

Abstract
Epstein-Barr virus (EBV) plays a major role in liver pathology. Similar to other members of the herpesvirus family, EBV establishes a persistent infection in more than 90% of adults. The aim of this study was to evaluate the impact of EBV and chronic hepatitis C co-infection (HCV) on biochemical and immunological responses in patients. The study was conducted in 62 patients and 33 apparently healthy controls. Patients were divided into three groups: group I, consisting of 31 patients with chronic hepatitis C infection (CHC), group II, consisting of eight patients with EBV infection and without HCV infection and group III, consisting of 23 patients with EBV and chronic HCV. The percentage of CD3+ cells, helper CD4+ cells and CD19+ B-cells was measured by flow cytometry. Human interferon-γ (IFN-γ) and interleukin (IL)-15 levels were measured by an ELISA. The levels of liver alanine aminotransferase and aspartate aminotransferase enzymes were higher in EBV/HCV patients compared to that in EBV and HCV mono-infected patients. EBV/HCV patients had significantly reduced percentages of CD3+ and CD4+ cells compared to EBV patients. Serum IFN-γ levels were significantly reduced in EBV/HCV patients (3.86 pg/mL) compared to CHC patients (6.76 pg/mL) and normal controls (4.69 pg/mL). A significant increase in serum IL-15 levels was observed in EBV/HCV patients (67.7 pg/mL) compared to EBV patients (29.3 pg/mL). Taken together, these observations suggest that HCV and EBV co-infection can potentiate immune response dampening in patients.
Keywords: Epstein-Barr virus, hepatitis C virus, IFN-γ, IL-15
Epstein-Barr virus (EBV) [human herpes virus (HHV)-4)] infects a narrow range of hosts and replicates slowly. It persists in a latent state in B-lymphocytes and is thought to lead to their immortalisation and malignant transformation (Henry et al. 2013). Similar to other herpes viruses, EBV tends to become latent (Petrova et al. 2010). Primary infection by EBV results in transitional viraemia followed by a powerful T-cell adaptive immune response, which, in immunocompetent subjects, maintains the infection in its latent state (Cohen et al. 2009).
In immunocompetent individuals, the immortalisation of B-lymphocytes is associated with EBV reactivation and the process is conducted by cytotoxic T lymphocytes specific for lytic and latent antigens (Petrova et al. 2010). EBV may also cause illness in immunocompetent people, such as viral hepatitis patients, in whom hepatitis C is the agent accountable for the majority of transfusion-associated hepatitis (Yeung et al. 2007). EBV and hepatitis C co-infection (HCV) leads to higher HCV production than HCV infection alone. It has been reported that the EBV-encoded nuclear antigen 1 (EBNA1) protein of EBV is responsible for higher HCV replication (Sugawara et al. 1999, Palma et al. 2010).
The aim of this study was (i) to investigate the cellular and humoral immune responses to EBV infection in chronic HCV patients by measuring the changes in serum levels of interleukin (IL)-15 and interferon (IFN)-γ and (ii) to study the changes in cellular immunity in these patients through the measurement of the percentage of total T lymphocytes CD3+, CD4+ helper cells and CD19+ B lymphocytes.
SUBJECTS, MATERIALS AND METHODS
Study population - This study (approved by the Ethical Committee of Ain Shams University) included 99 cases collected from Cairo, Egypt (El Demerdash and El Bakri hospitals), Al Qalubia (Banha hospital), Al Menia (El Menia hospital) and Al Monofia (Sheben El Kom hospital) from August 2011-February 2012. The 95 cases included 36 females and 59 males between 18-68 years of age. The subjects were divided into four groups: EBV patients with HCV infection (n = 23), EBV patients without HCV (n = 8), patients with chronic hepatitis C infection (CHC) infection (n = 31) and healthy controls [individuals negative for HCV, human immunodeficiency virus and hepatitis B virus antibodies, as determined from each hospital visit (n = 33)]. Ethics approval was obtained for the study and informed consent forms were signed by patients and healthy controls. Blood samples were drawn from all study participants and serum samples were separated and stored at -80ºC until further testing. Blood samples for lymphocyte subset staining (immunophenotyping) were processed the same day.
Detection of HCV IgG antibodies (Abs) - Abs were assayed in serum samples from all studied subjects using the HCV IgG Abs kit (Diagnostic Automation, Inc, USA).
