

Abstract
Retinoids and 1α,25-dihydroxyvitamin D3 (1,25(OH)2D3) induce differentiation of myeloid leukemia cells into granulocyte and macrophage lineages, respectively. All-trans retinoic acid (ATRA), which is effective in the treatment of acute promyelocytic leukemia, can induce differentiation of other types of myeloid leukemia cells, and combined treatment with retinoid and 1,25(OH)2D3 effectively enhances the differentiation of leukemia cells into macrophage-like cells. Recent work has classified macrophages into M1 and M2 types. In this study, we investigated the effect of combined treatment with retinoid and 1,25(OH)2D3 on differentiation of myeloid leukemia THP-1 and HL60 cells. 9-cis Retinoic acid (9cRA) plus 1,25(OH)2D3 inhibited proliferation of THP-1 and HL60 cells and increased myeloid differentiation markers including nitroblue tetrazolium reducing activity and expression of CD14 and CD11b. ATRA and the synthetic retinoic acid receptor agonist Am80 exhibited similar effects in combination with 1,25(OH)2D3 but less effectively than 9cRA, while the retinoid X receptor agonist HX630 was not effective. 9cRA plus 1,25(OH)2D3 effectively increased expression of M2 macrophage marker genes, such as CD163, ARG1 and IL10, increased surface CD163 expression, and induced interleukin-10 secretion in myeloid leukemia cells, while 9cRA alone had weaker effects on these phenotypes and 1,25(OH)2D3 was not effective. Taken together, our results demonstrate selective induction of M2 macrophage markers in human myeloid leukemia cells by combined treatment with 9cRA and 1,25(OH)2D3.
Introduction
Retinoids play roles in numerous biological functions, such as cellular proliferation and differentiation, embryogenesis, immunity and metabolism [1]. An active natural retinoid, all-trans retinoic acid (ATRA), is effective in differentiation therapy for acute promyelocytic leukemia (APL) [2]. APL is a subtype of acute myeloid leukemia, which is characterized by a specific chromosomal abnormality t(15,17) associated with a genetic rearrangement between retinoic acid receptor α (RARα) (gene symbol, RARA) and the promyelocytic leukemia gene PML [2]. RARα plays a role in granulocytic differentiation of hematopoietic cells and the abnormal chimeric receptor PML-RARα has been implicated in APL pathogenesis by blocking the myeloid differentiation program and enhancing self-renewal of leukemic cells [3], [4]. Pharmacological doses of ATRA induce differentiation of APL cells into granulocytes through degradation of PML-RARα and recovery of physiological RARα signaling [3], .
Retinoids, including ATRA, 9-cis retinoic acid (9cRA) and synthetic RAR ligands, exhibit anti-tumor effects not only on APL but also on other malignancies, such as breast cancer, lung cancer, and head and neck cancer [5]. With regard to leukemia, ATRA was first reported to induce the differentiation of human myeloid leukemia HL60 cells towards the granulocytic lineage [6], [7]. Importantly, HL60 cells are derived from non-APL leukemia without t(15,17) [8], and ATRA can also induce differentiation of leukemia cells from non-APL myeloid leukemia patients [9]. Retinoids in combination with other differentiation inducers, such as 1α,25-dihydroxyvitamin D3 (1,25(OH)2D3) and dibutyryl cAMP, synergistically induce differentiation of leukemia cells [10]–[13]. However, the underlying mechanisms of retinoid-induced differentiation of leukemia cells remain poorly understood and retinoids have not been utilized in the treatment of myeloid leukemia other than APL.
The active form of vitamin D3, 1,25(OH)2D3, regulates calcium and bone homeostasis, immunity, and cellular growth and differentiation through direct binding to the vitamin D receptor (VDR), and has been demonstrated to inhibit the proliferation and to induce the differentiation of various types of malignant cells, including breast, prostate and colon cancers as well as myeloid leukemia cells [14], [15]. The administration of 1,25(OH)2D3 and its analog has therapeutic effects in a mouse model of myeloid leukemia [16]. While ATRA induces granulocytic differentiation [6], [7], 1,25(OH)2D3 induces the differentiation of HL60 cells and other myeloid leukemia cells towards the monocyte and macrophage lineage [17], [18]. Interestingly, ATRA induces monocytic differentiation of monoblastic leukemia U937 and THP-1 cells [13], [19]. Combined treatment with 1,25(OH)2D3 and retinoids induces the differentiation of HL60 cells and human monoblastic leukemia cells, such as THP-1 cells, to monocyte/macrophage-lineage cells more effectively than 1,25(OH)2D3 alone [10], [12], [20]. Although 1,25(OH)2D3 has been shown to exert its biological effects on cellular proliferation and differentiation by genomic and/or non-genomic pathways [21], the detailed mechanisms remain unclear. Macrophages have been classified into two cell types, classically activated M1 macrophages and alternatively activated M2 macrophages [22], [23]. While M1 macrophages produce proinflammatory cytokines and enhance microbicidal and tumoricidal immunity, M2 macrophages are involved in wound healing and immune regulation. Although retinoids and 1,25(OH)2D3 play functional roles in monocytes and macrophages [4], [24], the macrophage cell type resulting from 1,25(OH)2D3 and/or retinoid differentiation of myeloid leukemia cells has not been further characterized. In this study, we examined the effects of 1,25(OH)2D3 in combination with retinoids on differentiation of myeloid leukemia cells and found that 1,25(OH)2D3 in combination with 9cRA and ATRA induce the differentiation of myeloid leukemia cells to macrophages with M2-like phenotype.
Materials and Methods
Compounds
1,25(OH)2D3, ATRA and 9cRA were purchased from Wako Pure Chemical Industries (Osaka, Japan). Am80 (4-[(5,6,7,8-tetrahydro-5,5,8,8-tetramethyl-2-naphthalenyl)carbamoyl]benzoic acid) [25] and HX630 (4-[2,3-(2,5-dimethyl-2,5-hexano)dibenzo[b,f][1], [4]-thiazepin-11-yl]benzoic acid) [26] were kindly provided by Dr. Koichi Shudo of Research Foundation ITSUU Laboratory (Tokyo, Japan).
