

Abstract
It is commonly accepted that summer cyanobacterial blooms cannot be efficiently utilized by grazers due to low nutritional quality and production of toxins; however the evidence for such effects in situ is often contradictory. Using field and experimental observations on Baltic copepods and bloom-forming diazotrophic filamentous cyanobacteria, we show that cyanobacteria may in fact support zooplankton production during summer. To highlight this side of zooplankton-cyanobacteria interactions, we conducted: (1) a field survey investigating linkages between cyanobacteria, reproduction and growth indices in the copepod Acartia tonsa; (2) an experiment testing relationships between ingestion of the cyanobacterium Nodularia spumigena (measured by molecular diet analysis) and organismal responses (oxidative balance, reproduction and development) in the copepod A. bifilosa; and (3) an analysis of long term (1999–2009) data testing relationships between cyanobacteria and growth indices in nauplii of the copepods, Acartia spp. and Eurytemora affinis, in a coastal area of the northern Baltic proper. In the field survey, N. spumigena had positive effects on copepod egg production and egg viability, effectively increasing their viable egg production. By contrast, Aphanizomenon sp. showed a negative relationship with egg viability yet no significant effect on the viable egg production. In the experiment, ingestion of N. spumigena mixed with green algae Brachiomonas submarina had significant positive effects on copepod oxidative balance, egg viability and development of early nauplial stages, whereas egg production was negatively affected. Finally, the long term data analysis identified cyanobacteria as a significant positive predictor for the nauplial growth in Acartia spp. and E. affinis. Taken together, these results suggest that bloom forming diazotrophic cyanobacteria contribute to feeding and reproduction of zooplankton during summer and create a favorable growth environment for the copepod nauplii.
Introduction
Toxic blooms of filamentous cyanobacteria are proliferating worldwide due to the climate change and eutrophication [1]. These cyanobacteria are commonly considered to impair survival, growth and reproduction of grazers [2]. The negative effects of cyanobacteria on zooplankton are usually related to a combination of (i) low nutritional value due to inadequate dietary fatty acid composition [3], [4], (ii) production of toxins and feeding deterrents [5], [6], and (iii) poor manageability of the colonies [7]. However, there are also studies showing that cyanobacteria have neutral or positive effects on zooplankton egg production and growth [8]–[12]. Application of stable isotopes, fatty acids and DNA-based methods [13]–[16] for zooplankton diet analysis suggest relatively high in situ grazing on colony-building cyanobacteria in freshwaters and estuaries, where summer cyanobacterial blooms are a regular feature. One can speculate that when preferred prey is scarce, zooplankton may increase feeding on the abundant cyanobacteria, despite their inadequate biochemical composition and toxicity. We know, for example, from terrestrial ecology that rainforest-dwelling parrots consume toxic foods during dry seasons when other, non-toxic, food is limited [17]. Supporting these contradictory reports, meta-analysis of cyanobacteria effects on various grazers [18], [19] suggest that cyanobacterial effects on biota are multifactorial and species- and system-specific [20].
In the Baltic Sea, summer blooms of diazotrophic filamentous cyanobacteria are dominated by Aphanizomenon sp., Nodularia spumigena and Dolichospermum spp. [21]. Biomass of Baltic mesozooplankton peaks during the same period as that of cyanobacteria, with calanoid copepods being the most important group [22], [23]. The evidence is accumulating that cyanobacterial blooms may have a more important role in the Baltic food webs than previously assumed, and it has been suggested that utilization of cyanobacteria as a food source by zooplankton in this system is underestimated [24]. For example, a strong nitrogen isotopic signal in of various size following summer bloom in the northern Baltic Sea [25], indicates that nitrogen (N) fixed by diazotrophic cyanobacteria is directly or indirectly utilized in the food web. Indeed, using both isotopic signals [26] and cyanobacterial pigments as tracers, copepods in the Baltic proper have been found to assimilate cyanobacteria in measurable quantities [24]. Also, lipid signal in the copepod Pseudocalanus acuspes in the southern Baltic Sea indicates that cyanobacteria contribute substantially to its diet during summer [27]. Finally, studies employing DNA-based analysis of stomach content provide unequivocal evidence that Baltic copepods and mysids ingest filamentous cyanobacteria in situ even when alternative food is present [14].
Similarly, effects of filamentous cyanobacteria on survival and development are often contradictory, with both negative [12], [28] and positive [9], [10] effects being observed. As a monospecific food, cyanobacteria do not seem to support neither egg production [11], [28], [29] nor egg hatching [28] in copepods, with nodularins and microcystins being the most commonly implicated in these effects. These hepatotoxins have been reported to cause oxidative stress in various invertebrates and fish [30], [31] by increasing formation of reactive oxygen species, decreasing the antioxidant capacity (e.g., inhibiting detoxification enzymes) and oxidation of macromolecules (proteins, lipids and DNA). However, when offered in mixtures with alternative prey, filamentous cyanobacteria may have positive effects on copepod egg production, hatching and juvenile development [9], [11], [32]. Due to this conflict of information, more studies on the occurrence and mechanisms of these effects in ecologically relevant settings are needed.
Here, we explored linkages between Baltic Sea diazotrophic filamentous cyanobacteria and fitness-related responses in copepods: reproductive output, juvenile development and growth, and oxidative status. To evaluate these connections, we conducted a set of interrelated studies in the northern Baltic proper. First, in a field survey, we used a correlative approach to relate reproduction (egg production and their viability) of the copepod Acartia tonsa to phytoplankton community structure, with specific focus on the effects of the bloom-forming cyanobacteria. Then, we conducted a follow-up laboratory experiment, where ingestion of the cyanobacterium was measured in A. bifilosa using molecular diet analysis and related to the copepod oxidative status, and recruitment. Finally, we analyzed long-term data on growth indices in nauplii of Acartia spp. and Eurytemora affinis in relation to cyanobacteria bloom intensity.
Methods
Ethics Statement
The sampling was conducted within Swedish National Marine Monitoring in the Baltic Sea and no specific permissions were required for the sampling locations of this study. Also, we did not require ethical approval to conduct this research as no animals considered in any animal welfare regulations and no endangered or protected species were involved in either field or experimental studies.
Summer field survey
Study sites
Sampling was conducted biweekly during cyanobacterial bloom (July 4 to September 26, 2007) at three stations in the north-west Baltic proper (58°49'N, 17°39'E; Figure 1). Two of these stations are located in the Himmerfjärden Bay (stns H2 and H4; SYVAB's marine monitoring program) and stn B1 (Swedish National Marine Monitoring Program, SNMMP) is outside the bay (Figure 1).
Figure 1. Schematic map over the Baltic Sea and the sampling sites.

(A) Himmerfjärden Bay (sampling stations H2 and H4) and a coastal area near Askö laboratory (station B1) in the western part of the northern Baltic proper, where field data for the summer field survey and the long term data (stn H4 and B1) were collected, and (B) Storfjärden Bay, at the entrance to the Gulf of Finland, where study animals for the laboratory experiment were collected.
Sampling
Sea surface temperature (SST) was measured on each sampling occasion. Integrated phytoplankton samples were collected by a hose (inner diameter 19 mm) at stns H2 and H4 (0–14 m) and stn B1 (0–20 m), and preserved with acidic Lugol's solution. Using an inverted microscope (Wild M40), cells counts (>2 µm) and biovolume analysis were conducted according to HELCOM monitoring guidelines [33], [34]; see also http://www.ices.dk/marine-data/Documents/ENV/PEG_BVOL.zip. Based on size measurements, phytoplankton were divided into size fractions of 2–5 µm, 5–15 µm, 15–30 µm and>30 µm (filamentous cyanobacteria excluded); each species of filamentous cyanobacteria was treated as a separate category regardless of size.
Copepods were sampled by vertical hauls (0–10 m) using a WP2 net (diameter 58 cm, mesh size 200 µm) and brought to the laboratory in 20 L insulated containers. Using a wide-mouth pipette, adult A. tonsa females were sorted upon arrival to the laboratory; the choice of species was based on relative abundance of copepod species in the samples and availability of adult females. They were incubated individually for 24 h in darkness at ambient (±2°C) temperature, in microplate wells (12 wells; Corning Costar, Corning NY, USA) filled with 5 mL of 6-µm filtered seawater. The mortality of the females during the incubation was <5%.
Reproductive variables
The egg production rate (EPR; eggs female−1 day−1) was recorded by counting number of eggs in each well under a stereomicroscope (Wild Heerbrugg, 6–50×). No crumbled or empty egg shells were found, indicating absence of egg cannibalism. Live females were individually placed in Eppendorf tubes, frozen at −80°C and stored for a few weeks before RNA analysis. For egg viability (EV%; percentage of viable eggs) analysis, all eggs from each well were transferred to a depression slide and stained with TO-PRO-1 iodide (Molecular Probes) [35]. The viable egg production rate (VEPR; viable eggs female−1 day−1) was calculated by multiplying EV% with EPR.
RNA quantification
As a proxy for growth rate, individual RNA content of the females was used [36] measured by the high-range RiboGreen (Molecular Probes, Inc., Eugene, OR, USA) assay after extraction with N-laurylsarcosine followed by RNase digestion [37]; fluorometer FLUOstar Optima (BMG Labtechnologies; 485 nm for excitation and 520 nm for emission) and black flat bottom microplates (Greiner Bio-One GmbH) were used. Before the analysis, the females (PL, prosome length mm) were measured using an inverted microscope (80×; Wild 40, Heerbrugg) with an ocular micrometer.
