

Abstract
Background
The JAK/STAT (Janus Tyrosine Kinase, Signal Transducers and Activators of Transcription) pathway is associated with cytokine or growth factor receptors and it is critical for growth control, developmental regulation and homeostasis. The use of porcine ocular cells as putative xenotransplants appears theoretically possible. The aim of this study was to investigate the response of various porcine ocular cells in vitro to human cytokines in regard to the activation of JAK-STAT signaling pathways.
Methods
Porcine lens epithelial cells, pigmented iris epithelial cells and pigmented ciliary body cells were used in this study. These cells were isolated from freshly enucleated porcine eyes by enzymatic digestion. Cultured cells between passages 3–8 were used in all experiments. Electromobility shift assay (EMSA), proliferation assay, immunofluorescence staining and flow cytometry were used to evaluate the JAK-STAT signaling pathway in these cells.
Results
JAK/STAT signaling pathways could be activated in porcine pigmented epithelial ciliary body cells, in pigmented iris epithelial cells and in lens epithelial cells in response to porcine and human interferons and cytokines. All cells showed very strong STAT1 activation upon stimulation with porcine interferon-gamma. Porcine ocular cells also respond to human cytokines; IFN-alpha induced strong activation of STAT1 in EMSA, flow cytometry and immunofluorescence experiments whereas activation of STAT3 was less strong in EMSA, but strong in flow cytometry and immunofluorescence. Human recombinant IL-6 activated STAT3 and human IL-4 activated STAT6. With the help of immunofluorescence assay and flow cytometry we observed nuclear localization of STAT proteins after activation of porcine ocular cells with cytokines and interferons. Human IFN-α had an inhibitory effect on porcine ocular cells in proliferation assays.
Conclusion
Our study demonstrated that some types of human cytokines and interferon activate intracellular JAK-STAT signaling pathways in porcine ocular cells. We hypothesize that direct stimulation of the JAK-STAT pathway in porcine cells in response to human cytokines will lead to complications or failure, if pig-to-human ocular tissue xenotransplantation were to be carried out. For successful xenotransplantation among other obstacles there must be new approaches developed to regulate signaling pathways.
Keywords: cytokine signaling, interferon, pig retina, STAT proteins, xenotransplantation
Introduction
The pig has been used extensively in experimental studies of human eye diseases and disorders 1–;3, as well as a possible source of xenotransplantation 4–;9. For example, glaucoma and retinitis pigmentosa models 1,10–;12 and retinal conditions such as proliferative vitreoretinopathy 13 and retinal detachment 14 have been modeled on pigs. Porcine ocular cells share many morphological features with the human eye 15–;17. Porcine eyes have also similar size with human eyes. Because there is limited availability of human organs and cells in terms of clinical transplantation, pigs may be an alternative source.
Age-related macular degeneration (AMD) is a degenerative disease of the macula that can cause irreversible loss of central vision. AMD is characterized by macular changes consisting of retinal pigment epithelium (RPE) abnormalities and drusen 18. Current investigations for developing treatments for retinal atrophy have shown the importance of RPE. Degeneration or dysfunction of the RPE can lead to photoreceptor degeneration and as a consequence to blindness. RPE cells are of great interest to researchers working in the field of tissue engineering and cell transplantation 19,20.
Studies in animal models demonstrated that the transplantation of RPE cells can delay the course of degenerative diseases. Although the first attempts to transplant RPE cells into the subretinal space in humans suffering from AMD were less successful, RPE cell transplantation is still favoured as a future therapeutic option, and much work has been carried out to develop cell transplants 20. In a recently published paper, Melville et al. 21 discuss the possibilities of stem cell-derived RPE transplantation into subretina space of patients with dry AMD.
The highly pigmented outer layer of the ciliary body continues into the RPE. Iris pigment epithelium and RPE possess the same embryologic origin. Rezai et al. 22 have shown that iris pigment epithelial cells were able to take over the functions of RPE cells, serving as an autologous graft for transplantation in a rat model. These results suggest that pigmented iris or pigmented ciliary body cells transplanted into the subretinal space might substitute the RPE. We speculate that also porcine iris or porcine pigmented ciliary body cells might substitute the RPE in humans.
In the last decade, the major progress has been made in the area of pig-to-human xenotransplantation. However, it is practically overlooked whether major players of inflammation, human cytokines, and interferons are able to activate signaling pathways in porcine ocular cells. Although the eye is known as an immunologically privileged organ, induction of cytokines in the ocular cells could be an early trigger of the inflammatory response, as it is known to occur after infection. Levels of interferon-gamma (IFN-γ), tumor necrosis factor-alpha (TNF-α), IL-1, IL-2, IL-6 were shown to be elevated in vitreous aspirates of patients with uveitis, proliferative vitreoretinopathy, and other inflammatory eye diseases 23–;25. However, there are only few studies researching the effect of human cytokines on pig tissues and vice versa 26. Thus, it has been shown that human IFN-γ and interleukin (IL)-1 beta do not have an effect on the expression of RANTES in porcine endothelial cells 26. Zhang et al. 27 reported that porcine IL-2 showed almost no effects on human T cells and practically did not promote porcine T-cell proliferation, although other research groups have shown the effects of human IL-2 on porcine cells 28,29. Recently, Manna et al. 30 have demonstrated that human dendritic cells (DC) activated by human IL-15 were cytotoxic toward allogeneic and xenogeneic endothelial cells via apoptotic reaction. Cellular assays remain the critical tool to elucidate the mechanism of xenogeneic rejection.
Signal transducers and activators of transcription (STAT) proteins are a class of transcription factors that become activated upon tyrosine phosphorylation. They are a family of cytoplasmic proteins with roles as signal messengers and transcription factors that participate in normal cellular responses to cytokines and growth factors 31–;34. To date, there are seven STAT family members identified in mammals. Frequently, the activity of certain STAT family members is associated with human malignancies, inflammatory responses, and acute rejection reactions 35,36. STAT1 has been described as upregulated during acute allograft rejection of different organs 37–;40, and there are also reports of roles of STAT4 and STAT6 during the allograft rejection 41 and during T helper cell differentiation 42. It has been shown that neutralizing the transcription factor STAT1 with a decoy oligodeoxynucleotide has a powerful, long-lasting anti-inflammatory effect 43,44 and may be considered for treating acute, and possibly, chronic graft rejections 45,46. Targeting these STAT transcription factors is considered as a useful approach to influence acute rejection reactions and prolongate the life of organ transplants.
This study was undertaken to delineate the molecular basis of human interferon and cytokine actions toward porcine ocular tissue.
Specifically, we examined whether STAT proteins could be activated in porcine ocular cells in response to human interferon and human cytokines, and whether either of them are involved in JAK–STAT signaling in these cells.
Materials and methods
Chemicals and reagents
Human recombinant IL-6 (hIL-6), human recombinant IL-4 (hIL-4), and porcine recombinant interferon-gamma (pIFN-γ) were from R&D Systems (Minneapolis, MN, USA). Roferon-A, human recombinant interferon-alpha (hIFN-α), was obtained from Hoffman-La Roche Ltd. (Reinach, Switzerland). Polyclonal antibodies against human STAT1, STAT2, STAT3, and STAT6 were from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Primary rabbit polyclonal antibodies against phosphorylated human STAT proteins for immunofluorescence experiments were from Cell Signaling (Danvers, MA, USA). Secondary antibodies conjugated with Alexa Fluor 488 (goat anti-mouse) and Alexa Fluor 555 (goat anti-rabbit) were purchased from Invitrogen (Carlsbad, CA, USA). Alexa Fluor 488 mouse anti-human phospho-STAT1 (epitope Y701), anti-phospho-STAT3 (epitope Y705), anti-phospho-STAT6 (epitope Y641), as well as isotype-matched IgG for flow cytometry experiments were purchased from BD Biosciences (San Jose, CA, USA). All cell culture reagents were obtained from Invitrogen. All cell culture plasticware was purchased from TPP (Trasadingen, Switzerland).