Cell culture, cell growth, nitroblue tetrazolium (NBT) reduction, and interleukin-10 (IL-10) production
Human myeloid leukemia HL60 and THP-1 cells (RIKEN Cell Bank, Tsukuba, Japan) were maintained in RPMI1640 medium containing 10% fetal bovine serum, 100 unit/ml penicillin, and 100 µg/ml streptomycin in a humidified atmosphere containing 5% CO2. Suspensions of cells (105 cells/ml) were cultured with or without test compounds at pharmacological concentrations (30−100 nM) according to our preliminary experiments and the previous reports [10]–[12], [27]. Cell numbers were counted in a Z1S Coulter Counter (Beckman Coulter, Fullerton, CA). Cell morphology was examined in cell smears stained with May-Grünwald-Giemsa. NBT reduction was assayed colorimetrically and NBT-reducing activity data were normalized to the cell numbers [28]. IL-10 levels in culture media were determined with the Human IL-10 ELISA MAX Standard kit (BioLegend, San Diego, CA).
Flow cytometry
Expression of cell surface antigens, CD14, CD11b and CD163, were determined with immunofluorescence staining and flow cytometry [29]. FITC mouse anti-human CD14, PE mouse anti-human CD11b, PE mouse anti-human CD163, and isotype control antibodies were purchased from Becton, Dickinson and Company (Franklin Lakes, NJ). The stained cells were assayed with a flow cytometer (BD FACSCalibur; Becton, Dickinson and Company) and analyzed with the BD CellQuest software (Becton, Dickinson and Company).
Reverse transcription and real-time quantitative polymerase chain reaction
Total RNAs from samples were prepared by the acid guanidine thiocyanate-phenol/chloroform method [30]. cDNAs were synthesized using the ImProm-II Reverse Transcription system (Promega Corporation, Madison, WI). Intron-spanning primers were as follows: CD163 (GenBank accession no. NM_004244), 5′-ACT GCA AGA ACT GGC AAT GG-3′ and 5′-CCA TGC TTC ACT TCA ACA CG-3′; ARG1 (GenBank accession no. NM_000045), 5′-TCC AAG GTC TGT GGG AAA AG-3′ and 5′-ATT GCC AAA CTG TGG TCT CC-3′; IL10 (GenBank accession no. NM_000572), 5′-CCA AGA CCC AGA CAT CAA GG-3′ and 5′-GGC CTT GCT CTT GTT TTC AC-3′; IL12B (GenBank accession no. NM_002187), 5′-ATT GAG GTC ATG GTG GAT GC-3′ and 5′-TTC TTG GGT GGG TCA GGT TT-3′; TGFB1 (GenBank accession no. NM_000660), 5′-CAA CAA TTC CTG GCG ATA CCT C-3′ and 5′-AAA GCC CTC AAT TTC CCC TC-3′; TNF (GenBank accession no. NM_000594), 5′-TGC TTG TTC CTC AGC CTC TT-3′ and 5′-TGA GGT ACA GGC CCT CTG AT-3′; IL6 (GenBank accession no. NM_000600), 5′-AAA GAG GCA CTG GCA GAA AA-3′ and 5′-AAA GCT GCG CAG AAT GAG AT-3′; NOS2 (GenBank accession no. NM_000625), 5′-TAC CCC TCC AGA TGA GCT TC-3′ and 5′-TCT CCT TTG TTA CCG CTT CC-3′. Other primer sequences have been reported previously [28]. The mRNA values were normalized to the amount of β-actin mRNA.
Statistical analysis
All values are shown as mean ± S.D. We performed one-way ANOVA followed by Tukey’s multiple comparisons or two-way ANOVA to assess significant differences using Prism 6 (Graphpad Software, La Jolla, CA).
Results
Induction of differentiation of human myeloid leukemia cells by retinoids plus 1,25(OH)2D3
We examined the effects of 9cRA and ATRA in the absence or presence of 1,25(OH)2D3 on NBT-reducing activity, a marker of myeloid differentiation, in monoblastic leukemia THP-1 cells and promyelocytic leukemia HL60 cells. 9cRA (100 nM) increased NBT-reducing activity in THP-1 and HL60 cells, while ATRA (100 nM) was not effective (Fig. 1A), consistent with the previous reports showing that 9cRA is more potent than ATRA in inducing differentiation of leukemia cells [20], [31], [32]. In combination with 1,25(OH)2D3, 9cRA and ATRA effectively increased NBT-reducing activity in these cells (Fig. 1A). Am80 (also called tamibarotene) is a potent synthetic RAR agonist that is used in the treatment of recurrent APL in Japan [33], [34]. HX630 is an RXR selective agonist derived from LE135, an RAR antagonist [26]. HX630 does not exhibit RAR antagonistic activity but enhances the differentiation-inducing activity of Am80 in HL60 cells [33]. We also examined the effects of Am80 and HX630 in the absence or presence of 1,25(OH)2D3. Although Am80 and HX630 at 100 nM were not effective, the combination of 1,25(OH)2D3 with Am80 but not HX630 significantly increased NBT-reducing activity in THP-1 and HL60 cells (Fig. 1A).
Figure 1. Induction of differentiation of THP-1 and HL60 cells by combined treatment with retinoid and 1,25(OH)2D3.

(A) NBT-reducing activities. Cells were treated with vehicle control (Cont), 100 nM 9cRA, ATRA, Am80 or HX630 in the absence or presence of 100 nM 1,25(OH)2D3 (D3) for 5 days. *, p<0.05; **, p<0.01; ***, p<0.001 (one-way ANOVA followed by Tukey’s multiple comparisons). (B) Morphological changes of THP-1 and HL60 cells treated with 9cRA and/or 1,25(OH)2D3. Cells were treated with vehicle control (Cont), 100 nM 9cRA and/or 100 nM 1,25(OH)2D3 for 5 days and the cell smears were stained with May-Grünwald-Giemsa. (C) Cell proliferations. Cells (1×105/ml) were cultured with vehicle control (Cont), 100 nM 9cRA and/or 100 nm 1,25(OH)2D3 (D3), and cell numbers were counted at indicated days. *, p<0.05; **, p<0.01; ***, p<0.001 vs Cont; ###, p<0.001 vs 9cRA; +++, p<0.001 vs D3 (one-way ANOVA followed by Tukey’s multiple comparisons). †††, p<0.001 (two-way ANOVA).
