

Abstract
Lymphatic filariasis affects 120 million people worldwide and another 1.2 billion people are at risk of acquiring the infection. Chemotherapy with mass drug administration is substantially reducing the incidence of the infection. Nevertheless, an effective vaccine is needed to prevent the infection and eradicate the disease. Previously we reported that a multivalent fusion protein vaccine (rBmHAT) composed of small heat shock proteins 12.6 (HSP12.6), abundant larval transcript-2 (ALT-2) and large extracellular domain of tetraspanin (TSP LEL) could confer >95% protection against the challenge infection with Brugia malayi infective larvae (L3) in mouse and gerbil models. In this study we evaluated the immunogenicity and efficacy of rBmHAT fusion protein vaccine in a rhesus macaque model. Our results show that rBmHAT is highly immunogenic in rhesus macaques. All the vaccinated monkeys developed significant titers of antigen-specific IgG antibodies against each of the component antigens (16,000 for rBmHSP12.6), (24,000 for rBmALT-2) and (16,000 for rBmTSP-LEL). An in vitro antibody dependent cellular cytotoxicity (ADCC) assay performed using the sera samples from vaccinated monkeys showed that the anti-rBmHAT antibodies are functional with 35% killing of B. malayi L3s. Vaccinated monkeys also had antigen responding cells in the peripheral blood. Vaccine-induced protection was determined after challenging the monkeys with 500 B. malayi L3. Following challenge infection, 3 out of 5 vaccinated macaques failed to develop the infection. These three protected macaques had high titers of IgG1 antibodies and their PBMC secreted significantly high levels of IFN-γ in response to the vaccine antigens. The two vaccinated macaques that picked the infection had slightly low titers of antibodies and their PBMC secreted high levels of IL-10. Based on these findings we conclude that the rBmHAT vaccine is highly immunogenic and safe and can confer significant protection against challenge infections in rhesus macaques.
Introduction
Human lymphatic filariasis caused mainly by Wuchereria bancrofti and Brugia malayi affects 120 million people and elicits a wide spectrum of pathological disorders of the lymphatic system with varied clinical manifestations. The filarial parasites can survive in the human for many years causing permanent disability due to chronic syndromes such as lymphoedema, elephantiasis, and hydrocoele. According to the World Health Organization, lymphatic filariasis is the second leading cause of physical disability in the world [1]. Certain individuals who live in the endemic regions remain retractile to filarial infection and carry high titer of circulating antibodies against select parasite antigens [2]. Previous studies showed that these serum antibodies are cytotoxic to infective larvae of the parasite (L3), suggesting that these individuals are naturally immune to the infection [3], [4]. These putatively immune individuals are called Endemic Normal (EN) [2]. Infected individuals do not carry these host protective antibodies [4]. Using an iterative screening of a phage cDNA expression library of the parasite with the sera from EN subjects, we identified several parasite antigens that were specifically recognized by the antibodies in the sera of EN subjects [5]. Subsequently, these antigens were cloned and their vaccine potential was evaluated in a mouse and/or jird models [3], [4], [6]–[8]. These studies identified three antigens [small heat shock proteins 12.6 (HSP12.6), abundant larval transcript-2 (ALT-2) and large extracellular domain of tetraspanin (TSP LEL)] as the most promising vaccine candidates. Vaccination trials with a multivalent fusion combination of the three proteins (rBmHAT) showed that approximately 95% protection can be achieved against challenge infections with B. malayi L3 in a mouse and jird model of lymphatic filariasis [9]. Although mice and jirds do not develop the typical lymphatic pathology during lymphatic filariasis infection, they are an excellent model to dissect out some of the key preclinical parasitological and immunological changes. Since rBmHAT vaccination gave close to sterile immunity in rodent models, there is great potential for developing this vaccine for human clinical trials. However, due to qualitative differences in the immune systems between rodents and human, vaccination trials in rodents are not easily translatable to clinical trial protocols. Also to understand the mechanisms of protective immune responses induced by vaccines, it is critical to use an animal model in which the immune system closely mimics that of human. Monkeys infected with B. malayi exhibited immunological changes in the lymphatic system similar to human lymphatic filariasis [10]. Therefore before testing the vaccine in human there is a need to evaluate its safety and immunogenicity in a non-human primate (NHP) model. Several other vaccines that are currently in use against infectious diseases were also developed through studies in NHPs [11], [12].
Rhesus macaques (Macaca mulatta) have close phylogenic relationship with human and are natural hosts for the lymphatic filarial infections [13]. In macaques filarial parasites develop into mature adult male and female worms and produce microfilaria similar to that in the human [14]. The infected macaques also develop fever, lymph node enlargement (around 5 weeks), lymphatic pathology (around 10–12 weeks) and chronic symptoms of elephantiasis very similar to that in the human lymphatic filariasis [15], [16]. Therefore, rhesus macaques are extremely valuable and an ideal animal model to evaluate the safety and efficacy of a vaccine against lymphatic filariasis [10], [17]. In fact, rhesus macaques have been used previously to test the vaccine potential of gamma-irradiation attenuated third stage B. malayi L3. In these previous studies, approximately 71% of vaccinated macaques were protected (5 out of 7 animals) against a challenge infection with B. malayi L3 [18] confirming that it is possible to evaluate vaccine-induced protection in macaques.
In the present study we evaluated the safety, immunogenicity and level of protection conferred following vaccination with rBmHAT in a rhesus macaque model. Protection was determined by evaluating the titer of protective antibodies, cytokine response of antigen responding peripheral blood mononuclear cells (PBMC), lack of establishment of challenge infection (as determined by the presence of circulating microfilariae, Mf), and absence of any clinical signs of lymphatic filariasis.
Materials and Methods
Ethics statement
Use of Rhesus Macaques (Macaca mulatta) in these studies was approved by the IACUC (Animal ethics) committee at the Bioqual Inc. (Rockville, MD). Ten 3- to 5-year old male rhesus macaques were purchased from PrimGen (Hines, IL). All macaques were quarantined for 6–8 weeks before entering them into the studies. Macaques were paired for housing, blood and stool samples were analyzed to ensure that the macaques are negative for any infections. Before entering each macaque into the study, serum samples were analyzed to confirm that the animals do not carry circulating antigens or antibodies against B. malayi adult worm antigens.
Animals were housed at Bioqual’s facility in Rockville, MD. Care and husbandry were provided in compliance with federal laws and guidelines as well as in accordance with recommendations provided in the NIH guide and other accepted standards of laboratory animal care and use. Bioqual is accredited by the Association for the Assessment and Accreditation of Laboratory Animal Care, (AAALAC file #624) and holds an Assurance on file with the National Institute of Health, Office for Protection of Research Risks as required by the US Public Health Service Policy on Humane Care and Use of Laboratory Animals. The PHS Animal Welfare Assurance File Number is #A-3086-01.
Animal are cared for in rooms that are maintained at a temperature range of 68°F–74°F and humidity between 30% and 70%. The air handling equipment provides 10 to 15 filtered fresh (non-recirculated) air changes per hour for all animal rooms. All animal rooms are negative to the corridors as required for housing nonhuman primates and animals exposed to infectious agents. The exhaust is passed through a HEPA filter system on the roof (the air is not recirculated). The exhaust system HEPA filters are certified twice each year by an outside vendor to ensure satisfactory operation of the system. To accommodate socialization of animals, Bioqual has designed a 6.0 sq. ft. over/under system to accommodate two animals.
The animals are fed Primate Biscuit (15% Primate Diet 8714), supplied by Harlan Teklad, Madison WI. Fresh fruit are supplied by the L&M Produce Company. Food and water are provided daily to all monkeys. Daily diets are supplemented with fresh fruit, including apples, bananas, pears, and oranges.
