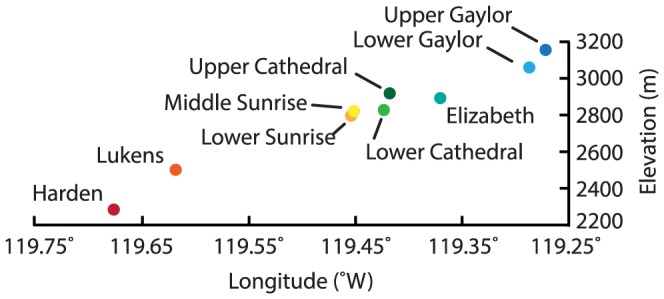
Table 1. Lake names, elevation, depth, and average nutrient concentrations.
| Lake | Elevatio (m) | Dept (m) | Average PO4 3- (nM) | Average NH4 + (nM) | Average NO2 - (nM) | Average NO3 - (nM) |
| Harden | 2289 | 4.5 | 80 | 15 | 44 | 297 |
| Lukens | 2506 | 6 | 98 | 32 | 41 | 374 |
| Lower Sunrise | 2801 | 5.5 | 74 | 13 | 38 | 432 |
| Middle Sunrise | 2826 | 5 | 116 | 25 | 52 | 212 |
| Lower Cathedral | 2832 | 10 | 73 | 13 | 28 | 281 |
| Upper Cathedral | 2923 | 3.5 | 91 | 20 | 67 | 419 |
| Elizabeth | 3050 | 9 | 84 | 10 | 36 | 232 |
| Lower Gaylor | 3064 | 11 | 81 | 41 | 33 | 398 |
| Upper Gaylor | 3160 | 7 | 79 | 20 | 26 | 307 |
Sampling
Water samples were collected from lakes located in Yosemite National Park (YNP) during the 2012 summer. Nine lakes, which ranged in elevation from 2289 m to 3160 m and in depth from 4.5 m to 11 m (Table 1), were sampled in June, July, August, and September. Water samples were collected from the middle of each lake at 1 m depth using a Van Dorn water sampler (Lamotte); this depth was selected to include the effects of UV radiation and high photosynthetically active radiation (PAR). Duplicate water samples were filtered in the field onto 0.22 µm PVDF Membrane Filters (Millipore) using sterile 60 mL Polycarbonate syringes (Cole-Parmer). After filtration, filter membranes were packed into bead tubes (MP Biomedicals) containing 800 µL of Sucrose-Tris-EDTA. Filtrate was collected in 60 mL HDPE Bottles (Nalgene) for subsequent nutrient analysis. Samples were transported on ice to UC Merced's main campus or Yosemite Field Station (within hours of collection) and stored at −80° or −20°C until extraction of DNA and nutrient analysis. Samples were collected under USA National Park Service permit YOSE-2012-SCI-0111.
DNA Extraction and quantification and real-time QPCR Analysis
DNA extraction followed Beman et al. [31] using Sucrose-Tris-EDTA (STE) lysis buffer, sodium dodecyl sulfate (SDS), and proteinase K with bead-beating. DNA was further purified using a DNeasy Blood & Tissue Extraction Kit (Qiagen) and resolubolized in 50 µL of ultra-pure DNA free water (Qiagen). DNA was quantified using a PicoGreen dsDNA quantification kit (Invitrogen) and an Mx3005P real-time thermocycler (Agilent Technologies). Total yield of DNA ranged from 23.5 to 922 ng.
Quantitative Polymerase Chain Reaction (QPCR) was used to quantify the abundance of amoA genes in lake samples. Primers, reaction chemistry, thermocycling, QPCR standards, quality control procedures, and data analysis exactly followed Beman et al. [31] and Beman et al. [43]. In brief, we used SYBR Green chemistry and the primers crenamoAF (5′-STAATGGTCTGGCTTAGACG-3′) and crenamoAR (5′-GCGGCCATCCATCTGTATGT-3′) for archaeal amoA (originally Arch-amoAF and R; [44]) and beta-amoA1F (5′-GGGGTTTCTACTGGTGGT-3′) and beta-amoA2R (5′-CCCCTCKGSAAAGCCTTCTTC-3′) for betaproteobacterial amoA (originally amoA-1F and amoA-2R [45]). QCPR efficiencies ranged from 89–91%, standard r2 values ranged from 0.98 to 0.99, and we tested for inhibition (which was not detected) by ‘spiking’ standards with samples [31], [43].