THP-1 cells and HL60 cells were treated with 1,25(OH)2D3 and/or 9cRA and the morphological features were examined. While untreated THP-1 cells had a basophilic cytoplasm and large nuclei with several nucleoli, cells treated with 9cRA (100 nM) had grayish, enlarged cytoplasm and slightly lobulated nuclei (Fig. 1B). Although 1,25(OH)2D3 (100 nM) treatment did not induce an apparent morphological change, the combination of 9cRA and 1,25(OH)2D3 enlarged the grayish cytoplasm area more effectively than 9cRA alone. Untreated HL60 cells have promyelocytic features, although they are not derived from APL having t(15;17) [8]. While HL60 treated with 1,25(OH)2D3 (100 nM) had slightly less basophilic cytoplasm and decreased nuclear-cytoplasmic ratio, 9cRA (100 nM) induced differentiation of HL60 cells into myelocytic cells having slightly lobulated nuclei and a decreased nuclear-cytoplasmic ratio (Fig. 1B). Combined treatment with 9cRA and 1,25(OH)2D3 enhanced the monocytic features of HL60 cells with enlarged grayish cytoplasm and a further decrease in the nuclear-cytoplasmic ratio. These findings are consistent with the previous reports showing monocytic differentiation of myeloid leukemia cells by 1,25(OH)2D3 plus 9cRA or ATRA [32], [35]. 9cRA (100 nM) suppressed the proliferation of THP-1 cells, and although 1,25(OH)2D3 (100 nM) was not effective, combined treatment with 1,25(OH)2D3 enhanced the anti-proliferative effect of 9cRA (Fig. 1C). While treatment with 9cRA (100 nM) or 1,25(OH)2D3 (100 nM) alone was not effective, the combined treatment with 9cRA and 1,25(OH)2D3 effectively suppressed HL60 proliferation (Fig. 1C).
We next examined the effects of combined treatment with retinoids and 1,25(OH)2D3 on expression of surface antigens, CD14 and CD11b, additional markers of myelomonocytic differentiation. 1,25(OH)2D3 at 100 nM slightly increased CD14 expression in THP-1 cells (Fig. 2A). 9cRA at 100 nM increased CD14 expression and the combination of 9cRA and 1,25(OH)2D3 increased CD14 expression more strongly than single use of these compounds. Interestingly, although ATRA and Am80 were not effective, these retinoids enhanced CD14 expression in combination with 1,25(OH)2D3. HX630 did not increase CD14 expression in the absence or presence of 1,25(OH)2D3. 1,25(OH)2D3 also slightly increased CD11b expression in THP-1 cells (Fig. 2B). 9cRA, ATRA and Am80 at 30 nM increased CD11b expression, and effectively enhanced CD11b expression induced by 1,25(OH)2D3. HX630 did not increase CD11b expression in the absence or presence of 1,25(OH)2D3. Transcriptional induction of CD14, a VDR target gene [36], is also associated with myeloid differentiation [28]. Retinoid treatment did not increase CD14 mRNA levels in THP-1 and HL60 cells (Fig. 2C). Interestingly, while the effect of 1,25(OH)2D3 (100 nM) alone was not significant, combinations of 1,25(OH)2D3 with 9cRA, ATRA and Am80 effectively increased CD14 mRNA expression in these cells. The combination of HX630 and 1,25(OH)2D3 was not effective in CD14 mRNA induction. Thus, when combined with 1,25(OH)2D3, a RAR/RXR agonist (9cRA) and RAR agonists (ATRA and Am80), but not a RXR agonist (HX630), effectively induce differentiation of myeloid leukemia THP-1 and HL60 cells.
Figure 2. Effects of combined treatment with retinoid and 1,25(OH)2D3 on cell surface CD14 and CD11b expression and CD14 mRNA expression.

Representative histograms of CD14 expression (A) and CD11b expression (B) in THP-1 cells. Cells were treated with vehicle control (Cont), 100 nM 9cRA, ATRA, Am80 or HX630 in the absence or presence of 100 nM 1,25(OH)2D3 (D3) for 96 hours. Filled curves, vehicle control. Similar results were obtained from repeated experiments. (C) CD14 mRNA levels in THP-1 and HL60 cells. Cells were treated with vehicle control (Cont), 30 nM 9cRA, ATRA, Am80 or HX630 in the absence or presence of 100 nM 1,25(OH)2D3 (D3) for 72 hours. **, p<0.01; ***, p<0.001 (one-way ANOVA followed by Tukey’s multiple comparisons).
Induction of M2 macrophage markers in human myeloid leukemia cells by retinoids plus 1,25(OH)2D3
As shown in Figure 1B and the previous reports [10], [20], [32], combined treatment with retinoid and 1,25(OH)2D3 induces differentiation of myeloid leukemia cells into the monocytic lineage rather than the granulocytic lineage. We examined whether leukemia cells treated with retinoid plus 1,25(OH)2D3 exhibit M1 or M2 macrophage phenotypes. Expression of the CD163, ARG1, IL10 and TGFB1 marker genes is associated with M2 macrophage activation, whereas IL12B, TNF, IL6 and NOS2 expression is increased in M1 macrophages [23]. Treatment of THP-1 cells with 9cRA, ATRA or 1,25(OH)2D3 alone did not induce CD163 mRNA expression, but the combination of 9cRA and 1,25(OH)2D3 effectively increased CD163 mRNA levels (Fig. 3A). ATRA plus 1,25(OH)2D3 also increased CD163 expression but less effectively than 9cRA plus 1,25(OH)2D3. Although CD163 mRNA expression was not detected in HL60 cells treated with 9cRA or ATRA alone, 9cRA plus 1,25(OH)2D3 and ATRA plus 1,25(OH)2D3 tended to increase CD163 mRNA levels. 9cRA plus 1,25(OH)2D3 also effectively increased ARG1 mRNA expression in THP-1 cells, while ATRA plus 1,25(OH)2D3 tended to increase its expression, an effect that did not reach statistical significance (Fig. 3B). Interestingly, combination of ATRA and 1,25(OH)2D3 effectively increased ARG1 mRNA levels in HL60 cells, while the combined effect of 9cRA and 1,25(OH)2D3 did not reach statistical significance. IL10 mRNA expression was also elevated in THP-1 cells treated with 9cRA plus 1,25(OH)2D3 and, to a lesser extent, ATRA plus 1,25(OH)2D3 (Fig. 3C). Although treatment of HL60 cells with 1,25(OH)2D3 decreased IL10 mRNA levels, the combination of 1,25(OH)2D3 with 9cRA or ATRA increased them to control levels. Combination of 1,25(OH)2D3 with 9cRA or ATRA did not increase IL12B mRNA levels in THP-1 and HL60 cells (Fig. 3D). IL12B mRNA levels in THP-1 cells treated with 9cRA were decreased by combined treatment with 1,25(OH)2D3. Addition of 9cRA increased TGFB1 mRNA levels in THP-1 cells treated with 1,25(OH)2D3 (Fig. 3E). 9cRA increased TNF mRNA expression in THP-1 cells but combined treatment with 1,25(OH)2D3 decreased its expression. IL6 mRNA expression was increased by the combination of 9cRA and 1,25(OH)2D3. 9cRA and/or 1,25(OH)2D3 induced no significant change in NOS2 mRNA expression. Thus, the combination of retinoid and 1,25(OH)2D3 increases expression of genes associated with M2 macrophages.