Cells
Porcine eyes from domestic pigs (4–6 months old, both males and females) were enucleated directly post mortem at the local abattoir. Ex vivo dissection of porcine lens epithelial cells (pLEC) were performed by isolation of anterior lens capsules with the adherent epithelium from freshly enucleated porcine eyes. Only clear lenses were used. Pigmented porcine iris epithelial cells (pIEC) as well as pigmented cells of ciliary bodies (pCBC) were isolated by enzymatic digestion in a trypsin/EDTA solution (0.05/0.02% w/v) for 15 min at 37 °C as previously described 47 and seeded in 75-cm2 culture flasks in Dulbecco’s minimal essential medium (DMEM), supplemented with 10% heat-inactivated fetal calf serum (FCS), 2 mm L-glutamine, and standard amounts of antibiotics. Cells were maintained as subconfluent cultures by passaging twice weekly. All experiments have been performed in passages 3–8. The ARPE-19, Colo 205, Daudi, and HepG2 cell lines were purchased from ATCC (Manassas, VA, USA) and cultivated according to the ATCC recommendations in 5% CO2/95% air at 37 °C.
Proliferation assay
Cell proliferation was evaluated using cultures of 10 000 cells per well in 24-well plates in the absence or presence of 500 U/ml of hIFN-α for the indicated time period (on days 2, 4, and 5) as it was previously described 48. Cells were trypsinized, and cell numbers were determined by live cell counting using a hematocytometer and 0.4% trypan blue reagent. Mean values and standard deviations were calculated. The experiments were carried out three times in triplicates for each cell type.
Electromobility shift assay
To examine the effects of cytokines on the activation of STAT proteins, electromobility shift assay (EMSA) was performed as it was previously described 49,50. Briefly, 1 day before the treatment, subconfluent cells were starved overnight in a serum-free medium. Cytokine treated and untreated cells (negative control) were lysed in a low-salt buffer (20 mm HEPES, pH 7.9, 10 mm KCl, 1 mm EDTA; 1 mm ethylene glycol tetraacetic acid (EGTA), 0.2% NP-40, 10% glycerol, 0.1 mm Na3VO4, 1 mm phenylmethanesulfonyl fluoride (PMSF), 1 mm dithiothreitol (DTT), 2 μg/ml aprotinin, 1 μg/ml leupeptin, 1 μg/ml pepstatin) at 4 °C for 10 min. After centrifugation for 1 min at 12 000× g, the supernatant (cytoplasmic extract) was removed, and the pelleted nuclei were incubated in a high-salt buffer (same as low-salt buffer), but with 420 mm NaCl and 20% glycerol for 30 min at 4 °C. Samples were cleared by centrifugation at 12 000× g at 4 °C. The supernatant was designated as the nuclear extract. The protein concentration was determined using the Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer’s instructions. Bovine serum albumin was used as a standard. All nuclear extracts were stored at –80 °C. For the EMSA, nuclear extracts were incubated for 20 min at room temperature in 20 mm HEPES, pH 7.9, 4% Ficoll, 1 mm MgCl2, 40 mm KCl, 0.1 mm EGTA, 0.5 mm DTT, 160 mg/ml poly(dI-dC)xpoly(dI-dC) with 1 ng of 32P-labeled oligonucleotides. Samples were separated on a 5% non-denaturing polyacrylamide gel at 400 V for 4 h at 4 °C. The oligonucleotide probes corresponding to STAT response element sequences are summarized in Table1. O15 probe binds to STAT1–STAT2–p48 complex; βCas contains the sequence of the β-casein promoter and binds to STAT1, STAT5, and STAT6 homodimers; Cε binds to STAT 6 homodimers and M67 probe binds to STAT1, STAT1:3, STAT3, and STAT4 homo-/heterodimers. The experiments were carried out five times.
Table 1.
Oligonucleotide sequences used for electromobility shift assay experiments
Immunofluorescence staining
Porcine ocular cells were cultured on cover glasses in 24-well tissue culture dishes. Cells were left untreated (negative control) or incubated with appropriate cytokines for 20 min at 37 °C. Then, cells were fixed in a freshly prepared (50 : 50) methanol/acetone mixture for 20 min on ice and were further incubated with 10% goat serum in PBS for 1 h at room temperature. Anti-phospho-STAT antibodies were used as primary antibodies. Goat anti-mouse Alexa Fluor 488 and goat anti-rabbit Alexa Fluor 555 were used as secondary antibodies, and the preparations were examined on confocal laser scanning microscope (Zeiss LSM 510, Oberkochen, Germany). Microscopic images were collected and analyzed using proprietary image acquisition LSM software, version 3.2. The experiments were carried out four times.
Analysis of activated STAT proteins by flow cytometry
Porcine cells were incubated for 20 min with appropriate cytokines or left untreated as a negative control. Cells were fixed with 2% paraformaldehyde for 20 min and then permeabilized in 90% methanol for 30 min. All cells were stained for 1 h with Alexa Fluor 488-conjugated anti-phospho-STAT1, anti-phospho-STAT3, or anti-phospho-STAT6 antibodies as it is indicated in figure legends. Mean fluorescence intensity of stained cells was quantitatively analyzed using a FACSCalibur flow cytometer and the CellQuest Pro Software (Becton Dickinson, Franklin Lakes, NJ, USA). The experiments were carried out four times.
Results
Interferon-alpha-induced growth inhibition
The effect of hIFN-α on the proliferation of pCBC, pLEC, and pIEC cells was analyzed by quantitation of cell viability using a trypan blue staining. Three independently performed experiments gave the results presented in Fig.1. The percentage of dead cells in cultures with and without IFN-α did not exceed 7%. As shown on Fig.1, exogenously added hIFN-α at a concentration of 500 U/ml suppressed the proliferation of pCBC, pLEC, and pIEC. In cultures without IFN-α the cell number increased whereas in the presence of IFN-α cell proliferation was inhibited between 19 and 37% on day 2, 30–50% on day 4, and 35–65% on day 5. pLEC were more sensitive to hIFN-α treatment compared with pCBC and pIEC.
Figure 1.