Bioqual employs four fulltime veterinarians who oversee animal enrichment in accordance with Bioqual’s Behavioral Environment Enhancement Program (BEEP). Bioqual has worked with OLAW and clients to socially house as many animals as possible. Any animal that requires additional enrichment due to abnormal behavior has a behavior and enrichment plan tailored to its needs, which is supervised by the behaviorist in conjunction with Bioqual’s veterinarians, assigned technicians and animal caretakers. All NHPs are observed twice per day and evaluated bi-monthly by a fulltime employed behaviorist. Additional behavioral observations are performed as needed when recommended/requested by the veterinary staff. All NHPs receive at least three commercially available pet toys in their cages at all times to manipulate as part of their enriched environment. NHPs are given destructible enrichment as a way for them to alter their environment, release aggression/tension, and forage through.
The destructible enrichment provided includes items such as cardboard, shredded paper, and phonebooks. Soft toys, such as fleece blankets and soft pet toys, are available for comfort to all NHPs, but are given more frequently to young, sick, and/or behaviorally distressed individuals. Various types of feeders/grooming boards, such as corn feeders, turf feeders, paint rollers, fruit feeders, and coconuts may be given.
To control of pain and discomfort, animals were sedated with ketamine, sometimes in combination with xylazine or telazol, for all technical procedures. Ketamine were given intramuscularly in the amount necessary for short-term procedures such as blood drawing. Animals were fully anesthetized for the challenge procedure.
All methods of euthanasia are performed in accordance with the 2000 Report of the AVMA Panel on Euthanasia and 2007 AVMA guidelines on euthanasia. The euthanasia procedure is performed by intravenous injection of a sodium pentobarbital overdose and subsequent auscultation of the heart”.
Macaques were paired for housing, blood and stool samples were analyzed to ensure that the macaques are negative for any infections. Before entering each macaque into the study, serum samples were analyzed to confirm that the animals do not carry circulating antigens (Hha I PCR, please see below for details) or antibodies against B. malayi or W. bancrofti adult worm soluble antigens or against SXP-1 as determined by ELISA.
Parasites
B. malayi infective third stage larvae (L3) were obtained from the NIAID/NIH Filariasis Research Reagent Resource Center (University of Georgia, Athens, GA).
Multivalent fusion protein rBmHAT
The multivalent fusion protein rBmHAT expressed in Escherichia coli BL21 (pLysS), was purified and endotoxin removed by Pierce High Capacity Endotoxin removal resin column (Thermo Fisher Scientific, Rockford, IL) as described previously [9].
Immunizations of rBmHAT
Five macaques each received 200 µg of rBmHAT vaccine mixed with 100 µg of AL007 alum (IDRI, Seattle, WA) under ABSL-2 conditions. Five (5) macaques that received alum (AL007) only remained as controls. Each animal was anesthetized with ketamine/xylazine and the vaccine was administered intramuscularly in each thigh (one injection site per thigh per vaccination). Animals were immunized at 4 weeks interval on days 0, 28 and 56. Intramuscular route is commonly used for clinical vaccine trials and hence we followed the same procedure for macaques. The injection sites were monitored daily for signs of fever, any adverse reactions (redness, swelling, etc.) for up to 7 days post immunization.
B. malayi L3 challenge
On day 84, one month after the final dose of vaccine, macaques were anesthetized with ketamine HCl and challenged subcutaneously with 400–500 Brugia malayi L3. To facilitate the production of the relatively large number of L3 (500 L3/animal) required for challenging 10 immunized macaques, the animals were divided into 2 subgroups within each group. The subgroups were challenged one week apart. Before challenge B. malayi L3 were counted and examined for viability under a microscope. Only viable parasites were used for challenge.
Monitoring of each animal after challenge
All animals were monitored daily for clinical signs after the challenge. Behavioral observations were similarly conducted during the entire post-challenge period. Clinical monitoring included serum chemistry, hematology, complete blood count (CBC) analysis (IDEXX) and CD4+/CD8+ T cell flow cytometry analysis. Body weights, body condition, lymphoedema and lymph node measurements were also recorded each time the animal was sedated for procedures (like immunizations, challenge, and blood collections).
Sample collections
Blood samples and peripheral blood mononuclear cells (PBMC) were collected at the Bioqual primate facility and shipped to UIC for sample analyses. Whole blood was collected into BD Vacutainer SST tubes according to manufacturer’s instructions. Heparinized blood (1 ml) was collected from the femoral vein of each animal during the immunization period and from the saphenous vein during the challenge period. The shift in blood collection site was to eliminate any potential interference with the inguinal lymph node measurements or assessments of edema. Blood samples were obtained at multiple time points during the entire follow-up period.
Isolation of PBMC
The blood pellets after plasma separation was diluted in phosphate buffered saline (PBS; 1∶2) and subjected to gradient density centrifugation for 30 min at 2200 rpm using a 90% Histopaque separation solution (Sigma, St. Louis, Mo.). The opaque interface containing mononuclear cells was collected, washed three times in PBS by centrifugation at 800 rpm. PBMC were enumerated using Trypan blue dye exclusion method and resuspended in RPMI 1640 medium containing 10% FBS (100 U/ml Penicillin/Streptomycin, and 2 mM L-glutamine). PBMC collected before the challenge was analyzed for T cell proliferation and IFN-γ secretions. PBMC collected after the challenge experiments were tested for T cell proliferation and ELISPOT assays. Proliferation assay was performed with PBMC isolated on the same day of blood collection. PBMC suspended in RPMI media with 10% FBS were used for Antibody Dependent Cellular Cytotoxicity (ADCC) assay and for cytokines analysis.
T cells proliferation and flow cytometry
Carboxyfluorescein diacetate succinimidyl ester (CFSE) based assay was used for assessment of antigen-specific proliferation within the T cell population [19]. A 5 mM CFSE stock solution (Invitrogen, Grand Island, NY) was prepared according to manufacturer’s instructions. PBMC collected four weeks after the final immunization were gently resuspended at 107 cells/ml in 5 µM CFSE and incubated in the dark at 37°C for 15 minutes. Cells were centrifuged and washed with RPMI containing 10% FBS (100 U/ml Penicillin/Streptomycin, and 2 mM L-glutamine) and incubated for an additional 30 minutes at 37°C. Cells were then washed, resuspended in RPMI containing FBS, plated in a 24-well plate at 2×106 cells/ml per well and incubated overnight at 37°C. The medium (∼500 µl) was removed the following day and cells were stimulated with 1 µg/mL of rBmHAT. Samples incubated only with RPMI medium served as negative controls. As a positive control for each animal, cells were stimulated with phytohemagglutinin (PHA). Cells were cultured and harvested after 5 days of stimulation. Following a washing step with PBS/0.2% FBS, cells were surface stained with an antibody cocktail of CD3-APC-Cy, CD4-PE and CD8-PerCP and incubated for 20 minutes at room temperature. After an additional washing step with PBS/0.2% FBS the cells were acquired on BD FACS Canto II flow cytometer (BD, San Jose, CA) and analyzed on a BD FACS Diva Software v6.1.2. At least 50,000 events within the live lymphocyte gate were acquired.
Cell Counts, serum chemistry and Complete Blood Count (CBC) analysis
CBC, serum chemistries and eosinophil counts were analyzed using commercial automated hematology and serum chemistry analyzers by IDEXX. Samples collected prior to the initiation of the study served as a normal reference baseline for each animal.
Secreted levels of IFN-γ were measured using an ELISA
PBMC (1×106 cells) were stimulated in vitro with 1 µg/ml of rBmHAT for 5 days at 37°C. Following stimulation the supernatants were harvested and assayed for secreted levels of IFN-γ using an ELISA kit (Mabtech AB, Ashburn, VA) according to manufacturer’s instructions.