Detection of HCV RNA by reverse transcription-polymerase chain reaction (RT-PCR) - In all subjects, HCV viraemia was detected by RT-PCR using nested primers from the highly conserved 5’ untranslated region. RNA was extracted from 200-μL serum samples using the acid guanidium thiocyanate-phenol-chloroform method (Chomczynski & Sacchi 1992). Primers used in the detection of HCV RNA were as follow. P1: 5’GGTGCACGGTCTACGAGACCTC3’, P2 forward primer: 5’AACTACTGTCTTCACGCAGAA3’, P3 reverse primer: 5’TGCTCATGGTGCACGGTCTA3’, nested reverse primer P4: 5’ACTCGGCTAGCAGTCTCGCG3’ and nested forward primer P5: 5’GTGCAGCCTCCAG-GACCC3’. All primers were purchased from Promega (Madison, USA). cDNA was synthesised by incubating 10 µL of RNA at 37ºC for 60 min with 20 U of cloned Avian myloblastosis virus reverse transcriptase, 1 × RT-buffer (Qbiogene, USA), 40 units of RNAsin (Clonetech, USA), 0.2 mmol/L each dNTP (Promega, USA) and 10 pmol primer (P1). First round amplification was performed in a total volume of 50 µL using 10 µL of cDNA, 10 pmol of each of the primers P2 and P3, 0.2 mmol/L of each dNTP (Promega), two units of Taq DNA polymerase (Promega) and 1 × Taq buffer. The second round of amplification was similar to the first, except for using the nested primers P4 and P5 and 10 µL of the first round PCR product as template. PCR cycling conditions for both rounds consisted of 30 cycles of 1 min at 94ºC, 1 min at 55ºC and 1 min at 72ºC. The nested PCR products were separated by electrophoresis on a 2% ethidium bromide stained agarose gel and visualised under ultraviolet light.
Serological analysis of EBV infection - Human EBV IgM antibodies were detected in all samples by the qualitative ELISA test using commercially available EBV kits (Diagnostic Automation, USA). EBV-IgG antibodies were detected using commercially available kits (ATLAS Medical EBV-IgG Kit, UK) according to the manufacturer’s instructions. The results of EBV IgM and IgG measurements were expressed as optical density units.
Detection of EBV-DNA - Viral nucleic acid DNA was extracted from 300 μL of serum using the Wizard® DNA purification mini kit (Promega) following the manufacturer’s instructions. For the detection of EBV DNA, nested PCR of the serum samples was performed according to previously established protocols (Kapranos et al. 2003). The 25-μL qualitative PCR reaction mixture contained 2.5 μL of 10x buffer (10 mM Tris-HCl pH 8.0, 50 mM KCl, 25 mM MgCl2), 0.5 μL of 50 mM dNTP mix, 10 pmol of primers E2P1 (5’ATCCTTGCACTTAGCCAAGC3’) and E2P2 (5’TCCAGATGTGTCTCCCTTCT3’) (Bioneer, USA) for the amplification of a 556-bp fragment in the EBNA-2 gene, 5 μL of DNA solution (DNA template), 14.15 μL of distilled water and 0.1 μL (2U) of Taq DNA polymerase (Bioneer). Nested PCR was performed according to the following thermal cycling protocol: pre-denaturation at 94ºC for 5 min, followed by 35 cycles of 94ºC for 30 s, annealing at 57ºC for 30 s and extension at 72ºC for 60 s and final extension at 72ºC for 4 min. The second round of the nested PCR was conducted using the same thermal cycling conditions as described above and by using 2 μL of the first PCR product with the same reaction mixture as mentioned above, except for internal primers AP1: 5’CCAGTAGCATCTCTGTCTGG3’ and AP2: 5’GAACCATCCTCGTCCTCATC3’ (Bioneer) for the amplification of a 190-bp fragment in the EBNA-2 gene. Nested amplification products were visualised by 2% agarose gel electrophoresis and ethidium bromide staining.
L iver enzyme levels - Alanine aminotransferase (ALT) (normal range is up to 40 U/L) and aspartate aminotransferase (AST) (normal range is up to 38 U/L) levels were measured in all samples using commercial kits (Siemens Healthcare Diagnostic Inc, USA) according to the manufacturer instructions.
Measurement of serum cytokine levels - Human IFN-γ and IL-15 were measured using commercially available ELISA kits (Human IFN-γ ELISA kit, Bender MedSystems, Austria, and Human IL-15 ELISA kit, Ray Biotech®, USA) according to the manufacturer’s instructions. The results are presented as the concentration (pg/mL).
