

Abstract
There is increasing evidence in the literature pointing to an important role of inflammation during initiation and progression of cancer. Glioblastoma is the most common malignant primary brain tumor with approximately 23,000 newly-diagnosed cases each year in the United States, and has a dismal median survival of only 15 months. Although the blood-brain barrier maintains an immune-privileged status of the brain under steady state, intracranial tumors including gliomas are invariably infiltrated with various types of immune cells. The T helper 17 (Th17) cells, a recently discovered interleukin (IL)-17-producing T cell subtype, have been reported in several extracranial and some intracranial tumors, where they have been implicated in either pro- or antitumor activity depending on the tumor type. Here, we present a succinct review of the current literature on the prevalence and potential role of IL-17 in malignant gliomas. Further mechanistic studies on IL-17 mediated inflammatory pathway in gliomas may provide with opportunities for novel immunotherapeutic interventions.
Keywords: Malignant glioma, Th17, IL-17, Inflammation, Brain cancer, T cells
Introduction
Infiltrating immune cells comprise a major component of solid tumors; and it has been well appreciated that the progression of malignant tumors is greatly influenced by the cytokine profile as well as the type and density of immune cells in the tumor microenvironment [1–6]. Glioblastoma is the most common malignant primary brain tumor with approximately 23,000 newly-diagnosed cases each year in the United States [7], and has a dismal median survival of only 15 months [8]. Although the blood-brain barrier maintains an immune-privileged status of the brain under steady state, intracranial tumors including gliomas are invariably infiltrated with various types of immune cells. Among these, the roles of CD8+ cytolytic T lymphocytes (CTL), regulatory T (Treg) cells and myeloid derived suppressor cells (MDSCs) in the progression of tumor have been quite well studied. The T helper 17 (Th17) cells, a recently discovered interleukin (IL)-17-producing T cell subtype [9], have also been reported in several extracranial tumors, where they have been implicated in either proor antitumor activity depending on the tumor type [10]. IL-17 is one of the most potent inflammatory cytokine and has been strongly involved in inflammatory autoimmune disorders such as rheumatoid arthritis and multiple sclerosis [11,12]. A mechanistic insight into the role of Th17 cells and cytokine IL-17 in the progression of malignant gliomas may provide additional avenues for investigation and therapeutic manipulation of this novel inflammatory axis, ultimately leading to improved survival. Here, we present a succinct review of the current literature on the prevalence and potential role of IL-17 in malignant gliomas.
Th17 cells
The Th17 cells were recently identified as an independent subtype of inflammatory T cells with a distinct cytokine (IL-17) and transcription factor retinoid-related orphan receptor (ROR)gt profile [9]. Naïve CD4+ T cells can be induced to differentiate towards Th1, Th2, Th17, and regulatory T cell (Treg) phenotypes according to the local cytokine milieu. Each T cell phenotype is characterized by unique signalling pathways and expression of specific transcription factors, notably T-bet for Th1, GATA-3 for Th2, forkhead box P3 (FoxP3) for Tregs, and RORa and RORgt and RORγt for Th17 cells [13]. Peripheral Th17 cells have been reported to efficiently migrate to the brain in autoimmune conditions [14]. There are very few studies investigating the recruitment or origin of Th17 cells in intracranial tumors.
Malignant gliomas have been known to secrete transforming growth factor (TGF)-β1 and IL-6 [15–18]; and in mouse models, Treg and Th17 cells have been demonstrated to arise from common precursors in a reciprocal manner based on exposure to TGF-β or TGF-β plus IL-6, respectively [9,19]. In humans however, TGF-β may not be necessary for the induction of Th17 phenotype [20]. A number of groups have now described conversion of Treg to the Th17 phenotype or vice-versa, induced by appropriate inflammatory stimuli [21]. Unlike the rapid advances in understanding their role in inflammation and autoimmunity, there are few studies on the activity of Th17 cells in cancer, and the results are controversial [10,22,23].
Myeloid (CD11b+) cells are predominant among immune cell that infiltrate malignant gliomas, constituting up to 30% of total cells and are often the first ones to be recruited [24–26]. IL-1β, along with IL-6 and IL-23 are critical in the induction and maintenance of Th17 phenotype in humans [27–31]. Tumor-associated monocytes have been shown to induce and expand Th17 cells via secretion of IL-1β and IL-23 in ovarian carcinoma [31] and hepatocellular carcinoma [32,33]. Tumor-associated macrophages expressed higher levels of IL-1β than normal tissue macrophages and normal monocyte-derived macrophages [22]. Thus, molecular mechanisms involved in inducing Th17 cells in patients with tumors can be different from those in patients with autoimmune disorders.