Laboratory experiment
Study animals and algal cultures
In August 2010, copepods were collected, with a 150 µm plankton net in the Storfjärden Bay, Western Finland (59°51'20''N, 23°15'42''E; Figure 1). Adult females and males of Acartia bifilosa which dominated the copepod community during the study period and area, were gently sorted under a stereo microscope and placed in 1.2 L bottles. Cultures of the green alga Brachiomonas submarina (strain TV15, collection of Tvärminne Zoological Station, University of Helsinki) and the cyanobacterium Nodularia spumigena (strain AV1 obtained from Prof. K. Sivonen, University of Helsinki) were grown under irradiance of 13.7 µmol photons m−2 s−1 for 16 h a day, and at 18°C, in f/2 medium (without silica) and Z8 (without nitrogen), respectively [38].
Experimental set-up
The copepods (17 females and 3 males per bottle, 12 replicates per treatment) were incubated in two alternative feeding regimes, with or without N. spumigena in the media, at 20°C. In the treatments without cyanobacteria, a monoculture of B. submarina (1061±87 µg C L−1; average ± SD) was used as a sole food, and in the treatments with cyanobacteria, B. submarina (971±208 µg C L−1) were mixed with N. spumigena (102±18 µg C L−1); this mixture approximates average N. spumigena contribution to the summer phytoplankton community in the northern Baltic proper (Figure 2). The bottles were incubated on a plankton wheel (1 rpm), with 16∶8 h light:dark cycle. Prior to the experimental incubation, the copepods were acclimatized under the same conditions for 36 h. At the end of the experiment, the copepods were filtered through a 120 µm mesh and examined under a stereo microscope. Live copepods were transferred to Eppendorf tubes and stored at −80°C for stomach content analysis using qPCR (quantitative real-time polymerase chain reaction) and oxidative status analysis. To get sufficient amount of material for these analyses, all material collected from consecutive replicates were pooled two and two within a treatment, giving 6 replicates for each treatment. Eggs and nauplii from each bottle were collected with a 48 µm mesh, stored overnight at 3°C in dark, and used to estimate EPR, EV% and development index (DI).
Figure 2. Contribution of the main taxonomic groups to the phytoplankton communities (by biovolume) at stations B1 (closed symbols) and H4 (open symbols) in June-August (mean ± SD; years 1986–2009).
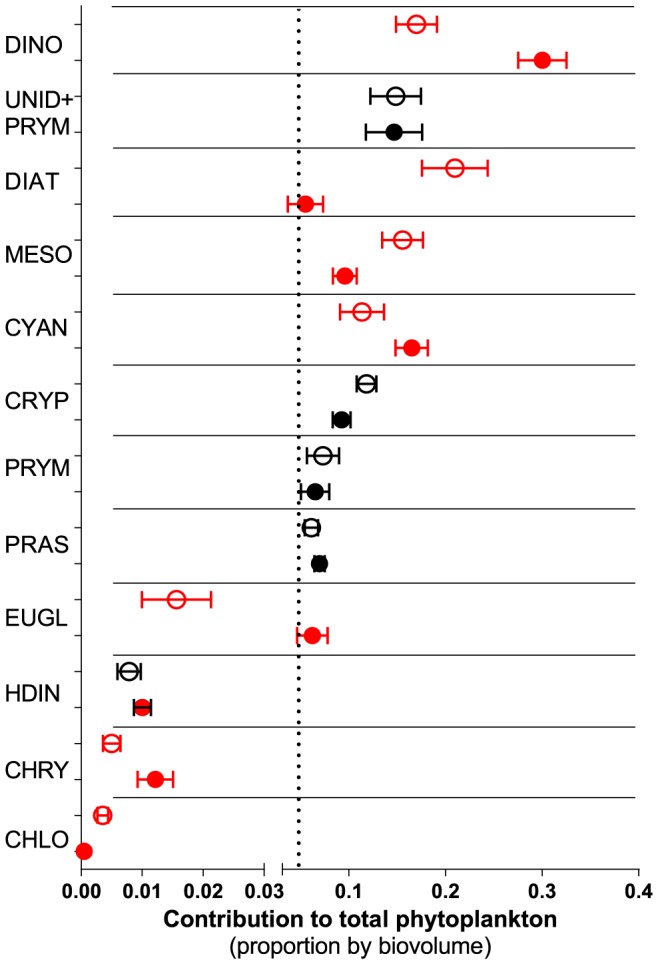
Significant differences between the stations (paired t-test, p<0.05) are indicated in red. Dotted line indicates 5% threshold for including a phytoplankton group in the GLM analysis of the field survey study.
Sample preparation for molecular and biochemical assays
To determine amount of N. spumigena in the copepod gut content and to characterize oxidative status of the copepods, adult animals recovered from the incubation bottles were placed into microcentrifuge tubes with 0.7 mL phosphate buffer saline (PBS) and 100 µm glass beads and homogenized for 4 minutes using FastPrep with cooling. The tubes were thereafter centrifuged at 4°C for 5 min with 10 000×g.
Molecular diet analysis
For DNA extraction, 50 µL of the homogenate were used by mixing with 50 µL 20% Chelex [39]. These samples were heated at 105°C for 30 min, centrifuged and the supernatant was used to quantify N. spumigena using qPCR. The Nodularia-specific primers were used to amplify ∼200 bp fragment of N. spumigena 16S rDNA in the guts of copepods [15]. All qPCR reactions were performed in a final volume of 25 µL reaction mixture with a StepOne real-time cycler (Applied Biosystems) using the KiCqStart SYBR Green qPCR Ready Mix (Sigma). To prepare a standard curve of five step 10-fold dilutions (8.2 to 8.2×10−4 ng), DNA extracted from a culture of N. spumigena was used [40]; the linearity of the standard curve was high (R2>0.99), with amplification efficiency of 95–100%. As a reference sample, the feeding media containing N. spumigena but no copepods following the same procedure as for the test samples was used. A standard curve and no template controls (water) in triplicates were included in all runs. To check for non-specific products, DNA melt-curve analysis was performed after each qPCR experiment.
Oxidative stress biomarkers
To measure the intracellular soluble antioxidant capacity, the homogenized copepod samples were analyzed using oxygen radical absorption capacity (ORAC; µM trolox equivalents) assay [41]; see Protocol S1. To measure oxidative damage, the lipid peroxidation assay using QuantiChrom thiobarbituric acid reactive substances (TBARS; mol MDA Assay Kit; DTBA-100; BioAssay Systems, USA) was used following the manufacturer's directions. The ratio between ORAC and TBARS (ORAC:TBARS ratio) was used as a proxy for oxidative balance [32].
Reproductive state variables
To estimate copepod reproductive output, the female EPR, the EV% and early nauplial development (development index; DI) were used. To determine EPR, all eggs were counted and related to the number of live females in the corresponding bottle. For EV% measurements, ∼50 eggs per bottle were analyzed with TO-PRO-1 iodide staining [35]. The remaining eggs and nauplii were preserved with acidic Lugol's solution for calculating DI, which incorporates survival and metamorphosis success in copepods:
where ki is assigned stage value (in our study: 0 for egg, 1 for nauplii NI, and 2 for nauplii NII; no nauplii had developed beyond NII), ni number of copepods at that stage, and NS – total number of individuals [42]. All nauplii from the incubations (i.e., Lugol-preserved samples for DI-analysis, and nauplii hatched in the TO-PRO-stained samples) were included in the EV calculation. VEPR was estimated by multiplying EV and EPR.
Long term data analysis
Zooplankton collected in the northern Baltic proper within SNMMP and SYVAB's marine monitoring program in Himmerfjärden Bay (Himmerfjärden Eutrophication Study; www.2.ecology.su.se) were used to study effects of various phytoplankton groups and physical factors on copepod growth and recruitment, with particular focus on the effects of cyanobacteria. As a proxy for copepod growth during early life stages, we used RNA:DNA ratio in the nauplii (feeding stages, NIII–NVI) of Acartia spp. (A. bifilosa, A. tonsa and A. longiremis) and Eurytemora affinis. In the study areas, A. bifilosa dominate in summer Acartia communities, contributing 62 to 94% on the long-term basis (E. Gorokhova, pers. obs.). Specifically, we considered that biotic (biovolumes of total phytoplankton, phytoplankton excluding filamentous cyanobacteria, and specific phytoplankton groups) and abiotic (North Atlantic Oscillation [NAO] indices, SST and salinity) variables integrate local environmental variability, and could define an adequate environmental framework for copepod growth.
Sampling
We used 34 and 36 samples for stations H4 and B1, respectively, collected every other week in July and August, i.e., when cyanobacteria are abundant in the study area, during 1999–2009, resulting in 3–4 samples per station and year. On each sampling occasion, zooplankton and phytoplankton were collected in concert; salinity and SST were measured by CTD (Meerestechnik Elektronik GmbH). Phytoplankton samples were collected as integrated hose samples, preserved and analyzed in the same way as in the summer field survey [33], [34]. Zooplankton samples were taken by vertical tows from near bottom to surface using a WP2 net (diameter 57 cm, 90 µm mesh size). From each tow, randomly selected zooplankton were preserved in bulk using RNAlater and stored for 12 to 24 months at −20°C until the nucleic acid analysis [43].