Measurement of CD4 + , CD3 + and CD19 + percentages in whole blood by flow cytometry - The percentage of total CD3+ T lymphocytes, CD4+ helper T cells and CD19+ B lymphocytes in whole blood was determined by direct staining with the following conjugated monoclonal antibodies: anti-CD3+ fluorescein isothiocyanate (FITC), anti-CD4+ FITC and anti-CD19+ phycoerythrin. The percentages were determined using a flow cytometer (Coulter® Epics® XLTM, USA) and the data were analysed using system IITM software (Flow cytometry Core Laboratory, El-Demerdash Hospital, Cairo)
Statistical analysis - All statistical analyses were performed using GraphPad InStat statistical software program. Differences were considered significant when p ≤ 0.05.
RESULTS
Based on the HCV-RNA RT-PCR (Fig. 1) and EBV-DNA PCR (Fig. 2) results, the study population was divided into four groups. The first group, representing 32.6% of the total, included 31 HCV infected patients (15 males and 16 females) between 17-63 years of age, with a mean age of 40.03 ± 11.1 years. The second group, representing 8.4% of the total, included eight EBV-infected patients (4 males and 4 females) between 18-58 years of age, with a mean age of 36.8 ± 15.1 years. The third group, representing 24.2% of the total, included 23 cases (18 males and 5 females) with both EBV and HCV infection in the age range of 22-68 years, with a mean age of 50.5 ± 15.1 years. The fourth group, representing 34.7% of the total, included 33 individuals (22 males and 11 females) negative for EBV and HCV infection (normal healthy control group) whose age range was between 18-46 years, with a mean age of 31.1 ± 8.2 years.
Fig. 1. : nested reverse transcription-polymerase chain reaction results of serum samples. Lanes 2, 3, 5, 7: positive for hepatitis C virus (HCV) RNA; 1, 4, 6: negative for HCV RNA.
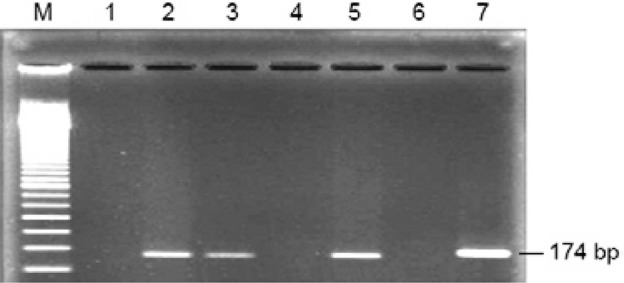
Fig. 2. : nested polymerase chain reaction results of serum samples. Lanes 2, 6-8: positive for Epstein-Barr virus (EBV) DNA; 1, 3-5: negative for EBV DNA.
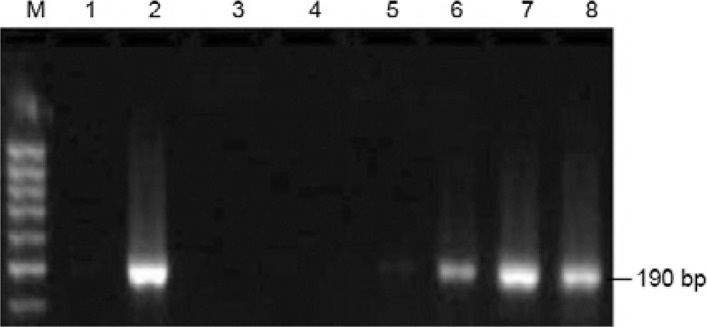
Comparison of liver enzyme levels in the study groups - Serum ALT levels were significantly higher (p < 0.001) in the EBV/HCV group (96.6 ± 10.4 U/mL) and the HCV group (77.1 ± 16.1 U/mL) when compared to the normal controls (NC) (30.5 ± 14.4 U/mL) and somewhat significantly higher (p < 0.01) in the EBV group (52.5 ± 7.7 U/mL) compared to the NC (Fig. 3). However, the increase in AST levels was highly significant (p < 0.001) in the EBV/HCV (93.9 ± 11.39 U/mL), HCV (75.4 ± 16.5 U/mL) and EBV (51.5 ± 8.3 U/mL) patients compared to the NC (29.5 ± 14.2) (Fig. 3).
Fig. 3. : alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activity levels among study subjects. EBV: Epstein-Barr virus; HCV: hepatitis C virus; NC: normal controls.