ELISPOT assay
An ELISPOT assay was performed to determine the antigen-specific IFN-γ and IL-10 secreting cells in the PBMC of vaccinated and control macaques. We used a monkey ELISPOT kit purchased from U-Cytech biosciences (Yalelaan, The Netherlands) to determine the spot forming units as per the manufacturer’s instruction. PBMC collected 20 weeks post challenge were plated in 96 well plates at 1×106 cells/ml and were stimulated with 100 ng/well of B. malayi adult soluble antigen (BmA) for 24 hours at 37°C and 5% CO2. Wells of ELISPOT plates were coated with 100 µl/well of capture antibodies (anti-IL-10 or anti- IFN-γ) diluted in sterile coating buffer and incubated overnight at 4°C. Plates were washed 2 times with sterile coating buffer. After blocking the plates with 200 µl/well of blocking buffer for 1 hour at room temperature, PBMC that were already stimulated with BmA antigens or only media (negative control) were added to the wells of the ELISPOT plates at 100 µl/well and incubated for 24 hrs at 37°C and 5% CO2. All the cells were removed from the plates and the membrane was washed 3 times with sterile PBS. Following wash, 100 µl of detection antibodies were added to each well and incubated at room temperature for 2 hr. After washing the plate 4 times with wash buffer, avidin-HRP reagent was added (100 µl/well) and incubated for 45 minutes at room temperature. After a final wash with PBS, freshly prepared 3-amino-9-ethylcarbazole (AEC) substrate solution was added (100 µl/well) and monitored for the development of spots at room temperature for 10–60 minutes. The substrate reaction was stopped by washing wells 3 times with 200 µl/well ultrapure water. The plates were air dried. Spots were counted using a dissecting microscope. The plates were stored in the dark prior to reading. Antigen-specific responses were determined by subtracting the number of spots in the negative control wells from the wells containing antigens. Results are shown as the mean value of spots obtained from triplicate wells.
Analysis of serum antibody titers in macaques
Levels of IgG, IgG1, IgG2, IgG3, IgA and IgE antibodies against rBmHSP, rBmALT-2, rBmTSP or rBmHAT were determined in the sera (collected one month after the final dose of vaccine) of each rhesus macaque using an indirect ELISA as described previously [9]. Briefly, wells of a 96 well microtiter ELISA plates were coated with 100 ng/well of antigens (rBmHSP, rBmALT-2, rBmTSP or rBmHAT) in 0.05 M carbonate-bicarbonate buffer, pH 9.6. The wells were blocked with 3% BSA in 0.05% Phosphate Buffer Saline-Tween 20 (PBS-T), and 100 µl of sera samples (diluted in the range of 1∶100–1∶50,000 in PBS-T) from each macaque were added to each well. Goat anti-monkey IgG antibodies conjugated to peroxidase (Rockland Immunochemicals, Gilbertsville, PA) was used as secondary antibodies to determine IgG titer antibodies. The color was developed using OPD substrate and absorbance was read at 492 nm in the ELISA reader (BioRad, Hercules, CA). To determine the levels of isotype antibodies, biotinylated anti-monkey IgG1 (1∶2000), IgG2 (1∶200), IgG3 (1∶2000), IgA (1∶2000) and IgE (1∶1000) antibodies (NHP Reagent Resources, Boston, MA) were used as secondary antibodies. After washing the plates, optimally diluted streptavidin conjugated horse radish peroxidase (HRP) was added and further incubated for 60 minutes at room temperature and the color was developed.
Antibody dependent cell mediated cytotoxicity assay (ADCC)
ADCC was performed according to the protocol described previously [9]. PBMC were prepared from heparinized whole blood from a naive healthy animal as described previously. Briefly, ten B. malayi L3 (suspended in 50 µl RPMI 1640 medium containing 10% FBS) were incubated with 2×105 PBMC (in 50 µl RPMI 1640) and 50 µl of serum from each animal (collected one month after the final dose of vaccine) in a 96-well round bottom tissue culture plate. 5 replicates were performed for each serum sample. Control wells contained B. malayi L3 incubated in media, with sera alone or cells alone. The plates were incubated at 37°C with 5% CO2 for 48 hours. Following incubation, B. malayi L3 were examined under a microscope at 24 and 48 hours to determine larval viability. Dead L3 were defined as those having a limpid or straight appearance with no movements for an additional observation period of 8 hours at 37°C. Live larvae were active, coiled and motile. The percentage larval death was expressed as the ratio of the number of dead L3 to that of the total number recovered within the experimental period multiplied by 100. Average larval death in 5 wells were calculated and expressed as percent protection in each animal.
Knott test to determine microfilaremia in macaques
The presence of Mf in the blood of macaques was detected using the Knott technique as described previously [20]. Peripheral blood of macaques was screened weekly for Mf starting from 5 weeks to 20 weeks post challenge. Briefly, whole blood was mixed with 9 ml of a 2% formalin solution (prepared in PBS) in a 15 ml conical centrifuge tube. The tubes were gently rocked for 2 min at room temperature and centrifuged at 1,500 rpm for 5 min. The supernatant was then thoroughly decanted by turning the tube completely upside down to remove all the liquid. Following this 5 ml of ACK lysis buffer (Quality Biologicals, Gaithersburg, MD) was added to the pellet and the tube was vortexed. Two to three drops of methylene blue solution (Fisher Scientific, Hannover Park, IL) was then added to the tubes, gently mixed, and smeared onto five glass slides. The samples were allowed to dry and read under a microscope using 40X lens objective. A comparison of Mf counts in blood collected from the saphenous and femoral veins showed similar results.
Detection of Mf in the peripheral blood by polymerase chain reaction (PCR)
PCR-based assays are more sensitive in detecting the presence of Mf in the blood samples [21], [22]. Therefore, we used the PCR based assay also to confirm the presence of Mf in the blood samples of all macaques 20 weeks after challenge. Whole blood samples were centrifuged at 10,000 rpm for 5 min and the supernatant containing serum was stored at −20°C. DNA was isolated from the pellet using DNeasy Blood & Tissue Kit (Qiagen, Valencia, CA) according to the manufacturer’s instruction. Primers were synthesized at Integrated DNA Technologies Inc., (Coralville, IA) for HhaI tandem repeats. Primer sequence for HhaI tandem repeat was (Forward 5′-GCG CAT AAA TTC ATC AGC-3′, Reverse 5′-GCG CAA AAC TTA ATT ACA AAA GC-3′). PCR parameters were initial denaturation of 94°C for 5 min, followed by 40 cycles of 1 min at 94°C, 1 min at 56°C, 1 min at 72°C and a final extension of 10 min at 72°C. Following PCR reaction, 10 µl of each PCR product was analyzed on a 1% agarose gel.
PBMC proliferations assay
PBMC collected 10 weeks post challenge were cultured in 96 well tissue culture plates at a concentration of 1×106 cells/well in RPMI 1640 supplemented with 10% FCS. Cells were stimulated either with rBmHAT antigen (1 mg/ml) or Concanavalin A (1 mg/ml) or with medium alone (unstimulated) in triplicate wells. PBMC were stimulated in triplicate wells and the plates were incubated at 37°C in 5% CO2. After 72 h, cell proliferation was measured using cell counting kit (CCK-8) (Dojindo Molecular Technologies, Inc, Gaithersburg). Stimulation index of PBMC proliferation was calculated using the formula: Absorbance of stimulated cells/Absorbance of unstimulated cells.
Statistical analysis
Data are represented as the mean ± standard error. One-way ANOVA tests (Kruskal-Wallis) was performed for the antibody titer and T cell proliferation using GraphPad Prism software. Student T test was performed for protection studies. A probability (P) value of ≤0.001 was considered statistically significant.
Results
rBmHAT vaccination did not induce any adverse reactions in macaques
The injection sites were monitored closely for signs of any adverse reactions (redness, swelling, etc.) for 7 days post immunization. There were no adverse reactions in any of the vaccinated or control animals. Clinical monitoring showed no dramatic loss of body weight (>10% of the original weight), changes in eating habits or any other behavioral changes. Temperature measurements obtained daily following immunizations did not show any significant variations. Temperature measurements were also performed at regular intervals using implanted transponders. There were no significant variations in the body temperature in vaccinated and control animals.