Nutrient Analyses
Filtered lake water samples were analyzed for orthophosphate (µmol l−1) nitrite (µmol l−1) and nitrate (µmol l−1) using flow-injection analysis on a QuikChem 8000 (Zellweger Analytics, Inc.) at the University of California, Santa Barbara Marine Sciences Institute Analytical Laboratory (standard curve r2 = 0.999 for all assays). Filtered lake water samples were analyzed for ammonium (NH4 +) following Holmes et al. [46]. 8 mL of filtered water was combined with 2 mL of reagent that consisted of 95% 0.1 M sodium tetraborate, 0.015 M O-pthaldialdehyde, 0.03 mM sodium sulfite, and 5% Ethanol. After aging, samples were measured in triplicate for fluorescence intensity using a fluorometer (Trilogy Laboratory Fluorometer, Turner Designs). Standards ranged from 31.2 to 186.8 nM and for different runs, standard curve r2 = 0.998–0.999.
15NH4 + oxidation rate measurements
Ammonia oxidation rates were measured by adding 99 atom percent (at%) 15NH4 + to a concentration of 200 nmol L−1, and measuring the accumulation of 15N label in the oxidized NO2 - + NO3 - pool after incubation for ∼24 hours [31], [47]. All samples were incubated within lakes to mimic in situ conditions as accurately as possible. δ 15N of NO2 - + NO3 - was measured at the UC Davis Stable Isotope Facility using the ‘denitrifier method’ [48], which produces N2O that can be analyzed on the mass spectrometer. Isotopic reference materials bracketed every 3–4 samples and coefficients of variation for these were 0.6%.
Initial at% enrichment of the substrate at the beginning of the experiment (noNH4 +, see Eq. 1) was calculated by isotope mass balance based on NH4 + concentrations determined fluorometrically [46] assuming that the 15N activity of unlabeled NH4 + was 0.3663 at% 15N. Rates of ammonia oxidation (15Rox) were calculated using equation 1 [31]:
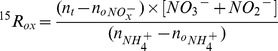 |
(eq.1) |
where nt is the at% 15N in the NO3 - + NO2 - pool measured at time t, noNOx -, is the measured at% 15N of unlabeled NO3 - + NO2 -, noNH4+ is the initial at% enrichment of NH4 + at the beginning of the experiment, n NH4+ is at% 15N of NH4 + at time t, and [NO3 - + NO2 -] is the concentration of the NOx - pool.
Statistical analyses
Statistical analyses were performed using the R statistical environment (http://www.r-project.org/) and the vegan package.
Results and Discussion
AOB amoA genes were detected in all lakes from June to September, and in 88% of samples, whereas AOA were found in 46% of all samples. AOB amoA gene copies ranged from 3.04×102 to 2.07×105 genes mL−1, however the majority (73%) of AOB values fell below 2×104 genes mL−1, with values exceeding this occurring in June, August and September at high elevations (Figure 3B). AOA ranged from 0 to 4.58×103 genes mL−1 (See Figure 3A) and there were zero instances where AOA outnumbered AOB. When AOA were detected, they were outnumbered by AOB by 2.8- to 1080-fold. Highest average abundances of both AOB and AOA were found in Middle Sunrise Lake (2826 m), while lowest average AOB abundances were found in Lukens Lake (2506 m), Lower Cathedral Lake (2832 m) and Lake Elizabeth (3019 m) (see Figure 3). For AOA, multiple lakes had low average values, including Lukens, Lower Cathedral, Elizabeth, and Lower Gaylor (3064 m). Over time, the highest numbers of amoA genes were present in September for AOB (4 out of 6 lakes above 2800 m), and in June for AOA (5 out of 6 lakes above 2800 m). AOB and AOA therefore appear to be present, and sometimes abundant, within the water columns of oligotrophic lakes in Yosemite National Park.
Figure 3. Boxplot comparison of number of amoA genes per milliliter (genes mL−1) for (A) ammonia-oxidizing archaea (AOA) and (B) ammonia-oxidizing bacteria (AOB).