Th17 cells associated with autoimmune disorders often convert to Th1-like phenotype via co-expression of interferon (IFN)-γ and are implicated in cytotoxicity [34,35]. In terms of intratumoral Th17 cells, they might not mediate direct antitumor activity but can promote antitumor immunity indirectly through the recruitment of dendritic cells and cytotoxic effector cells via production of the CCL20 chemokine [36,37]. Antitumor activity of a novel population of IL-17+/IFN-γ+ CD8 T cells (known as Tc17 cells) have also been reported in certain cancers [38,39] and interestingly, non-cytotoxic Tc17 cells could be rendered cytotoxic upon treatment with IL-12 [40]. These studies demonstrate that the polarity and functions of tumor-infiltrating immune cell subsets clearly depend on the cytokine profile in the tumor microenvironment. Th17 cells are very plastic and can switch to non-cytotoxic (low IFN-γ, high IL-10) functional phenotype depending on the cytokines present in the milieu [21,41,42]. Induction of Th17 cells in the presence of TGF-β1 may lead to generation of non-cytotoxic (low IFN-γ, high IL-10) and often pro-tumor Th17 cells [43,44]. We recently reported that factors derived from glioma and glioma-activated myeloid cells could mediate the generation or expansion of intratumoral Th17 cells, and glioma-associated Th17 cells were potentially non-cytotoxic and may also contribute to immune suppression via secretion of IL-10 [20].
Significance of IL-17 in Malignant Tumors
IL-17 (also called IL-17A) is the prototypic member of the IL-17 family composed of six cytokines (IL-17A–F). IL-17A shares the greatest homology with IL-17F. Both IL-17A and IL-17F signal through IL-17RA, a type I transmembrane receptor [45]. Unlike the rapid advances in understanding their role in inflammation and autoimmunity, there is a paucity of reports studying the activity of IL-17 in cancer and the results are quite conflicting.
Prevalence of Th17 cells in the tumor produced an antitumor effect in a murine model of pancreatic cancer [46]. Th17 cells were shown to represent an important fraction of the tumor-infiltrating lymphocytes (TILs) in head and neck squamous cell carcinoma, where they were implicated in antitumor activity [47]. Kryczek et al. analyzed Th17 cell phenotype and function in 201 ovarian cancer patients and proposed that Th17 cells may contribute to protective tumor immunity through recruitment and induction of Th1-type effector cells to the tumor [33]. In another retrospective study of clinical specimens from 104 patients with stage III–IV epithelial ovarian cancer, Lan et al. report a positive correlation between high IL-17 expression and progression-free survival (PFS) in advanced ovarian cancer, although no significant difference was observed in overall survival between the high and low IL-17 expression groups [48].
On the other hand, Chen et al. studied tumor specimens from 207 patients with breast carcinoma and observed that tumors with high numbers of IL-17 producing cells correlated with high histological grade and these patients had shorter disease-free survival than patients with tumors containing low number of IL-17 producing cells [49]. He et al. demonstrated that IL-17-mediated responses promote tumor development through the induction of a tumor-promoting microenvironment at tumor sites [50]. IL-17-mediated regulation of myeloid-derived suppressor cells (MDSCs) was found to be a primary mechanism for its tumor-promoting effects. In another study, IL-17 promoted multiple myeloma (MM) cell growth and colony formation in a murine xenograft model of human MM [51]. IL-17 also enhanced the formation and development of tumor in carcinogen-induced skin cancer models [1,52], in B16 melanoma and MB49 bladder carcinoma models [37], as well as in a prostate cancer model [53]. In an earlier study, IL-17 suppressed apoptosis of several tumor cell lines in vitro [54], suggesting that IL-17 could promote tumorigenesis directly. Consistent with this hypothesis, in the same study, knockdown of the IL-17 receptor in 4T1 mouse mammary cancer cells enhanced apoptosis and decreased tumor growth in vivo [54].
A recent report by Straus showed that IL-17 could positively interact with tumor necrosis factor (TNF)-α to stimulate glycolysis and growth factor production in colorectal cancer cells, which involved Akt and NF-kB signalling activity [55]. In another study, Chung et al. have implicated IL-17 mediated paracrine network in the promotion of tumor resistance to anti-angiogenic therapy. The report shows that IL-17 could induce secretion of granulocyte-colony stimulating factor (GCSF) in tumor cells via NF-αB and ERK signalling, which could lead to recruitment of immature myeloid cells into the tumor microenvironment [56]. Benevides et al. reported that both Treg and Th17 cells were synchronically increased in patients with invasive ductal carcinoma (IDC) of the breast, with positive correlation with tumor aggressiveness [57]. In this context, a small population of IL-17+ Foxp3+ cells, detected in patients with ulcerative colitis, have been implicated to ‘mechanistically link human chronic inflammation to tumor development’ via induction of inflammatory cytokines and inhibition of local T cell immunity [58]. In a murine model of colon carcinoma, blockade of IL-17A significantly suppressed the tumor growth by inhibiting angiogenesis via reduced expression of MMP9 and VEGF, while also activating the cytotoxic T lymphocytes at the tumor site [59].