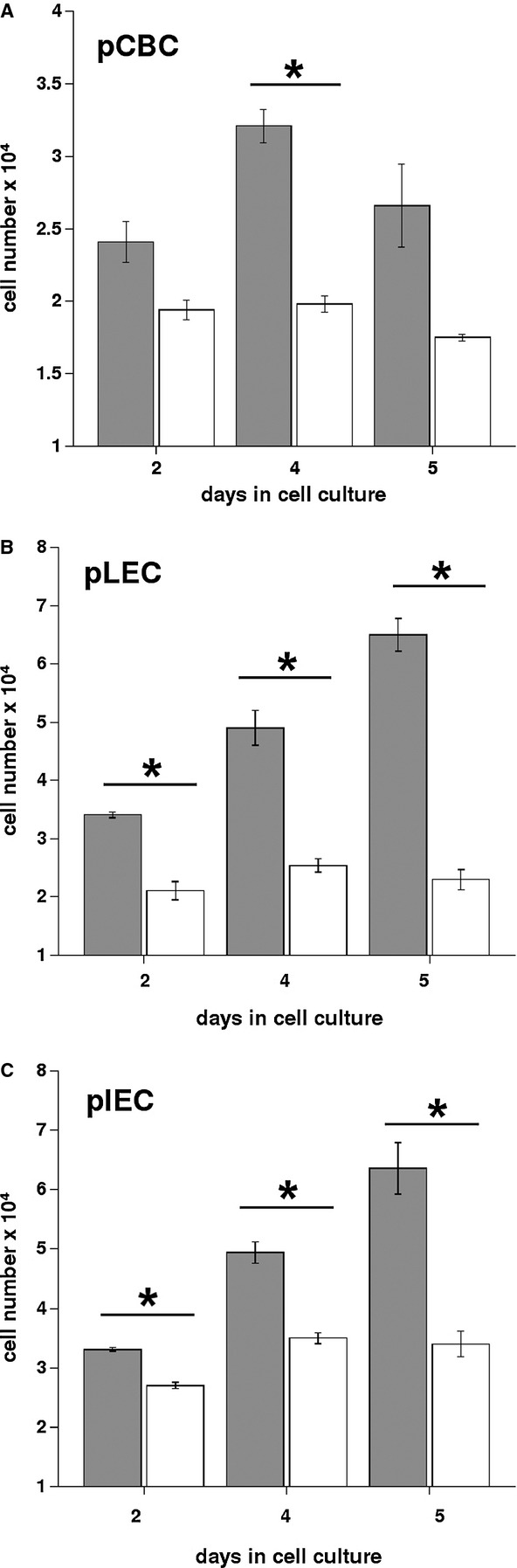
Interferon-alpha induced inhibition of proliferation in porcine ocular cells. Proliferation of porcine cells pCBC (A), pLEC (B), pIEC (C) over a 5-day period in the absence (gray bars) or presence (white bars) of 500 U/ml of human recombinant IFN-α. Three independent sets of experiments have been performed and are presented. Values are cell number means ± SD of n = 3 experiments. *statistically significant difference by Student’s t-test (P < 0.02) between untreated and treated cells.
JAK–STAT activation in porcine ocular cells
Electromobility shift assay experiments
To determine whether interferons and cytokines induce any known STAT protein in porcine eye, the EMSA was carried out with nuclear extracts of pCBC, pLEC, and pIEC and probed with four oligonucleotides (O15, βCas, Cε, and M67) that recognize all known STAT proteins. Untreated cells were used as negative controls, and for positive controls, ARPE-19, Daudi, HepG2, and Colo 668 cell lines were treated with appropriate cytokines. The cells were stimulated for 20 min with pIFN-γ or hIFN-α, or with hIL-6, or hIL-4. Five independently performed experiments gave the results presented in Figs5. Figure2A shows that stimulation with hIFN-α activates a STAT1–STAT2–p48 complex that binds to the O15 probe. All nuclear extracts of stimulated cells have been verified by supershift experiments using specific anti-STAT antibodies (Figs2B, 3B,C and 4B). The supershift experiment (Fig.2B) demonstrated that the O15 band disappears after incubation with antibodies specific for STAT1 and STAT2, whereas antibodies specific for STAT3 had no effect. IFN-α stimulated human retinal pigment epithelial cell line ARPE-19 served as a positive control (Fig.2B).
Figure 5.
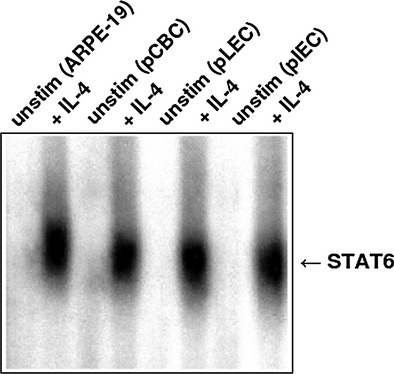
Electromobility shift assay analysis of nuclear extracts from pCBC, pLEC, and pIEC using the 32P-labeled Cε probe. Human retina pigment epithelial cells ARPE-19 stimulated with 10 ng/ml of IL-4 for 20 min were used as a positive control for the Cε probe. Porcine cells were left untreated (negative control) or treated with IL-4. The position of activated STAT6 is indicated by arrow.
Figure 2.
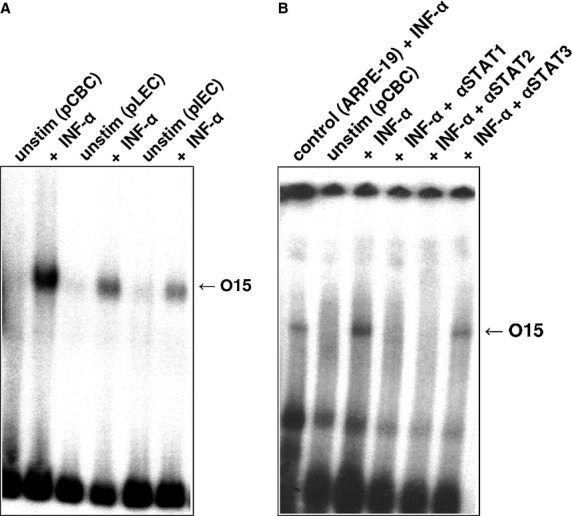
Electromobility shift assay analysis of nuclear extracts from pCBC, pLEC, and pIEC cells incubated with 32P-labeled O15 probe. (A) Nuclear extracts were prepared from unstimulated cells (negative control) and from cells stimulated with 500 U/ml of human recombinant IFN-α for 20 min. The position of the activated STAT1–STAT2–p48 complex is indicated by arrow. (B) Supershift experiment: IFN-α-treated pCBC cells were incubated with antisera specific for STAT1 (IFN-α + αSTAT1), with antisera specific for STAT2 (IFN-α + αSTAT2), or with antisera specific for STAT3 (IFN-α + αSTAT3). The nuclear extracts of human retinal pigment epithelial cell line ARPE-19, treated with 500 U/ml of IFN-α for 20 min were used as a positive control for the O15 probe.
Figure 3.