The lymph nodes in the left and right leg of all animals were monitored weekly starting approximately 2 weeks prior to challenge (to establish a baseline) and throughout the challenge period. The lymph nodes were measured with a caliper and observed for edema. The measurements showed an overall increase in the mean size of the inguinal lymph nodes in both legs during the 5–8 week post challenge period in all groups. Compared to the baseline (14.5 mm) the lymph node size in control animals were 22±1 mm and rBmHAT group were 26.2±1 mm. Following this period, the sizes of the lymph nodes decreased to near pre-challenge levels in all macaques.
Challenge with B. malayi L3 did not alter the body temperature in macaques (data not shown). Analyses of the serum chemistry and hematology (CBC) values showed that they were all in the normal range for all cell types (data not shown) except for a slight increase in the eosinophil counts following L3 challenge in infected animals.
All three antigens in the multivalent vaccine construct were immunogenic in macaques
Analysis of the IgG antibody titer in vaccinated macaques showed that all the macaques developed high titers (1∶40,000) of IgG antibodies after third immunization against rBmHAT (Fig. 1). We then analyzed the titer of antibodies against each of the three component antigens in the vaccine construct. All macaques developed high titers of IgG antibodies against rBmHSP12.6 (1∶16,000), rBmALT-2 (1∶24,000) and rBmTSP-LEL (1∶16,000) (Fig. 1). There were slight individual variations in the titer of antibodies between each vaccinated macaques. On a comparative basis, macaque #5242, #5258 and #5259 showed the highest titer of IgG antibodies against the component antigens (except anti-rBmHSP12.6 antibodies in macaque #5258 and anti-rBmTSP antibodies in macaque #5259). Macaque #4996 and 5254 developed only low titers of antibodies to rBmALT-2 and rBmTSP (Table 1).
Figure 1. rBmHAT specific IgG antibodies in the sera of macaques.
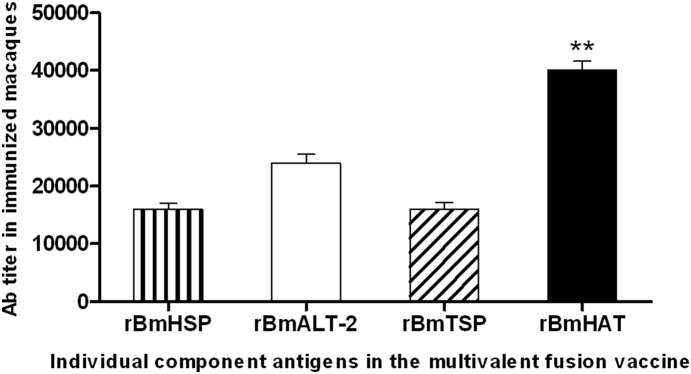
Macaques were immunized three times at four weeks interval with 200 µg of rBmHAT combined with alum (AL007) adjuvant. Sera collected one month after the final immunization was analyzed for IgG titer by ELISA. Approximately, 100 ng of each recombinant protein (100 ng/100 µl/well) were coated onto the wells of an ELISA plate and bound serum IgG was detected using a HRP labeled anti-monkey IgG secondary antibodies. Each bar represents mean ±S.D of 5 animals. Significant **P<0.001 titer of antibodies observed compared to other antigens.
Table 1. Titer of IgG antibodies.
| Animals immunized with rBmHAT | |||||
| Animal ID | 4996 | 5242 | 5254 | 5258 | 5259 |
| Antibody titre against r Bm HSP12.6 | 6400 | 16,000* | 6400 | 6400 | 16,000* |
| Antibody titre against r Bm ALT-2 | 3200 | 24000** | 800 | 24000** | 24000** |
| Antibody titre against r Bm TSP | 16,000* | 16,000* | 12,800* | 16,000* | 6400 |
| Antibody titre against r Bm HAT | 40,000 | 40,000 | 40,000 | 40,000 | 40,000 |
Macaques were immunized with 200 µg of rBmHAT with alum adjuvant. Anti- rBmHAT antibodies against rBmHSP12.6, rBmALT-2, rBmTSP LEL or rBmHAT were evaluated. Each animal differed in the antibody titer against each antigen.
*P<0.05 and **(P<0.001) statistically significant IgG antibody titer compared to other animals.
Isotype analysis showed that nearly all of the antibodies were of IgG1 isotype against all the four antigens tested (rBmHSP, rBmALT-2, rBmTSP and rBmHAT). Levels of IgG2, IgG3, IgA and IgE did not show any significant difference from the background values (Fig. 2).
Figure 2. rBmHAT specific antibodies isotypes in the sera of macaques.

Levels of IgG1, IgG2, IgG3, IgA and IgE antibodies against A) rBmHSP, B) rBmALT-2, C) rBmTSP and D) rBmHAT were determined in the sera (collected one month after the final dose of vaccine) of each rhesus macaque using an indirect ELISA. IgG1 antibodies were predominantly present in the immunized animals against all the four antigens tested. Each bar represents mean ±S.D of 5 animals. Significant **P<0.001 titer of antibodies observed compared to other animals.
rbmhat responding cells were present in the pbmc of immunized rhesus macaques
To determine the antigen specific proliferative responses, PBMC was collected four weeks after the final vaccination. Cell proliferation was determined after stimulating CFSE labeled PBMC with rBmHAT proteins for 5 days and counting the labeled cells in a flow cytometer. These results showed that the proliferation frequency of antigen-responding cells in the immunized animals were 3 folds higher (stimulation index 6.1±0.86) compared to the control animals (stimulation index 2.2±1.42). As expected, PBMC from all the animals showed robust proliferative responses (stimulation index 87.4±0) upon stimulation with pan-T mitogen, PHA. PBMC cultured in control medium had only low-level proliferation following 5-day incubation. The proliferation frequency value for each sample was obtained by subtracting the medium alone control value.
Frequency of CFSE labeled CD3+, CD4+ and CD8+ PBMC proliferating in response to antigen stimulation were determined by flow cytometry. These studies showed that there was an increase in the proliferation of antigen-responding T cells in all immunized macaques compared to control macaques (Fig. 3). Subset analysis showed that in immunized animals approximately 12.7% of the antigen responding T cells were CD4+ cells and 7.9% of T cells were CD8+ subsets (Fig. 3). Background proliferation in the presence of rBmHAT antigen in the PBMC of control animals were 1.4% for CD4+ cells and 2.3% for CD8+ cells.
Figure 3. T cells proliferative responses in rBmHAT vaccinated macaques.

PBMC from immunized macaques isolated at 4 weeks post final immunization were stimulated with rBmHAT or medium only (negative control). Flow cytometry analysis for the loss of CFSE labeling (indicating cell proliferation) was performed for cells within the live T cell (CD3+) or CD4+ or CD8+ T cell subsets. Shown are the mean ± S.I and results (% proliferating cells) of the five macaques in each group. The frequency for each sample is the value obtained following subtraction of the medium control. Significant **P<0.001 and *P<0.05 proliferation of cells observed compared to controls.
Antigen responding cells in the PBMC of immunized monkeys secrete IFN-γ
Antigen responding cells in the spleen of rBmHAT immunized mice and gerbils predominantly secreted high levels of IFN-γ [9]. Therefore, we wanted to determine if macaques also show a similar response after immunization but before challenge. These studies showed that PBMC from the 3 immunized macaques (#5242, #5258 and #5259) all secreted significant amounts of IFN-γ when stimulated with rBmHAT antigen (Table 2). Culture supernatants of PBMC from macaque #4996 and #5254 only had background levels of IFN-γ similar to that of the PBMC from control macaques (Table 2).