Zhang et al. have reported increased prevalence of Th17 cells in patients with gastric cancer, which was associated with poor progression [60]. The frequency of Th17 cells also showed negative correlation with time to disease progression in prostate cancer patients receiving a dendritic cell-based vaccine [61]. In another study, patients with medulloblastoma had a considerable population of Th17 cells in tumor-infiltrating T-cells, along with high expression of IL-17 in tumor tissues [62]. Moreover, the serum levels of IL-17 were significantly increased in patients with medulloblastoma compared to normal controls, indicating that Th17 cells and IL-17 may contribute to medulloblastoma pathogenesis [62]. Recently, Wainwright et al. demonstrated mRNA expression of IL-17A in human gliomas [63]. In the same study, the authors also identified Th17 cells in mouse brain tumors [63]. However, the role of IL-17 in gliomas was not investigated.
Collectively, these results demonstrate that tumor-infiltrating Th17 cells can have either pro- or antitumor activities depending on the type of neoplasm, while IL-17 by itself has been mostly associated with tumor promotion. The variable role of CD4+ Th17 cells from tumor to tumor may be because of their plasticity, which changes depending on the presence of other cytokines such as TGF-β, TNF-α, IL-1, IL-2 and IFN-γ in the tumor milieu [20,44,64,65]. Moreover, Th17 is not the sole source of IL-17 within tumors. Myeloid cells, especially macrophages, are also known to be a major source of IL-17 production [11]. Therefore, the role of IL-17 in the progression of malignant tumors should be considered independent of Th17.
Recently, a few investigators, including our group, have described a potential role of Th17 or the cytokine IL-17 in the progression of malignant gliomas. In a study with 41 tumors derived from patients with malignant glioma, Cui et al. reported a direct correlation between 2-year progression-free survival and low incidence of IL-17 producing cells, suggesting that presence of IL-17-producing cells may be a good prognostic marker in malignant gliomas [66]. However, the authors did not study the status of TGF-β1 or Treg cells vis-à-vis IL-17-producing cells in the patient samples. Moreover, the mechanism of how IL-17 could influence glioma progression was not studied. Hu et al. investigated IL-17 mRNA expression in tumor tissues derived from 24 patients with malignant gliomas. They reported higher expression of IL-17 mRNA in 18/24 glioma patients compared to that in tissues derived from trauma patients, while no difference was observed among various histologic grades of gliomas [67].
Two studies have investigated the prevalence of Th17 cells in murine models of malignant gliomas. Wainwright et al. demonstrated mRNA expression of IL-17A in human glioma [63]. The group also noted presence of Th17 cells in mouse brain tumors; however, they did not investigate the prevalence of Th17 cells in human gliomas, their mechanism of induction or expansion, or their immunologic functions [63]. In another study, Cantini et al. isolated Th17 cells from the spleen of naïve mice (nTh17) and glioma-bearing mice (gTh17) and co-transplanted them intracranially with GL261 glioma cells in immune-competent mice [68]. Mice co-injected with GL261 and nTh17 survived significantly longer compared to mice co-injected with GL261 and gTh17. Moreover, glioma co-transplanted with nTh17 expressed high levels of IFN-γ and TNF-α and low levels of IL-10 and TGF-β [68]. Studies from our group also demonstrated that Th17 cells co-cultured with glioma cells produced lower levels of IFN-γ and higher levels of IL-10 compared to naïve Th17 cells [20]. The tumor-cytolytic activity of Th17 cells was also inhibited following co-culture with gliomas [20].