RNA:DNA ratio
RNA and DNA contents were measured in nauplii (NIII–NVI) of Acartia spp. and E. affinis. The younger nauplial stages (NI–NII) were considered non-feeding [44] and thus not included in the analysis. Individual specimens were picked from the RNAlater preserved samples, rinsed and transferred to Eppendorf tubes (5–7 ind. sample−1); the two copepod genera were treated separately. Nucleic acids were quantified with fluorometric high-range RiboGreen (Molecular Probes, Inc., Eugene, OR assay) using the same instrumentation as for RNA analysis in the summer field survey [37]. Mean standard curve slope ratio (mDNA/mRNA) was 1.87.
Data analyses
The environmental data and phytoplankton biovolume data used in all analyses are available from www.smhi.se (SHARK database) and www.2.ecology.su.se/dbhfj/index.htm, and data on copepod growth- and reproduction-related variables are provided as Supporting Information (Summer field survey: Table S1; Laboratory experiment: Table S2; and Long-term data analysis: Table S3).
To evaluate effects of feeding environment with particular focus on cyanobacteria on copepod reproduction, growth and oxidative status, Generalized Linear Models (GLMs) with normal error structure and either identity or log-link function were applied using STATISTICA 10 (StatSoft, Inc., 2010). For all GLMs, the response variables and biovolume data were Box-Cox transformed to approach normal distribution. Akaike Information Criterion (AIC) was used to optimize the number and combination of predictive variables included. To validate the models, the Wald statistic was used to check the significance of the regression coefficient for each parameter, a likelihood ratio test was used to evaluate the statistical significance of including or not including each parameter and model goodness of fit was checked using deviance and Pearson χ2 statistics. Residual plots for each model were assessed visually to exclude remaining unattributed structure indicative of a poor model fit.
Summer field survey
GLMs were used to examine relationships between the dependent variables (reproductive/growth variables: VEPR, EPR, EV% and RNA) and the explanatory variables (biovolumes of filamentous cyanobacteria species [Aphanizomenon sp., N. spumigena and Dolichospermum spp.], autotrophs in each size fraction [2–5 µm; 5–15 µm; 15–30 µm and>30 µm], total phytoplankton biovolume, salinity and SST]. Copepod PL was also included as independent variable for RNA and EPR [45].
Laboratory experiment
Based on the positive effects of N. spumigena observed in the summer field survey and recent reports on high levels of antioxidants in cyanobacteria [46] as well as stimulation of antioxidant defenses in grazers feeding on hepatotoxic cyanobacteria [47], GLMs were used to test whether copepod reproductive output (EPR, EV%, VEPR and DI), antioxidant levels (ORAC), lipid peroxidation (TBARS), and oxidative balance (ORAC:TBARS ratio) were positively affected by grazing on N. spumigena. The amount of Nodularia in copepod guts determined by qPCR was used as a measure of grazing.
Long term data analysis
GLMs were used to examine relationships between the nauplial RNA:DNA ratio and biovolume of each phytoplankton group and abiotic variables that have been reported to affect zooplankton in the Baltic Sea (NAO, SST and salinity). Wilcoxon signed rank test was used to determine the significance of differences in nauplial RNA:DNA ratio between the stations (B1 and H4) and between the months (July and August) within the stations, whereas paired t-test was used to evaluate these differences in phytoplankton. The models were constructed for each taxa (Acartia spp. and E. affinis) and station (B1 and H4), because of the significant differences in RNA:DNA ratio between the species and differences in phytoplankton community structure between the stations. Only those phytoplankton groups that contributed>5% to the total phytoplankton biovolume were considered (Figure 2); these were: (1) dinoflagellates, (2) pooled group of small (<10 µm) unidentified flagellates and Prymnesiophyceae species, (3) diatoms, (4) Mesodinium rubrum (autotrophic ciliate), (5) nitrogen fixing filamentous Cyanobacteria, (6) Cryptophyceae, (7) large (>10 µm) Prymnesiophyceae, and (8) Prasinophyceae. We also included biovolumes of total phytoplankton and phytoplankton without cyanobacteria. The monthly NAO index values were taken from the Climate Prediction Center, Washington, DC (ftp://ftp.cpc.ncep.noaa.gov/wd52dg/data/indices/nao_index.tim). A high, positive winter NAO index (wNAO; December – March) indicates mild and rainy winters, while low, negative values occur during cold winters over Europe. A warmer winter is generally followed by a warmer spring (sNAO; March – May) with early onset of the growth season.
Results
Summer field survey
Phytoplankton communities differed among the stations, with the highest total phytoplankton (1.43 mm3 L−1) and highest contribution of filamentous cyanobacteria (29%) observed at stns H4 and H2, respectively (Figure 3). All cyanobacterial communities dominated by Aphanizomenon sp. (62–85% of total cyanobacteria; Figure 3). There were significant, albeit weak, negative correlations between the cyanobacteria and the two largest size classes of phytoplankton, 15–30 µm and>30 µm (Pearson r: −0.29 and −0.36 respectively; p<0.05), and a significantly positive correlation between the cyanobacteria and the smallest phytoplankton size class (2–5 µm; Pearson r: 0.38; p<0.05), whereas no correlation was found for the intermediate size class (5–15 µm; Pearson r: 0.13; p>0.05). Among the cyanobacteria, Aphanizomenon sp. had the highest significant negative correlation with phytoplankton>15 µm (Pearson r = −0.38; p<0.05) and N. spumigena had a moderate positive correlation with phytoplankton 2–5 µm (Pearson r = 0.53; p<0.05).
Figure 3. Composition of phytoplankton (A–C) and cyanobacterial (D–F) communities during the study period at: (A, D) stn B1, (B, E) stn H2, and (C, F) stn H4.

Number in the middle of each pie chart indicates total biovolume (mm3 L−1) of the contributing categories.
The maximum values for EPR and individual RNA content were observed in the first half of July, whereas their minima occurred in the end of August. The opposite trend was observed for EV%, with generally low values in July and high in the end of August (Table S4, Figure 4). The GLMs indicated that N. spumigena was a significant positive predictors for all reproductive variables (EPR, EV% and VEPR) but not the RNA content (Table 1). Aphanizomenon sp., on the other hand, significantly affected only EPR (stns B1 and H4) and the effect was negative. No statistically significant model for EPR was found for stn H2, which had significantly lower EPR compared to stns B1 and H4 (Table 1). Significant positive relationships were observed between phytoplankton 15–30 µm and all response variables. Also, phytoplankton>30 µm was significantly positively related to VEPR and RNA, albeit the relationships were relatively weak (Table 1).
Figure 4. Seasonal dynamics of reproductive and growth variables in the copepod Acartia tonsa in relation to cyanobacteria bloom progression (biovolume, mm3 L−1) at stations B1 (A), H2 (B) and H4 (C).

Egg production rate (EPR; eggs female−1 day−1), egg viability (EV%; % viable eggs), percentage of females producing eggs during 24-h incubation (%EP) and individual RNA content (µg ind−1).
Table 1. GLM results of the summer field study.
| Response variable | Stn | Explanatory variables | Beta | p | R 2 | Adj. R 2 | SE estimate |
| EPR | B1, H4* | Nodularia | 124.5 | <0.0001 | 0.39 | 0.35 | 3.29 |
| Aphanizomenon | −18.6 | <0.0001 | |||||
| 2–5 µm | 42.1 | 0.001 | |||||
| 15–30 µm | 7.6 | 0.0001 | |||||
| EV% | All | Nodularia | 7.0 | 0.006 | 0.28 | 0.19 | 0.21 |
| 0.008 | |||||||
| VEPR | All | Nodularia | 7.6 | <0.0001 | 0.39 | 0.35 | 3.29 |
| 15–30 µm | 6.4 | 0.034 | |||||
| >30 µm | 1.3 | <0.0001 | |||||
| 0.004 | |||||||
| RNA | All | 15–30 µm | 2.1 | <0.0001 | 0.64 | 0.62 | 0.04 |
| >30 µm | 0.6 | <0.0001 | |||||
| PL | 0.1 | <0.0001 | |||||
| 0.001 |
Response variables: egg production rate (EPR; eggs female−1 day−1), egg viability (EV%; % viable eggs), viable egg production rate (VEPR; viable eggs female−1 day−1), individual RNA content (µg ind−1). Explanatory variables: biovolume of different size fractions of phytoplankton and cyanobacteria species, copepod prosome length (PL) and station (B1: n = 28; H2: n = 29; and H4: n = 33). When a significant station effect was found, separate models were fitted for stations without significant differences. In EV% analysis, only females that produced eggs during incubation were included (n = 58).
*No significant model was found for stn H2.