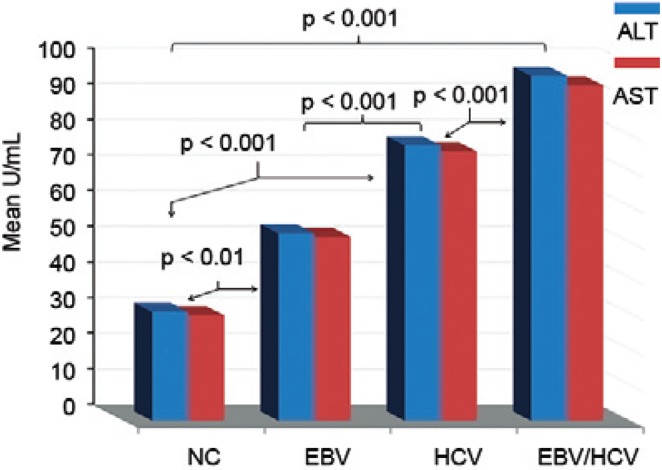
Serum levels of human IL-15 - Serum human IL-15 levels were significantly higher (p < 0.001) in the EBV/HCV (67.8 ± 21.1 pg/mL) and HCV-infected (47.2 ± 7.7 pg/mL) patients compared to the NC group (9.5 ± 11.1 pg/mL). In addition, IL-15 levels were significantly higher in the EBV/HCV patients than in the EBV-infected patients (29.3 ± 17.2 pg/mL). No significant difference was observed between the EBV-infected and HCV-infected patients (Fig. 4).
Fig. 4. : interleukin (IL)-15 activity levels among study subjects. EBV: Epstein-Barr virus; HCV: hepatitis C virus; NC: normal controls.
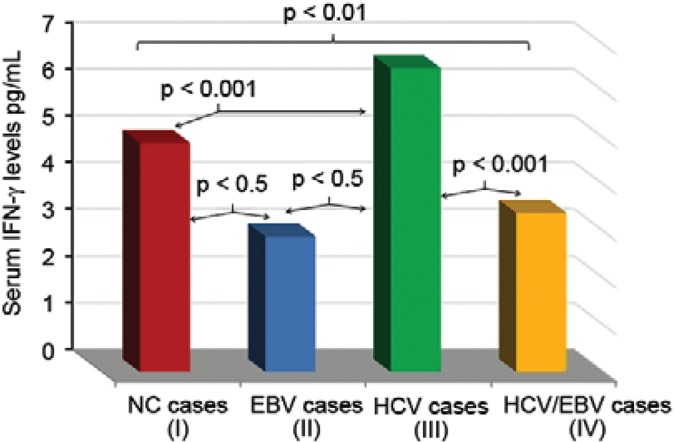
Serum level of human IFN-γ - Human IFN-γ levels in the serum were significantly (p < 0.001) elevated in HCV patients (6.5 ± 2.3 pg/mL) compared to the HCV/EBV patients (3.4 ± 1.01 pg/mL), the EBV patients (2.9 ± 0.8 pg/mL) and the NC (4.9 ± 1.2 pg/mL). IFN-γ serum levels were also significantly lower (p < 0.05) in the EBV and EBV/HCV patients compared to the NC. No significant difference was detected between the EBV and EBV/HCV groups (Fig. 5).
Fig. 5. : interferon-γ (IFN-γ) activity levels among study subjects. EBV: Epstein-Barr virus; HCV: hepatitis C virus; NC: normal controls.
Flow cytometry analysis - The percentage of CD3+, CD4+ and CD19+ lymphocytes was measured by flow cytometry and found to be lower by a statistically significant amount in the EBV/HCV patients compared to the healthy individuals. The mean percentage of total T lymphocytes (CD3+ cells) was significantly lower in the HCV/EBV patients (27.8 ± 11.07) compared to the chronic HCV patients (46.7 ± 4.6), the EBV patients (50.5 ± 4.3) and the NC (65.2 ± 8.1). The percentage of T-helper cells (CD4+ cells) was also significantly lower in the HCV/EBV (15.75 ± 1.3), chronic HCV (25.08 ± 2.4) and EBV patients (32.01 ± 3.01) compared to the NC (43.8 ± 3.6). Additionally, CD19+ B lymphocytes were significantly reduced in number in the HCV/EBV (2.41 ± 1.2), EBV (2.5 ± 1.3) and chronic HCV patients (4.7 ± 1.19) compared to the NC group (8.2 ± 1.5), as shown in Figs 6, 7.
Fig. 6. : examples of flow cytometric histograms of T lymphocytes CD3, helper cells CD4 and B-cells CD19. A: healthy control; B: Epstein-Barr virus (EBV)/hepatitis C virus group; C: EBV group.

Fig. 7. : mean percentage of positive CD3+, CD4+ and CD19+ cells in patients and normal control cases.