The vertical axes (logarithimic scale) denote the number of genes mL−1; the horizontal axes represent the elevation of the sampling sites, and are ordered from lowest elevation (Harden Lake) to highest elevation (Upper Gaylor Lake). In these plots, the box denotes the mean plus and minus one standard deviation; the line within the box represents the median value; and lines extending above and below the box span the full range of the data. Outliers were defined as sample values 1.5 times larger than the upper quartile and are represented by green and blue circles.
Previous studies that have quantified AOA abundance in high-altitude oligotrophic lakes reported amoA gene abundances as high as 3×104 genes mL−1 [15], while AOB were below detection limits in the same lakes [49]. Our results indicate that AOB are more prevalent and abundant than AOA within oligotrophic lakes of the Sierra Nevada, CA, and AOB were dominant regardless of lake nutrient concentrations (see below), date of sampling, or lake elevation. This contrasts with earlier work that found AOA were more prevalent (detected via PCR but not quantified by QPCR) than AOB in oligotrophic lakes of the Tibetan plateau [18], and dominant in oligotrophic lakes of the Spanish Pyrenees [49]. In two contrasting lower elevation (231–825 m) lakes in France, Hugoni et al. [28] reported AOA dominance under low ammonium concentrations and oligotrophic conditions, whereas AOB were dominant in nutrient-rich waters [28]. The lakes sampled in Yosemite are uniformly nutrient-poor and have particularly low ammonium concentrations (all <75 nM), however our data indicate that AOB amoA genes are more abundant and prevalent than those from AOA. One explanation for AOB dominance in these lakes is that the amoA primers used to detect AOB and AOA may over- or under-estimate their abundances. However, the AOB primers used here are specific for betaproteobacterial AOB and are widely used [50]; the AOA primers amplify a wide range of AOA groups from water, sediments, and soils [44]. It is therefore unlikely that the AOB primers severely overestimate AOB amoA genes or that the AOA primers severely underestimate AOA amoA genes. Nor would this explain the prevalence of AOB, which were frequently detected. AOA were also detected in nearly half the samples—despite high light levels and oligotrophic conditions that are presumably hostile to nitrifiers in general.
Ultimately AOB and AOA must oxidize N to persist under oligotrophic conditions, and we suggest that their populations could be sustained by N fluxes that are not reflected in depleted nutrient pools: that is, NH4 + may be rapidly regenerated, assimilated, and/or oxidized, but because of high demand, does not accumulate in oligotrophic lakes. In fact, lakes are watershed ‘integrators’ [51] that can function as hotspots of N-cycling, including nitrification [14], [25], [42], [52]. In Yosemite, lake N loading increases with elevation, and NO3 - concentrations are correlated with modeled N deposition rates [42]. N deposition to Sierra watersheds occurs primarily as NH4 + [37] and this flux of N—which can be comparatively large in these oligotrophic ecosystems [53]—must have been nitrified at least once if it accumulates as NO3 -. Ammonium concentrations therefore may not be the sole predictor for AOA and AOB abundances—elevation and nitrate concentrations may also be relevant—and we analyzed relationships between AOA and AOB and several types of spatial and nutrient concentration data using a variety of statistical approaches.
During the sampling period NO3 - concentrations in all lakes ranged from 150 nM to 990 nM, PO4 3- concentrations from 60 nM to 120 nM, NO2 - concentrations from 20 nM to 120 nM, and NH4 + concentrations from 2.8 nM to 72 nM (Figure 4). As expected in aquatic ecosystems, NO3 - levels were higher than either PO4 3- or NO2 -. We did not observe a significant trend with elevation (ANOVA P>0.05), but this could emerge with additional sampling. In nearly all lakes, the molar concentration of PO4 3- was higher than that of nitrite, which is typical, as NO2 - is quickly oxidized to NO3 - in the presence of oxygen [54]. Across our samples, AOB and AOA were significantly correlated with a few individual variables, including nutrient concentrations. For example, AOB abundance was most strongly correlated with PO4 3- concentrations (r2 = 0.32, p<0.1), consistent with work by Sundaweshar et al. [55] that showed P-limitation of N-cycling. AOB displayed an increasing trend with altitude, but this relationship was not significant (r2 = 0.21, p = 0.28); in contrast, AOA abundance was inversely correlated with altitude (r2 = 0.43, p<0.05). Both groups showed wide variation in Middle Sunrise Lake at 2826 m. These patterns are evident in Figure 3, where AOB were notably more abundant—but also variable—in the Gaylor Lakes at >3000 m elevation, whereas AOA were more abundant and variable at lower elevations. For all samples collected in the three lakes >3000 m elevation, AOA were only detected four times. This inverse relationship between AOA abundance and increasing elevation could reflect the effect of increased UV radiation at higher elevations, or other factors that vary with elevation.