Figure 3. Expression of marker genes of M1 and M2 macrophages.

mRNA levels of the M2 markers CD163 (A), ARG1 (B), IL10 (C), and the M1 marker IL12B (D) in THP-1 and HL60 cells. (E) mRNA levels of the M2 marker TGFB1, the M1 markers TNF, IL6, and NOS2 in THP-1 cells. Cells were treated with vehicle control (Cont), 30 nM 9cRA, or ATRA in the absence or presence of 100 nM 1,25(OH)2D3 (D3) for 72 hours. *, p<0.05; **, p<0.01; ***, p<0.001 (one-way ANOVA followed by Tukey’s multiple comparisons). n.d., not detected.
We further examined expression of CD163 as a cell surface marker of M2 macrophages in THP-1 cells. Figure 4A shows representative flow cytometric analysis of THP-1 cells treated with or without 9cRA and 1,25(OH)2D3 using anti-CD14 and anti-CD163 antibodies. 9cRA but not 1,25(OH)2D3 increased CD163 mean fluorescence intensity in THP-1 cells, and the combination of 9cRA and 1,25(OH)2D3 effectively enhanced the intensity values (Fig. 4B). 9cRA increased and 1,25(OH)2D3 slightly decreased surface CD14 mean fluorescence intensity, but 9cRA plus 1,25(OH)2D3 also strongly increased the CD14 intensity (Fig. 4B). 9cRA treatment increased the percentage of both CD163+/CD14+ cells and CD163−/CD14+ cells (Fig. 4B). Combination of 1,25(OH)2D3 with 9CRA increased the percentage of CD163+/CD14+ cells but not of CD163−/CD14+ cells. These findings are consistent with induction of the M2 macrophage phenotype in THP-1 cells by 9cRA plus 1,25(OH)2D3.
Figure 4. Cell surface expression of CD163 in THP-1 cells.
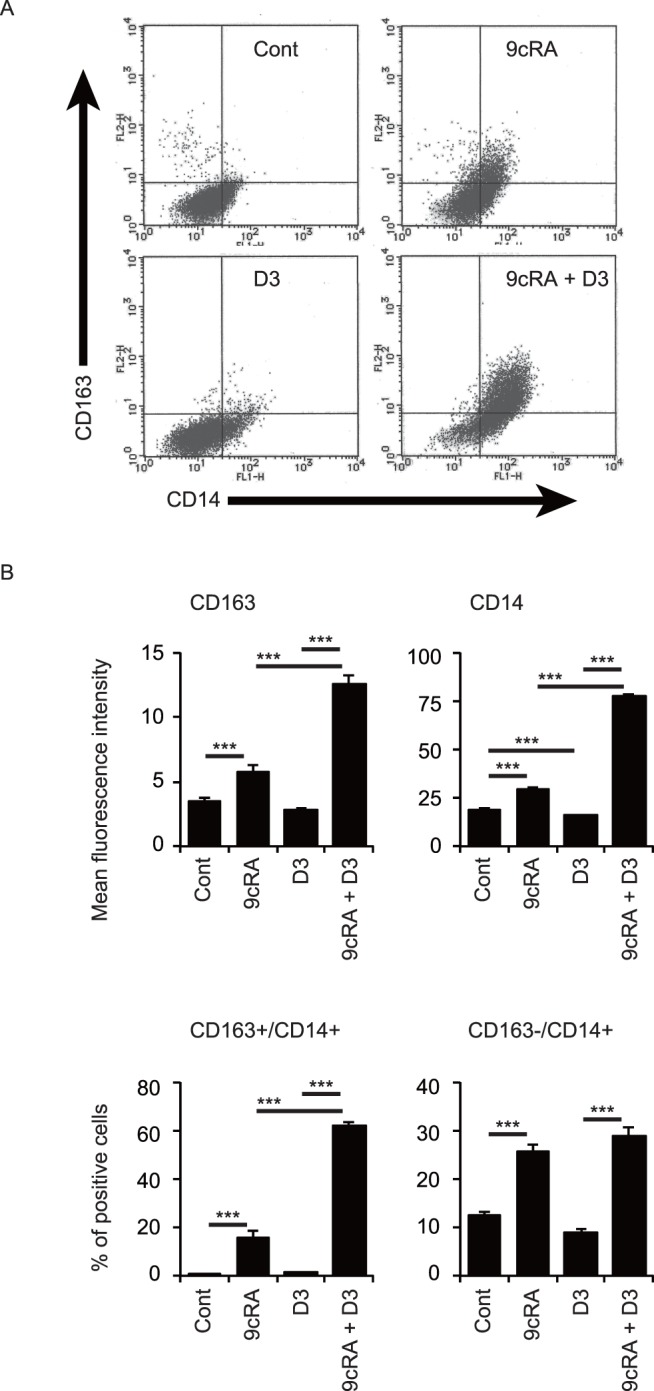
(A) Representative histograms of CD14 and CD163 expression. (B) Quantification of mean fluorescence intensity of CD163 and CD14 expression. (C) Quantification of percentages of CD163+/CD14+ cells and CD163−/CD14+ cells. Cells were treated with vehicle control (Cont), 100 nM 9cRA and/or 100 nM 1,25(OH)2D3 (D3) for 96 hours. **, p<0.01; ***, p<0.001 (one-way ANOVA followed by Tukey’s multiple comparisons).