IL-17R in Malignant Gliomas
The receptors for IL-17 family of cytokines consist of 5 members (IL-17RA, RB, RC, RD and RE), all of which share sequence homology to IL-17RA. The receptor for IL-17A, IL-17F, and the IL-17A/F heterodimer is composed of IL-17RA paired with IL-17RC. IL-17RA appears to be a component of at least one other receptor complex (IL-25R), and may in fact be the common signalling subunit of the entire family [11,69]. IL-17 has been shown to stimulate the expression of IαBα mRNA and the secretion of IL-6 and IL-8 in glioma cell, suggesting an expression of functional IL-17R in gliomas [57]. In a recent report by Schwartzbaum et al., a significant positive correlation was observed between the expression of CD133 mRNA and the levels for IL-17 and IL-17R mRNA in gliomas, whereas there was a negative correlation between most other inflammatory molecules and CD133 expression [70]. However, actual co-localization of IL-17R proteins along with CD133 in the tumor cells was not investigated. IL-17 induces expression of heparin-binding epidermal growth factor-like growth factor (HB-EGF) in airway smooth muscle cells [71] and in synovial fibroblasts [72]. IL-17, in combination with TNF-α, has been shown to induce CXCL8 and CXCL10 gene expression via EGFR transactivation [73]. It remains to be studied whether biological effects of IL-17 in cancer cells is mediated directly via IL-17R activation or, at least in part, via EGFR signalling. Unpublished observations in our laboratory have indicated the expression of functional IL-17R in CD133+ glioma stem cells (GSCs), as determined by immunohistochemistry and flow cytometry of primary glioma specimens. Moreover, preliminary results of our in vitro ‘neurospheroid’ assay and western blot analysis have suggested that IL-17–IL-17R interaction in GSCs could lead to proliferation and cytokine secretion via modulation of STAT-3 and GSK-3β/β-catenin signalling (Parajuli et al., unpublished observation). These data suggested that IL-17–IL-17R interaction may play an important role during the progression of gliomas.
Conclusion
Studies with extracranial tumors suggest a pleiotropic role of Th17 cells in tumor progression, which is dependent on cytokines present in the tumor microenvironment. Th17 cells expressing high IFN-γ could be tumor-cytolytic, while those expressing high IL-10 could promote tumor growth. On the other hand, the cytokine IL-17 itself seems to promote tumor growth, either via its inflammatory activity on other immune cells or via direct effect on the tumor (stem) cells (Figure 1). However, there are still vast lacunae in our knowledge regarding the interaction of tumor cells and various stromal immune components during the progression of malignant gliomas. Specifically, several aspects of the biology of intratumoral Th17 cells and the cytokine IL-17 still remain to be investigated in order to firmly understand their role in the progression or recurrence of malignant gliomas.
Figure 1.
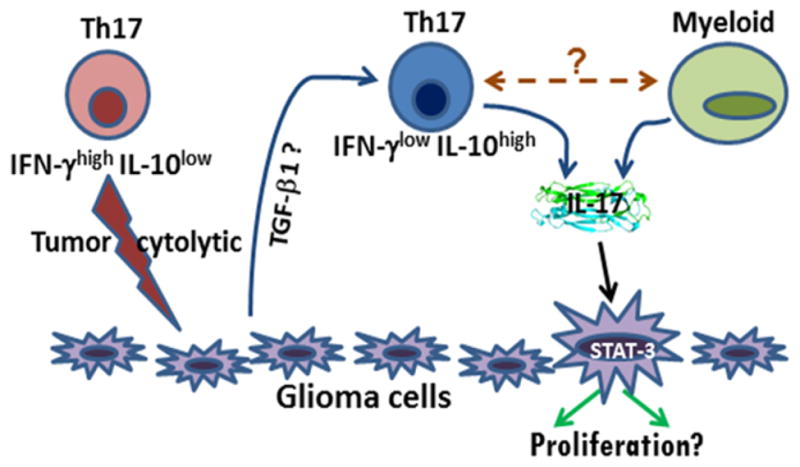
A schematic representation of possible mechanisms of glioma-Th17/IL-17 interaction.
Naïve Th17 cells (or Th17 cells in gliomas with low TGF-β levels) may express high IFN-γ, low IL-10, and show anti-glioma cytolytic activity. On the other hand, glioma derived factors, mainly TGF-β1; inhibit cytolytic activity of glioma-associated Th17 cells. Th17 cells may also influence glioma progression indirectly, via their effect on other immune cells, including MDSCs. Moreover, a small percentage of glioma cells (possibly GSCs) express IL-17R, which, upon interaction with IL-17, could lead to glioma progression via activation of STAT-3 signaling.