Figure 4. Variation in lake nutrient concentrations versus elevation for (A) phosphate (PO4 3-), (B) ammonium (NH4 +), (C) nitrite (NO2 -), and (D) nitrate (NO3 -).

Vertical axes show nutrient concentrations in nanomolar (nM) plotted on a logarithmic scale, and colors denote month of sampling. Note that some samples have highly similar nutrient concentrations and fall nearly on top of one another.
We therefore used redundancy analysis (RDA) to analyze multivariate relationships between AOA and AOB abundance and spatial (elevation and longitude), temporal (sampling date) and environmental (NO3 -, NO2 -, NH4 + and PO4 3-) data (Figure 5). 33% of the variability in AOA and AOB abundance was explained by these data, with nutrient concentrations accounting for 24% of the variability in AOA and AOB abundance, and site location and sampling date accounting for 9% of the variability. Collectively, nutrients explain nearly a quarter of the variation in AOA and AOB abundance in these lakes, and this includes ammonium, as well as nitrate, nitrite, and phosphate, concentrations. The modest percentage of constrained variance overall indicates that AOA and AOB populations are affected by other, un-measured factors, or this may reflect stochastic variation in populations. In lakes, AOB and AOA abundance may be modified by active growth, but also by transport of cells into lakes via air, water, or suspended particles; competition with other organisms for ammonium; and trophic interactions, such as grazing, viral infection, and lysis. None of these have been directly investigated.
Figure 5. Bi-plot of redundancy analysis of bacterial community composition.

Color key denotes different lakes, and arrows denote biplot scores for the constraining variables. The grey box encompasses 17 samples that fall within a narrow range of each other and are not visually distinguishable on the bi-plot; this includes 1–2 samples from every lake except Middle Sunrise and Upper Cathedral Lake. One sample from Middle Sunrise Lake with extremely high AOB amoA gene abundance is not shown, as it falls much farther along the RDA1 and RDA2 axes.
The prevalence and abundances of AOA and AOB suggest that active N cycling and nitrification may be occurring in high-altitude lakes, but is surprising given that high-altitude oligotrophic lakes have low N concentrations and experience high light levels. To determine whether AOA and AOB may be active under these conditions, we performed 15NH4 + incubations to detect ammonia oxidation rates in three lakes that span a range of elevations and AOB/AOA abundances (Lukens Lake, Lower Cathedral Lake and Lower Gaylor Lake). These were conducted in both light and dark bottles that were incubated within the lakes under in situ conditions. At low N concentrations found in oligotrophic waters, measuring ammonia oxidation is extremely challenging due to multiple factors: (1) measuring low-level N concentrations is difficult, and uncertainties in basic nutrient concentration values can strongly affect rate calculations; (2) addition of 15N label can significantly increase N concentrations and potentially introduce biases; (3) rates are expected to be low due to low N concentrations; and (4) measuring accurate isotopic values becomes difficult, which either introduces errors in the measurement or requires addition of N ‘carrier’—yet this can make detection of low rates difficult or impossible. For these reasons, we propagated uncertainties in nutrient measurements and isotopic values through our calculations, and found that rates were above detection (limit = 10.2 pmol L−1 d−1) only in dark bottles in Lower Cathedral Lake and Lower Gaylor Lake—where they were extremely low (Figure 6). Ammonia oxidation was below the limits of detection in most light bottles in these lakes, as well as in Lukens Lake. That rates were undetectable under in situ light levels is consistent with light inhibition of ammonia oxidation, but more importantly indicates that AOB and AOA were inactive—or active at extremely low levels—at the time and location of sampling. Our data do suggest that they respond to changing light conditions and oxidize ammonia at low levels in completely darkened bottles. This expands the range of habitats in which ammonia oxidation can potentially occur to include surface waters of high-altitude, oligotrophic, freshwater lakes—but when and where these organisms are active at appreciable levels is not yet clear.