The combination of 9cRA and 1,25(OH)2D3 induces IL-10 protein secretion in THP-1 and HL60 cells
Finally, we examined IL-10 protein levels in conditioned media of THP-1 and HL60 cells treated with 9cRA and/or 1,25(OH)2D3. IL-10 protein levels from THP-1 cells treated with 9cRA and 1,25(OH)2D3 alone were below detection limits, but the combination of these compounds effectively induced IL-10 protein secretion from these cells (Fig. 5). While IL-10 protein was detected in the culture media of untreated HL60 cells, 9cRA increased and 1,25(OH)2D3 decreased the protein level. 9cRA plus 1,25(OH)2D3 effectively increased the IL-10 protein levels in media. Thus, combined treatment of leukemia cells with 9cRA and 1,25(OH)2D3 induces IL-10 protein secretion as well as increased expression of M2 macrophage markers.
Figure 5. Secreted IL-10 production in THP-1 and HL60 cells.

Cells were treated with vehicle control (Cont), 30 nM 9cRA and/or 100 nM 1,25(OH)2D3 (D3) for 72 hours and secreted IL-10 levels in media were measured. **, p<0.01; ***, p<0.001 (one-way ANOVA followed by Tukey’s multiple comparisons). n.d., not detected.
Discussion
In this study, we found that combined treatment with retinoid and 1,25(OH)2D3 induces the differentiation of human myeloid leukemia THP-1 and HL60 cells into the monocytic lineage with a M2 macrophage phenotype. ATRA induces granulocytic differentiation of promyelocytic leukemia HL60 cells but monocytic differentiation of monoblastic U937 and THP-1 cells [6], [7], [13], [19]. Combination of ATRA or 9cRA with 1,25(OH)2D3 effectively induces monocyte/macrophage phenotypes, such as phagocytic activity, monocyte-specific esterase, lysozyme secretion, and CSF1R expression, in HL60 and U937 cells [10], [32], [37]. We observed a monocytic morphology and increased CD14 expression in HL60 and THP-1 cells treated with 9cRA plus 1,25(OH)2D3 (Figs. 1 and 2). ATRA plus 1,25(OH)2D3 also induces differentiation of promyelocytic AML-193 cells into cells that display both a typical neutrophilic morphology and monocyte-specific properties, such as CD14 expression and monocyte-specific esterase, a hybrid granulomonocytic phenotype [35]. RAR signaling plays an important role in hematopoiesis and RARα is involved in neutrophil development [4]. RAR and RXR signaling pathways have been reported to regulate monocyte/macrophage function [4]. However, it remains to be determined how retinoid signaling enhances monocytic differentiation induced by 1,25(OH)2D3 in myeloid leukemia cells.
Among retinoids, 9cRA, ATRA and Am80, but not HX630, in combination with 1,25(OH)2D3 exhibit effective differentiation-inducing activity in these cells (Figs. 1 and 2). 1,25(OH)2D3 acts as a ligand for the nuclear receptor VDR, which forms a heterodimer with RXR [38], and the VDR−RXR heterodimer is not permissive to RXR ligand activation [39]. RAR selective ligands exhibit stronger synergistic effects with 1,25(OH)2D3 than RXR selective ligands in inhibiting proliferation and inducing differentiation of monoblastic U937 cells [27]. Combined effects of retinoid and 1,25(OH)2D3 on differentiation of myelomonocytic leukemia cells are likely mediated by VDR and RAR activation. RXR also forms heterodimers with RAR and other nuclear receptors, including peroxisome proliferator-activated receptor (PPAR) and liver X receptor (LXR) [38]. The RAR−RXR heterodimer is activated by RXR ligand only in the presence of RAR ligand, a feature known as conditional permissivity [39]. 9cRA exhibits differentiation-inducing activity more effectively than ATRA in the absence or presence of 1,25(OH)2D3 (Figs. 1 and 2), in agreement with previous reports [20], [31], [32]. Since 9cRA acts as a ligand for both RAR and RXR [40], synergistic activation may be due to binding to both RAR and RXR in the RAR−RXR heterodimer. In addition, RXR ligands can activate permissive heterodimers, such as PPAR−RXR and LXR−RXR [38]. PPARγ ligand and LXR ligand have been reported to induce differentiation of myeloid leukemia cells [41], [42]. RXR ligand activation of these permissive heterodimers may also contribute to the effect of 9cRA. However, the pure RXR ligand HX630 alone and in combination with 1,25(OH)2D3 was not effective in inducing differentiation of THP-1 and HL60 cells, while the combination of 1,25(OH)2D3 with the RAR selective agonist Am80 induced the differentiation of these cells (Figs. 1 and 2). These findings suggest that cooperation between VDR signaling and RAR signaling, not RXR signaling, plays a role in the differentiation of myeloid leukemia cells. VDR activation changes expression of many genes, including those involved in cellular proliferation, differentiation and apoptosis [21]. 1,25(OH)2D3 treatment can modulate intracellular kinase pathways via a non-genomic mechanism, and it remains unknown whether the non-genomic actions are mediated through VDR or other proteins [21]. Although both genomic and non-genomic effects of 1,25(OH)2D3 have been shown to play roles in differentiation induction of leukemia cells, the detailed mechanisms remain to be elucidated. RAR signaling may modulate the vitamin D signaling pathway or regulate other differentiation mechanisms. Further studies are needed to elucidate molecular mechanisms involving VDR, RAR and RXR signaling pathways in the induced differentiation of leukemia cells.