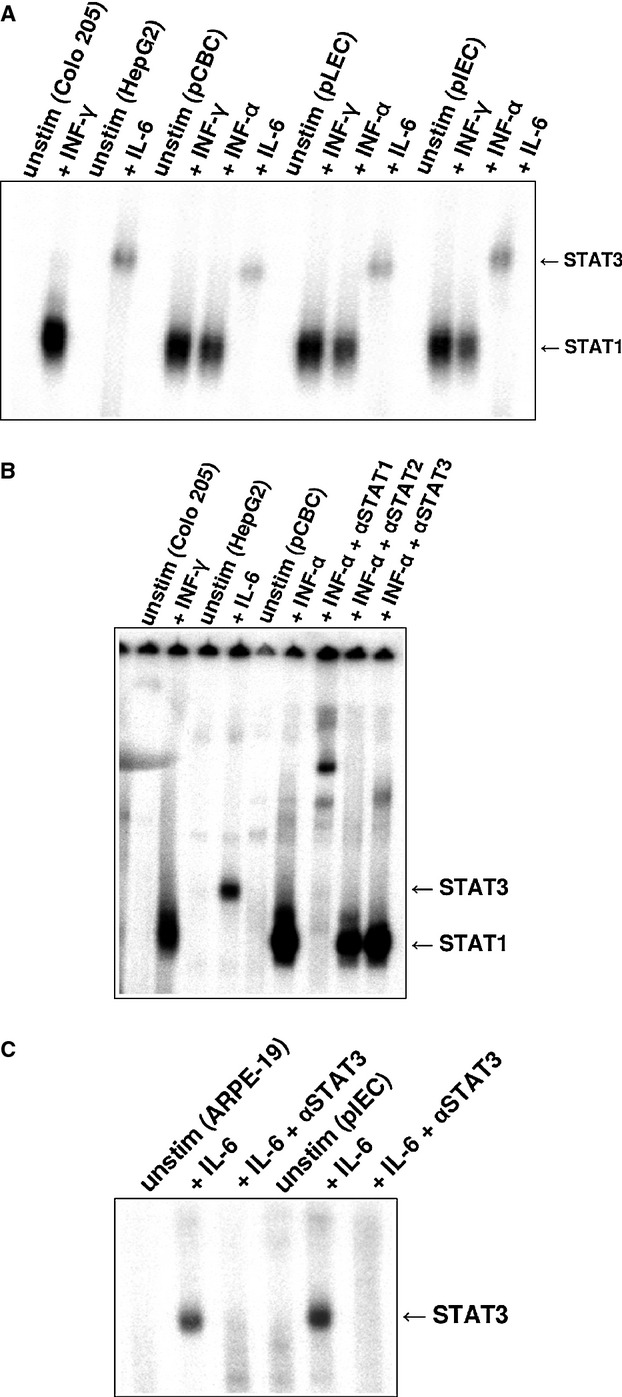
Electromobility shift assay (EMSA) analysis of nuclear extracts from pCBC, pLEC, and pIEC incubated with 32P-labeled M67 probe. (A) Colo 205 cells were left untreated or stimulated with 500 U/ml of human IFN-γ for 20 min STAT1 positive control). HepG2 cells stimulated with 10 ng/ml of human IL-6 were used as STAT3 positive control. pCBC, pLEC, and pIEC cell extracts were either untreated or treated with 10 ng/ml of porcine IFN-γ, with 500 U/ml of human IFN-α or with 10 ng/ml of human IL-6 for 20 min. The position of activated STAT1 and STAT3 proteins is indicated by arrows. (B) EMSA supershift experiment for STAT1 and STAT3 upon stimulation with human IFN-α using M67 probe. pCBC cells were left untreated or stimulated with human IFN-α. IFN-α treated pCBC cells were incubated with antisera specific for STAT1 (IFN-α + αSTAT1), with antisera specific for STAT2 (IFN-α + αSTAT2) or with antisera specific for STAT3 (IFN-α + αSTAT3). The nuclear extracts of human Colo 205 cell line treated with 10 ng/ml of human IFN-γ for 20 min were used as a positive control for the STAT1 and HepG2 cells treated with 10 ng/ml of human IL-6 as a positive control for STAT3. (C) EMSA supershift experiment for STAT3 upon stimulation with IL-6 on M67 probe. pLEC cells were left untreated or stimulated with human IL-6. IL-6-stimulated pLEC cells were incubated with antisera specific for STAT3 (Il-6 + αSTAT3). ARPE-19 cells were used as a positive STAT3 control. The cells were treated the same way as porcine cells: the cells left untreated or were stimulated with human IL-6 for 20 min. ll-6-treated ARPE-19 were incubated with antisera specific for STAT3 (Il-6 + αSTAT3).
Figure 4.
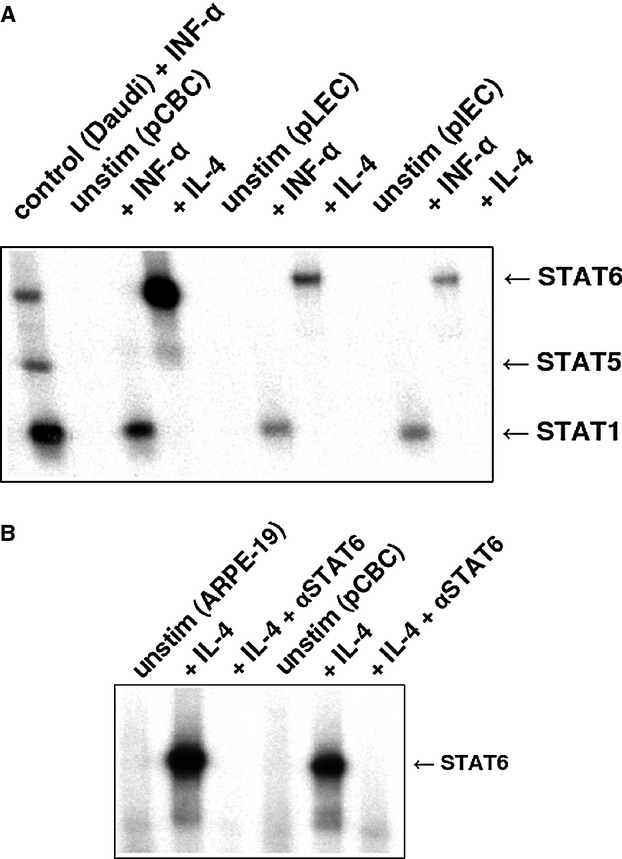
Electromobility shift assay (EMSA) analysis of nuclear extracts from pCBC, pLEC, and pIEC incubated with the 32P-labeled βCas probe. (A) Nuclear extracts were prepared from unstimulated porcine cells (negative control) and from cells stimulated with 500 U/ml of human IFN-α or with 10 ng/ml of human recombinant IL-4 for 20 min. Nuclear extracts from Daudi cells treated with 500 U/ml of human IFN-α were used as a positive control and compared with porcine cells. The position of the activated STAT1, STAT5, and STAT6 proteins is indicated by arrows. (B) EMSA supershift experiment for STAT6 upon stimulation with IL-4 on βCas probe. pCBC cells were left untreated or stimulated with human IL-4. IL-4-stimulated pCBC cells were incubated with antisera specific for STAT6 (IL-4 + αSTAT6). ARPE-19 cells were used as a positive STAT6 control. The cells were treated the same way as porcine cells: left untreated or were stimulated with human IL-4 for 20 min. lL-4-treated ARPE-19 were incubated with antisera specific for STAT6 (Il-4 + αSTAT6).
When tested on M67 probe, a very strong activation of STAT1 proteins was observed after pIFN-γ stimulation (Fig.3A). Untreated cells revealed no STAT signals. hIFN-γ-stimulated Colo 205 cell line served as a positive control for STAT1 and the HepG2 cell line stimulated with hIL-6 served as a positive control for STAT3 (Fig.3A,B). hIFN-α induced STAT1, STAT1:STAT3, and STAT3 activation. After IFN-α stimulation, STAT3 proteins were visible on all overexposed films as a very faint, but reproducible band. The STAT1:STAT3 heterodimers were visible as a band of intermediate intensity. This STAT1:STAT3 band was migrating between STAT1 and STAT3 bands (Fig.3B). Supershift experiment revealed that both STAT1 and STAT3 homodimers as well as STAT1:STAT3 heterodimers were supershifted after incubation with anti-STAT1 and anti-STAT3 antibodies, whereas STAT2-specific antisera had no effect (Fig.3B). hIL-6 activated STAT3 in all tested cells (Fig.3A). These STAT3 bands could be supershifted with antibodies specific for STAT3. hIL-6 stimulated ARPE-19 cell line served as a positive control for STAT3 supershift experiment (Fig.3C).
All nuclear extracts obtained from cells, stimulated with hIL-4 and hIFN-α, were tested on βCas probe. Strong STAT6 and STAT1 bands were observed (Fig.4A). The STAT6 band could be supershifted with antibodies specific for STAT6 (Fig.4B).