DISCUSSION
Virus-to-virus interactions are reported to modify the progression of viral infections in humans (Palma et al. 2010). The classic hepatotropic viruses, hepatitis A through E, are not the only viral agents capable of infecting the liver and causing hepatitis as a part of an organ-specific or systemic involvement with hepatic injury ranging from elevations in aminotransferases to acute hepatitis with or without acute liver failure and fulminant hepatitis. Their ability to cause chronic liver disease has not been fully proven. Cytomegalovirus (CMV), EBV, herpes simplex virus, varicella-zoster virus and adenoviruses have also been shown to be hepatotropic (Gallegos-Orozco et al. 2010). Epstein-Barr viral infection is characterised by alternating periods of latency and reactivation. The reactivation of the virus is observed during periods of immune system down-regulation, such as during drug treatment and illness-related stress or during co-infection with various pathogens (Gandhi et al. 2004, Gredmark et al. 2007).
Consistent with previous reports, our results showed that the activity of liver enzymes (ALT and AST) was significantly increased in the chronic HCV patients (p < 0.001) and in the EBV-infected group (p < 0.01) compared to the control group (Herrine 2002, Hinedi & Koff 2003, Missiha et al. 2008). The observed higher serum ALT and AST levels in the EBV/HCV patients compared to the chronic HCV or EBV mono-infected patients (p < 0.001) suggest a synergistic effect of EBV co-infection on liver cell damage. Similar results have also been reported for chronic HCV patients infected with other HHVs, such as CMV and HHV-6 (Claudio et al. 1999, Petrova et al. 2010).
IL-15 plays an important role in the immune system and is especially important in the activation of the innate and tissue-associated immune responses because it promotes the activation, proliferation and survival of natural killer and CD8+ memory T-cells (Golden-Mason & Rosen 2006). In this study, the highly significant (p < 0.001) increase in serum IL-15 levels in chronic HCV patients compared to the NC indicated the degree of liver tissue damage caused by HCV, which is in agreement with previous reports (Budhu & Wang 2006). These results also indicate the synergetic effect of double infection on IL-15 levels due to increased liver damage (Xu et al. 2000, Ohga et al. 2001, Kimura et al. 2005).
In this study, the highest serum levels of human IFN-γ were observed in CHC cases (p < 0.001). The introduction of EBV infection gradually decreased the IFN-γ levels in EBV/CHC co-infected patients, followed by EBV mono-infected patients. This observed increase in IFN-γ in CHC cases may be due to HCV viral replication and disease progression, as observed in previous reports (Lechmann et al. 1999, Budhu & Wang 2006, Posta et al. 2009). Other studies have reported a reduction in IFN-γ production in EBV-infected cases (Hislop et al. 2007, Saghafian et al. 2013); this reduction in production can account for the gradual decrease in IFN-γ levels upon the introduction of EBV infection. This observation is consistent with our results demonstrating that the highest level of IFN-γ was observed in the CHC group followed by the CHC/EBV co-infected group; the lowest IFN-γ level was observed in the EBV mono-infected cases. EBV infection has evolved multiple mechanisms for disrupting IFN-stimulated Janus kinases (JAK) signal transduction. EBV infection inhibits the IFN-γ response by decreasing the JAK1 protein, which is a necessary component of IFN-γ signalling. Using a variety of such mechanisms, EBV is able to escape antiviral immune responses throughout the primary infection period, as demonstrated by Morrison et al. 2001, Ramana et al. 2002 and Ressing et al. 2008.
In this study, total T lymphocytes (CD3+ cells) were significantly lower (p < 0.05) in CHC patients than in controls. These findings are in agreement with Iken et al. (2006) who observed an unusually low frequency of HCV-specific T-cells in the liver and peripheral blood of CHC patients. Moreover, Ciccaglione et al. (2007) deduced that HCV persistence in most infected individuals is associated with the ability of the virus to evade the host immune response at local and systemic levels and that the virus is capable of replicating in immune cells, effector cells and hepatocytes, which are the main target of HCV replication. Shete et al. (2010) previously reported that the CD3+ T-cell count was lower in EBV-infected patients compared to control individuals. Our results also showed a significant decrease in the percentage of CD4+ cells in the CHC, HCV/EBV, HCV and EBV patients compared to the NC group. These results were consistent with another study by Gonzalez et al. (2008), showing that the CD4+ count and percentage were significantly decreased in HCV infected patients compared to healthy individuals. Mozer-Lisewska et al. (2006) and Harcourt et al. (2006) reported that the percentage of CD4+ cells was decreased in HCV cases compared to control groups, while other studies reported a reduction in CD4+ T cells in patients with EBV-associated Hodgkin’s disease (Karcheva et al. 2008, Shete et al. 2010).