References
- 1.He D, Li H, Yusuf N, Elmets CA, Athar M, et al. IL-17 mediated inflammation promotes tumor growth and progression in the skin. PLoS One. 2012;7:e32126. doi: 10.1371/journal.pone.0032126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cole SW. Chronic inflammation and breast cancer recurrence. J Clin Oncol. 2009;27:3418–3419. doi: 10.1200/JCO.2009.21.9782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aggarwal BB, Gehlot P. Inflammation and cancer: how friendly is the relationship for cancer patients? Curr Opin Pharmacol. 2009;9:351–369. doi: 10.1016/j.coph.2009.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shankaran V, Ikeda H, Bruce AT, White JM, Swanson PE, et al. IFNgamma and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature. 2001;410:1107–1111. doi: 10.1038/35074122. [DOI] [PubMed] [Google Scholar]
- 5.Vega EA, Graner MW, Sampson JH. Combating immunosuppression in glioma. Future Oncol. 2008;4:433–442. doi: 10.2217/14796694.4.3.433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, et al. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science. 2006;313:1960–1964. doi: 10.1126/science.1129139. [DOI] [PubMed] [Google Scholar]
- 7.Dolecek TA, Propp JM, Stroup NE, Kruchko C. CBTRUS statistical report: primary brain and central nervous system tumors diagnosed in the United States in 2005–2009. Neuro Oncol. 2012;14:v1–49. doi: 10.1093/neuonc/nos218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009;10:459–66. doi: 10.1016/S1470-2045(09)70025-7. [DOI] [PubMed] [Google Scholar]
- 9.Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, et al. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–238. doi: 10.1038/nature04753. [DOI] [PubMed] [Google Scholar]
- 10.Bronte V. Th17 and cancer: friends or foes? Blood. 2008;112:214. doi: 10.1182/blood-2008-04-149260. [DOI] [PubMed] [Google Scholar]
- 11.Gaffen SL. Recent advances in the IL-17 cytokine family. Curr Opin Immunol. 2011;23:613–619. doi: 10.1016/j.coi.2011.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wilke CM, Bishop K, Fox D, Zou W. Deciphering the role of Th17 cells in human disease. Trends Immunol. 2011;32:603–611. doi: 10.1016/j.it.2011.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhu J, Paul WE. Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immunol Rev. 2010;238:247–262. doi: 10.1111/j.1600-065X.2010.00951.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Okada H, Khoury SJ. Type17 T-cells in central nervous system autoimmunity and tumors. J Clin Immunol. 2012;32:802–808. doi: 10.1007/s10875-012-9686-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Schneider J, Hofman FM, Apuzzo ML, Hinton DR. Cytokines and immunoregulatory molecules in malignant glial neoplasms. J Neurosurg. 1992;77:265–273. doi: 10.3171/jns.1992.77.2.0265. [DOI] [PubMed] [Google Scholar]
- 16.Parajuli P, Mathupala S, Mittal S, Sloan AE. Dendritic cell-based active specific immunotherapy for malignant glioma. Expert Opin Biol Ther. 2007;7:439–448. doi: 10.1517/14712598.7.4.439. [DOI] [PubMed] [Google Scholar]
- 17.Zou JP, Morford LA, Chougnet C, Dix AR, Brooks AG, et al. Human glioma-induced immunosuppression involves soluble factor(s) that alters monocyte cytokine profile and surface markers. J Immunol. 1999;162:4882–4892. [PubMed] [Google Scholar]
- 18.Gomez GG, Kruse CA. Cellular and functional characterization of immunoresistant human glioma cell clones selected with alloreactive cytotoxic T lymphocytes reveals their up-regulated synthesis of biologically active TGF-beta. J Immunother. 2007;30:261–73. doi: 10.1097/01.cji.0000211339.81211.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Morishima N, Mizoguchi I, Takeda K, Mizuguchi J, Yoshimoto T. TGF-beta is necessary for induction of IL-23R and Th17 differentiation by IL-6 and IL-23. Biochem Biophys Res Commun. 2009;386:105–110. doi: 10.1016/j.bbrc.2009.05.140. [DOI] [PubMed] [Google Scholar]
- 20.Paladugu M, Thakur A, Lum LG, Mittal S, Parajuli P. Generation and immunologic functions of Th17 cells in malignant gliomas. Cancer Immunol Immunother. 2013;62:75–86. doi: 10.1007/s00262-012-1312-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhu J, Paul WE. Heterogeneity and plasticity of T helper cells. Cell Res. 2010;20:4–12. doi: 10.1038/cr.2009.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zou W, Restifo NP. T(H)17 cells in tumour immunity and immunotherapy. Nat Rev Immunol. 2010;10:248–256. doi: 10.1038/nri2742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ji Y, Zhang W. Th17 cells: positive or negative role in tumor? Cancer Immunol Immunother. 2010;59:979–987. doi: 10.1007/s00262-010-0849-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hussain SF, Yang D, Suki D, Aldape K, Grimm E, et al. The role of human glioma-infiltrating microglia/macrophages in mediating antitumor immune responses. Neuro Oncol. 2006;8:261–279. doi: 10.1215/15228517-2006-008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Leung SY, Wong MP, Chung LP, Chan AS, Yuen ST. Monocyte chemoattractant protein-1 expression and macrophage infiltration in gliomas. Acta Neuropathol. 1997;93:518–527. doi: 10.1007/s004010050647. [DOI] [PubMed] [Google Scholar]
- 26.Takeshima H, Kuratsu J, Takeya M, Yoshimura T, Ushio Y. Expression and localization of messenger RNA and protein for monocyte chemoattractant protein-1 in human malignant glioma. J Neurosurg. 1994;80:1056–1062. doi: 10.3171/jns.1994.80.6.1056. [DOI] [PubMed] [Google Scholar]
- 27.Aggarwal S, Ghilardi N, Xie MH, de Sauvage FJ, Gurney AL. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J Biol Chem. 2003;278:1910–1914. doi: 10.1074/jbc.M207577200. [DOI] [PubMed] [Google Scholar]
- 28.Yu RY, Gallagher G. A naturally occurring, soluble antagonist of human IL-23 inhibits the development and in vitro function of human Th17 cells. J Immunol. 2010;185:7302–7308. doi: 10.4049/jimmunol.1002410. [DOI] [PubMed] [Google Scholar]
- 29.Aliahmadi E, Gramlich R, Grützkau A, Hitzler M, Krüger M, et al. TLR2- activated human langerhans cells promote Th17 polarization via IL-1beta, TGF-beta and IL-23. Eur J Immunol. 2009;39:1221–1230. doi: 10.1002/eji.200838742. [DOI] [PubMed] [Google Scholar]
- 30.Sha Y, Markovic-Plese S. A role of IL-1R1 signaling in the differentiation of Th17 cells and the development of autoimmune diseases. Self Nonself. 2011;2:35–42. doi: 10.4161/self.2.1.15639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Miyahara Y, Odunsi K, Chen W, Peng G, Matsuzaki J, et al. Generation and regulation of human CD4+ IL-17-producing T cells in ovarian cancer. Proc Natl Acad Sci U S A. 2008;105:15505–15510. doi: 10.1073/pnas.0710686105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kuang DM, Peng C, Zhao Q, Wu Y, Chen MS, et al. Activated monocytes in peritumoral stroma of hepatocellular carcinoma promote expansion of memory T helper 17 cells. Hepatology. 2010;51:154–164. doi: 10.1002/hep.23291. [DOI] [PubMed] [Google Scholar]
- 33.Kryczek I, Banerjee M, Cheng P, Vatan L, Szeliga W, et al. Phenotype, distribution, generation, and functional and clinical relevance of Th17 cells in the human tumor environments. Blood. 2009;114:1141–9. doi: 10.1182/blood-2009-03-208249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Domingues HS, Mues M, Lassmann H, Wekerle H, Krishnamoorthy G. Functional and pathogenic differences of Th1 and Th17 cells in experimental autoimmune encephalomyelitis. PLoS One. 2010;5:e15531. doi: 10.1371/journal.pone.0015531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Annunziato F, Romagnani S. The transient nature of the Th17 phenotype. Eur J Immunol. 2010;40:3312–3316. doi: 10.1002/eji.201041145. [DOI] [PubMed] [Google Scholar]
- 36.Wu S, Rhee KJ, Albesiano E, Rabizadeh S, Wu X, et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med. 2009;15:1016–1022. doi: 10.1038/nm.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang L, Yi T, Kortylewski M, Pardoll DM, Zeng D, et al. IL-17 can promote tumor growth through an IL-6-Stat3 signaling pathway. J Exp Med. 2009;206:1457–1464. doi: 10.1084/jem.20090207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jiang Li, Zhou-Feng H, Xiong G, Hao-Yuan M, Qiu F, et al. Distribution, characterization, and induction of CD8+ regulatory T cells and IL-17− producing CD8+ T cells in nasopharyngeal carcinoma. J Transl Med. 2011;9:189. doi: 10.1186/1479-5876-9-189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.de Garcia-Hernandez ML, Hamada H, Reome JB, Misra SK, Tighe MP, et al. Adoptive transfer of tumor-specific Tc17 effector T cells controls the growth of B16 melanoma in mice. J Immunol. 2010;184:4215–4227. doi: 10.4049/jimmunol.0902995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tajima M, Wakita D, Satoh T, Kitamura H, Nishimura T. IL-17/IFN-γ double producing CD8+ T (Tc17/IFN-γ) cells: a novel cytotoxic T-cell subset converted from Tc17 cells by IL-12. Int Immunol. 2011;23:751–759. doi: 10.1093/intimm/dxr086. [DOI] [PubMed] [Google Scholar]
- 41.Zhou L, Chong MM, Littman DR. Plasticity of CD4+ T cell lineage differentiation. Immunity. 2009;30:646–655. doi: 10.1016/j.immuni.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 42.Nistala K, Adams S, Cambrook H, Ursu S, Olivito B, et al. Th17 plasticity in human autoimmune arthritis is driven by the inflammatory environment. Proc Natl Acad Sci U S A. 2010;107:14751–14756. doi: 10.1073/pnas.1003852107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Volpe E, Touzot M, Servant N, Marloie-Provost MA, Hupé P, et al. Multiparametric analysis of cytokine-driven human Th17 differentiation reveals a differential regulation of IL-17 and IL-22 production. Blood. 2009;114:3610–3614. doi: 10.1182/blood-2009-05-223768. [DOI] [PubMed] [Google Scholar]
- 44.Alshaker HA, Matalka KZ. IFN-γ, IL-17 and TGF-β involvement in shaping the tumor microenvironment: The significance of modulating such cytokines in treating malignant solid tumors. Cancer Cell Int. 2011;11:33. doi: 10.1186/1475-2867-11-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Maniati E, Soper R, Hagemann T. Up for Mischief? IL-17/Th17 in the tumour microenvironment. Oncogene. 2010;29:5653–5662. doi: 10.1038/onc.2010.367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gnerlich JL, Mitchem JB, Weir JS, Sankpal NV, Kashiwagi H, et al. Induction of Th17 cells in the tumor microenvironment improves survival in a murine model of pancreatic cancer. J Immunol. 2010;185:4063–4071. doi: 10.4049/jimmunol.0902609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kesselring R, Thiel A, Pries R, Trenkle T, Wollenberg B. Human Th17 cells can be induced through head and neck cancer and have a functional impact on HNSCC development. Br J Cancer. 2010;103:1245–1254. doi: 10.1038/sj.bjc.6605891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lan C, Huang X, Lin S, Huang H, Cai Q, et al. High density of IL-17− producing cells is associated with improved prognosis for advanced epithelial ovarian cancer. Cell Tissue Res. 2013;352:351–359. doi: 10.1007/s00441-013-1567-0. [DOI] [PubMed] [Google Scholar]
- 49.Chen WC, Lai YH, Chen HY, Guo HR, Su IJ, et al. Interleukin-17- producing cell infiltration in the breast cancer tumour microenvironment is a poor prognostic factor. Histopathology. 2013;63:225–233. doi: 10.1111/his.12156. [DOI] [PubMed] [Google Scholar]
- 50.He D, Li H, Yusuf N, Elmets CA, Li J, et al. IL-17 promotes tumor development through the induction of tumor promoting microenvironments at tumor sites and myeloid-derived suppressor cells. J Immunol. 2010;184:2281–2288. doi: 10.4049/jimmunol.0902574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Prabhala RH, Pelluru D, Fulciniti M, Prabhala HK, Nanjappa P, et al. Elevated IL-17 produced by TH17 cells promotes myeloma cell growth and inhibits immune function in multiple myeloma. Blood. 2010;115:5385–5392. doi: 10.1182/blood-2009-10-246660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang L, Yi T, Zhang W, Pardoll DM, Yu H. IL-17 enhances tumor development in carcinogen-induced skin cancer. Cancer Res. 2010;70:10112–10120. doi: 10.1158/0008-5472.CAN-10-0775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Yang SX, Chen JH, Jiang XF, Wang QL, Chen ZQ, et al. Activation of chemokine receptor CXCR4 in malignant glioma cells promotes the production of vascular endothelial growth factor. Biochem Biophys Res Commun. 2005;335:523–528. doi: 10.1016/j.bbrc.2005.07.113. [DOI] [PubMed] [Google Scholar]
- 54.Nam JS, Terabe M, Kang MJ, Chae H, Voong N, et al. Transforming growth factor beta subverts the immune system into directly promoting tumor growth through interleukin-17. Cancer Res. 2008;68:3915–3923. doi: 10.1158/0008-5472.CAN-08-0206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Straus DS. TNF-α and IL-17 cooperatively stimulate glucose metabolism and growth factor production in human colorectal cancer cells. Mol Cancer. 2013;12:78. doi: 10.1186/1476-4598-12-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chung AS, Wu X, Zhuang G, Ngu H, Kasman I, et al. An interleukin-17-mediated paracrine network promotes tumor resistance to anti-angiogenic therapy. Nat Med. 2013;19:1114–1123. doi: 10.1038/nm.3291. [DOI] [PubMed] [Google Scholar]
- 57.Benevides L, Cardoso CR, Tiezzi DG, Marana HR, Andrade JM, et al. Enrichment of regulatory T cells in invasive breast tumor correlates with the upregulation of IL-17A expression and invasiveness of the tumor. Eur J Immunol. 2013;43:1518–1528. doi: 10.1002/eji.201242951. [DOI] [PubMed] [Google Scholar]
- 58.Kryczek I, Wu K, Zhao E, Wei S, Vatan L, et al. IL-17+ regulatory T cells in the microenvironments of chronic inflammation and cancer. J Immunol. 2011;186:4388–4395. doi: 10.4049/jimmunol.1003251. [DOI] [PubMed] [Google Scholar]
- 59.Hayata K, Iwahashi M, Ojima T, Katsuda M, Iida T, et al. Inhibition of IL-17A in tumor microenvironment augments cytotoxicity of tumor-infiltrating lymphocytes in tumor-bearing mice. PLoS One. 2013;8:e53131. doi: 10.1371/journal.pone.0053131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhang B, Rong G, Wei H, Zhang M, Bi J, et al. The prevalence of Th17 cells in patients with gastric cancer. Biochem Biophys Res Commun. 2008;374:533–537. doi: 10.1016/j.bbrc.2008.07.060. [DOI] [PubMed] [Google Scholar]
- 61.Derhovanessian E, Adams V, Hähnel K, Groeger A, Pandha H, et al. Pretreatment frequency of circulating IL-17+ CD4+ T-cells, but not Tregs, correlates with clinical response to whole-cell vaccination in prostate cancer patients. Int J Cancer. 2009;125:1372–1379. doi: 10.1002/ijc.24497. [DOI] [PubMed] [Google Scholar]
- 62.Zhou P, Sha H, Zhu J. The role of T-helper 17 (Th17) cells in patients with medulloblastoma. J Int Med Res. 2010;38:611–619. doi: 10.1177/147323001003800223. [DOI] [PubMed] [Google Scholar]
- 63.Wainwright DA, Sengupta S, Han Y, Ulasov IV, Lesniak MS. The presence of IL-17A and T helper 17 cells in experimental mouse brain tumors and human glioma. PLoS One. 2010;5:e15390. doi: 10.1371/journal.pone.0015390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kryczek I, Wei S, Vatan L, Escara-Wilke J, Szeliga W, et al. Cutting edge: opposite effects of IL-1 and IL-2 on the regulation of IL-17+ T cell pool IL-1 subverts IL-2-mediated suppression. J Immunol. 2007;179:1423–1426. doi: 10.4049/jimmunol.179.3.1423. [DOI] [PubMed] [Google Scholar]
- 65.Kryczek I, Wei S, Zou L, Altuwaijri S, Szeliga W, et al. Cutting edge: Th17 and regulatory T cell dynamics and the regulation by IL-2 in the tumor microenvironment. J Immunol. 2007;178:6730–6733. doi: 10.4049/jimmunol.178.11.6730. [DOI] [PubMed] [Google Scholar]
- 66.Cui X, Xu Z, Zhao Z, Sui D, Ren X, et al. Analysis of CD137L and IL-17 expression in tumor tissue as prognostic indicators for gliblastoma. Int J Biol Sci. 2013;9:134–141. doi: 10.7150/ijbs.4891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hu J, Mao Y, Li M, Lu Y. The profile of Th17 subset in glioma. Int Immunopharmacol. 2011;11:1173–1179. doi: 10.1016/j.intimp.2011.03.015. [DOI] [PubMed] [Google Scholar]
- 68.Cantini G, Pisati F, Mastropietro A, Frattini V, Iwakura Y, et al. A critical role for regulatory T cells in driving cytokine profiles of Th17 cells and their modulation of glioma microenvironment. Cancer Immunol Immunother. 2011;60:1739–1750. doi: 10.1007/s00262-011-1069-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chang SH, Dong C. Signaling of interleukin-17 family cytokines in immunity and inflammation. Cell Signal. 2011;23:1069–1075. doi: 10.1016/j.cellsig.2010.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Schwartzbaum JA, Huang K, Lawler S, Ding B, Yu J, et al. Allergy and inflammatory transcriptome is predominantly negatively correlated with CD133 expression in glioblastoma. Neuro Oncol. 2010;12:320–327. doi: 10.1093/neuonc/nop035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wang Q, Li H, Yao Y, Xia D, Zhou J. The overexpression of heparin-binding epidermal growth factor is responsible for Th17-induced airway remodeling in an experimental asthma model. J Immunol. 2010;185:834–841. doi: 10.4049/jimmunol.0901490. [DOI] [PubMed] [Google Scholar]
- 72.Honorati MC, Neri S, Cattini L, Facchini A. Interleukin-17, a regulator of angiogenic factor release by synovial fibroblasts. Osteoarthritis Cartilage. 2006;14:345–352. doi: 10.1016/j.joca.2005.10.004. [DOI] [PubMed] [Google Scholar]
- 73.Lee JW, Wang P, Kattah MG, Youssef S, Steinman L, et al. Differential regulation of chemokines by IL-17 in colonic epithelial cells. J Immunol. 2008;181:6536–6545. doi: 10.4049/jimmunol.181.9.6536. [DOI] [PubMed] [Google Scholar]