Table 2. Increase in IFN- γ production following rBmHAT antigen stimulation.
| Control (alum only) | rBmHAT(Immunized with rBmHAT + alum) | |||||||||
| Animal I.D | 4995 | 5240 | 5249 | 5252 | 5253 | 4996 | 5242 | 5254 | 5258 | 5259 |
| IFN-γ secretion (pg/ml) | 0 | 0 | 0 | 0 | 0 | 0 | 62.5 | 0 | 62.5 | 62.5 |
PBMC were isolated after the final immunization and subjected to rBmHAT antigen stimulation. Supernatants from PBMC cultures were harvested upon 5 days of stimulation and assayed in ELISA for IFN- production.
Anti-rBmHAT antibodies in the sera of immunized macaques can participate in the killing of B. malayi L3
To determine the protective ability of anti-rBmHAT antibodies in the sera of immunized macaques, we performed an in vitro ADCC assay. Results showed that the PBMC from vaccinated macaque were able to participate in the killing of 35% of B. malayi L3. When sera from individual macaques were evaluated maximum killing potential in the ADCC was 45% in the sera of macaque #5258 (Table 3). Sera from macaque #5242 and #5259 also showed significant killing potential with 38% and 35% killing respectively. Sera from macaque #4996 and #5259 had the least ADCC property with 25% and 31% killing respectively (Table 3). No larval death occurred when sera from control macaques were used in these assays (Table 3).
Table 3. Sera from macaques immunized with rBmHAT participated in the killing of B. malayi L3 in an ADCC mechanism.
| Animal I.D | Live L3 Mean ± S.Dof five wells | Dead L3 Mean ± S.Dof five wells | % Larval death Mean ± S.Dof five wells | Mean %Larval death |
| 4995 | 10 | 0 | 0 | 0 (Control) |
| 5240 | 10 | 0 | 0 | |
| 5249 | 10 | 0 | 0 | |
| 5252 | 10 | 0 | 0 | |
| 5253 | 10 | 0 | 0 | |
| 4996 | 7.5±0.5735 | 1.5±0.5735 | 25±5.16931* | 35% ±6.123724* (Immunized) |
| 5242 | 6.5±0.57735 | 4±0.57735 | 38±6.914* | |
| 5254 | 6.5±1 | 3±0.5735 | 31±7.3981* | |
| 5258 | 6.5±1.5275 | 5±0.57735 | 45±6.265021* | |
| 5259 | 7±1.154701 | 3.5±1.154701 | 35±11.54701* |
50 µl of sera samples from rBmHAT immunized macaques or control macaques were incubated with 2×105 PBMC from naive macaques and 10 B. malayi L3 for 48 hrs at 37°C. Larval viability was evaluated at the end of the incubation.
Significant larval death **(P<0.05) compare to other macaques. Control wells were L3 incubated with media, cells alone or sera alone.
Immunization with rBmHAT conferred partial protection in macaques
One month after the final vaccination, all 10 monkeys were challenged with 500 B. malayi L3 and screened for the appearance of Mf in the peripheral blood circulation. A Knott test and PCR analysis was used to detect Mf. The Knott test was performed weekly from week 5 post challenge until the animals became positive. In our studies, challenged macaques became positive for Mf starting from week 10 post challenge. During weeks 11–20 post challenge, 3 of the control macaques became positive for Mf. Unfortunately, the rest 2 control macaques remained negative through the end of the study. In the vaccinated group, 3 of the macaques (#5242, #5254 and #5259) remained negative throughout the study. However, 2 of the vaccinated macaques (#4996 and #5258) became positive for Mf (Table 4 and Fig. 4A). To further confirm the infection, we performed a PCR analysis, where Hha1 antigen specific primers were used to amplify for the presence of Mf specific DNA in the blood of infected monkeys. PCR analysis confirmed infections in macaque #5249 and #4996 (Table 4 and Fig. 4B). The other three positive animals identified by Knott technique were negative by PCR.
Table 4. Detection of Mf in the macaques challenged with B. malayi L3.
| Macaque groups | Animal ID | Presence of Mf | |
| Knott test | PCR test | ||
| Control (Immunized with only alum) | 4995 | + | − |
| 5240 | − | − | |
| 5249 | + | + | |
| 5252 | − | − | |
| 5253 | + | − | |
| rBmHAT (Immunized with rBmHAT mixed with alum) | 4996 | + | + |
| 5242 | − | − | |
| 5254 | + | − | |
| 5258 | − | − | |
| 5259 | − | − | |
The appearance of Mf in the peripheral blood of macaques during weeks 11–20 post challenge was detected by Knott and PCR techniques. 3 macaques in control group and 2 macaques in the immunized group were found to be microfilaremic.
Figure 4. All 10 monkeys challenged with B. malayi L3 were screened for the appearance of Mf in the peripheral blood circulation by Knott technique and PCR analysis.

A) Results from Knott technique showed the presence of mf in the blood that was stained with methylene blue. B) Mf specific Hha I gene was amplified and analyzed in 1% agarose gel.
rBmHAT responding cells were present in the PBMC of immunized rhesus macaques after challenge
PBMC collected 10 weeks post challenge was stimulated with rBmHAT to determine the antigen specific T cell response. PBMC of 3 animals #5242 (S.I- 0.928±0.01), #5258 (S.I- 1.091±0.16) and #5256 (S.I.-1.0181±0.13) from the vaccinated group that were negative for Mf showed significant proliferation upon rBmHAT stimulation. Whereas, 2 of the vaccinated animals #4996 (S.I- 0.258±0.12) and #5254 (S.I- 0.379±0.03) positive for Mf did not show significant proliferation upon rBmHAT stimulation (Table-5). No significant proliferation was observed in any of the control animals #4995 (S.I-0.280±0.03), 5240 (S.I. - 0.415±0.09), 5249 (S.I. - 0.300±0.26), 5252 (S.I. - 0.507±0.03) or 5253 (S.I - 0.475±0.25). S.I of PBMC stimulated with Concanavalin was in the range of 2.0–3.8.
Eosinophil numbers were high in infected macaques showing Mf
Microfilaremic individuals show high eosinophil counts in their blood [23], [24]. A similar finding was observed in rhesus macaques as well. Absolute counts of eosinophils were determined on weeks, 13, 9, and 5 prior to challenge, on the day of challenge and on weeks 1, 5, 10, and 14 post challenge. Our results show that there was an increase in the frequency of eosinophil numbers in the peripheral blood of microfilaremic macaques around 10 weeks post challenges (Fig. 5). One macaque (#5259) that was negative for Mf also showed some eosinophilia. Eosinophil counts were 10 fold higher in control macaques that had microfilariae in their peripheral blood.
Figure 5. Percent of eosinophils in the peripheral blood of control and vaccinated macaques.

Percent of eosinophils were evaluated in the peripheral blood of macaque pre- and post- challenge. Following challenge there was an increase in the frequency of eosinophils in animals that were microfilaremic as determined by Knott’s technique. Each line represents the eosinophil count of individual macaque on weeks −13, −9, −5, −1 before challenge, on day of challenge (week 0) and at weeks 1, 5, 10, and 14 post challenges. The levels of eosinophils in Mf positive animals are represented by a solid line. Levels of eosinophils in Mf negative animals are represented as a dotted line. # Macaques immunized with rBmHAT. Significant at **P<0.001 high number of eosinophils.