The enumeration of B-cells in this study indicated a significant difference in the percentage of B-cells (CD19+) between the CHC and control groups. Durand et al. (2010) suggested that CD19+ B-cells were significantly reduced in cirrhotic patients with or without hepatocellular carcinoma (HCC). However, HCV induces a powerful antibody response to its envelope glycoprotein E2. Because E2 binds to B-cells via CD81, it is possible that antibodies to E2 block the binding of HCV to cells, thus protecting against HCV infection in some cases (Merani et al. 2011). Once E2 binds to B-cells via CD81, it associates with CD19 and CD21, forming a complex that lowers the activation threshold (Ito et al. 2011). In this study, the percentage of CD19+ cells in HCV/EBV-infected cases was lower than that in the EBV-infected cases, the CHC patients and the controls. The differences in CD19+ counts between these cases were statistically significant. Yang et al. (2005) demonstrated a visible decrease in CD19+ expression in EBV-infected cases compared to normal B lymphocytes. Karcheva et al. (2008) showed that acute EBV infection is characterised by a decrease in CD19 (B-cell) counts.
Previous studies have reported that EBV is responsible for the increased replication of HCV in chronic HCV patients through EBNA1 (Sugawara et al. 1999, Morrison et al. 2001), suggesting that EBV could be involved in the development of HCC (Sugawara et al. 2000, Petrova et al. 2010). This observation suggests an urgent need for a management protocol for EBV/HCV co-infected patients and especially in the context of the study by Bader El-Din et al. (2011), who reported that co-infection with CMV, which is a member of the same family as EBV, complicates the effectiveness of antiviral therapy in the majority of chronic HCV cases.
In conclusion, we recommend introducing EBV treatment in chronic HCV patients to prevent rapid deterioration and to slow the progression to liver cirrhosis and HCC. It is important to measure the efficiency of immunological parameters (CD3+, CD4+ and CD19+ cells) and their related cytokines (IFN-γ and IL-15) in the peripheral blood and serum of HCV-infected patients before, during and after full-term antiviral therapy to assess their prognostic significance.
ACKNOWLEDGEMENTS
To the managers of the hospitals reported in this paper, for their permission to collect blood samples, and to the Department of Microbial Biotechnology, National Research Center, for support to execute this research work.
REFERENCES
- Bader El-Din NG, El-Meguid MA, Tabll AA, Anany MA, Esmat G, Zayed N, Helmy A, El-Zayady AR, Barakat A, El-Awady MK. Human cytomegalovirus infection inhibits response of chronic hepatitis C virus infected patients to interferon-based therapy. J Gastroentero Hepat. 2011:55–62.. doi: 10.1111/j.1440-1746.2010.06319.x. [DOI] [PubMed] [Google Scholar]
- Budhu A, Wang XW. The role of cytokines in hepatocellular carcinoma. J Leukoc Biol. 2006;80:1197–1213.. doi: 10.1189/jlb.0506297. [DOI] [PubMed] [Google Scholar]
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidium-thiocyanatephenol-chloroform extraction. Anal Biochem. 1992;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- Ciccaglione AR, Srellacci E, Marcantonio C, Muto V, Equestre M, Marsilli G, Rapicetta M, Battistini A. Repression of interferon regulatory factor 1 by hepatitis C virus core protein result in inhibition of antiviral and immunomodulatory genes. J Virol. 2007;81:202–214. doi: 10.1128/JVI.01011-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claudio C, Mauro C, Paola P, Marisa M, Anna M, Teresa S, Antonio D. Herpesvirus DNA is frequently detected in liver tissue from hepatitis C patients. J Clin Virol. 1999;14:9–16. doi: 10.1016/s1386-6532(99)00045-1. [DOI] [PubMed] [Google Scholar]
- Cohen JI, Kimura H, Nakamura S, Ko YH, Jaffe ES. Epstein-Barr virus-associated lymphoproliferative disease in non-immunocompromised hosts: a status report and summary of an international meeting, 8-9 September 2008. Ann Oncol. 2009;20:1472–1482. doi: 10.1093/annonc/mdp064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durand TG, Di-Liberto H, Colman E. Occult infection of peripheral B cells by hepatitis C variants which have low translational efficiency in cultured hepatocytes. Gut. 2010;59:934–942. doi: 10.1136/gut.2009.192088. [DOI] [PubMed] [Google Scholar]