The STAT6 proteins were also visible when the nuclear extracts were tested with the Cε probe derived from the immunoglobulin heavy chain ε gene promoter. When the porcine cells were stimulated with hIL-4, a strong band corresponding to STAT6 was observed (Fig.5).
Taken together, our gel shift data showed that the intracellular STAT pathways are activated in porcine cells in response to human cytokines, with hIFN-α activating STAT1 and STAT3. Similarly, hIl-4 activated STAT6, and hIL-6 activated STAT3.
Flow cytometry analysis
To further confirm the above observations, the expression of phosphorylated STAT proteins was measured by flow cytometry after incubating porcine cells with hIFN-α, hIL-4, or hIL-6. Four independently performed experiments gave the results presented in Fig.6. The samples were incubated with anti-phospho-STAT1, STAT3, or STAT6 antibodies. Strong anti-phospho-STAT1 and anti-phospho-STAT3 shifts were observed in response to hIFN-α (Fig.6, panels A and B, respectively). Representative result of hIL-6- induced activation of STAT3 is shown in Fig.6 (panel C). The treatment with hIL-4 induced a strong STAT6 shift as shown in Fig.6 (panel D).
Figure 6.

Flow cytometric analysis of phospho-STAT translocation in pCBC cells. Unstimulated cells (gray histograms) are compared with cells stimulated for 20 min with different cytokines (white histograms). (A) Stimulation with human IFN-α and analysis of phospho-STAT1. (B) Stimulation with IFN-α and analysis of phospho-STAT3. (C) Stimulation with human Il-6 and analysis of phospho-STAT3. (D) Stimulation with human IL-4 and analysis of phospho-STAT6.
Immunofluorescence analysis
In the next set of experiments, we analyzed the activation of STAT proteins by immunofluorescence. Four independently performed experiments gave the results presented in Fig.7. The pLEC, pIEC, and pCBC were cultured on cover glasses. After stimulation with the appropriate cytokine (or interferon), the cells were fixed in methanol/acetone mixture and probed with antibodies that react only with phosphorylated STAT proteins. These experiments confirmed data gathered from the EMSA and flow cytometry. Representative illustration out of four experiments of nuclear expression of STAT1 is shown in Fig.7. The pLEC were left untreated (left panel) or were treated with hIFN-α (Fig.7, right panel). Incubation with hIFN-α caused the nuclear translocation of STAT1 and STAT3, whereas the stimulation of cells with IL-6 caused the translocation of STAT3 and IL-4 caused the translocation of STAT6 into nuclei (data are not shown).
Figure 7.
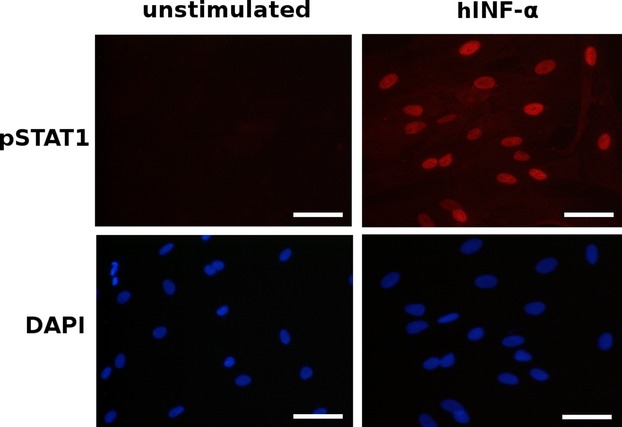
Confocal immunofluorescence analysis of phospho-STAT1 in pLEC. Unstimulated pLEC cells (left panel) or stimulated with 10 ng/ml of hIFN-α for 20 min (right panel). Both untreated and hIFN-α treated cells were incubated with anti-phospho-STAT1 antibodies. When pLEC cells were stimulated with hIFN-α there was intense nuclear immunostaining of phospho-STAT1 (right upper panel). DAPI staining was used for detection of nuclei. Magnification ×20. Scale bar 50 μm. This is a representative illustration out of four experiments.
Discussion
The compatibility of cross-species activation and costimulating molecules is a critical problem in the xenogeneic context 51. Severe shortage of allogeneic organ donors is the main reason for performing xenotransplantation of organs, and pigs are considered as the most suitable species. However, the compatibility between porcine organs and humans needs to be studied on tissue, cellular and molecular levels. The reports on the function of human cytokines, for example, IL-2 on porcine cells, are controversial 27–;29, although a number of cross-species interactions of porcine and human integrins and selectins 52,53, activation of porcine endothelial cells in response to human CD154 54, interaction of porcine CD86 and human CD28 55,56 and human CD2 with porcine CD28 are confirmed by different investigators 55–;57.
Type I interferons are central to innate and adaptive immunity, and type I porcine interferons are thoroughly characterized 58,59. In the context of cellular rejection of xenotransplant, IFN-γ is produced by the recipient (primate) effector cells 60. However, little is known about the STAT family in porcine ocular cells, although STAT4 and STAT6 recently have been cloned and described 61. We did not found in the literature publications the study of the JAK–STAT pathway in porcine cells in response to interferons and interleukins. Also, as is mentioned above, just a few studies were related to signal transduction in porcine cells in response to human cytokines and vice versa are available. Thus, it is known that stimulation of porcine vitreous-derived cells by human IL-1α, IL-1β, and TNF can increase expression of VEGF and IL-6 62 and lead to procoagulant changes in porcine endothelial cells 63.
To characterize the effects of human cytokines on porcine ocular cells, the JAK–STAT signaling pathway was studied in detail. For stimulation of porcine ocular cells pIFN-γ, several human recombinant cytokines and hIFN-α were used in current study.
The JAK–STAT pathway was originally discovered through the study of interferon-induced intracellular signal transduction 31,32,64. Subsequently, the STAT proteins have been shown to be induced by a wide variety of growth factors and cytokines. The STAT proteins have diverse biologic functions, which include roles in cell differentiation, proliferation, development, apoptosis, and inflammation 31,32,64,65. The STAT proteins are phosphorylated on tyrosine residues in response to growth factors and cytokines. On phosphorylation, STATs dimerize via SH2–phosphotyrosine interactions and become competent to bind to DNA. Phosphorylation causes the STATs to translocate from the cytoplasm to the nucleus 31,64,65. Because of their diverse biologic functions, aberrations in STAT signaling are predicted to have a wide variety of consequences 35,65.
Interferon-alpha activates the JAK–STAT pathway and exhibits immunomodulatory, antiviral and anti-proliferative activity 31–;35. IFN- α is a large family of proteins, which subdivided into 13 different subtypes. There IFNs share high sequence similarity. Currently, IFN-α is used in the treatment for certain forms of cancer 34,35,49. In current study, we have examined STAT1, STAT3, and STAT6 activation. Our data show that the JAK–STAT pathway could be activated in all porcine eye cells.
Proliferation assay revealed that hIFN-α substantially suppresses the proliferation of all tested porcine cells (Fig.1). In all wells without hIFN-α treatment cell number increased, in contrast hIFN-α-treated pCBC, pLEC, and pIEC cells have shown reduced proliferation (Fig.1). The anti-proliferative effect of hIFN-α was described for many human cells 34,48. In this study, we demonstrate that porcine ocular cells are sensitive to hIFN-α treatment.