Combined treatment with 9cRA and 1,25(OH)2D3 increased mRNA expression of CD163, ARG1, IL10, and TGFB1 genes (Fig. 3), surface expression of CD163 proteins (Fig. 4) and IL-10 secretion in THP-1 cells (Fig. 5). This combination also increased CD163, ARG1 and IL10 mRNA levels (Fig. 3) and IL-10 secretion in HL60 cells (Fig. 5). This phenotype has been characterized as M2 macrophages, although the classification of human macrophages remains controversial [23]. 9cRA plus 1,25(OH)2D3 did not increase expression of the M1 macrophage gene IL12B in THP-1 and HL60 cells (Fig. 3). Although TNF mRNA levels were not increased, IL6 mRNA expression was effectively induced by the combination of 9cRA and 1,25(OH)2D3 in THP-1 cells. ATRA plus 1,25(OH)2D3 has been reported to induce mRNA and protein levels of tumor necrosis factor (TNF) and IL-6 in U937 cells [43] and to increase expression of inducible nitric oxide synthase (encoded by NOS2) and nitric acid production in U937 cells [44], while we observed no significant change in NOS2 mRNA expression in THP-1 and HL60 cells after treatment with ATRA or 9cRA in combination with 1,25(OH)2D3 (Fig. 3, and data not shown). Tumor necrosis factor (TNF) and IL-6 are cytokines produced from M1 macrophages and NOS2 expression is a M1 macrophage marker [22], [23], [45]. Macrophages with a mixed phenotype expressing both M1 and M2 markers have been identified [46]. Thus, differentiated leukemia cells by 9cRA plus 1,25(OH)2D3 are macrophage-like cells expressing primarily M2 markers with some M1 markers. Further analysis is required to reveal their functional characteristics.
The RAR signaling pathways play an important role in hematopoiesis and granulocytic differentiation [4], [47]. With regard to macrophages, ATRA inhibits TNF production in mouse peritoneal macrophages activated by lipopolysaccharide and interferon γ [48]. ATRA also reduces the synthesis of IL-12 and TNF and enhances IL-10 production in lipopolysaccharide-stimulated human macrophages [49]. Although VDR is dispensable for normal myelopoiesis [50], the vitamin D signaling pathway is involved in the regulation of macrophage/monocyte function [4]. 1,25(OH)2D3 suppresses activation of mouse macrophages by interferon γ [51], and enhances the immunoglobulin- and complement-dependent phagocytosis activity of human blood monocytes [52]. Thus, both ATRA and 1,25(OH)2D3 induce the macrophage/monocyte function common to M2 macrophages. Combined effects of retinoid and 1,25(OH)2D3 on physiological monocyte/macrophage function remain to be elucidated.
In contrast to 9cRA treatment, 1,25(OH)2D3 reduced IL10 mRNA levels and IL-10 production in HL60 cells (Figs. 3 and 5). These findings agree with previous reports that show that 1,25(OH)2D3 suppresses IL10 expression through VDR recruitment to the IL10 promoter in monocytes [53], [54]. Interestingly, 1,25(OH)2D3 enhances IL10 expression of activated human B lymphocytes by recruiting VDR to the IL10 promoter [55]. The combination of 9cRA with 1,25(OH)2D3 effectively induced IL-10 transcription and secretion in THP-1 and HL60 cells (Figs. 3 and 5). Thus, VDR activation induces or suppresses IL10 expression in a manner dependent on cellular conditions. Treatment with 9cRA plus 1,25(OH)2D3 increased CD163 mRNA levels and surface CD163 expression (Figs. 3 and 4). CD163 mediates IL-10 secretion in human monocytes [56]. IL-10 plays an important role in immune regulation by macrophages [46], [57]. Our findings may provide an insight into mechanisms of IL-10 induction.
In conclusion, our results indicate that combined treatment with retinoid and 1,25(OH)2D3 induces differentiation of human myeloid leukemia THP-1 and HL60 cells into macrophage-like cells expressing M2 markers. Further study of human leukemia cell differentiation has the potential to extend differentiation-inducing therapy to the treatment of non-APL myeloid leukemia and to expand the understanding of human macrophage function.
Acknowledgments
The authors thank Dr. Shinya Suzu of Center for AIDS Research, Kumamoto University, Dr. Takayoshi Suganami and Dr. Yoshihiro Ogawa of Tokyo Medical and Dental University for helpful comments, members of the Makishima laboratory for technical assistance and helpful comments, and Dr. Andrew I. Shulman for editorial assistance.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.
Funding Statement
This work was supported by “Strategic Research Base Development” Program for Private Universities subsidized by MEXT (2008–2012) and funds from Nihon University School of Medicine. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Al Tanoury Z, Piskunov A, Rochette-Egly C (2013) Vitamin A and retinoid signaling: genomic and non-genomic effects. J Lipid Res 54: 1761–1775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Wang ZY, Chen Z (2008) Acute promyelocytic leukemia: from highly fatal to highly curable. Blood 111: 2505–2515. [DOI] [PubMed] [Google Scholar]
- 3. de Thé H, Chen Z (2010) Acute promyelocytic leukaemia: novel insights into the mechanisms of cure. Nat Rev Cancer 10: 775–783. [DOI] [PubMed] [Google Scholar]
- 4. Nagy L, Szanto A, Szatmari I, Szeles L (2012) Nuclear hormone receptors enable macrophages and dendritic cells to sense their lipid environment and shape their immune response. Physiol Rev 92: 739–789. [DOI] [PubMed] [Google Scholar]
- 5. Connolly RM, Nguyen NK, Sukumar S (2013) Molecular pathways: current role and future directions of the retinoic acid pathway in cancer prevention and treatment. Clin Cancer Res 19: 1651–1659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Honma Y, Takenaga K, Kasukabe T, Hozumi M (1980) Induction of differentiation of cultured human promyelocytic leukemia cells by retinoids. Biochem Biophys Res Commun 95: 507–512. [DOI] [PubMed] [Google Scholar]