Figure 6. Boxplot comparison of calculated 15NH4 + oxidation rates measured in Lower Cathedral and Lower Gaylor lakes.
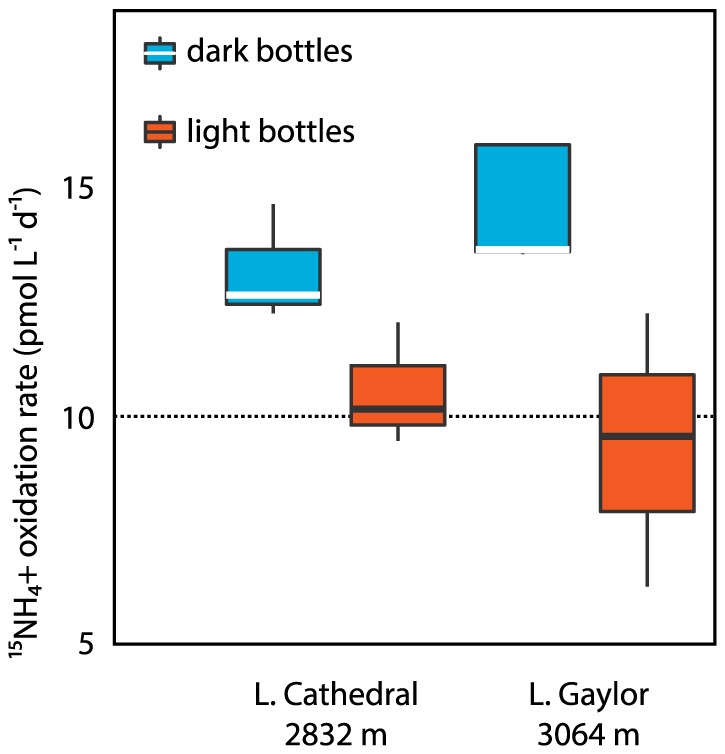
The vertical axis shows 15NH + oxidation rates (pmol L−1 D−1). The blue bars represent dark bottle incubations that were incubated under zero-light conditions, while the orange bars represent samples that were exposed to ambient light over a 24-hour cycle during in-situ incubation at the sampling site. The dashed line notes the detection limit. In these plots, the box denotes the mean plus and minus one standard deviation; the line within the box represents the median value; and lines extending above and below the box span the full range of the data.
AOB and AOA were frequently present, varied in abundance between lakes, and fluctuated over time within individual lakes. The presence of AOA is consistent with the idea that they are adapted to the low-nutrient conditions that are characteristic of these lakes—for example, the marine AOA Nitrosopumilus maritimus has a remarkably high affinity for ammonia and appears to be adapted to life under extreme nutrient limitation [23]. However, AOB were more commonly detected and were more abundant than AOA under all sampled conditions; this fits with the recent idea that AOB are more light-tolerant than AOA [24], [30]. High altitude lakes could also experience higher fluxes of N that would periodically favor AOB [42]. Both AOA and AOB may contribute to nitrification in freshwater oligotrophic lakes—as we detected low rates of ammonia oxidation in darkened bottles—but it is clear that they were inactive, or active only at low levels, during our incubations. Our data therefore add to the limited information available on microbial contributions to N cycling in lakes—particularly for AOA/AOB and nitrification—but additional work should expand these approaches to additional lakes and additional sampling periods. Nitrification under ice could be important during winter, for example, and nitrification may be especially significant following spring snowmelt, when pulses of N likely enter these lakes and may be metabolized by microbes. Altogether our findings are indicative of dynamic microbial communities and internal N cycling in high-altitude lakes.
Acknowledgments
We thank Susan Alford, Julie Cline, Matt Meyerhof, Elizabeth Perkins, Sang Park, and Victoria Velez for help in the field and lab.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.