Using all four labeled oligoprobes (M67, O15, βCas. and Cε), which allow for the detection of all known activated STAT proteins in EMSA, we were able to demonstrate that porcine eye cells are able to activate different STAT protein molecules in response to human cytokines. EMSA data have shown that porcine cells activate STAT1 in response to porcine IFN-γ, which is species specific. In addition, activation of STAT proteins was induced by human cytokines (IL-4, IL-6) and hIFN-α. Thus, STAT proteins activated by interferons, and human cytokines have also been verified using STAT protein-specific antibodies in the supershift experiments. As result of cellular activation, phosphorylated STATs were translocated to the nuclei. This was confirmed by confocal fluorescence assay and flow cytometry using phospho-specific STAT antibodies.
In this study, we showed that the direct biologic activities of human IFN-α and cytokines toward porcine eye cells are mediated through the JAK–STAT pathway. We hypothesize that activation of JAK–STAT pathway by human cytokines may modulate interaction of porcine cells with cells of human immune system. Thus, activation of STAT1 and STAT3 may lead to activation of many genes involved in immunosuppression and induction of tolerance 66–;68, and rebuilding the balance of STAT1 and STAT3 signaling may be explored as a useful approach to entertain survival of xenogeneic transplants. These results provide the first demonstration that porcine ocular cells can receive and transduce human interferon and cytokine signals by the JAK–STAT signaling pathway. Identification of the activated STAT molecules in other ocular cells and the target genes they regulate would provide important insights into the role of this family of transcription factors in the eye and in ocular diseases. The JAK–STAT pathway is not the only pathway, which could be activated in porcine ocular tissue in response to human cytokines; therefore, other signaling pathways must be investigated in detail.
Xenotransplantation approach provides the hope for a supply of organs and cells for patients awaiting transplantation. Also, in the field of ophthalmology, cell replacement is a promising approach. It has a potential in retinal degenerative diseases (retinitis pigmentosa and AMD), for which no pharmacological and surgical treatments are currently available 69. In this study, we have demonstrated that several STAT proteins could be activated in porcine ocular cells in response to human cytokines, and we speculate that xenotransplantation would fail due to activation of undesired intracellular signaling pathways. Prior performing xenotransplantation, researches must develop strategies to overcome these problems and consequently prevent xenograft rejection.
Acknowledgments
The authors would like to thank B. Erne (Confocal Core Unit, University of Basel, Switzerland) for technical assistance, A. Gorelova (Harvard University, USA) for editorial help and critical reading of the manuscript. The authors apologize to those investigators whose works were not cited because of space limitations.
Glossary
- AMD
age-related macular degeneration
- β-Cas
β-caseine
- DNA
deoxyribonucleic acid
- DAPI
diamidino-2-phenylindole
- DTT
dithiothreitol
- EDTA
ethylenediaminetetraacetic acid
- EGTA
ethylene glycol tetraacetic acid
- EMSA
electromobility shift assay
- FCS
fetal calf serum
- GTKO
alpha-1, 3-galactosyltransferase
- hCRP
human complement-regulatory proteins
- IFN
interferon
- IL
interleukin
- IRF
interferon regulatory factor
- JAK
janus kinase
- NK
natural killer cells
- pIFN-γ
porcine recombinant interferon-gamma
- pIEC
porcine iris epithelial cells
- pLEC
porcine lens epithelial cells
- pCBC
porcine pigmented cells of ciliary bodies
- PMSF
phenylmethylsulfanyl fluoride
- RANTES
regulated on activation, normal T cell expressed and secreted
- RPE
retinal pigment epithelium
- STAT
signal transducers and activators of transcription
- TNF
tumor necrosis factor
- VEGF
vascular endothelial growth factor.
Author contributions
EFK, NB provided the concept and design; AR, AN performed data analysis/interpretation; EFK, NB, JH –drafted the article; PM, JF provided critical revision of article, PM provided approval of article, JH performed statistics, and EFK, AR, SK, KW, AN collected the data. EFK was supported by University Hospital Basel and NB by Immune Disease Institute and Harvard Pilot Grant. PM received funding from the University Eye Clinic (University Hospital Basel, Switzerland).
Disclosure
No competing financial interests exist.
References
- Ruiz-Ederra J, Garcia M, Hernandez M, et al. The pig eye as a novel model of glaucoma. Exp Eye Res. 2005;81:561–569. doi: 10.1016/j.exer.2005.03.014. [DOI] [PubMed] [Google Scholar]
- Chader GJ. Animal models research on retinal degeneration: past progress and future hope. Vision Res. 2002;42:393–399. doi: 10.1016/s0042-6989(01)00212-7. [DOI] [PubMed] [Google Scholar]
- Kues WA, Niemann H. The contribution of farm animals to human health. Trends Biotechnol. 2004;22:286–294. doi: 10.1016/j.tibtech.2004.04.003. [DOI] [PubMed] [Google Scholar]
- Pan Z, Sun C, Jie Y, Wang N, Wang L. WZS-pig is a potential donor alternative in corneal xenotransplantation. Xenotransplantation. 2007;14:603–611. doi: 10.1111/j.1399-3089.2007.00432.x. [DOI] [PubMed] [Google Scholar]
- Hara H, Cooper DKC. The immunology of corneal xenotransplantation: a review of the literature. Xenotransplantation. 2010;17:338–349. doi: 10.1111/j.1399-3089.2010.00608.x. [DOI] [PubMed] [Google Scholar]
- Hara H, Cooper DK. Xenotransplantation – the future of corneal transplantation? Cornea. 2011;30:371–378. doi: 10.1097/ICO.0b013e3181f237ef. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim MK, Wee WR, Park C, Kim SJ. Xenocorneal transplantation. Curr Opin Organ Transplant. 2011;16:231–236. doi: 10.1097/MOT.0b013e328344870c. [DOI] [PubMed] [Google Scholar]
- Hara H, Koike N, Long C, et al. Initial in vitro investigation of the human immune response to corneal cells from genetically-engineering pigs. Invest Ophthalmol Vis Sci. 2011;52:5278–5286. doi: 10.1167/iovs.10-6947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekser B, Ezzelarab M, Hara H, et al. Clinical xenotransplantation: the next medical revolution? Lancet. 2012;379:672–683. doi: 10.1016/S0140-6736(11)61091-X. [DOI] [PubMed] [Google Scholar]
- Petters RM, Alexander CA, Wells KD, et al. Genetically engineered large animal model for studying cone photoreceptor survival and degeneration in retinitis pigmentosa. Nat Biotechnol. 1997;15:965–970. doi: 10.1038/nbt1097-965. [DOI] [PubMed] [Google Scholar]
- Tso MO, Li WW, Zhang C, et al. A pathologic study of degeneration of the rod and cone populations of the rhodopsin Pro347Leu transgenic pigs. Trans Am Ophthalmol Soc. 1997;95:467–479. [PMC free article] [PubMed] [Google Scholar]