Laboratory experiment
In copepods, incubated in the presence of N. spumigena in the feeding media, amount of the cyanobacterium per stomach estimated by qPCR varied ∼3-fold (0.47 to 1.32 ng Nodularia DW ind.−1); none of the controls were positive (Table S4). The amount of N. spumigena per copepod had significant positive effects on all response variables tested, except EPR where it was negative (Table 2). The concurrent opposite effects on EPR and EV% resulted in no significant influence on VEPR (Table 2). Also, the significant positive effect of N. spumigena on ORAC in combination with negative but not significant effect on TBARS resulted in the significant positive effect on the ORAC:TBARS ratio (Table 2).
Table 2. GLM results of the experimental data.
| Response variable | Estimate | Wald stat | p |
| EPR | −0.686 | 13.73 | 0.0002 |
| EV% | 1.560 | 10.643 | 0.0011 |
| VEPR | −0.293 | 1.783 | 0.1817 |
| DI | 1.687 | 4.279 | 0.0385 |
| ORAC | 0.004 | 7 | 0.0101 |
| TBARS | −0.814 | 2.391 | 0.1220 |
| ORAC:TBARS | 0.00003 | 4.299 | 0.0381 |
All models tested effects of grazing on Nodularia spumigena measured as amount of the cyanobacterium in the copepod guts (ng Nodularia DW ind−1) on reproductive output (EPR, EV%, and VEPR), nauplial development (DI), antioxidant (ORAC) and lipid peroxidation (TBARS) levels as well as the resulting oxidative balance (ORAC:TBARS ratio). Abbreviations and units for all response variables as in Tables 1 and 2. Significant models are in bold face.
Long term data
Significant differences were found between the stations in terms of the phytoplankton community structure over the years (Figures 2 and 5). Cyanobacteria contribution to the total phytoplankton biovolume during the bloom period (July-August) varied from less than 1%, with the lowest values observed in 2009 at both stations to 40% at B1 and 45% at H4, in 2007 and 2003, respectively. Other important phytoplankton groups were diatoms (up to 39% of the total biovolume), dinoflagellates (up to 47%) and prymnesiophyceans (up to 40%). Total phytoplankton biovolume also varied both over time and between the stations, reaching its peak in 2008–2009 and 2007–2008 at B1 and H4, respectively (Figure 5), with significantly higher values at stn H4 compared to stn B1 (Wilcoxon signed rank; p<0.0017, n = 33). Moreover, the nauplial RNA:DNA ratio was also significantly higher at stn H4 than at stn B1 (Wilcoxon signed rank; Acartia spp.: p<0.004; E. affinis: p<0.002). Therefore, to describe responses of nauplial RNA:DNA ratio (Figure 6) to variations in phytoplankton groups over time (Figure 5), the GLMs were fit for each station separately.
Figure 5. Long-term (1999–2009) dynamics of phytoplankton community structure at stations B1 (A) and H4 (B).
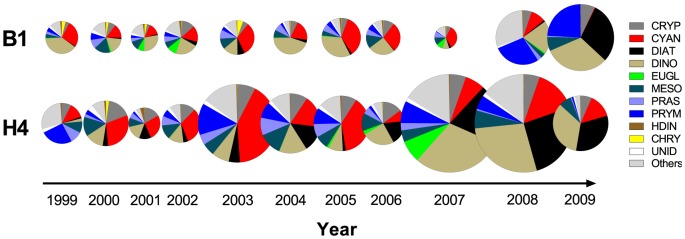
Data are averages for July and August, weeks 27 to 35 (e.g., period of the summer cyanobacterial bloom, n = 3 or 4). Circle diameter is proportional to the average total phytoplankton biovolume (mm3 L−1) observed during the same period (0.43±0.15, n = 70). Abbreviations: CHLO – Chlorophyceae, CHRY – Chrysophyceae, CRYP – Cryptophyceae, CYAN – Cyanophyceae, DIAT – Diatoms, DINO – Dinophyceae, EUGL – Euglenophyceae, MESO – Mesodinium rubrum (Myrionecta rubrum), PRAS – Prasinophyceae, PRYM – Prymnesiophyceae, UNID – unidentified flagellates, and HDIN – heterotrophic dinoflagellates.
Figure 6. Long-term (1999–2009) dynamics of RNA:DNA ratio (mean ± SD, n = 3 or 4) in the nauplii (NIII-NVI) of copepods Acartia spp. and Eurytemora affinis in relation to cyanobacteria biovolume at stations B1 (A) and H4 (B).
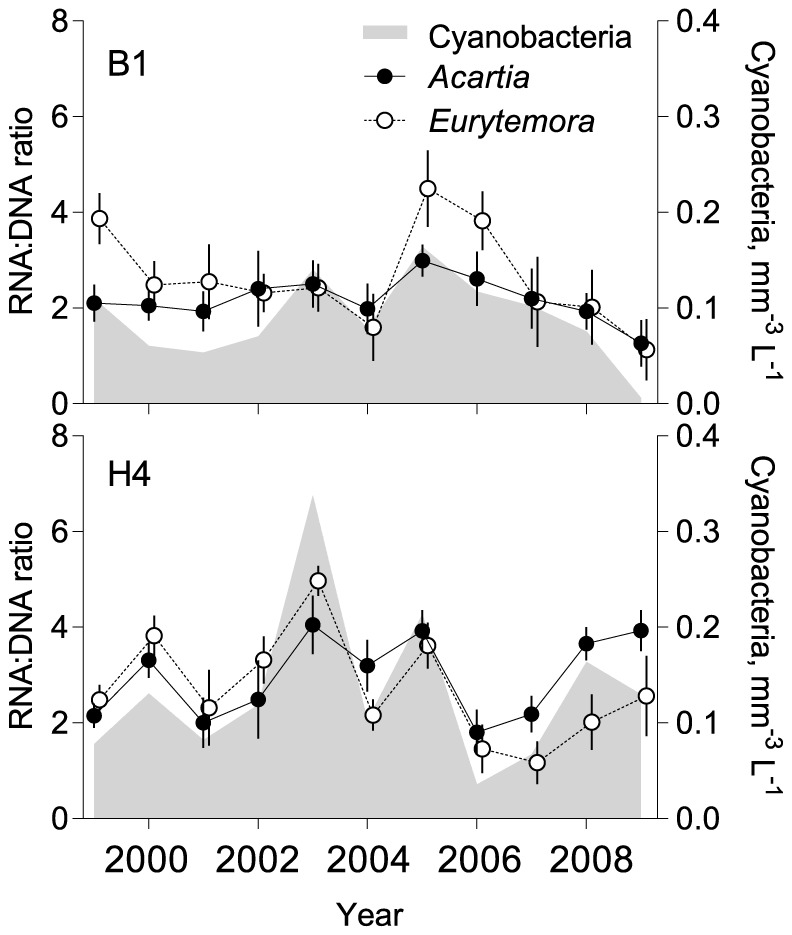
Data are averages for July and August, weeks 27 to 35.
For Acartia spp., only models based on filamentous cyanobacteria, diatoms, cryptophyceans, and abiotic parameters (SST, salinity, sNAO and wNAO) were significantly predictive. For E. affinis, only models incorporating total phytoplankton, filamentous cyanobacteria, diatoms and SST were significant (Table 3). Thus, in both species, the RNA:DNA ratio was significantly positively associated with the amount of cyanobacteria (Figure 6); moreover, parameter estimates indicated that these effects were the strongest in all models that included cyanobacteria (Table 3). Effects of other phytoplankton groups (cryptophyceans, total phytoplankton and diatoms) were all negative (Table 3). At stn B1, these groups correlated significantly negatively with cyanobacteria (Pearson r; −0.56, −0.44 and −0.62 for cryptophyceans, total phytoplankton and diatoms, respectively; Table S5). At stn H4, the effects of diatoms were also negative, but no significant correlations between the cyanobacteria and other phytoplankton groups were observed (Table S5).
Table 3. Best-fit GLMs relating RNA:DNA ratio in the copepod nauplii (Acartia spp. and E. affinis) to biovolumes of total phytoplankton (TotPhyto) and specific phytoplankton groups as well as climatic factors (SST, salinity and NAO indices) in the long-term dataset for stations B1 and H4.
| Station (n) | Phytoplankton group tested | Explanatory variables | Estimate | Wald statistics | p |
| Acartia spp. | |||||
| B1 | Cyanobacteria | Cyanobacteria | 3.788 | 170.892 | <0.0001 |
| (36) | SST | 0.045 | 21.118 | <0.0001 | |
| Cryptophyceans | Cryptophyceans | −7.002 | 18.845 | <0.0001 | |
| SST | 0.149 | 36.445 | <0.0001 | ||
| H4 | Cyanobacteria | Cyanobacteria | 1.583 | 9.736 | 0.0018 |
| (34) | wNAO | 0.566 | 17.104 | <0.0001 | |
| sNAO | −0.304 | 15.284 | <0.0001 | ||
| Salinity | 1.240 | 13.593 | 0.0003 | ||
| Diatoms | Diatoms | −0.820 | 5.838 | 0.0157 | |
| wNAO | 0.603 | 18.495 | <0.0001 | ||
| sNAO | −0.376 | 22.139 | <0.0001 | ||
| Salinity | 1.189 | 11.680 | 0.0007 | ||
| Eurytemora affinis | |||||
| B1 | TotPhyto | TotPhyto | −1.326 | 9.077 | 0.003 |
| (36) | sNAO | −0.346 | 26.641 | <0.0001 | |
| Cyanobacteria | Cyanobacteria | 5.164 | 22.059 | <0.0001 | |
| sNAO | −0.142 | 5.718 | 0.0282 | ||
| Diatoms | Diatoms | −3.248 | 4.796 | 0.0295 | |
| sNAO | −0.152 | 13.031 | 0.0012 | ||
| H4 | Cyanobacteria | Cyanobacteria | 2.611 | 47.899 | <0.0001 |
| (34) | wNAO | 0.327 | 10.008 | 0.0016 | |
| Salinity | −0.778 | 6.870 | 0.0087 | ||
| Diatoms | Diatoms | −1.646 | 18.088 | <0.0001 | |
| sNAO | −0.400 | 24.733 | <0.0001 | ||
| wNAO | 0.745 | 32.529 | <0.0001 | ||
| Salinity | −1.384 | 16.06 | <0.0001 | ||
sNAO and wNAO are NAO indices averaged for spring (March – May) and winter (December – March), respectively; n – number of observations.