Funding Statement
This work was supported University of California start-up funds (to JMB) and the UC Merced Sierra Nevada Research Institute (to JMB). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Elser JJ, Bracken MES, Cleland EE, Gruner DS, Harpole WS, et al. (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters 10: 1135–1142. [DOI] [PubMed] [Google Scholar]
- 2. Vrede T, Ballantyne A, Mille-Lindblom C, Algesten G, Gudasz C, et al. (2009) Effects of N: P loading ratios on phytoplankton community composition, primary production and N fixation in a eutrophic lake. Freshwater Biology 54: 331–344. [Google Scholar]
- 3. Schindler DW (1977) Evolution of Phosphorus Limitation in Lakes. Science 195: 260–262. [DOI] [PubMed] [Google Scholar]
- 4. Elser JJ, Andersen T, Baron JS, Bergstrom AK, Jansson M, et al. (2009) Shifts in Lake N: P Stoichiometry and Nutrient Limitation Driven by Atmospheric Nitrogen Deposition. Science 326: 835–837. [DOI] [PubMed] [Google Scholar]
- 5. Harpole WS, Ngai JT, Cleland EE, Seabloom EW, Borer ET, et al. (2011) Nutrient co-limitation of primary producer communities. Ecology Letters 14: 852–862. [DOI] [PubMed] [Google Scholar]
- 6. Cotner JB, Biddanda BA (2002) Small players, large role: Microbial influence on biogeochemical processes in pelagic aquatic ecosystems. Ecosystems 5: 105–121. [Google Scholar]
- 7. Fenn ME, Baron JS, Allen EB, Rueth HM, Nydick KR, et al. (2003) Ecological effects of nitrogen deposition in the western United States. Bioscience 53: 404–420. [Google Scholar]
- 8. Brochier-Armanet C, Boussau B, Gribaldo S, Forterre P (2008) Mesophilic crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nature Reviews Microbiology 6: 245–252. [DOI] [PubMed] [Google Scholar]
- 9. Konneke M, Bernhard AE, de la TorreJR, Walker CB, Waterbury JB, et al. (2005) Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437: 543–546. [DOI] [PubMed] [Google Scholar]
- 10. Small GE, Bullerjahn GS, Sterner RW, Beall BFN, Brovold S, et al. (2013) Rates and controls of nitrification in a large oligotrophic lake. Limnology and Oceanography 58: 276–286. [Google Scholar]
- 11. Erguder TH, Boon N, Wittebolle L, Marzorati M, Verstraete W (2009) Environmental factors shaping the ecological niches of ammonia-oxidizing archaea. Fems Microbiology Reviews 33: 855–869. [DOI] [PubMed] [Google Scholar]
- 12. Francis CA, Beman JM, Kuypers MMM (2007) New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation. Isme Journal 1: 19–27. [DOI] [PubMed] [Google Scholar]
- 13. Hatzenpichler R (2012) Diversity, Physiology, and Niche Differentiation of Ammonia-Oxidizing Archaea. Applied and Environmental Microbiology 78: 7501–7510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Auguet JC, Casamayor EO (2008) A hotspot for cold crenarchaeota in the neuston of high mountain lakes. Environmental Microbiology 10: 1080–1086. [DOI] [PubMed] [Google Scholar]
- 15. Auguet JC, Nomokonova N, Camarero L, Casamayor EO (2011) Seasonal Changes of Freshwater Ammonia-Oxidizing Archaeal Assemblages and Nitrogen Species in Oligotrophic Alpine Lakes. Applied and Environmental Microbiology 77: 1937–1945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Callieri C, Corno G, Caravati E, Rasconi S, Contesini M, et al. (2009) Bacteria, Archaea, and Crenarchaeota in the epilimnion and hypolimnion of a deep holo-oligomictic lake. Applied and Environmental Microbiology 75: 7298–7300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Whitby CB, Saunders JR, Pickup RW, McCarthy AJ (2001) A comparison of ammonia-oxidiser populations in eutrophic and oligotrophic basins of a large freshwater lake. Antonie Van Leeuwenhoek 79: 179–188. [DOI] [PubMed] [Google Scholar]
- 18. Hu AY, Yao TD, Jiao NZ, Liu YQ, Yang Z, et al. (2010) Community structures of ammonia-oxidising archaea and bacteria in high-altitude lakes on the Tibetan Plateau. Freshwater Biology 55: 2375–2390. [Google Scholar]