- Gallegos-Orozco JF, Rakela-Broder J. Hepatitis viruses: not always what it seems to be. Rev Med Chile. 2010:1302–1311. [PubMed] [Google Scholar]
- Gandhi MK, Tellam JT, Khanna R. Epstein-Barr virus-associated Hodgkin’s lymphoma. Br J Haematol. 2004;125:267–281. doi: 10.1111/j.1365-2141.2004.04902.x. [DOI] [PubMed] [Google Scholar]
- Golden-Mason L, Rosen HR. Natural killer cells: primary target for hepatitis C virus immune evasion strategies? Liver Transplant. 2006;12:363–372. doi: 10.1002/lt.20708. [DOI] [PubMed] [Google Scholar]
- Gonzalez VD, Falconer K, Michaelsson J, Moll M, Rechard O, Alaeus A, Sandberg JK. Expansion of CD56- NK cells in chronic HCV/HIV-1 co-infection: reversion by antiviral treatment with pegylated interferon-gamma and ribavirin. Clin Immunol. 2008;128:46–56. doi: 10.1016/j.clim.2008.03.521. [DOI] [PubMed] [Google Scholar]
- Gredmark S, Jonasson L, Van Gosliga D, Ernerudh J, Soderberg-Naucler C. Active cytomegalovirus replication in patients with coronary disease. Scand Cardiovasc J. 2007;41:230–234. doi: 10.1080/14017430701383755. [DOI] [PubMed] [Google Scholar]
- Harcourt G, Gomperts E, Donfield S, Klenerman P. Profound loss of HCV-specific interferon-gamma secreting CD4+ T-cells in HIV/HCV co-infected patients. Gut. 2006;55:1484–1487. doi: 10.1136/gut.2005.083758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henry H, Balfour JR, Priya V. Primary Epstein–Barr virus infection: impact of age at acquisition, co-infection and viral load. J Infect Dis. 2013;207:1787–1789. doi: 10.1093/infdis/jit096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrine SK. Approach to the patient with chronic hepatitis C virus infection. Ann Intern Med. 2002;136:747–757. doi: 10.7326/0003-4819-136-10-200205210-00010. [DOI] [PubMed] [Google Scholar]
- Hinedi TB, Koff RS. Cholestatic hepatitis induced by Epstein-Barr virus infection in an adult. Dig Dis Sci. 2003;48:539–541. doi: 10.1023/a:1022592801060. [DOI] [PubMed] [Google Scholar]
- Hislop AD, Taylor G, Sauce D, Rickinson AB. Cellular responses to viral infection in humans: lessons from Epstein-Barr virus. Annu Rev Immunol. 2007;25:587–617. doi: 10.1146/annurev.immunol.25.022106.141553. [DOI] [PubMed] [Google Scholar]
- Iken K, Huang L, Bekele H, Schmidt EV, Koziel MJ. Apoptosis of activated CD4+ and CD8+ T-cells in enhanced by co-culture with hepatocytes expressing hepatitis C virus structural proteins through FasL induction. Virol. 2006;346:363–372. doi: 10.1016/j.virol.2005.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito M, Kusunoki H, Mochida K, Yamaguchi K, Mizuochi T. HCV infection and B-cell lymphomagenesis. 835314Adv in Hematol. 20112011 doi: 10.1155/2011/835314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapranos N, Petrakou E, Anastasiadou C, Kotronias D. Detection of herpes simplex virus, cytomegalovirus and Epstein-Barr virus in the semen of men attending an infertility clinic. Fertil Steril. 2003;79(Suppl.):1566–1570. doi: 10.1016/s0015-0282(03)00370-4. [DOI] [PubMed] [Google Scholar]
- Karcheva M, Tz Lukanov, Gecheva S, Slavcheva V, Veleva G, Nachev R. Infectious mononucleosis - Diagnostic potentials. J Imab. 2008;93:431–438. [Google Scholar]
- Kimura H, Hoshino Y, Hara S, Sugaya N, Kawada J, Shibata Y, Kojima S, Nagasaka T, Kuzushima K, Morishima T. Differences between T cell-type and natural killer cell-type chronic active Epstein-Barr virus infection. J Infec Dis. 2005;191:531–539. doi: 10.1086/427239. [DOI] [PubMed] [Google Scholar]
- Lechmann M, Woitas RI, Langhans B, Kaiser R, Ihlenfeldt HG, Jung G, Sauerbruch T, Spengler U. Decreased frequency of HCV core-specific peripheral blood mononuclear cells with type 1 cytokine secretion in chronic hepatitis C. J Hepatol. 1999;31:971–978. doi: 10.1016/s0168-8278(99)80307-9. [DOI] [PubMed] [Google Scholar]