Among the abiotic parameters, wNAO and sNAO indices were repeatedly indicated as significant, with positive effects of wNAO and negative of sNAO (Table 3). In addition, salinity had positive effects on RNA:DNA ratio in Acartia spp. and negative in E. affinis, whereas SST effect was significant only for Acartia spp. at B1 (Table 3).
Discussion
Both positive and negative effects of Baltic filamentous cyanobacteria on copepod biochemical and physiological responses were observed in the field and laboratory settings (Table S6). Moreover, these effects were species-specific and differed, in some cases, among the studies. However, contrary to the widely reported harmful effects of these cyanobacteria on the copepod reproduction [2], no negative effects on the net reproductive output, i.e. viable egg production, were found. In fact, viable egg production in A. tonsa was positively related to N. spumigena abundance in the field (Table 1). Although no effect of N. spumigena on viable egg production was observed in the laboratory experiment with A. bifilosa, the development of nauplii (non-feeding stages) was significantly advanced if their mothers were feeding on the cyanobacterium, thus effectively increasing recruitment (Table 2). Also, the long term data analysis also identified diazotrophic filamentous cyanobacteria as significant positive growth predictors for the copepod nauplii (Table 3).
Specific mechanisms behind the observed stimulating and suppressing effects of diazotrophic filamentous cyanobacteria on copepod reproduction and early development are largely unknown. We suggest three, not mutually exclusive, pathways involved in stimulating effects of cyanobacteria on copepod reproduction: (1) direct increase of macro- and micronutrient intake by copepods that use cyanobacteria as an additional food source; (2) supplementing copepod's diet with phytochemicals, such as polyphenols, vitamins and various antioxidants, that may enhance physiological responses [46], including hormesis effect, which is a generally favorable biological response to low exposures of toxins and other stressors [47]; and (3) indirect increase of nutrient intake by stimulation of the microbial loop through cyanobacterial exudates, resulting in increase of bacterio- and nanoplankton prey for smaller copepods [48]. The qPCR-based gut content analysis in the laboratory experiment confirmed that the copepods were actively feeding on N. spumigena (Table S4), and thus, the observed variations in the reproductive variables are – at least partly – the result of the direct grazing. Some filamentous cyanobacteria could be nutritionally valuable for grazers, particularly in combination with other food sources [49]. Moreover, as pointed out by Jiang and colleagues [50], “a putatively harmful alga is not always deleterious to grazers, and its ecological effects may be distinctly different during bloom and non-bloom periods”. They found that toxicity and nutritional value of the dinoflagellate Cochlodinium polykrikoides went from deleterious to beneficial for A. tonsa when the algal density decreased. Cyanobacteria may contain complimentary nutrients and microelements, e.g. amino acids, antioxidants, vitamins, proteins, phosphorus and nitrogen [3], [49], [51], [52] that are of particular value for copepods. Nitrogen, for instance, is frequently depleted in the pelagia during summer in the Baltic Sea [53] and can thus be limiting for copepod reproduction [54]. Therefore, feeding on nitrogen fixing cyanobacteria may relax this limitation.
In addition to the positive effects on egg viability and nauplial development, the presence of N. spumigena increased the antioxidant defenses and improved oxidative status as indicated by the ORAC:TBARS ratio (Table 2). This is in contrast to other studies implicating hepatotoxic cyanobacteria as pro-oxidants for grazers [31]. However, the nature of cyanobacteria-induced oxidative stress is poorly understood; in fact, cyanobacteria produce dietary antioxidants that may directly improve antioxidative capacity of consumers [52]. Moreover, microcystins have been shown to increase activity of antioxidant enzymes in estuarine crabs [55]. The observed increase in antioxidant capacity may be indicative of the hormetic response to the cyanobacterial toxins and bioactive compounds, known as The Xenohormesis Hypothesis [56]. The latter has been suggested to be an evolutionary adaptation to sustain fitness in a changing environment, where the presence of a toxin in low concentrations acts as a signal for the organism to mobilize metabolic reserves to prepare itself for higher environmental stress [47], [56], [57]. Some of phytochemicals have evolved as toxins to intimidate grazers, triggering adaptive stress responses by, for instance, stimulating the production and/or activity of antioxidant enzymes in the consumer [56], [58]. Nodularins and microcystins produced by N. spumigena and Dolichospermum spp., respectively, could have contributed to the observed positive linkages between these cyanobacteria and copepod egg viability. One can speculate that an increase in the antioxidant capacity of a female triggered by a hepatotoxin and/or another secondary metabolite could enhance allocation of maternal antioxidants to the eggs, which improved embryonic and post-embryonic development.
The positive linkages between nauplial growth inferred from the RNA:DNA ratio and total diazotrophic filamentous cyanobacteria in the long term data set (Table 3) are not likely to be due to the grazing on cyanobacteria by the nauplii. Although some grazing on N. spumigena colonies by copepod nauplii has been observed [59], the pathway involving fueling of microbial communities is much more likely. During summer, when inorganic nitrogen is depleted, bloom-forming cyanobacteria are responsible for a significant proportion of N2-fixation in the Baltic Sea [60]–[63]. As much as one third of the fixed N2 leaks out as ammonium NH4 + from these cyanobacteria [62], [63] and is further utilized by other organisms, such as heterotrophic bacteria and picoautotrophs [64]. The microbial loop is also stimulated by dissolved organic matter (DOM) and detritus derived from cyanobacteria and utilized by bacteria [64], [65]. Nauplii and, to a lesser extent, copepodites, graze on pico- and bacterioplankton [48], [65], [66], which also support dietary nano- and microzooplankton, and these communities flourish in association with N. spumigena [67]. Therefore, N-leakage from diazotrophic cyanobacteria, which stimulate microbial prey, could explain the positive association between nauplial growth and filamentous cyanobacteria during summer blooms (Table 3). Remarkably, cyanobacteria were the only phytoplankton group positively related to the nauplial RNA:DNA ratio (Table 3, Figure 6). Although cryptophyceans, total phytoplankton and diatoms were all negatively associated with nauplial growth at stn B1 (Table 3), these negative effects may, at least partially, be explained by the negative cross-correlations between these groups and the cyanobacteria (Table S5); see also [68]. However, the negative effect of the diatoms on the nauplial growth at stn H4 cannot be explained by such correlation (Table S5). In this case, diatoms may have affected nauplii either directly, e.g., via deleterious effects [69], or by interspecific interaction with some other prey that was beneficial for growth. The observed effects of salinity (Table 3) are rather expected as A. bifilosa is a species of marine origin, whereas E. affinis is a brackish water copepod thriving in a broad salinity range [22]. Moreover, high genetic diversity of Baltic E. affinis [70] may have contributed to the varying responses to winter and spring NAO (Table 3).
While stimulation of the microbial food web is possible in the field, it cannot explain the positive effects of N. spumigena on copepod egg viability and non-feeding nauplial development in the experiment (Table 2), because the copepods were incubated in 0.2 µm filtered sea water, where bacteria were heavily reduced and flagellates and ciliates were largely eliminated. Hence, both the direct and indirect pathways are likely to be responsible for the positive relationships between N. spumigena and copepod reproduction observed in the summer field survey (Table 1). The microbial communities thriving in cyanobacterial bloom may nourish the copepods either via the microbial loop [65], [71] or as epibions on the ingested cyanobacterial colonies. Moreover, the mechanisms may differ between the cyanobacteria species as well as among zooplankton grazers.
Although the net effects of cyanobacteria on copepod reproduction (i.e., VEPR) appear either neutral or positive, there is some variation among the species and studies. For example, in the summer field survey, effects of N. spumigena on egg production in A. tonsa were positive (Table 1), whereas in the experimental study, this effect in A. bifilosa was negative, albeit these opposite responses resulted in no significant net effect on viable egg production (Table 2). This discrepancy between the laboratory experiment and the summer field survey suggest that variations in the feeding environment may be crucial for cyanobacteria-copepod interactions. In the field, N. spumigena had a significant positive correlation with biovolume of edible phytoplankton (15–30 µm), which was also a positive predictor for egg production; whereas these were not available for the copepods in the experiment. There might also be differences between the test species, A. tonsa (field survey) and A. bifilosa (experiment), in their sensitivity to the cyanobacterium effects and dependency on other food sources and environmental factors. In particular, A. bifilosa might have been avoiding to ingest larger filaments in the experiment due to food selection against this cyanobacterium [72]. This would effectively decrease food availability in the incubations as food (Brachiomonas with or without Nodularia) was provided at the same amount. As a result, lower food intake when feeding in mixtures containing cyanobacteria would translate into lower egg production, which was observed in this copepod species.