- Li ZY, Wong F, Chang YH, et al. Rhodopsin transgenic pigs as a model for human retinitis pigmentosa. Invest Ophthalmol Vis Sci. 1998;39:808–819. [PubMed] [Google Scholar]
- Garcia-Layana A, Pastor JC, Saomil MA, Gonzalez G. Porcine model of proliferative vitreoretinopathy with platelets. Curr Eye Res. 1997;16:556–563. doi: 10.1076/ceyr.16.6.556.5082. [DOI] [PubMed] [Google Scholar]
- Scholda C, Biowski R, Simader C, et al. A new sponge profile for retinal detachment surgery: design and in vitro-effectiveness. Acta Ophthalmol Scand. 1999;77:700–703. doi: 10.1034/j.1600-0420.1999.770620.x. [DOI] [PubMed] [Google Scholar]
- Ghosh F, Arner K. Transplantation of full-thickness retina in the normal porcine eye: surgical and morphological aspects. Retina. 2002;22:478–486. doi: 10.1097/00006982-200208000-00013. [DOI] [PubMed] [Google Scholar]
- Warfvinge K, Kiilgaard JF, Lavik EB. Retinal progenitor cell xenografts to the pig retina: morphologic integration and cytochemical differentiation. Arch Ophthalmol. 2005;123:1385–1393. doi: 10.1001/archopht.123.10.1385. [DOI] [PubMed] [Google Scholar]
- Lassota N, Kiilgaard JF, Prause JU, La Cour M. Correlation between clinical and histological features in a pig model of choroidal neovascularization. Graefes Arch Clin Exp Ophthalmol. 2006;244:394–398. doi: 10.1007/s00417-005-0061-0. [DOI] [PubMed] [Google Scholar]
- Klein R, Klein BP, Linton KLP. Prevalence of age-related maculopathy: the beaver eye study. Ophthalmology. 1999;299:933–943. doi: 10.1016/s0161-6420(92)31871-8. [DOI] [PubMed] [Google Scholar]
- Del Priore LV, Tezel TH, Kaplan HJ. Survival of allogeneic porcine retinal pigment epithelial sheets after subretinal transplantation. Invest Ophthalmol Vis Sci. 2004;45:985–992. doi: 10.1167/iovs.03-0662. [DOI] [PubMed] [Google Scholar]
- Valtinik M, Engelmann K. Culturing of retinal pigment epithelium cells. Dev Ophthalmol. 2009;43:109–119. doi: 10.1159/000223844. [DOI] [PubMed] [Google Scholar]
- Melville H, Carpiniello M, Hollis K, Staffaroni A, Golestaneli N. Stem cells: a new paradigm for disease modeling and developing therapies for age-related macular degeneration. J Transl Med. 2013;11:53. doi: 10.1186/1479-5876-11-53. DOI 10.1186/1479-5876-11-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rezai KA, Kohen L, Wiedermann P, Heimann K. Iris pigment epithelium transplantation. Graefes Arch Clin Exp Ophthalmol. 1997;235:558–562. doi: 10.1007/BF00947084. [DOI] [PubMed] [Google Scholar]
- Deshenes J, Char HD, Kaleta S. Activated T-lymphocytes in uveitis. Br J Ophthalmol. 1988;72:83–87. doi: 10.1136/bjo.72.2.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franks WA, Limb GA, Stanford MR, et al. Cytokines in human intraocular inflammation. Curr Eye Res. 1992;11:187–191. doi: 10.3109/02713689208999531. [DOI] [PubMed] [Google Scholar]
- Limb GA, Little BC, Meager A, et al. Cytokines in proliferative vitreoretinopathy. Eye (Lond) 1991;5:686–693. doi: 10.1038/eye.1991.126. [DOI] [PubMed] [Google Scholar]
- Yang J, Cho B, Choi I, et al. Molecular characterization of miniature porcine RANTES and its chemotactic effect on human mononuclear cells. Transplantation. 2006;82:1229–1233. doi: 10.1097/01.tp.0000235603.19527.05. [DOI] [PubMed] [Google Scholar]
- Zhang B, Ge B, Xia X, et al. Compatibility of porcine and human interleukin 2: implications for xenotransplantation. Xenotransplantation. 2006;13:423–432. doi: 10.1111/j.1399-3089.2006.00329.x. [DOI] [PubMed] [Google Scholar]
- Knoblock KF, Canning PC. Modulation of in vitro porcine natural killer cell activity by recombinant interleukin-1 alpha, interleukin-2 and interleukin-4. Immunology. 1992;76:299–304. [PMC free article] [PubMed] [Google Scholar]
- Collins RA, Tayton HK, Gelder KI, Britton P, Oldham G. Cloning and expression of bovine and porcine interleukin-2 in baculovirus and analysis of species cross-reactivity. Vet Immunol Immunopathol. 1994;40:313–324. doi: 10.1016/0165-2427(94)90042-6. [DOI] [PubMed] [Google Scholar]
- Manna PP, Hira SK, Das AA, Bandyopadhyay S, Gupta KK. IL-15 activated human peripheral blood dendritic cell kill allogeneic and xenogeneic endothelial cells via apoptosis. Cytokine. 2013;61:118–126. doi: 10.1016/j.cyto.2012.09.004. [DOI] [PubMed] [Google Scholar]
- Darnell JE, Jr, Kerr IM, Stark GR. JAK-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science. 1994;264:1415–1421. doi: 10.1126/science.8197455. [DOI] [PubMed] [Google Scholar]
- Darnell JE., Jr STATs and gene regulation. Science. 1997;277:1630–1635. doi: 10.1126/science.277.5332.1630. [DOI] [PubMed] [Google Scholar]
- Schindler C. Cytokines and JAK-STAT signaling. Exp Cell Res. 1999;253:7–14. doi: 10.1006/excr.1999.4670. [DOI] [PubMed] [Google Scholar]
- Heim MH. The Jak-STAT pathway: cytokine signalling from the receptor to the nucleus. J Recept Signal Transduct Res. 1999;19:75–120. doi: 10.3109/10799899909036638. [DOI] [PubMed] [Google Scholar]
- Frank DA. STAT signaling in the pathogenesis and treatment of cancer. Mol Med. 1999;5:432–456. [PMC free article] [PubMed] [Google Scholar]
- Spivey TL, Uccellini L, Ascierto ML, et al. Gene expression profiling in acute allograft rejection: challenging the immunologic constant of rejection hypothesis. J Transl Med. 2011;9:174. doi: 10.1186/1479-5876-9-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgun A, Shulzenko N, Perez-Diez A, et al. Molecular profiling improves diagnosis of rejection and infection in transplanted organs. Circ Res. 2006;98:E74–E83. doi: 10.1161/01.res.0000228714.15691.8a. [DOI] [PubMed] [Google Scholar]
- Saint-Mezard P, Berthler CC, Zhang H, et al. Analysis of independent microarray datasets of renal biopsies identifies a robust transcript signature of acute allograft rejection. Transpl Int. 2009;22:293–302. doi: 10.1111/j.1432-2277.2008.00790.x. [DOI] [PubMed] [Google Scholar]
- Patil J, Lande JD, Li N, et al. Bronchoalveolar lavage cell gene expression in acute lung rejection: development of a diagnostic classifier. Transplantation. 2008;85:224–231. doi: 10.1097/TP.0b013e318160268a. [DOI] [PubMed] [Google Scholar]
- Chen R, Sigdel TK, Li L, et al. Differentially expressed RNA from public microarray data identifies serum protein biomarkers for cross-organ transplant rejection and other conditions. PLoS Comput Biol. 2010;6:e1000940. doi: 10.1371/journal.pcbi.1000940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou P, Szot G, Guo Z, et al. Role of STAT4 and STAT6 signaling in allograft rejection and CTLA4-Ig-mediated tolerance. J Immunol. 2000;165:5580–5587. doi: 10.4049/jimmunol.165.10.5580. [DOI] [PubMed] [Google Scholar]