High titer of antigen-specific IgG antibodies and elevated antigen specific secretion of IFN-γ from PBMC correlated with protection in the immunized macaques
Since two of the macaques in the immunized group showed presence of infection following challenge, we compared the vaccine induced immune responses in the 2 infected macaques with similar responses in the 3 uninfected macaques within the immunized group. We compared the values before and after challenge. Values before challenge will eliminate any bias due to the challenge of parasites. A chart showing comparative immunological values are presented in Table 5. Results show that the titer of IgG antibodies was significantly high in the 3 immunized macaques that did not develop the infection after the challenge. Similarly PBMC from the same 3 macaques secreted higher levels of IFN-γ when stimulated with the rBmHAT antigen. PBMC from the 2 immunized macaques that developed the infection after challenge were unable to secrete similar levels of IFN-γ in response to rBmHAT stimulation. An ELISPOT assay was performed using PBMC from vaccinated and control macaques. Results show that in all the infected macaques there was a significant increase in the number of antigen-specific IL-10 secreting cells compared to IFN-γ secreting cells (Fig. 6). When we compared the ratio of IFN-γ to IL-10 secreting cells in the PBMC of immunized macaques, there was a significant increase in the IL-10 secreting cells in the two vaccinated macaques that showed infection (Table 6). These findings suggest a clear correlation between the type immune responses elicited and the failure to establish infection in the vaccinated macaques.
Table 5. PBMC were isolated 10 weeks post challenge from each macaque and cultured at 2×105 cells per well in triplicate wells of a 96 wells plate.
| Macaque groups | Animal ID | PBMC Proliferation Mean S.I ± S.D (N = 3) | |
| Stimulated withConcanavalin A | Stimulated with rBmHAT | ||
| Control (Immunized with only alum) | 4995 | 3.260±0.01 | 0.280±0.03 |
| 5240 | 3.090±0.58 | 0.415±0.09 | |
| 5249 | 2.982±0.24 | 0.300±0.26 | |
| 5252 | 3.674±0.83 | 0.507±0.03 | |
| 5253 | 2.582±0.72 | 0.475±0.25 | |
| rBmHAT (Immunized with rBmHAT mixed with alum) | 4996 | 3.874±0.47 | 0.258±0.12 |
| 5242 | 2.170±0.43 | 0.928±0.01** | |
| 5254 | 2.068±0.18 | 0.379±0.03 | |
| 5258 | 3.304±0.64 | 1.091±0.16** | |
| 5259 | 2.883±0.27 | 1.0181±0.13** | |
PBMC were stimulated in vitro with 1 µg/ml of rBmHAT for 72 hrs at 37°C and analyzed for proliferation using cell counting kit (CCK-8). Con A was used as positive control. Stimulation index (S.I.) was calculated from the unstimulated control wells.
Significant proliferation of PBMC **(P<0.001) compare to PBMC from other macaques.
Figure 6. Antigen specific IFN-γ and IL-10 secreting cells in macaques.

To determine the antigen-specific IFN-γ and IL-10 secreting cells ELISPOT was performed in the PBMC collected 20 weeks post challenge from vaccinated and control macaques. 1×106 cells/ml were stimulated with 100 ng/well of B. malayi adult soluble antigen (BmA) for 48 hrs at 5% CO2 and used in the ELISPOT assays. Spot forming units were counted using a dissecting microscope. Antigen-specific cells were counted by subtracting the number of spots in the negative control wells from the wells containing antigens. Results are shown as the mean spot forming units obtained from triplicate wells. Each bar represents mean ± S.D of 5 animals. Significant **P<0.001 number of spots compared to other groups.
Table 6. Correlates of protection in macaques vaccinated with rBmHAT.
| Macaque groups | Animal ID | Immunological values before L3 challenge | Immunological values after L3 challenge | ||||
| Antibody titer of>12,000 againstrBmHSP | Antibody titerof >12,000 againstrBmALT-2 | Antibody titer of>12,000 againstrBmTSP LEL | IFN-γproduction | Mfdetection | Ratio of IFNγ:IL-10 secreting cells | ||
| Control (Immunizedwith only alum) | 4995 | − | − | − | − | + | 1∶3 |
| 5240 | − | − | − | − | − | 1∶1 | |
| 5249 | − | − | − | − | + | 1∶11 | |
| 5252 | − | − | − | − | − | 1∶0.01 | |
| 5253 | − | − | − | − | + | 1∶13 | |
| rBmHAT (Immunized withrBmHAT mixed with alum) | 4996 | − | − | + | − | + | 1∶4 |
| 5242 | + | + | + | + | − | 1∶0.003 | |
| 5254 | − | − | + | − | + | 1∶2 | |
| 5258 | + | + | + | + | − | 1∶0.001 | |
| 5259 | + | + | − | + | − | 1∶0.02 | |
The three vaccinated macaques that did not show any evidence of infection had high titer of anti- rBmHAT IgG antibodies and their PBMC secreted significantly high levels of IFN-γ but low levels of IL-10 as determined by an ELISPOT assay. Ratio of IFN-γ: IL-10 secreting cells was calculated for each macaque. These analyses also confirmed that there is a clear difference between the infected macaques and the three vaccinated macaques that did not show any infection. These findings coupled with the ADCC data clearly suggest that the three vaccinated macaques that did not show any signs of infection were protected.
#PBMC collected before challenge and stimulated with rBmHAT.
Discussion
Filarial parasites have complex life cycle that involve several discrete developmental stages within the human host [25]. L3 transmitted by mosquitoes develop into adult parasites that live in the lymphatic system for several years often more than 30 years evading the host immune system. Microfilariae that are released from these adult parasites that circulate in the blood can be cleared by anti-filarial drugs such as DEC, Albendazole and Ivermectin [26]–[28]. Current mass drug treatment administration (MDA) strategy uses these drugs to clear the parasite from the circulation to reduce the incidence of the infection and thus leading the way for elimination of lymphatic filariasis from the endemic regions. However, for sustained control strategy and for eradication of lymphatic filariasis, there is a need for an effective vaccine that can be combined with the current successful MDA approach. Previous studies from our laboratory showed that a multivalent fusion protein vaccine rBmHAT, could confer over 95% protection against challenge infections in rodents. In this study we show that rBmHAT vaccine confers approximately 35% protection in vaccinated macaques, as confirmed by the absence of circulating microfilaria, presence of protective antibodies and cells, absence of circulating antigen and lack of any visible pathology following a challenge infection. Our results showed that the protection in vaccinated macaques correlated with high titer of IgG antibodies against the vaccine antigen and higher frequency of IFN-γ secreting antigen responding cells in the peripheral blood and significant L3 killing potential in the sera of immune macaques. These findings suggested that the rBmHAT vaccine is partially protective in macaques.
None of the vaccinated animals showed any adverse reactions to rBmHAT vaccination. There were no abnormal clinical signs such as increase in the body temperature, pain, irritability or any altered behavioral changes in vaccinated macaques. Similarly the social behavior, feeding and sleeping habits were not altered in vaccinated macaques suggesting that the vaccination was safe in these macaques.
All the vaccinated macaques developed significantly high titers of IgG antibodies against rBmHAT. This suggests that the vaccine antigens are highly immunogenic in macaques. These findings were similar to that observed in mice and gerbils following vaccination with rBmHAT [9], [29]. Subsequent analysis of the isotype of IgG antibodies showed that anti-BmHAT antibodies in the sera of vaccinated macaques were predominantly of IgG1 isotype. Surprisingly the vaccinated macaques did not show any IgG2 or IgG3 antibodies against the vaccine antigen. Previous studies in EN human subjects showed that these individuals carry significantly high titers of IgG1, IgG2 and IgG3 antibodies against the three vaccine antigens in their sera and these antibodies have protective function [3], [9]. Depletion of total IgG, IgG1, IgG2 or IgG3 antibodies from the EN sera significantly reduced the L3 killing ability of EN sera. Reconstitution of the antibody depleted sera with the antibodies recovered the L3 killing effect suggesting that the IgG1, IgG2 and IgG3 antibodies have an important role in the protection against lymphatic filariasis in the human [9]. Lack of elicitation of anti-rBmHAT IgG2 and IgG3 antibodies in the sera of vaccinated macaques might explain the low protection rate observed following the challenge infections. Thus, it is possible that generating a balanced IgG1/IgG2 response in the macaque might significantly improve the protection rate after vaccination. Typically macaques do not produce IgG3 antibodies [30]. This might explain why we did not see any detectable anti-rBmHAT IgG3 antibodies in our vaccinated macaques. Similarly, there were no detectable levels of anti-rBmHAT IgE antibodies in the sera of vaccinated macaques. This is really advantageous if this vaccine were to be developed further for human use.