The observed differences among the cyanobacteria species could also be related to their relative abundance, morphology, nutritional value and/or biochemistry of toxins and metabolites. N. spumigena colonies do not form bundles, are not as rigid [73], and could, therefore, be easier to handle for mesozooplankton grazers than Aphanizomenon sp., which was found to negatively affect EPR (Table 1), and which has been reported to impair copepod reproduction to a higher extent than N. spumigena [29]. These cyanobacteria species also differ in their abundance (Figure 3) and nutritional value [49], [51], including the value of their microbial epibionts [67], [74]. Aphanizomenon sp. has lower colonization by heterotrophic bacteria compared to N. spumigena [59], [63], which hosts a lucrative microenvironment for microorganisms [62], [67], [75]. This was also supported by significant positive correlation between N. spumigena and phytoplankton 2–5 µm observed in the summer field survey. This size fraction might be more N-sufficient than larger phytoplankton and thereby provide the more balanced food for the copepods. In line with this, significant positive effects of the phytoplankton 2–5 µm on the copepod reproductive variables were observed (Table 1). The highest egg production has been reported to occur when the diet consisted of alternative food with small contribution of cyanobacteria, mainly N. spumigena [29]; this resembles the in situ feeding conditions in our field study. The negative correlation between Aphanizomenon sp. and EPR (Table 2) could also be a result of poor feeding conditions for the copepods, due to scarcity of edible phytoplankton during the cyanobacteria bloom. Indeed in the field survey, Aphanizomenon sp., among the filamentous cyanobacteria, had the highest significant negative correlation with phytoplankton>15 µm. For A. tonsa, the optimal food size is 2–5% of the prosome length [76], which implies that optimal food size for the females sampled here would be 15–40 µm, explaining the observed positive relationships between phytoplankton 15–30 µm and growth related variables (RNA content and EPR; Table 1). Moreover, station had a significant influence on egg production, indicating presence of other factors, not accounted for in our analysis (Table 1).
In conclusion, our results indicate that summer blooms of diazotrophic filamentous cyanobacteria in the Baltic Sea could be important for copepod growth and reproduction by providing complementary food, supporting high antioxidant levels and fueling growth of microbial prey. In particular, N. spumigena, which are the most conspicuous bloom forming toxic filamentous cyanobacteria that build surface accumulations [20], is grazed upon and have positive effects on copepod recruitment and growth (Table S6). This was however not the case for the most abundant cyanobacterium, Aphanizomenon sp., which appears to decrease total egg production, with, however, neutral effects on the production of viable eggs. Given that dominant Baltic zooplankters are selectively feeding on non-toxic cyanobacteria or can avoid cyanobacterial filaments altogether [72], it is possible that by supporting growth and recruitment in grazer populations, these cyanobacteria may gain a competitive advantage over other phytoplankton [77]. In this case, the effects of cyanobacteria on grazers will be highly system-specific, depending on evolutionary trajectories of the species and populations in question. Our results, together with findings reporting high incorporation of diazotrophic nitrogen in pelagic and benthic food webs in this and other systems experiencing blooms of these cyanobacteria [78], [79], have important implications for understanding impacts of these blooms on secondary, and, ultimately, fish production in the Baltic Sea. Further investigations on the associated ecological and evolutionary trade-offs behind these interactions are needed, if we are to understand and manage eutrophication and fishery in this system.
Supporting Information
Acartia tonsa : Data on reproductive and growth variables obtained in the summer field survey conducted in July - September 2007, at three coastal stations (B1, H2 and H4) in the northern Baltic proper. Variables measured: EPR (egg production rate), VEPR (viable egg production rate), RNA (individual ribonucleic acid content), and PL (prosome length).
(XLSX)
Acartia bifilosa : Data on feeding, reproduction and nauplii development obtained in the laboratory experiment conducted in August 2010, in the Storfjärden Bay, Western Finland. Variables measured: EPR (egg production rate), VEPR (viable egg production rate), RNA (individual ribonucleic acid content), and PL (prosome length).
(XLSX)
Acartia sp. and Eurytemora affinis : Long-term data (mean ± SD; period 1999–2009) on RNA:DNA ratio in nauplii at stns B1 and H4 in the northern Baltic proper. The mean values are based on 3–4 replicate samples analyzed per year (July-August).
(XLSX)
Growth and reproduction indices observed in the (A) field survey, (B) experimental study, and (C) long-term dataset. Data are presented as means with their standard deviations; abbreviations as in Tables S1 and S2.
(DOC)
Pearson r for correlations among phytoplankton groups at stn B1 and stn H4. Significant correlations are in bold face (p<0.05). Abbreviations: CHLO – Chlorophyceae, CHRY – Chrysophyceae, CRYP – Cryptophyceae, CYAN – Cyanophyceae, DIAT – Diatomea, DINO – Dinophyceae, EUGL – Euglenophyceae, MESO – Mesodinium rubrum (Myrionecta rubrum), PRAS – Prasinophyceae, PRYM – Prymnesiophyceae, UNID – unidentified flagellates, and HDIN – heterotrophic dinoflagellates.
(DOC)
Summary of the observed effects linking filamentous cyanobacteria to reproductive output, growth indices and oxidative status in copepods. Abbreviations as in Tables S1 and S2.
(DOC)
Oxidative stress biomarkers.
(DOCX)
Acknowledgments
We thank Askö Laboratory (Sweden) and Tvärminne Zoological Station (Finland) for permission to use facilities and for technical assistance in the field and laboratory studies. We also thank Dr. Ulf Larsson (Stockholm University, Sweden) for providing scientific advice on data interpretation, Leif Lundgren, Bengt Abrahamsson, Stefan Svensson, Lisa Mattsson, Carl Mattsson and Marika Tirén (Stockholm University, Sweden) for collecting samples within Swedish National Monitoring Programme and assistance in the laboratory, and Prof. K. Sivonen (University of Helsinki, Finland) for providing Nodularia spumigena (AV1) strain for our experiments.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. The environmental data and phytoplankton biovolume data used in all analyses are available from www.smhi.se (SHARK database) and www.2.ecology.su.se/dbhfj/index.htm, and data on copepod growth- and reproduction-related variables are provided as Supporting Information (Summer field survey: Table S1; Laboratory experiment: Table S2; and Long-term data analysis: Table S3).
Funding Statement
This work was financially supported by Stockholm University's strategic marine environmental research program Baltic Ecosystem Adaptive Management, the Swedish Environmental Protection Agency, SYVAB (The Southwestern Stockholm Region Sewage Company), Ivar Bendixsons Stipendiefond and the Swedish Research Council for the Environment, Agricultural Sciences and Spatial Planning (FORMAS), the Academy of Finland (projects 125251 and 255566), the Maj and Tor Nessling Foundation, Walter and Andrée de Nottbeck Foundation, and the Research and Development Institute ARONIA. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Paerl HW, Fulton RS (2006) Ecology of harmful cyanobacteria, In: Granéli E, Turner J (Eds.) Ecology of harmful marine algae. Springer-Verlag, Berlin, pp. 95–107. [Google Scholar]
- 2. Wiegand C, Pflugmacher S (2005) Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicol Appl Pharm 203: 201–218. [DOI] [PubMed] [Google Scholar]
- 5. DeMott WR, Moxter F (1991) Foraging on cyanobacteria by copepods – responses to chemical defenses and resource abundance. Ecology 72: 1820–1834. [Google Scholar]
- 6. Carmichael WW (1992) Cyanobacteria secondary metabolites – the cyanotoxins. J Appl Bacteriol 72: 445–459. [DOI] [PubMed] [Google Scholar]
- 7. Webster EK, Peters RH (1978) Some size-dependent inhibitions of larger cladoceran filterers in filamentous suspensions. Limnol Oceanogr 23: 1238–1245. [Google Scholar]
- 8. DeMott WR (1998) Utilization of a cyanobacterium and a phosphorus-deficient green alga as complementary resources by daphnids. Ecology 79: 2463–2481. [Google Scholar]
- 9. Koski M, Schmidt K, Engström-Öst J, Viitasalo M, Jónasdóttir S, et al. (2002) Calanoid copepods feed and produce eggs in the presence of toxic cyanobacteria Nodularia spumigena . Limnol Oceanogr 47: 878–885. [Google Scholar]
- 10. Schmidt K, Koski M, Engström-Öst J, Atkinson A (2002) Development of Baltic Sea zooplankton in the presence of a toxic cyanobacterium: a mesocosm approach. J Plankton Res 24: 979–992. [Google Scholar]
- 11. Kozlowsky-Suzuki B, Karjalainen M, Lehtiniemi M, Engström-Öst J, Koski M, et al. (2003) Feeding, reproduction and toxin accumulation by the copepods Acartia bifilosa and Eurytemora affinis in the presence of the toxic cyanobacterium Nodularia spumigena . Mar Ecol Prog Ser 249: 237–249. [Google Scholar]
- 12. Schmidt K, Jónasdóttir SH (1997) Nutritional quality of two cyanobacteria: How rich is 'poor' food? Mar Ecol Prog Ser 151: 1–10. [Google Scholar]
- 13. Oberholster PJ, Botha AM, Cloete TE (2006) Use of molecular markers as indicators for winter zooplankton grazing on toxic bentic cyanobacteria colonies in an urban Colorado lake. Harmful Algae 5: 705–716. [Google Scholar]
- 14. Gorokhova E (2009) Toxic cyanobacteria Nodularia spumigena in the diet of Baltic mysids: Evidence from molecular diet analysis. Harmful Algae 8: 264–272. [Google Scholar]
- 15. Gorokhova E, Engström-Öst J (2009) Toxin concentration in Nodularia spumigena is modulated by mesozooplankton grazers. J Plankton Res 31: 1235–1247. [Google Scholar]
- 16. Davis TW, Gobler CJ (2011) Grazing by mesozooplankton and microzooplankton on toxic and non-toxic strains of Microcystis in the Transquaking River, a tributary of Chesapeake Bay. J Plankton Res 33: 415–430. [Google Scholar]
- 17. Gilardi JD, Toft CA (2012) Parrots eat nutritious foods despite toxins. PLoS One 7: e38293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Wilson AE, Sarnelle O, Tillmanns AR (2006) Effects of cyanobacterial toxicity and morphology on the population growth of freshwater zooplankton: Meta-analyses of laboratory experiments. Limnol Oceanogr 51: 1915–1924. [Google Scholar]
- 19.Ibelings BW, Havens KE (2008) Cyanobacterial toxins: a qualitative meta-analysis of concentrations, dosage and biota, Chapter 32, In: Hudnell HK (Ed.) Cyanobacterial harmful algal blooms: State of the science and research needs, pp. 675–732. [DOI] [PubMed]
- 20.El-Shehawy R, Gorokhova E (2013) The bloom-forming cyanobacterium Nodularia spumigena: a peculiar nitrogen-fixer in the Baltic Sea food webs, In: Cyanobacteria: Ecology, Toxicology and Management. Nova publisher, USA. [Google Scholar]
- 21. Wasmund N (1997) Occurrence of cyanobacterial blooms in the Baltic sea in relation to environmental conditions. Int Rev Ges Hydrobiol 82: 169–184. [Google Scholar]
- 22. Viitasalo M, Katajisto T, Vuorinen I (1994) Seasonal dynamics of Acartia bifilosa and Eurytemora affinis (Copepoda: Calanoida) in relation to abiotic factors in the northern Baltic Sea. Hydrobiologia 292/293: 415–422. [Google Scholar]
- 23. Johansson M, Gorokhova E, Larsson U (2004) Annual variability in ciliate community structure, potential prey and predators in the open northern Baltic Sea proper. J Plankton Res 26: 67–80. [Google Scholar]
- 24. Meyer-Harms B, Reckermann M, Voss M, Siegmund H, von Bodungen B (1999) Food selection by calanoid copepods in the euphotic layer of the Gotland Sea (Baltic Proper) during mass occurrence of N2-fixing cyanobacteria. Mar Ecol Prog Ser 191: 243–250. [Google Scholar]
- 25. Rolff C (2000) Seasonal variation in δ13C and δ15N of size-fractionated plankton at a coastal station in the northern Baltic proper. Mar Ecol Prog Ser 203: 47–65. [Google Scholar]
- 26. Loick-Wilde N, Dutz J, Miltner A, Gehre M, Montoya JP, et al. (2012) Incorporation of nitrogen from N2 fixation into amino acids of zooplankton. Limnol Oceanog 57: 199–210. [Google Scholar]
- 27. Peters J, Renz J, van Beusekom J, Boersma M, Hagen W (2006) Trophodynamics and seasonal cycle of the copepod Pseudocalanus acuspes in the Central Baltic Sea (Bornholm Basin): evidence from lipid composition. Mar Biol 149: 1417–1429. [Google Scholar]
- 28. Koski M, Engström J, Viitasalo M (1999) Reproduction and survival of the calanoid copepod Eurytemora affinis fed with toxic and non-toxic cyanobacteria. Mar Ecol Prog Ser 186: 187–197. [Google Scholar]
- 29. Sellner KG, Olson MM, Olli K (1996) Copepod interactions with toxic and non-toxic cyanobacteria from the Gulf of Finland. Phycologia 35: 177–182. [Google Scholar]
- 30. Smith JL, Boyer GL, Zimba PV (2008) A review of cyanobacterial odorous and bioactive metabolites: Impacts and mangagement alternatives in aquaculture. Aquaculture 280: 5–20. [Google Scholar]
- 31. Amado LL, Monserrat JM (2010) Oxidative stress generation by microcystins in aquatic animals: Why and how. Environ Int 36: 226–235. [DOI] [PubMed] [Google Scholar]
- 32. Vehmaa A, Hogfors H, Gorokhova E, Brutemark A, Holmborn T, et al. (2013) Projected marine climate change: effects on copepod oxidative status and reproduction. Ecol Evol 3: 4548–4557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.HELCOM (2008) Manual for marine monitoring in the COMBINE Programme of HELCOM, Annex 6: Guidelines concerning phytoplankton species composition, abundance and biomass.
- 34. Olenina I, Hajdu S, Edler L, Andersson A, Wasmund N, et al. (2006) Biovolumes and size-classes of phytoplankton in the Baltic Sea. Balt Sea Environ Proc 106: 144 pp [Google Scholar]
- 35. Gorokhova E (2010) A single-step staining method to evaluate egg viability in zooplankton. Limnol Oceanogr-Meth 8: 414–423. [Google Scholar]
- 36. Gorokhova E (2003) Relationships between nucleic acid levels and egg production rates in Acartia bifilosa: implications for growth assessment of copepods in situ . Mar Ecol Prog Ser 262: 163–172. [Google Scholar]
- 37. Gorokhova E, Kyle M (2002) Analysis of nucleic acids in Daphnia: development of methods and ontogenetic variations in RNA-DNA content. J Plankton Res 24: 511–522. [Google Scholar]
- 38.Guillard RRL (1975) Culture of phytoplankton for feeding marine invertebrates, In: Smith WL, Chanley MH (Eds.) Culture of marine invertebrate animals. Plenum publishing corp, New York, pp. 29–60. [Google Scholar]
- 39. Giraffa G, Rossetti L, Neviani E (2000) An evaluation of Chelex-based DNA purification protocols for the typing of lactic acid bacteria. J Microbiol Meth 42: 175–184. [DOI] [PubMed] [Google Scholar]
- 40. Engström-Öst J, Hogfors H, El-Shehawy R, De Stasio B, Vehmaa A, et al. (2011) Toxin producing cyanobacterium Nodularia spumigena, potential competitors and grazers: testing mechanisms of reciprocal interactions in mixed plankton communities. Aquat Microb Ecol 62: 39–48. [Google Scholar]
- 41. Prior RL, Hoang H, Gu LW, Wu XL, Bacchiocca M, et al. (2003) Assays for hydrophilic and lipophilic antioxidant capacity (oxygen radical absorbance capacity (ORAC(FL))) of plasma and other biological and food samples. J Agr Food Chem 51: 3273–3279. [DOI] [PubMed] [Google Scholar]
- 42. Knuckey RM, Semmens GL, Mayer RJ, Rimmer MA (2005) Development of an optimal microalgal diet for the culture of the calanoid copepod Acartia sinjiensis: Effect of algal species and feed concentration on copepod development. Aquaculture 249: 339–351. [Google Scholar]
- 43. Gorokhova E (2005) Effects of preservation and storage of microcrustaceans in RNAlater on RNA and DNA degradation. Limnol Oceanogr-Meth 3: 143–148. [Google Scholar]
- 44. Baud A, Barthelemy RM, Nival S, Brunet M (2002) Formation of the gut in the first two naupliar stages of Acartia clausi and Hemidiaptomus roubaui (Copepoda, Calanoida): comparative structural and ultrastructural aspects. Can J Zool 80: 232–244. [Google Scholar]