- Wei L, Vahedi G, Sun HW, et al. Discrete roles of STAT4 and STAT6 transcription factors in tuning epigenetic modifications and transcription during T helper cell differentiation. Immunity. 2010;32:840–851. doi: 10.1016/j.immuni.2010.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stojanovic T, Scheele L, Wagner AH, et al. STAT-1 decoy oligonucleotide improves microcirculation and reduces acute rejection in allogeneic rat small bowel transplants. Gene Ther. 2007;14:883–890. doi: 10.1038/sj.gt.3302931. [DOI] [PubMed] [Google Scholar]
- Stojanovic T, Wagner AH, Wang S, et al. STAT-1 decoy oligodeoxynucleotide inhibition of acute rejection in mouse heart transplants. Basic Res Cardiol. 2009;104:719–729. doi: 10.1007/s00395-009-0028-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoelschenmann H, Stadbauer THW, Wagner AH, et al. STAT-1 and AP-1 decoy oligonucleotide therapy delays acute rejection and prolongs cardiac allograft survival. Cardiovasc Res. 2006;71:527–536. doi: 10.1016/j.cardiores.2006.05.021. [DOI] [PubMed] [Google Scholar]
- Stadbauer THW, Wagner AH, Hoelschermann H, et al. AP-1 and STAT-1 decoy oligodeoxynucleotides attenuate transplant vasculopathy in rat cardiac allografts. Cardiovasc Res. 2008;79:698–705. doi: 10.1093/cvr/cvn135. [DOI] [PubMed] [Google Scholar]
- Wunderlich K, Knorr M, Dartsch P. Serum-free cultivation of bovine lens epithelial cells. Graefes Arch Clin Exp Ophthalmol. 1994;232:355–360. doi: 10.1007/BF00175987. [DOI] [PubMed] [Google Scholar]
- Fasler-Kan E, Barteneva N, Ketterer S, et al. Activation of the JAK-STAT intracellular pathway in human retinal pigment epithelial cell line ARPE-19. Int J Interferon Cytokine Mediator Res. 2010;2:127–136. [Google Scholar]
- Pansky A, Hildebrand P, Fasler-Kan E, et al. Defective JAK-STAT signal transduction pathway in melanoma cells resistant to growth inhibition by interferon-α. Int J Cancer. 2000;85:720–725. doi: 10.1002/(sici)1097-0215(20000301)85:5<720::aid-ijc20>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- Fasler-Kan E, Pansky A, Wiederkehr M, Battegay M, Heim M. Interferon-a activates signal transducers and activators of transcription 5 and 6 in Daudi cells. Eur J Biochem. 1998;254:514–519. doi: 10.1046/j.1432-1327.1998.2540514.x. [DOI] [PubMed] [Google Scholar]
- Le Bas-Bernardet S, Blancho G. Current cellular immunological hurdles in pig-to-primate xenotransplantation. Transpl Immunol. 2009;21:60–64. doi: 10.1016/j.trim.2008.10.006. [DOI] [PubMed] [Google Scholar]
- Simon AR, Warrens AN, Yazzie NP, et al. Cross-species interaction of porcine and human integrins with their respective ligands: implications for xenogeneic tolerance induction. Transplantation. 1998;66:385–394. doi: 10.1097/00007890-199808150-00017. [DOI] [PubMed] [Google Scholar]
- Warrens AN, Simon AR, Theodore PR, Sykes M. Human-porcine receptor-ligand compatibility within the immune system: relevance for xenotransplantation. Xenotransplantation. 1999;6:75–78. doi: 10.1034/j.1399-3089.1999.00020.x. [DOI] [PubMed] [Google Scholar]
- Rusworth SA, Bravery CA, Thompson S. Human CD154 induces activation of porcine endothelial cells and up-regulation of MHC class II expression. Transplantation. 2001;72:127–132. doi: 10.1097/00007890-200107150-00025. [DOI] [PubMed] [Google Scholar]
- Murray AG, Khodadoust MM, Pober JS, Bothell ALM. Porcine aortic endothelial cells activate human T-cells: direct presentation of MHC antigens and costimulation by ligands for human CD2 and Cd28. Immunity. 1994;1:57–63. doi: 10.1016/1074-7613(94)90009-4. [DOI] [PubMed] [Google Scholar]
- Maher SE, Karmann K, Min W, et al. Porcine endothelial CD86 is a major costimulator of xenogeneic human T cells: cloning, sequencing, and functional expression in human endothelial cells. J Immunol. 1996;157:3838. [PubMed] [Google Scholar]
- Rollins SA, Kennedy SP, Chodera AJ, et al. Evidence that activation of human T cells by porcine endothelium involves direct recognition of porcine SLA and costimulation by porcine ligands for LFA-1 and CD2. Transplantation. 1994;57:1709–1716. [PubMed] [Google Scholar]
- Sang Y, Rowland RRR, Hesse RA, Blecha F. Differential expression and activity of the porcine type I interferon family. Physiol Genomics. 2010;42:248–258. doi: 10.1152/physiolgenomics.00198.2009. [DOI] [PubMed] [Google Scholar]
- Zhao X, Cheng G, Jiao Y. Cloning and characterization of porcine interferon-delta-related genes identified by genomic database screening. J Interferon Cytokine Res. 2012;32:378–385. doi: 10.1089/jir.2011.0095. et al.. [DOI] [PubMed] [Google Scholar]
- Ezzelrab C, Ayares D, Cooper DKC, Ezzelrab MB. Human T-cell proliferation in response to thrombin-activated GTKO pig endothelial cells. Xenotransplantation. 2012;19:311–316. doi: 10.1111/j.1399-3089.2012.00718.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang J, Ma G, Zhu M, et al. Molecular characterization of the porcine STAT4 and STAT6 genes. Mol Biol Rep. 2012;39:6959–6965. doi: 10.1007/s11033-012-1523-1. [DOI] [PubMed] [Google Scholar]
- Tojo N, Kashiwagi Y, Nishitsuka K, et al. Interactions between vitreous-derived cells and vascular endothelial cells in vitreoretinal diseases. Acta Ophthalmol. 2010;88:564–570. doi: 10.1111/j.1755-3768.2008.01466.x. [DOI] [PubMed] [Google Scholar]
- Lee KG, Lee H, Ha JM, et al. Increased human tumor necrosis factor-alpha induce procoagulant change in porcine endothelial cells in vitro. Xenotransplantation. 2012;19:186–195. doi: 10.1111/j.1399-3089.2012.00704.x. [DOI] [PubMed] [Google Scholar]
- Ihle JN, Kerr IM. Jaks and Stats in signaling by the cytokine receptor superfamily. Trends Genet. 1995;11:69–74. doi: 10.1016/s0168-9525(00)89000-9. [DOI] [PubMed] [Google Scholar]
- Stark GR, Darnell JE., Jr The JAK-STAT pathway at twenty. Immunity. 2012;36:503–514. doi: 10.1016/j.immuni.2012.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang T, Niu G, Kotylewski M, et al. Regulation of the innate and adaptive immune responses by STAT-3 signaling in tumor cells. Nat Med. 2004;10:48–54. doi: 10.1038/nm976. [DOI] [PubMed] [Google Scholar]
- Yagil Z, Nechustan H, Kay G, et al. The enigma of the role of protein inhibitor of activated STAT 3 (PIA S3) in the immune response. Trends Immunol. 2010;31:199–204. doi: 10.1016/j.it.2010.01.005. [DOI] [PubMed] [Google Scholar]
- Hix LM, Karavatis J, Khan MW, et al. Tumor STAT1 transcription factor activity enhances breast tumor growth and immune suppression mediated by myeloid-derived suppressor cells. J Biol Chem. 2013;288:11676–11688. doi: 10.1074/jbc.M112.441402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guduric-Fuchs J, Chen W, Price H, Archer DB, Cogliati T. RPE and neuronal differentiation of allotransplantated porcine ciliary epithelium-derived cells. Mol Vis. 2011;17:2580–2595. [PMC free article] [PubMed] [Google Scholar]