- Merani SD, Petrovic I, James A. Effect of immune pressure on hepatitis C virus evolution: insights from a single-source outbreak. Hepatol. 2011;53:396–405. doi: 10.1002/hep.24076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Missiha SB, Ostrowski M, Heathcote EJ. Disease progression in chronic hepatitis C: modifiable and non-modifiable factors. Gastroenterol. 2008;134:1699–1714. doi: 10.1053/j.gastro.2008.02.069. [DOI] [PubMed] [Google Scholar]
- Morrison TS, Mauser A, Wong A. Inhibition of IFN-γ signaling by EBV immediate-early protein. Immunity. 2001;15:787–797. doi: 10.1016/s1074-7613(01)00226-6. [DOI] [PubMed] [Google Scholar]
- Mozer-Lisewska I, Dworackiy G, Kaczmarekz E, Sluzewski W, Kaczmarky M, Wozniakz A, Zeromskiy J. Significance of alterations in PBMC immunophenotype of children with chronic viral hepatitis C - the role of dendritic cells. Scand J Immun. 2006;63:311–319. doi: 10.1111/j.1365-3083.2006.01741.x. [DOI] [PubMed] [Google Scholar]
- Ohga S, Nomura A, Takada H. Epstein-Barr virus (EBV) load and cytokine gene expression in activated T cells of chronic active EBV infection. J Infect Dis. 2001;183:1–7. doi: 10.1086/317653. [DOI] [PubMed] [Google Scholar]
- Palma T, Doonan PB, Trager NN, Kasman LM. A systematic approach to virus-virus interactions. Virus Res. 2010;149:1–9. doi: 10.1016/j.virusres.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrova M, Kamburov V, Nikolovska D, Kosseva O, Nikolova M, Krastev Z. Epstein-Barr virus: is there any contribution to chronic hepatitis B and C? Liver Int. 2010;30:488–489. doi: 10.1111/j.1478-3231.2009.02138.x. [DOI] [PubMed] [Google Scholar]
- Posta JJ, Ratnarajah S, Lloyd AR. Immunological determinants of the outcomes from primary hepatitis C infection. Cell Mol Life Sci. 2009;66:733–756. doi: 10.1007/s00018-008-8270-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramana CV, Gil MP, Schreiber RD, Stark GR. Stat1-dependent and independent pathways in IFN-gamma-dependent signaling. Trends Immunol. 2002;23:96–101. doi: 10.1016/s1471-4906(01)02118-4. [DOI] [PubMed] [Google Scholar]
- Ressing ME, Horst D, Griffin BD, Tellam J, Zuo J, Khanna R, Rowe M, Wiertz EJ. Epstein-Barr virus evasion of CD8(+) and CD4(+) T cell immunity via concerted actions of multiple gene products. Semin Cancer Biol. 2008;18:397–408. doi: 10.1016/j.semcancer.2008.10.008. [DOI] [PubMed] [Google Scholar]
- Saghafian HS, Sundstro Y, Sohlberg E, Nilsson C, Linde A, Troye BM, Berg L, Sverremark EE. Herpesvirus seropositivity in childhood associates with decreased monocyte-induced NK cell IFN-γ production. J Immunol. 2013;182:2511–2517. doi: 10.4049/jimmunol.0801699. [DOI] [PubMed] [Google Scholar]
- Shete A, Thakar M, Abraham PR, Paranjape R. A review on peripheral blood CD4+ T lymphocyte counts in healthy adult Indians. Ind J Med Res. 2010;132:667–675. [PMC free article] [PubMed] [Google Scholar]
- Sugawara Y, Makuuchi M, Kato N, Shimotohno K, Takada K. Enhancement of hepatitis C virus replication by Epstein-Barr virus-encoded nuclear antigen 1. EMBO. 1999;18:5755–5760. doi: 10.1093/emboj/18.20.5755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugawara Y, Makuuchi M, Takada K. Detection of Epstein-Barr virus DNA in hepatocellular carcinoma tissues from hepatitis C-positive patients. Scand J Gastroenterol. 2000;35:981–984. doi: 10.1080/003655200750023075. [DOI] [PubMed] [Google Scholar]
- Xu J, Ahmad A, Jones JF. Elevated serum transforming growth factor b1 levels in Epstein-Barr virus-associated diseases and their correlation with virus-specific immunoglobulin A (IgA) and IgM. J Virol. 2000;74:2443–2446. doi: 10.1128/jvi.74.5.2443-2446.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang W, Agrawal N, Patel J, Edinger A, Osei E, Thut D, Powers J, Meyerson H. Diminished expression of CD19 in B-cell lymphomas. Cytometry - Part B. Clin Cyto. 2005;63 B:28–35. doi: 10.1002/cyto.b.20030. [DOI] [PubMed] [Google Scholar]
- Yeung LT, To T, King SM, Roberts EA. Spontaneous clearance of childhood hepatitis C virus infection. J Viral Hepat. 2007;14:797–805. doi: 10.1111/j.1365-2893.2007.00873.x. [DOI] [PubMed] [Google Scholar]