- 45. Holmborn T, Lindell K, Holeton C, Hogfors H, Gorokhova E (2009) Biochemical proxies for growth and metabolism in Acartia bifilosa (Copepoda, Calanoida). Limnol Oceanogr Methods 7: 785–794. [Google Scholar]
- 46. Guedes AC, Gião MS, Seabra R, Ferreira ACS, Tamagnini P, et al. (2013) Evaluation of the antioxidant activity of cell extracts from microalgae. Mar Drugs 11: 1256–1270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Parsons PA (2001) The hormetic zone: An ecological and evolutionary perspective based upon habitat characteristics and fitness selection. Q Rev Biol 76: 459–467. [DOI] [PubMed] [Google Scholar]
- 48. Motwani NH, Gorokhova E (2013) Metazooplankton grazing on picocyanobacteria as inferred from molecular diet analysis. PLoS ONE 8(11): e79230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Singh S, Kate BN, Banerjee UC (2005) Bioactive compounds from cyanobacteria and microalgae: An overview. Crit Rev Biotechnol 25: 73–95. [DOI] [PubMed] [Google Scholar]
- 50. Jiang XD, Lonsdale DJ, Gobler CJ (2010) Density-dependent nutritional value of the dinoflagellate Cochlodinium polykrikoides to the copepod Acartia tonsa . Limnol Oceanogr 55: 1643–1652. [Google Scholar]
- 51. Vargas MA, Rodriguez H, Moreno J, Olivares H, Del Campo JA, et al. (1998) Biochemical composition and fatty acid content of filamentous nitrogen-fixing cyanobacteria. J Phycol 34: 812–817. [Google Scholar]
- 52. Pandey U, Pandey J (2009) Enhanced production of delta-aminolevulinic acid, bilipigments, and antioxidants from tropical algae of India. Biotechnol Bioproc E 14: 316–321. [Google Scholar]
- 53. Stal LJ, Staal M, Villbrandt M (1999) Nutrient control of cyanobacterial blooms in the Baltic Sea. Aquat Microb Ecol 18: 165–173. [Google Scholar]
- 54. Kiørboe T (1989) Phytoplankton growth rate and nitrogen content: implications for feeding and fecundity in a herbivorous copepod. Mar Ecol Prog Ser 55: 229–234. [Google Scholar]
- 55. Pinho GLL, da Rosa CM, Maciel FE, Bianchini A, Yunes JS, et al. (2005) Antioxidant responses and oxidative stress after microcystin exposure in the hepatopancreas of an estuarine crab species. Ecotox Environ Safe 61: 353–360. [DOI] [PubMed] [Google Scholar]
- 56. Howitz KT, Sinclair DA (2008) Xenohormesis: Sensing the Chemical Cues of Other Species. Cell 133: 387–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Forbes VE (2000) Is hormesis an evolutionary expectation? Funct Ecol 14: 12–24. [Google Scholar]
- 58. Mattson MP, Cheng AW (2006) Neurohormetic phytochemicals: low-dose toxins that induce adaptive neuronal stress responses. Trends Neurosci 29: 632–639. [DOI] [PubMed] [Google Scholar]
- 59. Sellner KG (1997) Physiology, ecology, and toxic properties of marine cyanobacteria blooms. Limnol Oceanogr 42: 1089–1104. [Google Scholar]
- 60. Degerholm J, Gundersen K, Bergman B, Söderbäck E (2008) Seasonal significance of N2 fixation in coastal and offshore waters of the northwestern Baltic Sea. Mar Ecol Prog Ser 360: 73–84. [Google Scholar]
- 61. Larsson U, Hajdu S, Walve J, Elmgren R (2001) Baltic Sea nitrogen fixation estimated from the summer increase in upper mixed layer total nitrogen. Limnol Oceanogr 46: 811–820. [Google Scholar]
- 62. Ploug H, Adam B, Musat N, Kalvelage T, Lavik G, et al. (2011) Carbon, nitrogen and O2 fluxes associated with the cyanobacterium Nodularia spumigena in the Balric Sea. ISME J 5: 1549–1558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Ploug H, Musat N, Adam B, Moraru CL, Lavik G, et al. (2010) Carbon and nitrogen fluxes associated with the cyanobacterium Aphanizomenon sp. in the Baltic Sea. ISME J 4: 1215–1223. [DOI] [PubMed] [Google Scholar]
- 64. Ohlendieck U, Stuhr A, Siegmund H (2000) Nitrogen fixation by diazotrophic cyanobacteria in the Baltic Sea and transfer of the newly fixed nitrogen to picoplankton organisms. J Mar Syst 25: 213–219. [Google Scholar]
- 65. de Kluijver A, Yu J, Houtekamer M, Middelburg JJ, Liu Z (2012) Cyanobacteria as a carbon source for zooplantkon in eutrophic Lake Taihu, China, measured by 13C labeling and fatty acid biomarkers. Limnol Oceanogr 57: 1245–1254. [Google Scholar]
- 66. Wilson SE, Steinberg DK (2010) Autotrophic picoplankton in mesozooplankton guts: evidence of aggregate feeding in the mesopelagic zone and export of small phytoplankton. Mar Ecol Prog Ser 412: 11–27. [Google Scholar]
- 67. Hoppe HG (1981) Blue-green algae agglomeration in surface water: a microbiotope of highly bacterial activity. Kieler Meeresforsch Sonderh 5: 291–303. [Google Scholar]
- 68. Suikkanen S, Laamanen M, Huttunen M (2007) Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuar Coast Shelf Sci 71: 580–592. [Google Scholar]
- 69. Miralto A, Barone G, Romano G, Poulet SA, Ianora A, et al. (1999) The insidious effect of diatoms on copepod reproduction. Nature 402: 173–176. [Google Scholar]
- 70. Gorokhova E, Lehtiniemi M, Motwani NH (2013) Trade-offs between predation risk and growth benefits in copepods Eurytemora affinis with contrasting pigmentation. PLoS ONE 8(8): e71385 doi:10.1371/journal.pone.0071385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Gifford DJ (1991) The protozoan-metazoan trophic link in pelagic ecosystems. J Protozool 38: 81–86. [Google Scholar]
- 72. Engström J, Koski M, Viitasalo M, Reinikainen M, Repka S, et al. (2000) Feeding interactions of the copepods Eurytemora affinis and Acartia bifilosa with the cyanobacteria Nodularia sp. J Plankton Res 22: 1403–1409. [Google Scholar]
- 73. Komárek J, Anagnostidis K (1989) Modern approach to the classification system of Cyanophytes 4 — Nostocales. Algol Stud Arch Hydrobiol Suppl56: 247–345. [Google Scholar]
- 74. Breteler WCM, Schogt N, Baas M, Schouten S, Kraay GW (1999) Trophic upgrading of food quality by protozoans enhancing copepod growth: role of essential lipids. Mar Biol 135: 191–198. [Google Scholar]
- 75. Tuomainen J, Hietanen S, Kuparinen J, Martikainen PJ, Servomaa K (2006) Community structure of the bacteria associated with Nodularia sp. (cyanobacteria) aggregates in the Baltic Sea. Microb Ecol 52: 513–522. [DOI] [PubMed] [Google Scholar]
- 76. Berggreen U, Hansen B, Kiørboe T (1988) Food size spectra, ingestion and growth of the copepod Acartia tonsa during development — implications for determination of copepod production. Mar Biol 99: 341–352. [Google Scholar]
- 77. Holland DP, van ErpI, Beardall J, Cook PLM (2012) Environmental controls on the nitrogen-fixing cyanobacterium Nodularia spumigena in a temperate lagoon system in SE Australia. Mar Ecol Prog Ser 461: 47–57. [Google Scholar]
- 78. Wannicke N, Korth F, Liskow I, Voss M (2013) Incorporation of diazotrophic fixed N2 by mesozooplankton — Case studies in the southern Baltic Sea. J Mar Syst 117–118: 1–13. [Google Scholar]
- 79. Woodland RJ, Holland DP, Beardall J, Smith J, Scicluna T, et al. (2013) Assimilation of diazotrophic nitrogen into pelagic food webs. PLoS ONE 8(6): e67588. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Acartia tonsa : Data on reproductive and growth variables obtained in the summer field survey conducted in July - September 2007, at three coastal stations (B1, H2 and H4) in the northern Baltic proper. Variables measured: EPR (egg production rate), VEPR (viable egg production rate), RNA (individual ribonucleic acid content), and PL (prosome length).
(XLSX)
Acartia bifilosa : Data on feeding, reproduction and nauplii development obtained in the laboratory experiment conducted in August 2010, in the Storfjärden Bay, Western Finland. Variables measured: EPR (egg production rate), VEPR (viable egg production rate), RNA (individual ribonucleic acid content), and PL (prosome length).
(XLSX)
Acartia sp. and Eurytemora affinis : Long-term data (mean ± SD; period 1999–2009) on RNA:DNA ratio in nauplii at stns B1 and H4 in the northern Baltic proper. The mean values are based on 3–4 replicate samples analyzed per year (July-August).
(XLSX)
Growth and reproduction indices observed in the (A) field survey, (B) experimental study, and (C) long-term dataset. Data are presented as means with their standard deviations; abbreviations as in Tables S1 and S2.
(DOC)
Pearson r for correlations among phytoplankton groups at stn B1 and stn H4. Significant correlations are in bold face (p<0.05). Abbreviations: CHLO – Chlorophyceae, CHRY – Chrysophyceae, CRYP – Cryptophyceae, CYAN – Cyanophyceae, DIAT – Diatomea, DINO – Dinophyceae, EUGL – Euglenophyceae, MESO – Mesodinium rubrum (Myrionecta rubrum), PRAS – Prasinophyceae, PRYM – Prymnesiophyceae, UNID – unidentified flagellates, and HDIN – heterotrophic dinoflagellates.
(DOC)
Summary of the observed effects linking filamentous cyanobacteria to reproductive output, growth indices and oxidative status in copepods. Abbreviations as in Tables S1 and S2.
(DOC)
Oxidative stress biomarkers.
(DOCX)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. The environmental data and phytoplankton biovolume data used in all analyses are available from www.smhi.se (SHARK database) and www.2.ecology.su.se/dbhfj/index.htm, and data on copepod growth- and reproduction-related variables are provided as Supporting Information (Summer field survey: Table S1; Laboratory experiment: Table S2; and Long-term data analysis: Table S3).