- 7. Breitman TR, Selonick SE, Collins SJ (1980) Induction of differentiation of the human promyelocytic leukemia cell line (HL-60) by retinoic acid. Proc Natl Acad Sci USA 77: 2936–2940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Dalton WJ, Ahearn M, McCredie K, Freireich E, Stass S, et al. (1988) HL-60 cell line was derived from a patient with FAB-M2 and not FAB-M3. Blood 71: 242–247. [PubMed] [Google Scholar]
- 9. Honma Y, Fujita Y, Kasukabe T, Hozumi M, Sampi K, et al. (1983) Induction of differentiation of human acute non-lymphocytic leukemia cells in primary culture by inducers of differentiation of human myeloid leukemia cell line HL-60. Eur J Cancer Clin Oncol 19: 251–261. [DOI] [PubMed] [Google Scholar]
- 10. Brown G, Bunce CM, Rowlands DC, Williams GR (1994) All-trans retinoic acid and 1α,25-dihydroxyvitamin D3 co-operate to promote differentiation of the human promyeloid leukemia cell line HL60 to monocytes. Leukemia 8: 806–815. [PubMed] [Google Scholar]
- 11. Taimi M, Chateau MT, Cabane S, Marti J (1991) Synergistic effect of retinoic acid and 1,25-dihydroxyvitamin D3 on the differentiation of the human monocytic cell line U937. Leuk Res 15: 1145–1152. [DOI] [PubMed] [Google Scholar]
- 12. Makishima M, Kanatani Y, Yamamoto-Yamaguchi Y, Honma Y (1996) Enhancement of activity of 1α,25-dihydroxyvitamin D3 for growth inhibition and differentiation induction of human myelomonocytic leukemia cells by tretinoin tocoferil, an α-tocopherol ester of all-trans retinoic acid. Blood 87: 3384–3394. [PubMed] [Google Scholar]
- 13. Olsson IL, Breitman TR (1982) Induction of differentiation of the human histiocytic lymphoma cell line U-937 by retinoic acid and cyclic adenosine 3′:5′-monophosphate-inducing agents. Cancer Res 42: 3924–3927. [PubMed] [Google Scholar]
- 14. Nagpal S, Na S, Rathnachalam R (2005) Noncalcemic actions of vitamin D receptor ligands. Endocr Rev 26: 662–687. [DOI] [PubMed] [Google Scholar]
- 15. Haussler MR, Whitfield GK, Kaneko I, Haussler CA, Hsieh D, et al. (2013) Molecular mechanisms of vitamin D action. Calcif Tissue Int 92: 77–98. [DOI] [PubMed] [Google Scholar]
- 16. Honma Y, Hozumi M, Abe E, Konno K, Fukushima M, et al. (1983) 1α,25-Dihydroxyvitamin D3 and 1α-hydroxyvitamin D3 prolong survival time of mice inoculated with myeloid leukemia cells. Proc Natl Acad Sci USA 80: 201–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Abe E, Miyaura C, Sakagami H, Takeda M, Konno K, et al. (1981) Differentiation of mouse myeloid leukemia cells induced by 1α,25-dihydroxyvitamin D3 . Proc Natl Acad Sci USA 78: 4990–4994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Mangelsdorf DJ, Koeffler HP, Donaldson CA, Pike JW, Haussler MR (1984) 1,25-Dihydroxyvitamin D3-induced differentiation in a human promyelocytic leukemia cell line (HL-60): receptor-mediated maturation to macrophage-like cells. J Cell Biol 98: 391–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mehta K, Lopez-Berestein G (1986) Expression of tissue transglutaminase in cultured monocytic leukemia (THP-1) cells during differentiation. Cancer Res 46: 1388–1394. [PubMed] [Google Scholar]
- 20. Defacque H, Sevilla C, Piquemal D, Rochette-Egly C, Marti J, et al. (1997) Potentiation of VD-induced monocytic leukemia cell differentiation by retinoids involves both RAR and RXR signaling pathways. Leukemia 11: 221–227. [DOI] [PubMed] [Google Scholar]
- 21.Okamoto R, Koeffler HP (2011) Hematological malignancy. In: Feldman D, Pike JW, Adams JS, editors. Vitamin D. 3 ed. Waltham: Academic Press. 1731–1750.
- 22. Mosser DM, Edwards JP (2008) Exploring the full spectrum of macrophage activation. Nat Rev Immunol 8: 958–969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, et al. (2014) Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity 41: 14–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Hewison M (2012) Vitamin D and immune function: an overview. Proc Nutr Soc 71: 50–61. [DOI] [PubMed] [Google Scholar]
- 25. Kagechika H, Kawachi E, Hashimoto Y, Himi T, Shudo K (1988) Retinobenzoic acids. 1. Structure-activity relationships of aromatic amides with retinoidal activity. J Med Chem 31: 2182–2192. [DOI] [PubMed] [Google Scholar]
- 26. Umemiya H, Fukasawa H, Ebisawa M, Eyrolles L, Kawachi E, et al. (1997) Regulation of retinoidal actions by diazepinylbenzoic acids. Retinoid synergists which activate the RXR-RAR heterodimers. J Med Chem 40: 4222–4234. [DOI] [PubMed] [Google Scholar]
- 27. Makishima M, Shudo K, Honma Y (1999) Greater synergism of retinoic acid receptor (RAR) agonists with vitamin D3 than that of retinoid X receptor (RXR) agonists with regard to growth inhibition and differentiation induction in monoblastic leukemia cells. Biochem Pharmacol 57: 521–529. [DOI] [PubMed] [Google Scholar]
- 28. Amano Y, Cho Y, Matsunawa M, Komiyama K, Makishima M (2009) Increased nuclear expression and transactivation of vitamin D receptor by the cardiotonic steroid bufalin in human myeloid leukemia cells. J Steroid Biochem Mol Biol 114: 144–151. [DOI] [PubMed] [Google Scholar]
- 29. Iriyama N, Yuan B, Hatta Y, Horikoshi A, Yoshino Y, et al. (2012) Granulocyte colony-stimulating factor potentiates differentiation induction by all-trans retinoic acid and arsenic trioxide and enhances arsenic uptake in the acute promyelocytic leukemia cell line HT93A. Oncol Rep 28: 1875–1882. [DOI] [PubMed] [Google Scholar]
- 30. Tavangar K, Hoffman AR, Kraemer FB (1990) A micromethod for the isolation of total RNA from adipose tissue. Anal Biochem 186: 60–63. [DOI] [PubMed] [Google Scholar]
- 31. Sakashita A, Kizaki M, Pakkala S, Schiller G, Tsuruoka N, et al. (1993) 9-cis-retinoic acid: effects on normal and leukemic hematopoiesis in vitro . Blood 81: 1009–1016. [PubMed] [Google Scholar]
- 32. Nakajima H, Kizaki M, Ueno H, Muto A, Takayama N, et al. (1996) All-trans and 9-cis retinoic acid enhance 1,25-dihydroxyvitamin D3-induced monocytic differentiation of U937 cells. Leuk Res 20: 665–676. [DOI] [PubMed] [Google Scholar]