Alum adjuvants in general promote IgG1 antibodies and are poor inducers of IgG2 antibody responses [31]. Thus, the high levels of anti-rBmHAT IgG1 antibodies observed in our vaccinated macaques may be largely alum driven. Therefore, we hypothesize that addition of another adjuvant such as Toll-like receptor (TLR)-4 agonists along with alum can promote both IgG1 and IgG2 responses. In fact, one of our recent studies in mice showed that a combination of alum plus TLR-4 agonist can induce a balanced Th1/Th2 response in mice and this correlated with significant protection [32]. Based on these studies, it is possible that an inclusion of TLR-4 agonist along with alum as adjuvants for rBmHAT vaccination in macaques could substantially increase the protection rate.
Previous studies from our laboratory and others showed that ADCC assay could be used as a surrogate to measure protective responses to lymphatic filariasis in vitro [3], [9]. Our results showed that sera samples from all vaccinated macaque were able to kill B. malayi L3. However, the mean larval killing potential of the sera was only 35%, which was significantly lower than what we observed in the putatively immune EN human subjects and in the rodent models. Since both IgG1 and IgG2 antibodies appears to be critical for the killing of L3, we believe that the reduced ADCC function observed in the sera of our vaccinated macaque may be due to the lack of IgG2 antibodies.
Acute lymphatic filariasis infection is confirmed in the infected human subjects by demonstrating the presence of microfilariae in the peripheral blood using a Knott technique [33]. A similar analysis in our infected control macaques showed that microfilariae could be demonstrated in the peripheral blood of infected macaques. In addition we also used a PCR based method to confirm the infection [21]. PCR method detects the circulating antigens in the peripheral blood of infected animals. Both these analyses are mutually exclusive but provided a confirmative diagnosis. However, the PCR approach in our hands appeared to be less sensitive than actual demonstration of the parasite. Out of the three animals that were positive for microfilariae by Knott technique in the control group only one animal was found to be positive by the PCR method. Similarly out of the 2 positive samples identified by Knott’s method in the vaccinated animals, only one animal was positive in the PCR assay. It is not clear why we observed this discrepancy. One possible explanation may be that we identified one Mf in 50 ml of blood by the Knott’s technique and the PCR method failed to detect low levels of infections. These findings suggest that multiple diagnostic approaches may be needed to confirm the presence of low level infections. Nevertheless, samples positive by PCR method was also positive by Knott’s technique. Demonstrating Mf in the peripheral blood inconclusively proves infection.
Two of the control macaques in our study did not pick the infection. At this time we do not know why the infection failed to establish in these two macaques. Prior to entering the animals into the study, serological and parasitological analyses were performed in all the animals and all of them were negative for any parasitic infection and did not carry any antibodies against B. malayi.
A distinctive characteristic of lymphatic filariasis infection in the human is the appearance of eosinophilia [24]. All the infected macaques in the control and vaccinated groups showed significant eosinophilia, whereas, eosinophilia was not a prominent finding in uninfected animals. One vaccinated animal with no infection also showed eosinophilia. Clinical data did not show any signs of other infections or allergy in this particular animal. Reason for elevated eosinophils in this animal could not be determined.
PBMC hypo-responsiveness to parasite antigen is another characteristic in human subjects who have circulating microfilaria [34], [35]. Analysis of the proliferative responses of PBMC from the 2 infected macaques in the vaccinated group with BmA and rBmHAT antigens showed minimal proliferation similar to the control animals further confirming that these animals were infected, whereas, the PBMC from all the three uninfected animals in the vaccinated group proliferated significantly suggesting that these animals are immune.
PBMC from lymphatic filariasis infected patients do not secrete interferon (IFN)-γ in response to parasite antigen [35]–[37] but secrete significant amounts of the anti-inflammatory cytokines primarily IL-10 because of circulating microfilariae [38]. Analysis of the cytokine response of PBMC from the two infected macaques in the vaccinated group showed that these cells secrete high levels of IL-10 and low levels of IFN-γ compared to PBMC from the three uninfected macaques which secreted significantly high levels of IFN-γ. A similar dichotomy in the IFN-γ secretion was seen in the mouse model as well [9]. These findings clearly showed that the three Mf negative macaques in the vaccinated group have developed significant immunity against the challenge infection and prevented establishment of the infection.
Thus, based on high titer of antibody responses elicited after vaccination in macaques, ability of the serum antibodies to participate in the ADCC function to kill B. malayi L3, absence of Mf in the peripheral circulation, significant proliferative response of PBMC and their ability to secrete significant amounts of IFN-γ it could be concluded that the three Mf negative macaques in the vaccinated group are truly protected. Unfortunately, two of the macaques in the control group failed to show any infection. Analysis of the various parameters in these two uninfected control macaques definitely demonstrated that the challenge infection failed to establish in these two macaques. Therefore, we conclude that vaccination with rBmHAT can confer partial protection in macaques against B. malayi L3 challenge infection. Further studies are needed to improve the vaccine formulation or adjuvant formulation to increase the vaccine induced protection against B. malayi infections in macaques before moving this vaccine to human clinical trials.
Acknowledgments
We would like to thank Andy Morehouse at the NIH/NIAID Filariasis research reagent resource center, College of Veterinary Medicine, University of Georgia, Athens, GA for supply of B. malayi third stage infective larvae (L3) for these studies.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.
Funding Statement
Funding for these studies was provided through PIPB by the Division of Microbial and Infectious Diseases contract number HHSN27220100006I/HHSN2720003 to Bioqual, and additional funding was from NIH RO1 grant AI064745 to RK. Bioqual Inc. provided support in the form of salaries for authors AVG, HA, and ML, but did not have any additional role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Molyneux DH, Zagaria N (2002) Lymphatic filariasis elimination: progress in global programme development. Ann Trop Med Parasitol 96 Suppl 2: S15–40. [DOI] [PubMed] [Google Scholar]
- 2. Day KP (1991) The endemic normal in lymphatic filariasis: A static concept. Parasitol Today 7(12): 341–343. [DOI] [PubMed] [Google Scholar]
- 3. Dakshinamoorthy G, Samykutty AK, Munirathinam G, Shinde GB, Nutman T, et al. (2012) Biochemical characterization and evaluation of a Brugia malayi small heat shock protein as a vaccine against lymphatic filariasis. PLoS One 7(4): e34077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Veerapathran A, Dakshinamoorthy G, Gnanasekar M, Reddy MV, Kalyanasundaram R (2009) Evaluation of Wuchereria bancrofti GST as a vaccine candidate for lymphatic filariasis. PLoS Negl Trop Dis 3(6): e457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Gnanasekar M, Rao KV, He YX, Mishra PK, Nutman TB, et al. (2004) Novel phage display-based subtractive screening to identify vaccine candidates of Brugia malayi. Infect Immun 72(8): 4707–4715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Anand SB, Gnanasekar M, Thangadurai M, Prabhu PR, Kaliraj P, et al. (2007) Immune response studies with Wuchereria bancrofti vespid allergen homologue (WbVAH) in human lymphatic filariasis. Parasitol Res 101(4): 981–988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Thirugnanam S, Pandiaraja P, Ramaswamy K, Murugan V, Gnanasekar M, et al. (2007) Brugia malayi: comparison of protective immune responses induced by Bm-alt-2 DNA, recombinant Bm-ALT-2 protein and prime-boost vaccine regimens in a jird model. Exp Parasitol 116(4): 483–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Dakshinamoorthy G, Munirathinam G, Stoicescu K, Reddy MV, Kalyanasundaram R (2013) Large extracellular loop of tetraspanin as a potential vaccine candidate for filariasis. PLoS One 8(10): e77394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Dakshinamoorthy G, Samykutty AK, Munirathinam G, Reddy MV, Kalyanasundaram R (2013) Multivalent fusion protein vaccine for lymphatic filariasis. Vaccine 31(12): 1616–1622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Giambartolomei GH, Lasater BL, Villinger F, Dennis VA (1998) Diminished production of T helper 1 cytokines and lack of induction of IL-2R+ T cells correlate with T-cell unresponsiveness in rhesus monkeys chronically infected with Brugia malayi . Exp Parasitol 90(1): 77–85. [DOI] [PubMed] [Google Scholar]
- 11. Gardner MB, Luciw PA (2008) Macaque models of human infectious disease. Ilar J 49(2): 220–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Ahmad G, Zhang W, Torben W, Ahrorov A, Damian RT, et al. (2011) Preclinical prophylactic efficacy testing of Sm-p80-based vaccine in a nonhuman primate model of Schistosoma mansoni infection and immunoglobulin G and E responses to Sm-p80 in human serum samples from an area where schistosomiasis is endemic. J Infect Dis 204(9): 1437–1449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Shedlock DJ, Silvestri G, Weiner DB (2009) Monkeying around with HIV vaccines: using rhesus macaques to define ‘gatekeepers’ for clinical trials. Nat Rev Immunol 9(10): 717–728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. el-Bihari S, Ewert A (1971) Distribution of developing and mature Brugia malayi in Rhesus monkeys and in the jird, Meriones unguiculatus, after a single infection. J Parasitol 57(6): 1170–1174. [PubMed] [Google Scholar]
- 15. Crandall RB, Crandall CA, Neilson JT, Fletcher JT, Kozek WW, et al. (1983) Antibody responses to experimental Brugia malayi infections in patas and rhesus monkeys. Acta Trop 40(1): 53–64. [PubMed] [Google Scholar]
- 16. Wong MM, Guest MF, Lim KC, Sivanandam S (1977) Experimental Brugia malayi infections in the rhesus monkey. Southeast Asian J Trop Med Public Health 8(2): 265–273. [PubMed] [Google Scholar]
- 17. Folse DS, Ewert A (1988) Edema resulting from experimental filariasis. Lymphology 21(4): 244–247. [PubMed] [Google Scholar]
- 18. Wong MM, Fredericks HJ, Ramachandran CP (1969) Studies on immunization against Brugia malayi infection in the rhesus monkey. Bull World Health Organ 40(4): 493–501. [PMC free article] [PubMed] [Google Scholar]
- 19.Parish CR, Glidden MH, Quah BJ, Warren HS (2009) Use of the intracellular fluorescent dye CFSE to monitor lymphocyte migration and proliferation. Curr Protoc Immunol Chapter 4: Unit4 9. [DOI] [PubMed]
- 20. Liu JY1, Wang GQ, Chen YN, Tu ZP, Fang Z (1989) Observations on the clinical manifestations and treatment of an experimental infection with Brugia malayi in man. J Trop Med Hyg. 92(2): 93–96. [PubMed] [Google Scholar]
- 21. Mishra K, Raj DK, Dash AP, Hazra RK (2005) Combined detection of Brugia malayi and Wuchereria bancrofti using single PCR. Acta Trop 93(3): 233–237. [DOI] [PubMed] [Google Scholar]
- 22. Rao RU, Weil GJ, Fischer K, Supali T, Fischer P (2006) Detection of Brugia parasite DNA in human blood by real-time PCR. J Clin Microbiol 44(11): 3887–3893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Pearlman E, Kroeze WK, Hazlett FE Jr, Chen SS, Mawhorter SD, et al. (1993) Brugia malayi: acquired resistance to microfilariae in BALB/c mice correlates with local Th2 responses. Exp Parasitol 76(2): 200–208. [DOI] [PubMed] [Google Scholar]
- 24. Pearlman E, Kazura JW, Hazlett FE Jr, Boom WH (1993) Modulation of murine cytokine responses to mycobacterial antigens by helminth-induced T helper 2 cell responses. J Immunol 151(9): 4857–4864. [PubMed] [Google Scholar]
- 25. Choi YJ, Ghedin E, Berriman M, McQuillan J, Holroyd N, et al. (2011) A deep sequencing approach to comparatively analyze the transcriptome of lifecycle stages of the filarial worm, Brugia malayi. PLoS Negl Trop Dis 5(12): e1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Farid HA, Hammad RE, Hassan MM, Ramzy RM, El Setouhy M, et al. (2005) Effects of combined diethylcarbamazine and albendazole treatment of bancroftian filariasis on parasite uptake and development in Culex pipiens L. Am J Trop Med Hyg. 73(1): 108–114. [PubMed] [Google Scholar]
- 27. Critchley J, Addiss D, Ejere H, Gamble C, Garner P, et al. (2005) Albendazole for the control and elimination of lymphatic filariasis: systematic review. Trop Med Int Health 10(9): 818–825. [DOI] [PubMed] [Google Scholar]
- 28. Hotez PJ (2009) Mass drug administration and integrated control for the world’s high-prevalence neglected tropical diseases. Clin Pharmacol Ther 85(6): 659–664. [DOI] [PubMed] [Google Scholar]
- 29. Joseph SK, Ramaswamy K (2013) Single multivalent vaccination boosted by trickle larval infection confers protection against experimental lymphatic filariasis. Vaccine 31(33): 3320–3326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Shearer MH, Dark RD, Chodosh J, Kennedy RC (1999) Comparison and characterization of immunoglobulin G subclasses among primate species. Clin Diagn Lab Immunol 6(6): 953–958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Jin BR, Kim SJ, Lee JM, Kang SH, Han HJ, et al. (2013) Alum Directly Modulates Murine B Lymphocytes to Produce IgG1 Isotype. Immune Netw 13(1): 10–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Dakshinamoorthy G, Kalyanasundaram R (2013) Evaluating the efficacy of rBmHATalphac as a multivalent vaccine against lymphatic filariasis in experimental animals and optimizing the adjuvant formulation. Vaccine 32(1): 19–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Turner P, Copeman B, Gerisi D, Speare R (1993) A comparison of the Og4C3 antigen capture ELISA, the Knott test, an IgG4 assay and clinical signs, in the diagnosis of Bancroftian filariasis. Trop Med Parasitol 44(1): 45–48. [PubMed] [Google Scholar]
- 34. de Almeida AB, Carmelita M, Silva Me, Maciel MAl, Freedman DO (1996) The Presence or Absence of Active Infection, Not Clinical Status, Is Most Closely Associated with Cytokine Responses in Lymphatic Filariasis. Journal of Infectious Diseases 173(6): 1453–1459. [DOI] [PubMed] [Google Scholar]
- 35. Maizels RM, Sartono E, Kurniawan A, Partono F, Selkirk ME, et al. (1995) T-cell activation and the balance of antibody isotypes in human lymphatic filariasis. Parasitol Today 11(2): 50–56. [DOI] [PubMed] [Google Scholar]
- 36. Mahanty S, Nutman TB (1995) Immunoregulation in human lymphatic filariasis: the role of interleukin 10. Parasite Immunol 17(8): 385–392. [DOI] [PubMed] [Google Scholar]
- 37. Maizels RM, Holland MJ, Falcone FH, Zang XX, Yazdanbakhsh M (1999) Vaccination against helminth parasites–the ultimate challenge for vaccinologists? Immunol Rev 171: 125–147. [DOI] [PubMed] [Google Scholar]
- 38. Mahanty S, Luke HE, Kumaraswami V, Narayanan PR, Vijayshekaran V, et al. (1996) Stage-specific induction of cytokines regulates the immune response in lymphatic filariasis. Exp Parasitol 84(2): 282–290. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.