- 19. Cebron A, Coci M, Garnier J, Laanbroek HJ (2004) Denaturing gradient gel electrophoretic analysis of ammonia-oxidizing bacterial community structure in the lower Seine River: impact of Paris wastewater effluents. Appl Environ Microbiol 70: 6726–6737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Tourna M, Freitag TE, Nicol GW, Prosser JI (2008) Growth, activity and temperature responses of ammonia‚Äêoxidizing archaea and bacteria in soil microcosms. Environmental Microbiology 10: 1357–1364. [DOI] [PubMed] [Google Scholar]
- 21. Nicol GW, Leininger S, Schleper C, Prosser JI (2008) The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environmental Microbiology 10: 2966–2978. [DOI] [PubMed] [Google Scholar]
- 22. Auguet JC, Casamayor EO (2013) Partitioning of Thaumarchaeota populations along environmental gradients in high mountain lakes. Fems Microbiology Ecology 84: 154–164. [DOI] [PubMed] [Google Scholar]
- 23. Martens-Habbena W, Berube PM, Urakawa H, de la TorreJR, Stahl DA (2009) Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 461: 976–U234. [DOI] [PubMed] [Google Scholar]
- 24. French E, Kozlowski JA, Mukherjee M, Bullerjahn G, Bollmann A (2012) Ecophysiological Characterization of Ammonia-Oxidizing Archaea and Bacteria from Freshwater. Applied and Environmental Microbiology 78: 5773–5780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Finlay JC, Sterner RW, Kumar S (2007) Isotopic evidence for in-lake production of accumulating nitrate in lake superior. Ecological Applications 17: 2323–2332. [DOI] [PubMed] [Google Scholar]
- 26. Hall GH, Jeffries C (1984) The Contribution of Nitrification in the Water Column and Profundal Sediments to the Total Oxygen Deficit of the Hypolimnion of a Mesotrophic Lake (Grasmere, English Lake District). Microbial Ecology 10: 37–46. [DOI] [PubMed] [Google Scholar]
- 27. Rudd JWM, Kelly CA, Schindler DW, Turner MA (1988) Disruption of the Nitrogen-Cycle in Acidified Lakes. Science 240: 1515–1517. [DOI] [PubMed] [Google Scholar]
- 28. Hugoni M, Etien S, Bourges A, Lepere C, Domaizon I, et al. (2013) Dynamics of ammonia-oxidizing Archaea and Bacteria in contrasted freshwater ecosystems. Research in Microbiology 164: 360–370. [DOI] [PubMed] [Google Scholar]
- 29. Lomas MW, Lipschultz F (2006) Forming the primary nitrite maximum: Nitrifiers or phytoplankton? Limnology and Oceanography 51: 2453–2467. [Google Scholar]
- 30. Merbt SN, Stahl DA, Casamayor EO, Marti E, Nicol GW, et al. (2012) Differential photoinhibition of bacterial and archaeal ammonia oxidation. FEMS Microbiology Letters 327: 41–46. [DOI] [PubMed] [Google Scholar]
- 31. Beman JM, Popp BN, Alford SE (2012) Quantification of ammonia oxidation rates and ammonia-oxidizing archaea and bacteria at high resolution in the Gulf of California and eastern tropical North Pacific Ocean. Limnology and Oceanography 57: 711–726. [Google Scholar]
- 32. Church MJ, Wai B, Karl DM, DeLong EF (2010) Abundances of crenarchaeal amoA genes and transcripts in the Pacific Ocean. Environmental Microbiology 12: 679–688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Santoro AE, Casciotti KL, Francis CA (2010) Activity, abundance and diversity of nitrifying archaea and bacteria in the central California Current. Environmental Microbiology 12: 1989–2006. [DOI] [PubMed] [Google Scholar]
- 34. Sommaruga R (2001) The role of solar UV radiation in the ecology of alpine lakes. Journal of Photochemistry and Photobiology B-Biology 62: 35–42. [DOI] [PubMed] [Google Scholar]
- 35. Sterner RW (2008) On the Phosphorus Limitation Paradigm for Lakes. International Review of Hydrobiology 93: 433–445. [Google Scholar]
- 36.Eilers JM, Kanciruk P, McCord RA, Overton WS, Hook L, et al.. (1987) Characteristics of Lakes in the Western United States. Volume II: Data Compendium of Selected Physical and Chemical Variables. In: Agency UEP, editor.
- 37.Clarisse L, Shephard MW, Dentener F, Hurtmans D, Cady-Pereira K, et al.. (2010) Satellite monitoring of ammonia: A case study of the San Joaquin Valley. Journal of Geophysical Research-Atmospheres 115.
- 38. Bytnerowicz A, Tausz M, Alonso R, Jones D, Johnson R, et al. (2002) Summer-time distribution of air pollutants in Sequoia National Park, california. Environmental Pollution 118: 187–203. [DOI] [PubMed] [Google Scholar]
- 39. Baron JS (2006) Hindcasting nitrogen deposition to determine an ecological critical load (vol 16, pg 433, 2006). Ecological Applications 16: 1629–1629. [DOI] [PubMed] [Google Scholar]
- 40. Sickman JO, Leydecker A, Melack JM (2001) Nitrogen mass balances and abiotic controls on N retention and yield in high-elevation catchments of the Sierra Nevada, California, United States. Water Resources Research 37: 1445–1461. [Google Scholar]
- 41. Meixner T, Gutmann C, Bales R, Leydecker A, Sickman J, et al. (2004) Multidecadal hydrochemical response of a Sierra Nevada watershed: sensitivity to weathering rate and changes in deposition. Journal of Hydrology 285: 272–285. [Google Scholar]
- 42.Clow DW, Nanus L, Huggett B (2010) Use of regression-based models to map sensitivity of aquatic resources to atmospheric deposition in Yosemite National Park, USA. Water Resources Research 46.
- 43. Beman JM, Sachdeva R, Fuhrman JA (2010) Population ecology of nitrifying Archaea and Bacteria in the Southern California Bight. Environmental Microbiology 12: 1282–1292. [DOI] [PubMed] [Google Scholar]
- 44. Francis CA, Roberts KJ, Beman JM, Santoro AE, Oakley BB (2005) Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proceedings of the National Academy of Sciences of the United States of America 102: 14683–14688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Rotthauwe JH, Witzel KP, Liesack W (1997) The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl Environ Microbiol 63: 4704–4712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Holmes RM, Aminot A, Kerouel R, Hooker BA, Peterson BJ (1999) A simple and precise method for measuring ammonium in marine and freshwater ecosystems. Canadian Journal of Fisheries and Aquatic Sciences 56: 1801–1808. [Google Scholar]
- 47.Ward BB (2008) Chapter 5 - Nitrification in Marine Systems. In: Capone DG, Bronk DA, Mulholland MR, Carpenter EJ, editors. Nitrogen in the Marine Environment (2nd Edition). San Diego: Academic Press.pp.199–261. [Google Scholar]
- 48. Sigman DM, Casciotti KL, Andreani M, Barford C, Galanter M, et al. (2001) A bacterial method for the nitrogen isotopic analysis of nitrate in seawater and freshwater. Analytical Chemistry 73: 4145–4153. [DOI] [PubMed] [Google Scholar]
- 49. Auguet JC, Triado-Margarit X, Nomokonova N, Camarero L, Casamayor EO (2012) Vertical segregation and phylogenetic characterization of ammonia-oxidizing Archaea in a deep oligotrophic lake. Isme Journal 6: 1786–1797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Junier P, Kim OS, Hadas O, Imhoff JF, Witzel KP (2008) Evaluation of PCR primer selectivity and phylogenetic specificity by using amplification of 16S rRNA genes from betaproteobacterial ammonia-oxidizing bacteria in environmental samples. Applied and Environmental Microbiology 74: 5231–5236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Williamson CE, Saros JE, Vincent WF, Smol JP (2009) Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnology and Oceanography 54: 2273–2282. [Google Scholar]
- 52. Baron JS, Hall EK, Nolan BT, Finlay JC, Bernhardt ES, et al. (2013) The interactive effects of excess reactive nitrogen and climate change on aquatic ecosystems and water resources of the United States. Biogeochemistry 114: 71–92. [Google Scholar]
- 53.Murphy DD, Knopp CM (2000) Lake Tahoe watershed assessment: volume I. General Technical Report-Pacific Southwest Research Station, USDA Forest Service.
- 54.Goldman CR, Horne AJ (1983) Limnology: McGraw-Hill.
- 55. Sundareshwar PV, Morris JT, Koepfler EK, Fornwalt B (2003) Phosphorus limitation of coastal ecosystem processes. Science 299: 563–565. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.