- 33. Kagechika H (2002) Novel synthetic retinoids and separation of the pleiotropic retinoidal activities. Curr Med Chem 9: 591–608. [DOI] [PubMed] [Google Scholar]
- 34. Tobita T, Takeshita A, Kitamura K, Ohnishi K, Yanagi M, et al. (1997) Treatment with a new synthetic retinoid, Am80, of acute promyelocytic leukemia relapsed from complete remission induced by all-trans retinoic acid. Blood 90: 967–973. [PubMed] [Google Scholar]
- 35. Masciulli R, Testa U, Barberi T, Samoggia P, Tritarelli E, et al. (1995) Combined vitamin D3/retinoic acid induction of human promyelocytic cell lines: enhanced phagocytic cell maturation and hybrid granulomonocytic phenotype. Cell Growth Differ 6: 493–503. [PubMed] [Google Scholar]
- 36. Carlberg C, Seuter S, de Mello VDF, Schwab U, Voutilainen S, et al. (2013) Primary vitamin D target genes allow a categorization of possible benefits of vitamin D3 supplementation. PLoS ONE 8: e71042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Bunce CM, Wallington LA, Harrison P, Williams GR, Brown G (1995) Treatment of HL60 cells with various combinations of retinoids and 1α,25 dihydroxyvitamin D3 results in differentiation towards neutrophils or monocytes or a failure to differentiate and apoptosis. Leukemia 9: 410–418. [PubMed] [Google Scholar]
- 38. Evans RM, Mangelsdorf DJ (2014) Nuclear receptors, RXR, and the big bang. Cell 157: 255–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Shulman AI, Larson C, Mangelsdorf DJ, Ranganathan R (2004) Structural determinants of allosteric ligand activation in RXR heterodimers. Cell 116: 417–429. [DOI] [PubMed] [Google Scholar]
- 40. Allenby G, Bocquel MT, Saunders M, Kazmer S, Speck J, et al. (1993) Retinoic acid receptors and retinoid X receptors: interactions with endogenous retinoic acids. Proc Natl Acad Sci USA 90: 30–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Tsao T, Kornblau S, Safe S, Watt JC, Ruvolo V, et al. (2010) Role of peroxisome proliferator-activated receptor-γ and its coactivator DRIP205 in cellular responses to CDDO (RTA-401) in acute myelogenous leukemia. Cancer Res 70: 4949–4960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Sanchez PV, Glantz ST, Scotland S, Kasner MT, Carroll M (2014) Induced differentiation of acute myeloid leukemia cells by activation of retinoid X and liver X receptors. Leukemia 28: 749–760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Taimi M, Defacque H, Commes T, Favero J, Caron E, et al. (1993) Effect of retinoic acid and vitamin D on the expression of interleukin-1β, tumour necrosis factor-α and interleukin-6 in the human monocytic cell line U937. Immunology 79: 229–235. [PMC free article] [PubMed] [Google Scholar]
- 44. Dugas N, Mossalayi MD, Calenda A, Leotard A, Becherel P, et al. (1996) Role of nitric oxide in the anti-tumoral effect of retinoic acid and 1,25-dihydroxyvitamin D3 on human promonocytic leukemic cells. Blood 88: 3528–3534. [PubMed] [Google Scholar]
- 45. Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, et al. (2004) The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol 25: 677–686. [DOI] [PubMed] [Google Scholar]
- 46. Noy R, Pollard JW (2014) Tumor-associated macrophages: from mechanisms to therapy. Immunity 41: 49–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Chanda B, Ditadi A, Iscove NN, Keller G (2013) Retinoic acid signaling is essential for embryonic hematopoietic stem cell development. Cell 155: 215–227. [DOI] [PubMed] [Google Scholar]
- 48. Mehta K, McQueen T, Tucker S, Pandita R, Aggarwal BB (1994) Inhibition by all-trans-retinoic acid of tumor necrosis factor and nitric oxide production by peritoneal macrophages. J Leukoc Biol 55: 336–342. [DOI] [PubMed] [Google Scholar]
- 49. Wang X, Allen C, Ballow M (2007) Retinoic acid enhances the production of IL-10 while reducing the synthesis of IL-12 and TNF-α from LPS-stimulated monocytes/macrophages. J Clin Immunol 27: 193–200. [DOI] [PubMed] [Google Scholar]
- 50. O’Kelly J, Hisatake J, Hisatake Y, Bishop J, Norman A, et al. (2002) Normal myelopoiesis but abnormal T lymphocyte responses in vitamin D receptor knockout mice. J Clin Invest 109: 1091–1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Helming L, Bose J, Ehrchen J, Schiebe S, Frahm T, et al. (2005) 1α,25-Dihydroxyvitamin D3 is a potent suppressor of interferon gamma-mediated macrophage activation. Blood 106: 4351–4358. [DOI] [PubMed] [Google Scholar]
- 52. Xu H, Soruri A, Gieseler RK, Peters JH (1993) 1,25-Dihydroxyvitamin D3 exerts opposing effects to IL-4 on MHC class-II antigen expression, accessory activity, and phagocytosis of human monocytes. Scand J Immunol 38: 535–540. [DOI] [PubMed] [Google Scholar]
- 53. Matilainen JM, Husso T, Toropainen S, Seuter S, Turunen MP, et al. (2010) Primary effect of 1α,25(OH)2D3 on IL-10 expression in monocytes is short-term down-regulation. Biochim Biophys Acta 1803: 1276–1286. [DOI] [PubMed] [Google Scholar]
- 54. Matilainen JM, Rasanen A, Gynther P, Vaisanen S (2010) The genes encoding cytokines IL-2, IL-10 and IL-12B are primary 1α,25(OH)2D3 target genes. J Steroid Biochem Mol Biol 121: 142–145. [DOI] [PubMed] [Google Scholar]
- 55. Heine G, Niesner U, Chang HD, Steinmeyer A, Zugel U, et al. (2008) 1,25-dihydroxyvitamin D3 promotes IL-10 production in human B cells. Eur J Immunol 38: 2210–2218. [DOI] [PubMed] [Google Scholar]
- 56. Philippidis P, Mason JC, Evans BJ, Nadra I, Taylor KM, et al. (2004) Hemoglobin scavenger receptor CD163 mediates interleukin-10 release and heme oxygenase-1 synthesis: antiinflammatory monocyte-macrophage responses in vitro, in resolving skin blisters in vivo, and after cardiopulmonary bypass surgery. Circ Res 94: 119–126. [DOI] [PubMed] [Google Scholar]
- 57. Ng TH, Britton GJ, Hill EV, Verhagen J, Burton BR, et al. (2013) Regulation of adaptive immunity; the role of interleukin-10. Front Immunol 4: 129. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper.