

Abstract
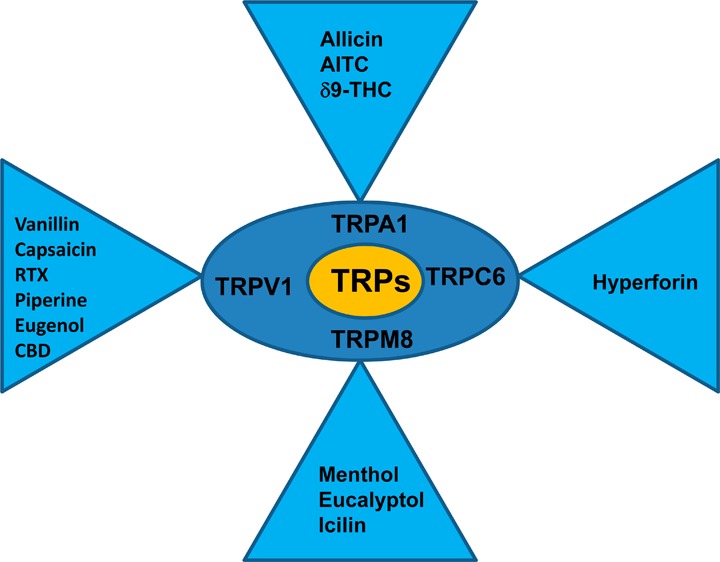
To date, 28 mammalian transient receptor potential (TRP) channels have been cloned and characterized. They are grouped into six subfamilies on the basis of their amino acid sequence homology: TRP Ankyrin (TRPA), TRP Canonical (TRPC), TRP Melastatin (TRPM), TRP Mucolipin (TRPML), TRP Polycystin (TRPP), and TRP Vanilloid (TRPV). Most of the TRP channels are nonselective cation channels expressed on the cell membrane and exhibit variable permeability ratios for Ca2+ versus Na+. They mediate sensory functions (such as vision, nociception, taste transduction, temperature sensation, and pheromone signaling) and homeostatic functions (such as divalent cation flux, hormone release, and osmoregulation). Significant progress has been made in our understanding of the specific roles of these TRP channels and their activation mechanisms. In this Review, the emphasis will be on the activation of TRP channels by phytochemicals that are claimed to exert health benefits. Recent findings complement the anecdotal evidence that some of these phytochemicals have specific receptors and the activation of which is responsible for the physiological effects. Now, the targets for these phytochemicals are being unveiled; a specific hypothesis can be proposed and tested experimentally to infer a scientific validity of the claims of the health benefits. The broader and pressing issues that have to be addressed are related to the quantities of the active ingredients in a given preparation, their bioavailability, metabolism, adverse effects, excretion, and systemic versus local effects.
Keywords: TRP channel, phytochemical, neuropeptide, botanical
David Julius and colleagues from the University of California, San Francisco, cloned the receptor for the active ingredient in hot chili pepper, capsaicin, and named it as vanilloid receptor 1 (VR1) because capsaicin has a vanillyl moiety in its structure.1 It was recognized that the VR1 had a sequence homology to a receptor cloned from a mutant fly (Drosophila melanogaster), in which the electroretinogram exhibited a transient response to continuous light;2 therefore, it was renamed as transient receptor potential (TRP) Vanilloid 1 (TRPV1).3,4 Several TRP channels have been cloned, and some of them are considered as targets for active ingredients in botanicals. For example, TRP Ankyrin 1 (TRPA1), a receptor that carries sensory information from the periphery, is coexpressed with TRPV1, activated by the active ingredient in mustard, allyl isothiocyanate (AITC),5,6 and TRP Melastatin 8 (TRPM8), involved in sensing cold temperatures, is activated by menthol extracted from mint leaves.7,8 Other TRP channels activated by plant ingredients include TRPC6 by hyperforin, TRPV3 by incensole, and TRPM5 indirectly by glucose (sweet taste receptor-mediated increase in intracellular Ca2+). All the subunits of TRP channels that have been cloned so far have six transmembrane domains and a loop between domains five and six, which forms the pore. The channels have a stoichiometry of homo- or heterotetramers. Recently, high resolution structural studies using electron cryomicroscopy have confirmed the tetrameric structure.9,10
Intense research is ongoing to identify the active ingredients in botanicals and their targets to explain the physiological effects they claim to exert. The active ingredient in turmeric is curcumin, which is claimed to be effective in conditions ranging from relieving flatulence to treating Alzheimer’s disease and cancer. However, the bioavailability of curcumin is very low to produce effects systemically either because it is not absorbed or because it is metabolized rapidly by the liver (first-pass metabolism). However, it is important to emphasize that these ingredients can cause significant effects locally in the gastrointestinal (GI) tract. Activation of TRP channels can modulate or promote the release of peptide hormones and neurotransmitters from the sensory and enteric nerve endings and from enteroendocrine cells. It is fascinating to learn that, following a specific type of Bariatric surgery (Roux-en-Y) that involves transposition of the ileum, where the ileum is directly connected to the stomach, unexpectedly, the glucagon-like peptide-1 (GLP-1) levels increased significantly.11 It is inferred from this effect that when food is directly exposed to the ileum, certain ingredients are able to stimulate specialized cells in the lower GI tract (enteroendocrine cells) to cause release of GLP-1. In an intact GI tract, these ingredients may be degraded because of the high acidic environment (pH 2–3) of the stomach and by the gastric enzymes that are released during digestion. This further emphasizes the local effects of phytochemicals in the GI tract, although some of them are not well absorbed. The purpose of this Review is to provide scientific bases for the effects of plant products by identifying the phytochemicals and their TRP channel targets.
Phytochemicals That Activate Transient Receptor Potential Ankyrin (TRPA)
TRPA1 is the only identified member of this family, which is a Ca2+ permeable nonselective cation channel, predominately expressed in a population of sensory neurons that also express TRPV1.5,12 TRPA1 is activated by the phytochemicals such as allyl isothiocyanate (AITC), allicin, diallyldisulfide (DADS), cinnamaldehyde, methylsalicylate, Δ-9-tetrahydrocannabinoid (THC), and synthetic compounds such as icilin, acrolein, N-methylmaleimide (NMM), and (R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinylmethyl)pyrrolo[1,2,3-de)-1,4-benzoxazin-6-yl]-1-apthalenylmethanone (WIN55,212-2).6,13−16 TRPA1 can be activated by multiple products of oxidative stress, which include hydrogen peroxide (H2O2), hydroxyalkenyl aldehyde (4-hydroxynonenal, 4-HNE), and 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2)17,18 (Figure 1). TRPA1 can also be activated by bradykinin (BK).13 Recent studies show that TRPA1 is activated by methylglyoxal (MG).19 MG is formed from triose phosphate during secondary glucose metabolism in hyperglycemic conditions. It is well-known that MG covalently modifies arginine, lysine, and cysteine residues and forms advanced glycation end products.20 TRPA1 has been shown to be involved in various sensory processes, such as detection of noxious cold, mechanosensation, and inflammatory hyperalgesia.5,13,16,21−23
Figure 1.
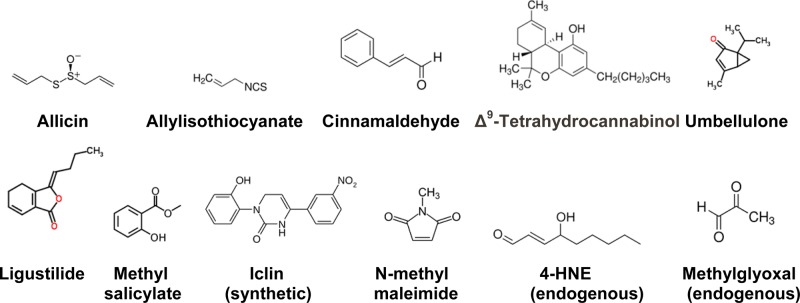
TRPA1 agonists (phytochemicals, synthetic chemicals, and endogenous molecules). Allicin, 2-propene-1-sulfinothioic acid S-2-propenyl ester; allylisothiocyanate (AITC), 3-isothiocyanato-1-propene, is an organosulfur compound; cinnamaldehyde, (2E)-3-phenylprop-2-enal; Δ9-tetrahydrocannabinol, (−)-(6aR,10aR)-6,6,9-trimethyl-3-pentyl-6a,7,8,10a-tetrahydro-6H-benzo[c]chromen-1-ol; umbellulone, 1-isopropyl-4-methylbicyclo[3.1.0]hex-3-en-2-one; ligustilide, (3Z)-3-butylidene-4,5-dihydro-2-benzofuran-1(3H)-one; methyl salicylate, methyl 2-hydroxybenzoate; icilin, 1-(2-hydroxyphenyl)-4-(3-nitrophenyl)-3,6-dihydropyrimidin-2-one; N-methylmaleimide (oxidizing agent); 4-hydroxynonenal, 4-hydroxy-2-nonenal, an α,β-unsaturated hydroxyalkenal produced by lipid peroxidation, is an endogenous agonist; methylglyoxal, an aldehyde from pyruvic acid, acts both as an aldehyde and ketone, and reacts with free amino acids such as lysine, arginine and thiol groups of cysteine. MG is an endogenous agonist.
TRPA1 can be activated by three different mechanisms, including a mechanism of covalent modification of cysteine residues, which is unique among ion channel activation mechanisms: (1) AITC, allicin, DADS, acrolein, and NMM activate the channel by covalent modification of cysteine residues in the cytoplasmic N-terminals; (2) THC and WIN55,212-2 activate the channel possibly by binding to a site; and (3) BK by activating phospholipase C (PLC). Simultaneous mutations of C619, C639, and C663 significantly reduced NMM- and AITC-induced current. It was further demonstrated that additional mutation of K708 prevented the activation by AITC but THC could still activate the channel.23 Cysteine residues are involved in covalent modification, yet the membrane current responses induced by TRPA1 agonists such as AITC and NMM are readily reversible.24
The active ingredients in cinnamon (Cinnamomum zeylanicum), which belongs to the family Lauraceae, are cinnamaldehye, cinnamyl alcohol, and cinnamyl acetate. Cinnamon is a sweet-smelling spice obtained from the bark of the tree. In early days it was used in perfumes, as an appetite stimulant, and to flavor wines. It is considered to improve digestion and acts as an aphrodisiac and is found to be effective in treating sore throat and common cold.
Cinnamaldehyde ((2E)-3-phenylprop-2-enal) is a TRPA1 agonist (Figure 1, EC50 = 100 μM).13 The pungency of cinnamon, when it comes in contact with the tongue, is due to its ability to activate TRPA1 expressed at the nerve terminals. Further, the activation of TRPA1 can cause the release of vasoactive peptides, such as calcitonin gene-related peptide (CGRP) and substance P (SP) from the nerve terminals. It is intriguing that fibers that carry pain sensation also innervate the blood vessels, although the blood vessels are considered to be insensate.25 It is likely that the vasoactive substances released from the nerve terminals have beneficial effects on the cardiovascular functions. Activation of these receptors in the nerve terminals innervating the GI tract sends signals to satiety centers and releases neuropeptides/neurotransmitters locally. It has been shown that cinnamon can decrease blood glucose levels in type 2 diabetes.26,27 Diabetic animals treated with cinnamon showed decrease in blood glucose levels, which could be brought about by the release of incretins (glucose-dependent insulinotropic hormone (GIP) and GLP-1) and insulin release caused by activation of TRPA1 receptors.19
Garlic (Allium sativum) belongs to the family Alliaceae. There are several claims that consumption of garlic imparts good health. Beneficial effects of garlic in fighting common cold, sore throat, and cough have been reported.28,29 Allicin, the active ingredient in garlic, is an organosulfur compound found to have potent antibacterial and antifungal properties30 (Figure 1). Allicin activates TRPA1 and TRPV1.15 It is important to note that allicin has a very short half-life (1–5 s), because it rapidly decomposes. When garlic is crushed, the pungent smell is due to formation of allicin from alliin by the enzyme allicinase. When allicin is degraded, it forms 2-propenesulfenic acid, which can bind to free radicals. Other sulfur compounds present in garlic are ajoene, allyl sulfides, and vinyldithiins.
Anticancer effects of ingredients in garlic have been shown in cell lines and in animal experiments.31 Local application of ingredients in garlic can prevent certain forms of skin cancer.32 The anticancer effects of ingredients in garlic could be due to a local effect of the phytochemicals in the gastrointestinal tract activating TRPA1 and promoting Ca2+ influx, rather than being absorbed and acting systemically. Excessive Ca2+ flux leads to cell death.
Mustard belongs to the family Brassicaceae, genus Brassica and species alba (yellow mustard); or genus Snapis and species nigra (black mustard). Mustard seed contains several ingredients, such as glucosinolates (sinigrin), that can be broken down by the enzyme myrosinase to yield isothiocyanates. The active ingredient in pure mustard oil, AITC, activates TRPA1 and is involved in several functions (see cinnamon and garlic) (Figure 1). While eating mustard-laced food, the effect is more of an olfactory sensation (smell) rather than gustatory sensation (taste). Derivatives of AITC have been used as a war gas. Nitrogen mustard (mechlorethamine) is used as an anticancer agent. As discussed earlier, TRPA1 is a highly Ca2+-permeable channel, the activation of which can cause neuropeptide/neurotransmitter release and can mediate intracellular Ca2+-induced cellular functions.
AITC also is the active phytochemical in horseradish (Armoracia rusticana), which belongs to the family Brassicaceae. It is considered to be pungent; the pungency is due to activation of TRPA1 expressed at the nerve terminals. Wasabi is a common condiment in Japanese cuisine, which has high levels of AITC. When taken orally, it is considered as a remedy for sinusitis, sore throat, and nasal congestion. It is considered to be anthelmintic and bactericidal. Thiopropanal S-oxide is the pungent ingredient in onion (Allium cepa) which belongs to the family Alliaceae. Similar to allicin and AITC, isothiocyanate and thiopropanal S-oxide activate the ion channels TRPA1.33 AITC activates the channel with an EC50 of 33 μM.13 When ingested orally, the bioavailability of AITC is high and is considered as a cancer chemopreventive compound.33
Curcumin ((E,E)-1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione), obtained from turmeric (Curcuma longa), which belongs to the family Zingiberaceae, is an activator of TRPA1.34 The spice comes from the root of the plant and has a bright-yellow color. There are several claimed effects of curcumin, which include anti-inflammatory, antioxidant, anticancer, antidiabetic, antimicrobial, and so forth.35 It has been claimed to be effective in a wide variety of conditions, including flatulence, jaundice, menstrual pain, toothache, and colic. The bioavailability of curcumin is very low. In a study, an oral dose of 8 g resulted in blood levels of around 200 ng.35 However, efforts are being made to improve curcumin bioavailability by various stable preparations, including packaging curcumin in lipid nanoparticles. Longvida is a solid lipid curcumin particle (SLCP) that has a high bioavailability. As discussed earlier, it is possible that turmeric could produce its effects locally by direct contact with the cells in the lumen of the GI tract.
Cystic fibrosis is a condition caused by mutations in the cystic fibrosis transmembrance conductance regulator (CFTR). Curcumin has been shown to be effective in the disease caused by ΔF508 mutation, which results in the production of misfolded CFTR protein.36−38 Curcumin activates TRPA1, which is expressed in the bronchial mucosa and relieves symptoms of cystic fibrosis.
The phytochemical umbellulone is obtained from Umbellularia californica, which belongs to the family Lauraceae (Figure 1). The tree is called “headache tree” because the vapors from the tree can cause severe headache. Umbellulone is a reactive molecule that binds to cysteine residues in TRPA1, thereby activating the receptor (EC50 = 11.6 μM). The headache may be due to activation of TRPA1 in the trigeminal system. Stimulation of sensory nerve terminals causes neuropeptide release. Since CGRP has been shown to play an important role in migraine type headaches by causing vasodilation of the meningeal vessels, it is likely that umbellulone causes headache by causing CGRP release by activating TRPA1.39
Ligustilide, a dihydrophthalide, is a reactive molecule obtained from Angelica acutiloba, which belongs to the family Apiaceae (Figure 1). Ligustilide can bind to thiol groups, and this property could be responsible for activating TRPA1 (EC50 = 44 μM). As this plant ages, it produces dehydroligustilide (EC50 = 539 μM), but this is an antagonist of TRPA1 at lower concentrations (IC50 = 23 μM). The specificity and the reactive nature of these molecules were demonstrated by mutating specific residues required for TRPA1 activation by electrophilic and reactive TRPA1 agonists. Celery contains ligustilide, which could activate TRPA1 and bring about its beneficial gustatory effects.40
Paclitaxel obtained from the Pacific yew (Taxus brevifolia), which belongs to the family Taxaceae, is being used to treat certain forms of cancer (Figure 2). One of the side effects of paclitaxel is peripheral neuropathy that can be explained by its ability to activate TRPA1, which mediates tactile and cold allodynia.41
Figure 2.
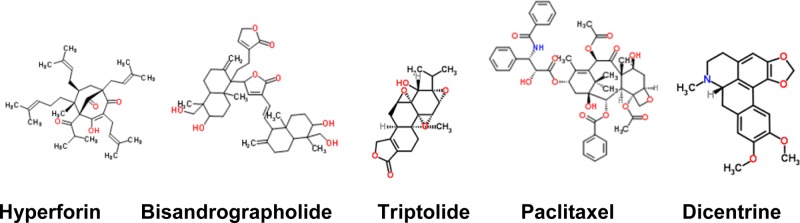
Other TRP channel activators. Hyperforin, (1R,5S,6R,7S)-4-hydroxy-5-isobutyryl-6-methyl-1,3,7-tris(3-methyl-2-buten-1-yl)-6-(4-methyl-3-penten-1-yl)bicyclo[3.3.1]non-3-ene-2,9-dione (TRPC6); bisandrographolide, 3-{(E)-2-[6-hydroxy-5-(hydroxymethyl)-5,8a-dimethyl-2-methylenedecahydro-1-naphthalenyl]vinyl}-5-{6-hydroxy-5-(hydroxymethyl)-5,8a-dimethyl-2-methylene-1-[2-(2-oxo-2,5-dihydro-3-furanyl)ethyl]decahydro-1-naphthalenyl}-2(5H)-furanone (TRPV4); paclitaxel, (2α,5β,7β,10β,13α)-4,10-diacetoxy-13-{[(2R,3S)-3-(benzoylamino)-2-hydroxy-3-phenylpropanoyl]oxy}-1,7-dihydroxy-9-oxo-5,20-epoxytax-11-en-2-yl benzoate (TRPA1, TRPV4); dicentrine, (7aS)-10,11-dimethoxy-7-methyl-6,7,7a,8-tetrahydro-5H-[1,3]benzodioxolo[6,5,4-de]benzo[g]quinoline (TRPA1).
Δ(9)-Tetrahydrocannabinol (THC) is a psychoactive compound in Cannabis sativa, which belongs to the family Cannabaceae (Figure 1). As described earlier TRPA1 is activated by covalent modification of cysteine residues. However, when the cysteine residues were mutated, the activation by NMM was abolished, but THC and WIN55,212-2 could still activate the channel possibly by binding to a site.22 The phytochemicals in cannabis, cannabichromene and cannabigerol, activate TRPA1 with an EC50 of 60 nM and 3.4 μM, respectively. Cannabidiol acid was least potent (EC50 ∼ 12 μM)42 (Table 1).
Table 1. Phytochemicals That Modulate TRP Channels.
| plant name | phytochemical | TRP target | EC50 (μM)a | ref |
|---|---|---|---|---|
| Cinnamomum zeylanicum | cinnamaldehyde | TRPA1 | 6.8 | (6) |
| 61 | (13) | |||
| Allium sativum | allicin | TRPA1 | 7.5 | (22) |
| DADS | TRPA1 | 192 | (22) | |
| Brassica alba | allyl isothiocyanate | TRPA1 | 64.5 | (23) |
| Snapis nigra | 22 | (13) | ||
| 11 | (6) | |||
| Curcuma longa | curcumin | TRPA1 | ND | (34) |
| Umbellularia californica | umbelluline | TRPA1 | 11.6 | (39) |
| Angelica acutiloba | ligustilide | TRPA1 | 44 | (40) |
| dehydroligustilide | TRPA1 | 539 | (40) | |
| Taxus brevifolia | paclitaxel | TRPA1 | ND | (41) |
| TRPV4 | ND | (41) | ||
| Cannabis sativa | tetrahydrocannabinol | TRPA1 | 12 | (6) |
| TRPM8 | 0.1 (ant) | (42) | ||
| cannabidiol | TRPA1 | 12 | (42, 119) | |
| TRPV1 | 3.2 | (42) | ||
| TRPM8 | 0.1 (ant) | (42) | ||
| TRPV3 | 3.7 | (42) | ||
| cannabichromene | TRPA1 | 0.06 | (42) | |
| cannabigerol | TRPM8 | 0.1 (ant) | (42) | |
| TRPA1 | 3.4 | (144) | ||
| cannabidivarin | TRPV4 | 0.9 | (144) | |
| tetrahydrocannabivarin | TRPV3 | 3.7 | (144) | |
| TRPV4 | 6.4 | (144) | ||
| Lindera megaphylla | dicentrine | TRPA1 | ND | (43) |
| Nicotiana tabacum | nicotine | TRPA1 | ∼10 | (44) |
| Zingiberaceae aframomum melegueta | linalool | TRPA1 | 117 | (45) |
| TRPM8 | 6700 | (116) | ||
| α-sanshool | TRPA1 | 69 | (45) | |
| TRPV1 | 1.1 | |||
| shogaol | TRPA1 | 11.2 | (45) | |
| TRPV1 | 0.2 | |||
| paradol | TRPA1 | 71 | (45) | |
| TRPV1 | 1.8 | |||
| Hypericum perforatum | hyperforin | TRPC6 | 0.7 (Na+) | (48) |
| 1.2 (Ca2+) | ||||
| Mentha longifolia | menthol | TRPM8 | 80 | (8) |
| TRPA1 | 68 (ant) | (142) | ||
| Eucalyptus globulus | eucalyptol | TRPM8 | 7700 | (66) |
| Tripterygium wilfordii | triptolide | TRPP | ND | (67) |
| Capsicum annuum | capsaicin | TRPV1 | 0.71 | (1) |
| 0.3 | (119) | |||
| Euphorbia resinifera | Euphorbia resinifera | TRPV1 | 0.04 | (1) |
| Ocimum basilicum | eugenol | TRPV1 | ND | (13, 115) |
| Cinnamonium tamala | TRPV3 | ND | (123) | |
| Artemisia dracunculus | TRPA1 | 262 | (13, 116) | |
| TRPM8 | ND | (13, 116) | ||
| Piper nigrum | piperine | TRPV1 | 38 | (117) |
| Cinnamomum camphora | camphor | TRPV1 | 4500 | (120, 142) |
| Rosmarinus officinalis | TRPA1 | 660 (ant) | (120) | |
| 68 (ant) | (142) | |||
| Euodia ruticarpa | evodiamine | TRPV1 | 0.86 | (121) |
| Zingiber officinale | gingerols | TRPV1 | ND | (13, 122) |
| TRPA1 | ND | (13) | ||
| Origanum vulgare | thymol | TRPV1 | ND | (123) |
| carvacrol | TRPV3 | ND | (123) | |
| TRPA1 | ND | (123) | ||
| Thymus vulgarism | thymol | TRPV3 | ND | (123) |
| Tasmannia lanceolata | polygodial | TRPV1 | ND | (124) |
| Vanilla planifolia | vanillin | TRPV1 | ND | (123, 126) |
| TRPA1 | ND | (123) | ||
| Vernonia tweedieana | α-spinasterol | TRPV1 | 40 (ant) | (127) |
| Boswellia thurifera | incensole | TRPV3 | 16 | (143) |
| Andrographis paniculata | bisandrographolide | TRPV4 | 0.87 | (159) |
(Na+), sodium flux; (Ca2+), calcium flux; (ant), antagonist.
Dicentrine is a naturally occurring aporphine type isoquinoline alkaloid, isolated from the root Lindera megaphylla Hemsl., which belongs to the Lauraceae family (Figure 2). In animal models, dicentrine induced antinociceptive effects. Cinnamaldehyde-induced nocifensive behavior was abolished by dicentrine, but not the capsaicin-induced nocifensive behavior. Based on these studies, it is proposed that the dicentrine effect may involve interaction with TRPA1 channels.43
Nicotine is obtained from Nicotiana tabacum of the Solanaceae family. It brings about its stimulatory actions by activating neuronal nicotinic acetylcholine receptors. However, it also produces irritation while smoking, chewing, or snorting. It has been shown that irritation is caused by the activation of TRPA1. Nicotine activates TRPA1 in lower concentrations (EC50 ∼ 10 μM), but inhibits at higher concentrations (>1 mM).44
Extracts of Sichuan and melegueta peppers evoke pungent sensations that are mediated by different alkylamides, such as sanshool and shogaol; both activate TRPA1 and TRPV1 channels. Linalool, a terpene in Sichuan peppers, is able to activate TRPA1 but not TRPV145 (Table 1).
Phytochemicals That Activate Transient Receptor Potential Canonical (TRPC)
TRPC channels have been classified as TRPC1, TRPC2, TRPC3/6/7 and TRPC4/5 on the basis of structural similarities and functions.46 The channel is formed as a homo- or heterotetramer. The activation mechanism of TRPC channels is not fully clarified. These channels are associated with G-protein coupled receptors and activation of G-protein coupled receptor results in transactivation of TRPC channels and facilitates their openings.47
St. John’s wort (Hypericum perforatum) extract is used as an antidepressant, and the mechanism of action is still elusive. One of the active ingredients in the extract has been found to be hyperforin, a bicyclic polyprenylated acylphloroglucinol compound (Figure 2). Hyperforin has been shown to activate TRPC6.48 In general, antidepressants are selective serotonin and norepinephrine uptake inhibitors, thereby increasing the levels of serotonin and norepinephrine. The neurotransmitter levels can also be increased by promoting their release by causing Ca2+ influx at the nerve terminals. Generally, neurotransmitter release occurs in response to an action potential arriving at the nerve terminal and activating voltage-gated Ca2+ channels. It is becoming increasingly apparent that Ca2+ permeable TRP channels expressed at the presynaptic terminals can cause transmitter release and modulate synaptic transmission, independent of action potentials. TRPV1 and TRPA1 expressed in the presynaptic terminals of the sensory neurons can cause neurotransmitter release.49−52 The effect of hyperforin could be due to its activation of TRPC6 expressed in central neurons. The effect appears to be specific because TRPC3 is unaffected by hyperforin. TRPC6, like most TRP channels, is a nonspecific cation channel that has a high Ca2+ permeability. Hyperforin also exhibits neurotrophic effects leading to axonal sprouting and neurite extension. In confirmation of this effect, neurons from TRPC6 overexpressing animals exhibit enhanced dendritic growth and synapse formation that may play a role in learning and memory formation.53
Phytochemicals That Activate Transient Receptor Potential Melastatin (TRPM)
There are eight members in TRPM family. There are no ankyrin domains in the N-terminus of these channels. These channels are Ca2+ and Mg2+ permeable, and the permeability to Ca2+ ranges from impermeable (TRPM4 and TRPM5) to significantly Ca2+ permeable (TRPM6 and TRPM7).54
TRPM5
TRPM5 is a channel activated by increases in intracellular Ca2+. TRPM5 activation by sweet tastants is by an indirect mechanism of causing an increase in intracellular Ca2+ levels by the phytochemicals activating the sweet-taste receptor. Several structurally diverse phytochemicals have been shown to activate the sweet-taste receptor. But the degree of sweetness differs; glucose is less sweet in comparison to sucrose, which is a disaccharide formed by the combination of fructose and glucose. Fructose is the sweetest (73% sweeter than glucose). The plant product stevioside, obtained from Stevia rebaudiana, activates the sweet receptors. Interestingly, although the whole-plant extract can be used as a sweetening agent, it has been shown to cause infertility. However, the pure ingredient from this plant, rebaudioside A, is devoid of this action.55
The type 2 taste receptor (T2R) is alpha-gustducin, which is a sweet- and bitter-taste receptor. A knockout of the alpha-gustducin gene causes animals to lose both sweet and bitter taste sensations. T2Rs sense bitter taste. For umami and sweet taste perception, taste receptors type 1 (T1R1), type 2 (T1R2), and type 3 (T1R3) form heterodimers. T1R1 and T1R3 form a complex to sense the umami taste. T1R2 and T1R3 form a complex to taste sweetness. The signal transduction involves the activation of the G-protein-coupled receptor alpha-gustducin, which is coupled to phospholipase Cβ 2 (PLCβ 2). Activation of PLCβ 2 promotes the hydrolysis of PIP2 to form IP3 and DAG. IP3 releases Ca2+ from intracellular stores and activates TRPM5, which depolarizes the cell and causes ATP release. ATP acts as a neurotransmitter and mediates signal transduction.56
It is becoming evident that the receptors that sense sweet-taste on the tongue are also expressed throughout the GI tract and act as chemosensors. Taste receptors are present in the cells lining the stomach, pancreas, and enteroendocrine cells of the GI tract. Stimulation of brush border cells with tastants releases GLP-1 and peptide YY (PYY). Both alpha-gustducin and TRPM5 receptors are predominately expressed in these cells. It is possible that phytochemicals are able to activate these receptors and promote release of neuropeptides, neurotransmitters and hormones.57−60
TRPM8
Mint (Mentha longifolia) belongs to the family Lamiaceae, and peppermint belongs to the species piperita. The decoction of mint leaves is used for stomach aches and for some painful conditions. The active ingredient is menthol (Figure 3); it is used as an ingredient in various balms that are used to relieve pain.
Figure 3.
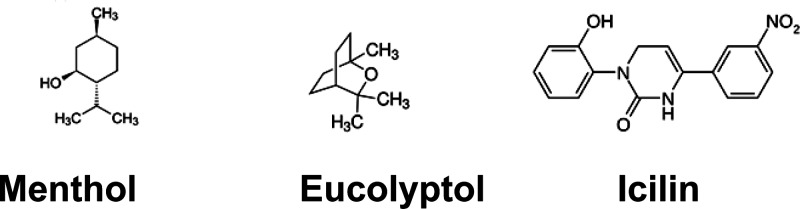
TRPM8 agonists. Menthol, (1R,2S,5R)-2-isopropyl-5-methylcyclohexanol; eucalyptol, 1,3,3-trimethyl-2-oxabicyclo[2.2.2]octane; icilin, 1-(2-hydroxyphenyl)-4-(3-nitrophenyl)-3,6-dihydropyrimidin-2-one.
Transient receptor potential melastatin 8 (TRPM8), previously known as menthol and cold receptor 1 (CMR1), is a Ca2+ permeant nonspecific cation channel, which is expressed in a subpopulation of primary afferent neurons. TRPM8 is activated by cold (<25 °C), phytochemicals such as menthol and eucalyptol, and the synthetic chemical icilin. Activation of TRPM8 induces a cool/soothing sensation.7,8 TRPM8 expressed at the central terminals modulates synaptic transmission.61,62 TRPM8 (earlier identified as Trp-p8) is upregulated in prostate cancer and is involved in urinary bladder functions, which broadens the horizon of the involvement of TRPM8 in other physiological and pathophysiological conditions.63
Generally, phosphorylation enhances the activity of ion channels. However, it has been demonstrated that a functional downregulation of TRPM8 occurs when PKC is stimulated resulting in an inhibition of TRPM8-mediated channel activity, in contrast to TRPV1, which is robustly potentiated by PKC activation. These effects are due to dephosphorylation of TRPM8 by activation of protein phosphatases.61
The activation of TRPM8 sends the cool and soothing sensation to alleviate pain. Therefore, it is expected to be upregulated by phosphorylation in inflammatory conditions. Paradoxically, phosphorylation downregulates TRPM8, thereby compromising the much needed cool and soothing sensation.
It has also been shown that mentholated cigarette smoke exerts a cool and soothing sensation while inhaling; because of this, mentholated cigarettes may encourage the smoking habit. Recently, the United States Food and Drug Administration has issued a warning that mentholated cigarettes are more addictive.64 Further, menthol can directly interact with the nicotinic acetylcholine receptor and inhibit its function, a mechanism that may explain the reason for smoking a greater number of cigarettes to get the same effect, thereby increasing the addictive potential of nicotine.65
Eucalyptol is obtained from the leaves of Eucalyptus globulus in an oil form (Figure 3). It has the structure of a cyclic ether and a monoterpenoid. It has a smell resembling that of camphor. It is added as one of the additives in cigarettes. Eucalyptol is a TRPM8 agonist (EC50 = 7.7 mM).66
Phytochemicals That Activate Transient Receptor Potential Polycystin (TRPP)
The TRPP family is made up of three channel members, namely, TRPP1, TRPP2, and TRPP3. TRPP1 is an ion channel, which is considered to be involved in polycystic kidney disease. The disease is characterized by the formation of multiple cysts, hence the name polycystic kidney disease, eventually leading to kidney failure. In this disease, cysts are also found in liver, pancreas, and other inner surfaces covered by tubular epithelial cells. In tubular epithelial cells, ciliary action transduces a mechanical stimulus and opens a Ca2+ permeable ion channel, such as polycystin-2 (PC2 or TRPP1), and increases the intracellular Ca2+ levels and causes cell cycle arrest. Mutations in TRPP and/or the associated protein, polycystin 1 (PC1), result in autosomal dominant polycystic kidney disease (ADPKD).
Triptolide, a diterpene (Figure 2) obtained from Tripterygium wilfordii, induces Ca2+ influx in tubular epithelial cells and controls their proliferation, resulting in reduced cyst formation and alleviation of symptoms associated with kidney damage in a murine model of ADPKD.67
Phytochemicals That Activate Transient Receptor Potential Vanilloid (TRPV)
There are six members in the TRPV family, the name is derived by the activation of TRP Vanilloid 1 (TRPV1) by molecules consisting of a vanillyl moiety. The other members of this family are TRPV2, TRPV3, TRPV4, TRPV5, and TRPV6.68−70 While comparing the Ca2+ permeability, it has become clear that TRPV5 and TRPV6 channels are purely Ca2+ permeable. They form homo- or heterotetrameric structures.71 The high resolution structure of TRPV1 has been recently published, aided by electron cryomicroscopy.9,10
TRVP1
TRPV1, formerly known as vanilloid receptor 1 (VR1), is a nonselective cation channel with high Ca2+ permeability, which is expressed predominantly in a population of small-diameter sensory neurons. It functions as a polymodal receptor in the peripheral sensory nerve terminals and modulates synaptic transmission at the first sensory synapse.1,49,51,72−74 A recent study has shown its expression in the dorsal horn inhibitory interneurons.75 Capsaicin has been shown to modulate synaptic transmission in other brain regions.76−80 Activation of TRPV1 results in two functional components: (1) sending the impulses to the brain by generating an action potential at the nerve endings (afferent function) and (2) releasing vaso/neuroactive substances by virtue of its Ca2+ permeability, such as histamine, bradykinin, CGRP, and SP, from the peripheral nerve terminals (efferent action).
TRPV1 is activated by heat (>42 °C) and phytochemicals such as capsaicin, resiniferatoxin (RTX), tinyatoxin (TNX), camphor, carvacrol, and thymol. It is activated by endogenous ligands such as protons, anandamide, arachidonic acid metabolites, and N-arachidonyl dopamine (NADA).1,27,49,54,68,81−86
Although TRPV1 is considered mainly to be involved in thermal sensory perception, its distribution in regions that are not exposed to its activation temperature ranges raises the possibility of its involvement in other functions. TRPV1 can be detected using RT-PCR and radioligand binding throughout the neuroaxis, and the identification of specific ligands such as NADA in certain brain regions further suggests its roles in the CNS.51,75,87−90 TRPV1 is present in the smooth muscles of the blood vessels and bronchi, where activation of the receptor leads to vasodilation by releasing CGRP, acetylcholine, or nitric oxide from nerve terminals and bronchoconstriction by promoting Ca2+ influx, respectively.91−93 TRPV1 is found in the nerve terminals, supplying the urinary bladder and the urothelium, indicating a role in urinary bladder function, such as micturition.94,95 Interestingly, TRPV1 is also involved in the regulation of body temperature. Subcutaneous injection of capsaicin decreases the body temperature by 2–3 °C. TRPV1 antagonists increase the body temperature to the same extent.96,97
TRPV1 has emerged as a potential target for developing analgesics. Potent TRPV1 antagonists have been developed and shown to be effective in alleviating pain in several animal models. Unfortunately, development of hyperthermia following their administration has halted the clinical trials.94 However, findings suggest that certain compounds may be devoid of the effect of elevating core body temperature.98
Capsaicin (8-methyl-N-vanillyl-6-nonenamide) (Figure 4), an active ingredient in hot chili pepper (Capsicum annuum or frutescens), which belongs to the family Solanaceae. The nonpungent bell peppers belong to the species annuum, and hot peppers belong to the species frutescens.
Figure 4.
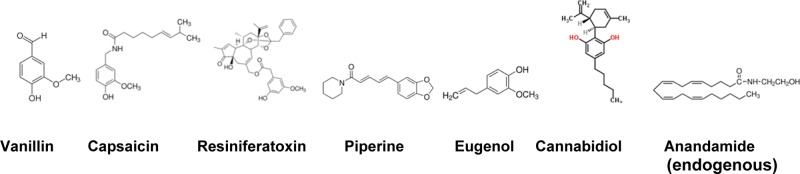
TRPV1 agonists. Vanillin, has the vanillyl moiety, that is essential for activating TRPV1 channels; Capsaicin, (E)-N-[(4-hydroxy-3-methoxyphenyl)methyl]-8-methyl-6-nonenamide, has a vanilloid and an aceyl moiety; dihydrocapsaicin, the structure of which is a 6,7-dihydro derivative of capsaicin; resiniferatoxin, has a complex structure, but shares a homovanillyl group, which is necessary for the activity of all vanilloids; eugenol, 2-methoxy-4-(2-propenyl)phenol and is a member of the allylbenzene class of chemical compounds; Cannabidiol, 2-[(1R,6R)-6-isopropenyl-3-methyl-3-cyclohexen-1-yl]-5-pentyl-1,3-benzenediol; anandamide or arachidonylethanolamide or arachidonic acid N-(hydroxyethyl)amide consists of the acyl moiety and is an edogenous ligand of TRPV1 and cannabinoid receptor 1 (CB1).
The hotness of chili peppers is due to the chemical content of capsaicin. Purified capsaciin activated TRPV1 with a EC50 of 711.9 nM.1 Other capsinoids such as capsiate, dihydrocapsiate, and nordihydrocapsiate have been isolated and purified. The degree of hotness can be quantified by using the Scoville scale. A Scoville unit is the “number” of times the alcoholic extract has to be diluted to lose the pungency. A sweet bell pepper has a score of 0 Scoville units and the Habanero, Savina, and Naga Jolokia (ghost peppers) are calibrated to have scores of 400 000, 600 000, and 1 000 000 Scoville units, respectively. For example, ghost pepper extract has to be diluted one million times to lose its pungency. Capsaicin and dihydrocapsaicin have the Scoville scores of 16 000 000 and 15 000 000, respectively. Shogaol from ginger has the score of 160 000 units, piperine from pepper has the score of 100 000 units, and gingerol from ginger has a score of 60 000 units. Commercially available pepper spray has the score of 2 500 000 units, and police-grade pepper spray has the score of 5 500 000 Scoville units.
When capsaicin binds to its receptor, the ion channel opens, but when constantly activated, the receptor enters a desensitization state. On the other hand, sensitization is a phenomenon, where the receptor activity is enhanced by phosphorylation. Overexpression and overactivation of TRPV1 is observed in various painful conditions. Topical capsaicin application has been useful to treat conditions such as arthritis, diabetic peripheral neuropathy, shingles, and psoriasis by exerting a local effect.85,86,99,100 The mechanism of pain relief has been proposed to be due to desensitization of the receptor or degeneration/ablation of the nerve terminals.101
Altered expression of TRPV1 is found in cancers involving prostate, bladder, pancreas, tongue, skin, liver, and colon. Capsaicin can induce its effects by causing Ca2+ influx through TRPV1 overexpressed in cancerous cells, which can lead to cell death by apoptosis or necrosis.85
In diabetes, consuming a hot chili pepper containing meal showed a decrease in the amount of insulin required to combat the postprandial increase in glucose. TRPV1 may play a role in this effect.102,103 Capsaicin has been reported to increase oxygen consumption and thermogenesis, which might lead to weight loss. There are studies to support a potential neurogenic mechanism by which TRPV1-sensitive sensory neurons may regulate energy and fat metabolism. Capsaicin prevents adipogenesis by apoptotic mechanism. Capsaicin or N-oleoylethanolamide, an endogenous ligand of TRPV1, reduced food intake by conveying information through the vagus nerve and affecting satiety centers. TRPV1 knockout animals, when fed a high-fat diet, although the food intake was the same, they gained less weight as compared to their wild-type counterparts.104,105
Although there is no direct evidence that hot chili pepper containing spicy food increases acid secretion in the stomach, in certain conditions, such as gastroesophageal reflux disease (GERD), increased expression of TRPV1 in the esophagus can induce a burning sensation.106 Capsaicin has been shown to worsen the condition in patients with irritable bowel syndrome (IBS) and Crohn’s disease. Activation of TRPV1 has been shown to cause the release of gastric acid in the stomach, but other studies have reported otherwise.107
Urinary bladder hyperreflexia is a condition that has been shown to be related to overexpression of TRPV1 in the nerve terminals innervating the bladder. Excretion of capsaicin through the kidneys can accumulate in the bladder and exert an effect on the urinary bladder.85
Resiniferatoxin (RTX) and tinyatoxin (TNX) are the most potent among all the known natural, synthetic, and endogenous agonists of TRPV1. These pure chemicals obtained from a cactus-like spurge Euphorbia resinifera/poissonii. Purified RTX activates TRPV1 with an EC50 of 39.1 nM.1 In fact, RTX can maximally activate single channel TRPV1 currents in picomolar ranges.108 RTX/TNX have the Scoville scores of 16 000 000 000 and 5 300 000 000, respectively. RTX, a phorbol related diterpene (resiniferonol 9,13,14-orthophenylacetate 20-homovanillate), has a complex structure with a phorbol and a vanillyl moiety (Figure 4). Its ultrapotency was thought to be due to the phorbol moiety, which could activate protein kinase C (PKC) and promote phosphorylation of TRPV1. This notion was abandoned because of the higher concentrations of RTX required to activate PKC. The tritiated form ([3H]RTX) has been used as a tool in ligand-binding assays.68,109 Binding of capsaicin and RTX to TRPV1 involves amino acid residues, which have been shown to reside within N- and C-cytosolic and transmembrane domains of the channel.110−112 RTX could induce nerve terminal ablation by sustained Ca2+ influx and prevent nociceptive transmission. Intrathecal administation of RTX provides a long-lasting pain relief.51,113,114 A clinical trial is ongoing to determine the effectiveness of intrathecal administation of RTX in debilitating terminal cancer pain conditions (NCT00804154). Intravesicular irrigation of RTX containing solution has yielded a significant improvement in urinary bladder hyperreflexia by its ability to ablate TRPV1 expressing nerve terminals in the urinary bladder.95 Resiniferatoxin and tinyatoxin are used as pesticides.
Basil (Ocimum basilicum) belongs to the family Lamiaceae. Basil and oregano have the ingredient β-caryophyllene, a natural bicyclic sesquiterpene, which is an agonist of cannabinoid receptor 2 (CB2) and has been shown to be involved in anti-inflammatory actions. Eugenol, a phenylpropene and an allyl chain-substituted guaiacol is one of the active ingredients in basil and clove (Figure 4). Other ingredients include citral that provides the citrus smell to basil, camphene in the African blue basil, and anethole in licorice and basil. Eugenol has anti-inflammatory properties by blocking the cyclooxygenase enzyme. Eugenol activates TRPV1 and TRPV3115 (Table 1).
Bay leaves (Cinnamonium tamala) belong to the family Lauraceae. The active ingredients include β-caryophyllene, eugenol, and linalool, a naturally occurring terpene alcohol found in many flowers and spice plants. As discussed above, eugenol can activate TRPV1 and TRPV3 ion channels and linalool can activate TRPA1.116
Black pepper (Piper nigrum) belongs to the family Piperaceae. The active ingredients have been isolated. The main ingredient is piperine. Other alkaloids present in black pepper include chavicine and piperidine (Figure 3). Piperine activates TRPV1.117 The pungency of pepper is due to the alkaloid piperine and is quantified to have a score of 100 000 Scoville units as compared to the ghost pepper, which has a score of 1 000 000 Scoville units. It is also used to treat conditions such as sore throat and bronchitis. It improves digestion and acts as a carminative. Piperine is known to inhibit the liver metabolizing enzyme CYP3A4, thereby increasing the bioavailability of other drugs. Piperine has been shown to significantly increase the bioavailability of curcumin by interfering with its metabolism.118
As discussed earlier, several ingredients in Cannabis sativa can activate TRP channels.119 Cannabidiol (CBD) does not exhibit any psychotropic effects. Recently, CBD has gained attention because of its effectiveness in treating refractory epilepsies in children. CBD and other active ingredients in cannabis are considered as activators of TRPV1 (Figure 4). CBD activates TRPV1 with EC50 of 3.2 μM as compared to activation by capsaicin (EC50 of 0.3–0.7 μM).1,119
Camphor, a terpenoid, is a transparent solid obtained from an evergreen tree Cinnamomum camphora. Another source of camphor is from dried rosemary (Rosmarinus officinalis). Camphor is an activator of TRPV1.120
Clove (Eugenia caryophyllis or Syzgium aromaticum) belongs to the family Myrtaceae. Clove oil is commonly used to treat toothaches and used as a local anesthetic in dental procedures. The principal active ingredients in clove are eugenol and salicylic acid. The effects of both of these active ingredients are useful in painful conditions. Eugenol activates TRPV1 and TRPV3 channels.115
Evodiamine, an active ingredient from the Rutadeae family of plants (Euodia ruticarpa), is a TRPV1 agonist. It is being used as a dietary supplement; it has been shown to induce thermogenesis.121
Ginger (Zingiber officinale) belongs to the family Zingiberaceae. The active ingredients in ginger are gingerols, shogaols, and zingiberene. They exist in different forms (6, 8, 10) depending on the alkyl carbon chain. The pungency is quantified to be 60 000 Scoville units. Structurally, it is related to capsaicin and piperine. Heat converts gingerol to zingerone. When gingerol is dehydrated, it is converted to a more potent compound, shogaol (EC50 = 0.2 μM; 160 000 Scoville units). All these ingredients can activate TRPV1 and TRPA1.122 The alcoholic extract has been shown to possess antioxidant properties.
Oregano (Origanum vulgare) belongs to the family Lamiaceae. It contains several phytochemicals, such as thymol, carvacrol, and rosmarinic acid. The antioxidant properties are stronger than those of synthetic antioxidants, such as butylated hydroxytoluene (BHT) and butylated hydroxyanisole (BHA). Thymol and carvacrol can activate TRPV1 and TRPV3 ion channels123 (Table 1).
Polygodial, a drimane-type sesquiterpene dialdehyde, is an active ingredient obtained from the Dorrigo pepper (Tasmannia lanceolata). Polygodial activates TRPV1 and TRPA1.124,125
Tarragon (Artemisia dracunculus) is a perennial herb that belongs to the family Asteraceae. The active ingredient that is responsible for the characteristic taste is considered to be cis-pellitorin. The ingredients in tarragon oil include methyl eugenol (36%) and methyl chavicol (16%). Eugenol is a TRPV1 and TRPV3 channel agonist.115
Thyme (Thymus vulgarism) belongs to the family Lamiaceae. The major ingredient in thyme oil is thymol; other ingredients are borneol, linalool, myrcene, and p-cymene. It is incorporated as an antiseptic in mouthwash and in toothpaste. Thymol and linalool have been shown to activate TRPV3 and TRPA1, respectively.45,116,123
Vanilla (Vanilla planifolia) belongs to the family Orchidaceae (Figure 4). The smell is due to the active ingredient vanillin, which is structurally related to eugenol or guaiacol. It is used for flavoring ice creams, confectionaries, tobacco, beverages, and so forth. The natural flavor is due to vanillic aldehyde, but the artificial flavor ethylvanillin is as potent as the natural vanilla. It has mild CNS effects, is regarded as an aphrodisiac, and is useful to treat impotence. Vanillin can activate TRPV1 and TRPV3 ion channels.126
Most phytochemicals have been shown to activate TRP channels. The active phytochemical in the dichloromethane fraction from the leaves of the medicinal plant Vernonia tweedieana that belongs to the family Asteraceae was identified to be α-spinasterol, which acts as a potent antagonist of TRPV1. The antagonistic effects were demonstrated by the displacement of tritiated RTX ([3H]RTX) and inhibition of capsaicin-induced Ca2+ influx. α-Spinasterol exhibited an antinociceptive effect to noxious heat, but the mechanical threshold was unaffected. The specific action involving TRPV1 was confirmed by the lack of antinociceptive effect in mice systemically treated with RTX, which is known to ablate TRPV1 expressing neurons. Its effectiveness was demonstrated by its ability to reduce inflammatory hypersensitivity induced by complete Freund’s adjuvant (CFA). Interestingly, the body temperature was unaffected by its antagonistic action.127
TRPV3
TRPV3 is a thermosensitive channel that has a high sequence homology with TRPV1. TRPV3 was initially found to be exclusively expressed in the keratinocytes; however, further studies have shown its expression in sensory and central neurons, nasal mucosa, tongue, kidney, and testis. TRPV3 expression has been shown to be enhanced in painful conditions and is being pursued as a target for developing analgesics.129−135
Nerve terminals of the sensory neurons in the periphery are surrounded by keratinocytes; therefore substances such as ATP, prostaglandins, and nerve growth factor released from keratinocytes can make the nerve terminals more excitable.136,137 TRPV3 knockout animals lacked ATP release from peripheral terminals.138 Activation of PLC has been shown to modulate the function of TRPV3 by PIP2-mediated mechanism and by IP3-mediated increase in intracellular Ca2+ levels.138,139 Overexpression of TRPV3 in animals results in a “hairless” phenotype, indicating the involvement in functions other than nociception.140,141
Several of the TRPV1 activating phytochemicals also activate TRPV3. This may be due to the possibility of coassembly of channels withTRPV1 and TRPV3 subunits. These compounds include thymol, carvacrol, camphor, and eugenol. The channel is also activated by menthol and moderate heat (between 30 and 35 °C).70,115,123,142 Frankincense is a resin obtained from the frankincense tree (Boswellia thurifera), which belongs to the family Buseraceae. The active ingredient is boswellia acid. Myrrh is another resin from the species commiphora. It has been shown that the active ingredient in these resins, incensole, has psychoactive properties. Incensole smoke is used in religious ceremonies to attain higher levels of meditation. Incensole is an activator of TRPV3.143
Compounds in Cannabis sativa, CBD and tetrahydrocannabivarin, caused TRPV3-mediated Ca2+ influx with a EC50 of ∼3.7 μM. Cannabigerovarin and cannabigerolic acid interacted with the channel by causing reduced carvacrol induced Ca2+ influx.144
TRPV4
TRPV4 is expressed in hypothalamus, sensory neurons, trachea, kidney, cochlear hair cells, vascular smooth muscle cells, endothelial cells, and keratinocytes.145−148 TRPV4 is activated by cell-swelling induced by hypotonicity, shear stress, heat (>27 °C), diacyl glycerol (DAG), phorbol esters, 5′,6′-epoxyeicosatrienoic acid (5′,6′-EET), and 4-α-phorbol 12,13-didecanoate (4-α-PDD).146−151 TRPV4 mediates mechanical sensitivity by direct activation of the channel as well as by second messengers produced by mechanical stimuli.152−154 The role of TRPV4 in nociception is confirmed by the administration of antisense oligodeoxynucleotide.155−158
As discussed under activators of TRPA1, paclitaxel is obtained from the Pacific yew and is used to treat certain forms of cancer (Figure 2). The tactile and cold allodynia induced by paclitaxel have been attributed to its ability to activate both TRPV4 and TRPA1. When antagonists of these TRP channels were administered individually, tactile allodynia induced by paclitaxel was alleviated partially. However, a combination of both completely alleviated tactile allodynia. Paclitaxel-induced CGRP release from mouse esophagus was abolished by TRPA1 and TRPV4 antagonists, suggesting that TRPA1 and TRPV4 contribute to paclitaxel-induced neuropathy.41 Bisandrographolide and andrographolide are diterpenoids purified from Andrographis paniculata, which belongs to the family Acanthaceae (Figure 2). Bisandrographolide is able to selectively activate TRPV4 without having any effects on TRPV1, TRPV2, and TRPV3. The abundant phytochemical in the extract, andrographolide, failed to activate TRPV4.159
Cannabidivarin and tetrahydrocannabivarin from cannabis induced TRPV4-mediated Ca2+ influx with an EC50 of 0.9–6.4 μm. whereas cannabigerolic acid, cannabigerovarin, cannabinol, and cannabigerol interacted with TRPV4 causing reduced 4-α-PDD induced responses.144
Concluding Remarks
The TRP family of ion channels has emerged as targets for phytochemicals in botanicals. From the scientific studies, it is becoming apparent that specific and potent active ingredients are being isolated and identified from botanicals. More interestingly, specific receptors for the active ingredients are also being identified, cloned, and characterized. Further, there are endogenous ligands for some of these receptors. The classic examples are endorphin and enkephalin for opioid receptors and anandamide for cannabinoid and TRPV1 receptors. The presence of endogenous ligands strengthens the argument that phytochemicals can fulfill the deficiency of the endogenous ligands or can overactivate the receptor to exert unphysiological responses. TRP channels are expressed in neuronal and nonneuronal cells. Activation of these receptors at the nerve terminals can initiate an afferent sensory signal by depolarizing the nerve terminal and generating an action potential. On the other hand, activation of TRP channels at the nerve terminals can cause an efferent function of releasing peptide hormones that can act locally on other cells in a paracrine fashion (local release) and stimulate the cells to release hormones in an endocrine fashion (release into the blood). Expression of TRP channels in nonneuronal cells, such as pancreatic beta cells and enteroendocrine cells, can release insulin and GLP-1, respectively, which can play a role in glucose homeostasis. It is important to consider the specificity of action, potency, and the bioavailability of these phytochemicals. It is certain that more phytochemicals and their TRP channel targets will be identified in the future to attribute scientific bases for the physiological effects and the health benefits produced by botanicals.
Acknowledgments
I thank Somaja Louis for critically reading the manuscript.
This work is supported by a grant from NIDA (DA028017).
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
References
- Caterina M. J.; Schumacher M. A.; Tominaga M.; Rosen T. A.; Levine J. D.; Julius D. (1997) The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389, 816–824. [DOI] [PubMed] [Google Scholar]
- Cosens D. J.; Manning A. (1969) Abnormal electroretinogram from a Drosophila mutant. Nature 224, 285–287. [DOI] [PubMed] [Google Scholar]
- Montell C.; Rubin G. M. (1989) Molecular characterization of the Drosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron 2, 1313–1323. [DOI] [PubMed] [Google Scholar]
- Montell C. (2005) The TRP superfamily of cation channels. Sci. STKE re3. [DOI] [PubMed] [Google Scholar]
- Story G. M.; Peier A. M.; Reeve A. J.; Eid S. R.; Mosbacher J.; Hricik T. R.; Earley T. J.; Hergarden A. C.; Andersson D. A.; Hwang S. W.; McIntyre P.; Jegla T.; Bevan S.; Patapoutian A. (2003) ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 112, 819–829. [DOI] [PubMed] [Google Scholar]
- Jordt S. E.; Bautista D. M.; Chuang H. H.; McKemy D. D.; Zygmunt P. M.; Hogestatt E. D.; Meng I. D.; Julius D. (2004) Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 427, 260–265. [DOI] [PubMed] [Google Scholar]
- Peier A. M.; Moqrich A.; Hergarden A. C.; Reeve A. J.; Andersson D. A.; Story G. M.; Earley T. J.; Dragoni I.; McIntyre P.; Bevan S.; Patapoutian A. (2002) A TRP channel that senses cold stimuli and menthol. Cell 108, 705–715. [DOI] [PubMed] [Google Scholar]
- McKemy D. D.; Neuhausser W. M.; Julius D. (2002) Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 416, 52–58. [DOI] [PubMed] [Google Scholar]
- Cao E.; Liao M.; Cheng Y.; Julius D. (2013) TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 504, 113–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao M.; Cao E.; Julius D.; Cheng Y. (2013) Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature 504, 107–112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bikman B. T.; Zheng D.; Pories W. J.; Chapman W.; Pender J. R.; Bowden R. C.; Reed M. A.; Cortright R. N.; Tapscott E. B.; Houmard J. A.; Tanner C. J.; Lee J.; Dohm G. L. (2008) Mechanism for improved insulin sensitivity after gastric bypass surgery. J. Clin. Endocrinol. Metab. 93, 4656–4663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corey D. P.; Garcia-Anoveros J.; Holt J. R.; Kwan K. Y.; Lin S. Y.; Vollrath M. A.; Amalfitano A.; Cheung E. L.; Derfler B. H.; Duggan A.; Geleoc G. S.; Gray P. A.; Hoffman M. P.; Rehm H. L.; Tamasauskas D.; Zhang D. S. (2004) TRPA1 is a candidate for the mechanosensitive transduction channel of vertebrate hair cells. Nature 432, 723–730. [DOI] [PubMed] [Google Scholar]
- Bandell M.; Story G. M.; Hwang S. W.; Viswanath V.; Eid S. R.; Petrus M. J.; Earley T. J.; Patapoutian A.; Bautista D. M.; Movahed P.; Hinman A.; Axelsson H. E.; Sterner O.; Hogestatt E. D.; Julius D.; Jordt S. E.; Zygmunt P. M. (2004) Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 41, 849–857. [DOI] [PubMed] [Google Scholar]
- Bautista D. M.; Movahed P.; Hinman A.; Axelsson H. E.; Sterner O.; Hogestatt E. D.; Julius D.; Jordt S. E.; Zygmunt P. M. (2005) Pungent products from garlic activate the sensory ion channel TRPA1. Proc. Natl. Acad. Sci. U.S.A. 102, 12248–12252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macpherson L. J.; Geierstanger B. H.; Viswanath V.; Bandell M.; Eid S. R.; Hwang S.; Patapoutian A. (2005) The pungency of garlic: activation of TRPA1 and TRPV1 in response to allicin. Curr. Biol. 15, 929–934. [DOI] [PubMed] [Google Scholar]
- Kwan K. Y.; Allchorne A. J.; Vollrath M. A.; Christensen A. P.; Zhang D. S.; Woolf C. J.; Corey D. P. (2006) TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 50, 277–289. [DOI] [PubMed] [Google Scholar]
- Andersson D. A.; Gentry C.; Moss S.; Bevan S. (2008) Transient receptor potential A1 is a sensory receptor for multiple products of oxidative stress. J. Neurosci. 28, 2485–2494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bessac B. F.; Sivula M.; von Hehn C. A.; Escalera J.; Cohn L.; Jordt S. E. (2008) TRPA1 is a major oxidant sensor in murine airway sensory neurons. J. Clin. Invest. 118, 1899–1910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao D. S.; Zhong L.; Hsieh T. H.; Abooj M.; Bishnoi M.; Hughes L.; Premkumar L. S. (2012) Expression of transient receptor potential ankyrin 1 (TRPA1) and its role in insulin release from rat pancreatic beta cells. PloS One 7, e38005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brownlee M. (2001) Biochemistry and molecular cell biology of diabetic complications. Nature 414, 813–820. [DOI] [PubMed] [Google Scholar]
- Nagata K.; Duggan A.; Kumar G.; Garcia-Anoveros J. (2005) Nociceptor and hair cell transducer properties of TRPA1, a channel for pain and hearing. J. Neurosci. 25, 4052–4061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obata K.; Katsura H.; Mizushima T.; Yamanaka H.; Kobayashi K.; Dai Y.; Fukuoka T.; Tokunaga A.; Tominaga M.; Noguchi K. (2005) TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Invest. 115, 2393–2401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hinman A.; Chuang H. H.; Bautista D. M.; Julius D. (2006) TRP channel activation by reversible covalent modification. Proc. Natl. Acad. Sci. U.S.A. 103, 19564–19568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raisinghani M.; Zhong L.; Jeffry J. A.; Bishnoi M.; Pabbidi R. M.; Pimentel F.; Cao D. S.; Evans M. S.; Premkumar L. S. (2011) Activation characteristics of transient receptor potential ankyrin 1 and its role in nociception. Am. J. Physiol.: Cell Physiol. 301, C587–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Premkumar L. S.; Raisinghani M. (2006) Nociceptors in cardiovascular functions: complex interplay as a result of cyclooxygenase inhibition. Mol. Pain 2, 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan A.; Safdar M.; Ali Khan M. M.; Khattak K. N.; Anderson R. A. (2003) Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care 26, 3215–3218. [DOI] [PubMed] [Google Scholar]
- Hlebowicz J.; Darwiche G.; Bjorgell O.; Almer L. O. (2007) Effect of cinnamon on postprandial blood glucose, gastric emptying, and satiety in healthy subjects. Am. J. Clin. Nutr. 85, 1552–1556. [DOI] [PubMed] [Google Scholar]
- Lissiman E.; Bhasale A. L.; Cohen M. (2009) Garlic for the common cold. Cochrane Database Syst. Rev. 3, CD006206. [DOI] [PubMed] [Google Scholar]
- Lissiman E.; Bhasale A. L.; Cohen M. (2012) Garlic for the common cold. Cochrane Database Syst. Rev. 14, CD006206. [DOI] [PubMed] [Google Scholar]
- Cutler R. R.; Odent M.; Hajj-Ahmad H.; Maharjan S.; Bennett N. J.; Josling P. D.; Ball V.; Hatton P.; Dall’Antonia M. (2009) In vitro activity of an aqueous allicin extract and a novel allicin topical gel formulation against Lancefield group B streptococci. J. Antimicrob. Chemother. 63, 151–154. [DOI] [PubMed] [Google Scholar]
- Khanum F.; Anilakumar K. R.; Viswanathan K. R. (2004) Anticarcinogenic properties of garlic: a review. Crit. Rev. Food. Sci. Nutr. 44, 479–488. [DOI] [PubMed] [Google Scholar]
- Shenoy N. R.; Choughuley A. S. (1992) Inhibitory effect of diet related sulphydryl compounds on the formation of carcinogenic nitrosamines. Cancer Lett. 65, 227–232. [DOI] [PubMed] [Google Scholar]
- Koizumi K.; Iwasaki Y.; Narukawa M.; Iitsuka Y.; Fukao T.; Seki T.; Ariga T.; Watanabe T. (2009) Diallyl sulfides in garlic activate both TRPA1 and TRPV1. Biochem. Biophys. Res. Commun. 382, 545–548. [DOI] [PubMed] [Google Scholar]
- Leamy A. W.; Shukla P.; McAlexander M. A.; Carr M. J.; Ghatta S. (2011) Curcumin ((E,E)-1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) activates and desensitizes the nociceptor ion channel TRPA1. Neurosci. Lett. 503, 157–162. [DOI] [PubMed] [Google Scholar]
- Gupta S. C.; Patchva S.; Aggarwal B. B. (2013) Therapeutic roles of curcumin: lessons learned from clinical trials. AAPS J. 15, 195–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song Y.; Sonawane N. D.; Salinas D.; Qian L.; Pedemonte N.; Galietta L. J.; Verkman A. S. (2004) Evidence against the rescue of defective DeltaF508-CFTR cellular processing by curcumin in cell culture and mouse models. J. Biol. Chem. 279, 40629–40633. [DOI] [PubMed] [Google Scholar]
- Gao J.; Zhou H.; Lei T.; Zhou L.; Li W.; Li X.; Yang B. (2011) Curcumin inhibits renal cyst formation and enlargement in vitro by regulating intracellular signaling pathways. Eur. J. Pharmacol. 654, 92–99. [DOI] [PubMed] [Google Scholar]
- Sohma Y.; Yu Y. C.; Hwang T. C. (2013) Curcumin and genistein: the combined effects on disease-associated CFTR mutants and their clinical implications. Curr. Pharm. Des. 19, 3521–3528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nassini R.; Materazzi S.; Vriens J.; Prenen J.; Benemei S.; De Siena G.; la Marca G.; Andre E.; Preti D.; Avonto C.; Sadofsky L.; Di Marzo V.; De Petrocellis L.; Dussor G.; Porreca F.; Taglialatela-Scafati O.; Appendino G.; Nilius B.; Geppetti P. (2012) The ‘headache tree’ via umbellulone and TRPA1 activates the trigeminovascular system. Brain 135, 376–390. [DOI] [PubMed] [Google Scholar]
- Zhong J.; Pollastro F.; Prenen J.; Zhu Z.; Appendino G.; Nilius B. (2011) Ligustilide: a novel TRPA1 modulator. Pfluegers Arch. 462, 841–849. [DOI] [PubMed] [Google Scholar]
- Materazzi S.; Fusi C.; Benemei S.; Pedretti P.; Patacchini R.; Nilius B.; Prenen J.; Creminon C.; Geppetti P.; Nassini R. (2012) TRPA1 and TRPV4 mediate paclitaxel-induced peripheral neuropathy in mice via a glutathione-sensitive mechanism. Pfluegers Arch. 463, 561–569. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L.; Vellani V.; Schiano-Moriello A.; Marini P.; Magherini P. C.; Orlando P.; Di Marzo V. (2008) Plant-derived cannabinoids modulate the activity of transient receptor potential channels of ankyrin type-1 and melastatin type-8. J. Pharmacol. Exp. Ther. 325, 1007–1015. [DOI] [PubMed] [Google Scholar]
- Montrucchio D. P.; Cordova M. M.; Santos A. R. (2013) Plant derived aporphinic alkaloid S-(+)-dicentrine induces antinociceptive effect in both acute and chronic inflammatory pain models: evidence for a role of TRPA1 channels. PloS One 8, e67730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talavera K.; Gees M.; Karashima Y.; Meseguer V. M.; Vanoirbeek J. A.; Damann N.; Everaerts W.; Benoit M.; Janssens A.; Vennekens R.; Viana F.; Nemery B.; Nilius B.; Voets T. (2009) Nicotine activates the chemosensory cation channel TRPA1. Nat. Neurosci. 12, 1293–1299. [DOI] [PubMed] [Google Scholar]
- Riera C. E.; Menozzi-Smarrito C.; Affolter M.; Michlig S.; Munari C.; Robert F.; Vogel H.; Simon S. A.; Le Coutre J. (2009) Compounds from Sichuan and Melegueta peppers activate, covalently and non-covalently, TRPA1 and TRPV1 channels. Br. J. Pharmacol. 157, 1398–1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vazquez G.; Wedel B. J.; Aziz O.; Trebak M.; Putney J. W. Jr. (2004) The mammalian TRPC cation channels. Biochim. Biophys. Acta 1742, 21–36. [DOI] [PubMed] [Google Scholar]
- Birnbaumer L. (2009) The TRPC class of ion channels: a critical review of their roles in slow, sustained increases in intracellular Ca(2+) concentrations. Annu. Rev. Pharmacol. Toxicol. 49, 395–426. [DOI] [PubMed] [Google Scholar]
- Leuner K.; Kazanski V.; Muller M.; Essin K.; Henke B.; Gollasch M.; Harteneck C.; Muller W. E. (2007) Hyperforin--a key constituent of St. John’s wort specifically activates TRPC6 channels. FASEB J. 21, 4101–4111. [DOI] [PubMed] [Google Scholar]
- Sikand P.; Premkumar L. S. (2007) Potentiation of glutamatergic synaptic transmission by protein kinase C-mediated sensitization of TRPV1 at the first sensory synapse. J. Physiol. 581, 631–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao D. S.; Yu S. Q.; Premkumar L. S. (2009) Modulation of transient receptor potential Vanilloid 4-mediated membrane currents and synaptic transmission by protein kinase C. Mol. Pain 5, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeffry J. A.; Yu S. Q.; Sikand P.; Parihar A.; Evans M. S.; Premkumar L. S. (2009) Selective targeting of TRPV1 expressing sensory nerve terminals in the spinal cord for long lasting analgesia. PloS One 4, e7021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans M. S.; Cheng X.; Jeffry J. A.; Disney K. E.; Premkumar L. S. (2012) Sumatriptan inhibits TRPV1 channels in trigeminal neurons. Headache 52, 773–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou J.; Du W.; Zhou K.; Tai Y.; Yao H.; Jia Y.; Ding Y. (2008) Critical role of TRPC6 channels in the formation of excitatory synapses. Nat. Neurosci. 11, 741–743. [DOI] [PubMed] [Google Scholar]
- Nilius B.; Owsianik G. (2011) The transient receptor potential family of ion channels. Genome Biol. 12, 218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carakostas M. C.; Curry L. L.; Boileau A. C.; Brusick D. J. (2008) Overview: the history, technical function and safety of rebaudioside A, a naturally occurring steviol glycoside, for use in food and beverages. Food Chem. Toxicol. 46(Suppl 7), S1–S10. [DOI] [PubMed] [Google Scholar]
- Medler K. F. (2011) Multiple roles for TRPs in the taste system: not your typical TRPs. Adv. Exp. Med. Biol. 704, 831–846. [DOI] [PubMed] [Google Scholar]
- Sprous D.; Palmer K. R. (2010) The T1R2/T1R3 sweet receptor and TRPM5 ion channel taste targets with therapeutic potential. Prog. Mol. Biol. Transl. Sci. 91, 151–208. [DOI] [PubMed] [Google Scholar]
- Palmer R. K. (2007) The pharmacology and signaling of bitter, sweet, and umami taste sensing. Mol. Interv. 7, 87–98. [DOI] [PubMed] [Google Scholar]
- Depoortere I. (2014) Taste receptors of the gut: emerging roles in health and disease. Gut 63, 179–190. [DOI] [PubMed] [Google Scholar]
- Kaji I.; Karaki S. I.; Kuwahara A. (2014) Taste Sensing in the Colon. Curr. Pharm. Des. 20, 2766–2774. [DOI] [PubMed] [Google Scholar]
- Premkumar L. S.; Raisinghani M.; Pingle S. C.; Long C.; Pimentel F. (2005) Downregulation of transient receptor potential melastatin 8 by protein kinase C-mediated dephosphorylation. J. Neurosci. 25, 11322–11329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuzuki K.; Xing H.; Ling J.; Gu J. G. (2004) Menthol-induced Ca2+ release from presynaptic Ca2+ stores potentiates sensory synaptic transmission. J. Neurosci. 24, 762–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsavaler L.; Shapero M. H.; Morkowski S.; Laus R. (2001) Trp-p8, a novel prostate-specific gene, is up-regulated in prostate cancer and other malignancies and shares high homology with transient receptor potential calcium channel proteins. Cancer Res. 61, 3760–3769. [PubMed] [Google Scholar]
- Uhl G. R.; Walther D.; Behm F. M.; Rose J. E. (2011) Menthol preference among smokers: association with TRPA1 variants. Nicotine Tob. Res. 13, 1311–1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hans M.; Wilhelm M.; Swandulla D. (2012) Menthol suppresses nicotinic acetylcholine receptor functioning in sensory neurons via allosteric modulation. Chem. Senses 37, 463–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willis D. N.; Liu B.; Ha M. A.; Jordt S. E.; Morris J. B. (2011) Menthol attenuates respiratory irritation responses to multiple cigarette smoke irritants. FASEB J. 25, 4434–4444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leuenroth S. J.; Okuhara D.; Shotwell J. D.; Markowitz G. S.; Yu Z.; Somlo S.; Crews C. M. (2007) Triptolide is a traditional Chinese medicine-derived inhibitor of polycystic kidney disease. Proc. Natl. Acad. Sci. U.S.A. 104, 4389–4394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szallasi A.; Blumberg P. M. (1999) Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol. Rev. 51, 159–212. [PubMed] [Google Scholar]
- Vennekens R.; Owsianik G.; Nilius B. (2008) Vanilloid transient receptor potential cation channels: an overview. Curr. Pharm. Des. 14, 18–31. [DOI] [PubMed] [Google Scholar]
- Vriens J.; Nilius B.; Vennekens R. (2008) Herbal compounds and toxins modulating TRP channels. Curr. Neuropharmacol. 6, 79–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellwig N.; Albrecht N.; Harteneck C.; Schultz G.; Schaefer M. (2005) Homo- and heteromeric assembly of TRPV channel subunits. J. Cell Sci. 118, 917–928. [DOI] [PubMed] [Google Scholar]
- Tominaga M.; Caterina M. J.; Malmberg A. B.; Rosen T. A.; Gilbert H.; Skinner K.; Raumann B. E.; Basbaum A. I.; Julius D. (1998) The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 21, 531–543. [DOI] [PubMed] [Google Scholar]
- Nakatsuka T.; Furue H.; Yoshimura M.; Gu J. G. (2002) Activation of central terminal vanilloid receptor-1 receptors and alpha beta-methylene-ATP-sensitive P2X receptors reveals a converged synaptic activity onto the deep dorsal horn neurons of the spinal cord. J. Neurosci. 22, 1228–1237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baccei M. L.; Bardoni R.; Fitzgerald M. (2003) Development of nociceptive synaptic inputs to the neonatal rat dorsal horn: glutamate release by capsaicin and menthol. J. Physiol. 549, 231–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim Y. H.; Back S. K.; Davies A. J.; Jeong H.; Jo H. J.; Chung G.; Na H. S.; Bae Y. C.; Kim S. J.; Kim J. S.; Jung S. J.; Oh S. B. (2012) TRPV1 in GABAergic interneurons mediates neuropathic mechanical allodynia and disinhibition of the nociceptive circuitry in the spinal cord. Neuron 74, 640–647. [DOI] [PubMed] [Google Scholar]
- Doyle M. W.; Bailey T. W.; Jin Y. H.; Andresen M. C. (2002) Vanilloid receptors presynaptically modulate cranial visceral afferent synaptic transmission in nucleus tractus solitarius. J. Neurosci. 22, 8222–8229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli S.; Di Marzo V.; Berretta N.; Matias I.; Maccarrone M.; Bernardi G.; Mercuri N. B. (2003) Presynaptic facilitation of glutamatergic synapses to dopaminergic neurons of the rat substantia nigra by endogenous stimulation of vanilloid receptors. J. Neurosci. 23, 3136–3144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli S.; Vaughan C. W.; Christie M. J.; Connor M. (2002) Capsaicin activation of glutamatergic synaptic transmission in the rat locus coeruleus in vitro. J. Physiol. 543, 531–540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson H. E.; Edwards J. G.; Page R. S.; Van Hook M. J.; Kauer J. A. (2008) TRPV1 channels mediate long-term depression at synapses on hippocampal interneurons. Neuron 57, 746–759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grueter B. A.; Brasnjo G.; Malenka R. C. (2010) Postsynaptic TRPV1 triggers cell type-specific long-term depression in the nucleus accumbens. Nat. Neurosci. 13, 1519–1525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Premkumar L. S.; Ahern G. P. (2000) Induction of vanilloid receptor channel activity by protein kinase C. Nature 408, 985–990. [DOI] [PubMed] [Google Scholar]
- Caterina M. J.; Julius D. (2001) The vanilloid receptor: a molecular gateway to the pain pathway. Annu. Rev. Neurosci. 24, 487–517. [DOI] [PubMed] [Google Scholar]
- Minke B.; Cook B. (2002) TRP channel proteins and signal transduction. Physiol. Rev. 82, 429–472. [DOI] [PubMed] [Google Scholar]
- Clapham D. E. (2003) TRP channels as cellular sensors. Nature 426, 517–524. [DOI] [PubMed] [Google Scholar]
- Premkumar L. S.; Bishnoi M. (2011) Disease-related changes in TRPV1 expression and its implications for drug development. Curr. Top. Med. Chem. 11, 2192–2209. [DOI] [PubMed] [Google Scholar]
- Julius D. (2013) TRP channels and pain. Annu. Rev. Cell Dev. Biol. 29, 355–384. [DOI] [PubMed] [Google Scholar]
- Mezey E.; Toth Z. E.; Cortright D. N.; Arzubi M. K.; Krause J. E.; Elde R.; Guo A.; Blumberg P. M.; Szallasi A. (2000) Distribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and human. Proc. Natl. Acad. Sci. U.S.A. 97, 3655–3660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang S. M.; Bisogno T.; Trevisani M.; Al-Hayani A.; De Petrocellis L.; Fezza F.; Tognetto M.; Petros T. J.; Krey J. F.; Chu C. J.; Miller J. D.; Davies S. N.; Geppetti P.; Walker J. M.; Di Marzo V. (2002) An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. U.S.A. 99, 8400–8405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishra S. K.; Tisel S. M.; Orestes P.; Bhangoo S. K.; Hoon M. A. (2011) TRPV1-lineage neurons are required for thermal sensation. EMBO J. 30, 582–593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanaugh D. J.; Chesler A. T.; Braz J. M.; Shah N. M.; Julius D.; Basbaum A. I. (2011) Restriction of transient receptor potential vanilloid-1 to the peptidergic subset of primary afferent neurons follows its developmental downregulation in nonpeptidergic neurons. J. Neurosci. 31, 10119–10127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundberg J. M.; Martling C. R.; Saria A. (1983) Substance P and capsaicin-induced contraction of human bronchi. Acta Physiol. Scand. 119, 49–53. [DOI] [PubMed] [Google Scholar]
- Mitchell J. A.; Williams F. M.; Williams T. J.; Larkin S. W. (1997) Role of nitric oxide in the dilator actions of capsaicin-sensitive nerves in the rabbit coronary circulation. Neuropeptides 31, 333–338. [DOI] [PubMed] [Google Scholar]
- Zygmunt P. M.; Petersson J.; Andersson D. A.; Chuang H.; Sorgard M.; Di Marzo V.; Julius D.; Hogestatt E. D.; Wang Y. (1999) Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 400, 452–457. [DOI] [PubMed] [Google Scholar]
- Birder L. A.; Nakamura Y.; Kiss S.; Nealen M. L.; Barrick S.; Kanai A. J.; Wang E.; Ruiz G.; De Groat W. C.; Apodaca G.; Watkins S.; Caterina M. J. (2002) Altered urinary bladder function in mice lacking the vanilloid receptor TRPV1. Nat. Neurosci. 5, 856–860. [DOI] [PubMed] [Google Scholar]
- Cruz F.; Dinis P. (2007) Resiniferatoxin and botulinum toxin type A for treatment of lower urinary tract symptoms. Neurourol. Urodyn. 26, 920–927. [DOI] [PubMed] [Google Scholar]
- Varga A.; Nemeth J.; Szabo A.; McDougall J. J.; Zhang C.; Elekes K.; Pinter E.; Szolcsanyi J.; Helyes Z. (2005) Effects of the novel TRPV1 receptor antagonist SB366791 in vitro and in vivo in the rat. Neurosci. Lett. 385, 137–142. [DOI] [PubMed] [Google Scholar]
- Gavva N. R. (2008) Body-temperature maintenance as the predominant function of the vanilloid receptor TRPV1. Trends Pharmacol. Sci. 29, 550–557. [DOI] [PubMed] [Google Scholar]
- Lehto S. G.; Tamir R.; Deng H.; Klionsky L.; Kuang R.; Le A.; Lee D.; Louis J. C.; Magal E.; Manning B. H.; Rubino J.; Surapaneni S.; Tamayo N.; Wang T.; Wang J.; Wang J.; Wang W.; Youngblood B.; Zhang M.; Zhu D.; Norman M. H.; Gavva N. R. (2008) Antihyperalgesic effects of (R,E)-N-(2-hydroxy-2,3-dihydro-1H-inden-4-yl)-3-(2-(piperidin-1-yl)-4-(trifluorom ethyl)phenyl)-acrylamide (AMG8562), a novel transient receptor potential vanilloid type 1 modulator that does not cause hyperthermia in rats. J. Pharmacol. Exp. Ther. 326, 218–229. [DOI] [PubMed] [Google Scholar]
- Pabbidi R. M.; Yu S. Q.; Peng S.; Khardori R.; Pauza M. E.; Premkumar L. S. (2008) Influence of TRPV1 on diabetes-induced alterations in thermal pain sensitivity. Mol. Pain 4, 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Buren J. J.; Bhat S.; Rotello R.; Pauza M. E.; Premkumar L. S. (2005) Sensitization and translocation of TRPV1 by insulin and IGF-I. Mol. Pain 1, 17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bishnoi M.; Bosgraaf C. A.; Abooj M.; Zhong L.; Premkumar L. S. (2011) Streptozotocin-induced early thermal hyperalgesia is independent of glycemic state of rats: role of transient receptor potential vanilloid 1(TRPV1) and inflammatory mediators. Mol. Pain 7, 52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akiba Y.; Kato S.; Katsube K.; Nakamura M.; Takeuchi K.; Ishii H.; Hibi T. (2004) Transient receptor potential vanilloid subfamily 1 expressed in pancreatic islet beta cells modulates insulin secretion in rats. Biochem. Biophys. Res. Commun. 321, 219–225. [DOI] [PubMed] [Google Scholar]
- Razavi R.; Chan Y.; Afifiyan F. N.; Liu X. J.; Wan X.; Yantha J.; Tsui H.; Tang L.; Tsai S.; Santamaria P.; Driver J. P.; Serreze D.; Salter M. W.; Dosch H. M. (2006) TRPV1+ sensory neurons control beta cell stress and islet inflammation in autoimmune diabetes. Cell 127, 1123–1135. [DOI] [PubMed] [Google Scholar]
- Wang X.; Miyares R. L.; Ahern G. P. (2005) Oleoylethanolamide excites vagal sensory neurones, induces visceral pain and reduces short-term food intake in mice via capsaicin receptor TRPV1. J. Physiol. 564, 541–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahern G. P. (2013) Transient receptor potential channels and energy homeostasis. Trends Endocrinol. Metab. 24, 554–560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kindt S.; Vos R.; Blondeau K.; Tack J. (2009) Influence of intra-oesophageal capsaicin instillation on heartburn induction and oesophageal sensitivity in man. Neurogastroenterol. Motil. 21, 1032–e1082. [DOI] [PubMed] [Google Scholar]
- Okumi H.; Tashima K.; Matsumoto K.; Namiki T.; Terasawa K.; Horie S. (2012) Dietary agonists of TRPV1 inhibit gastric acid secretion in mice. Planta Med. 78, 1801–1806. [DOI] [PubMed] [Google Scholar]
- Raisinghani M.; Pabbidi R. M.; Premkumar L. S. (2005) Activation of transient receptor potential vanilloid 1 (TRPV1) by resiniferatoxin. J. Physiol. 567, 771–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts J. C.; Davis J. B.; Benham C. D. (2004) [3H]Resiniferatoxin autoradiography in the CNS of wild-type and TRPV1 null mice defines TRPV1 (VR-1) protein distribution. Brain Res. 995, 176–183. [DOI] [PubMed] [Google Scholar]
- Chou M. Z.; Mtui T.; Gao Y. D.; Kohler M.; Middleton R. E. (2004) Resiniferatoxin binds to the capsaicin receptor (TRPV1) near the extracellular side of the S4 transmembrane domain. Biochemistry 43, 2501–2511. [DOI] [PubMed] [Google Scholar]
- Jung J.; Lee S. Y.; Hwang S. W.; Cho H.; Shin J.; Kang Y. S.; Kim S.; Oh U. (2002) Agonist recognition sites in the cytosolic tails of vanilloid receptor 1. J. Biol. Chem. 277, 44448–44454. [DOI] [PubMed] [Google Scholar]
- Gavva N. R.; Klionsky L.; Qu Y.; Shi L.; Tamir R.; Edenson S.; Zhang T. J.; Viswanadhan V. N.; Toth A.; Pearce L. V.; Vanderah T. W.; Porreca F.; Blumberg P. M.; Lile J.; Sun Y.; Wild K.; Louis J. C.; Treanor J. J. (2004) Molecular determinants of vanilloid sensitivity in TRPV1. J. Biol. Chem. 279, 20283–20295. [DOI] [PubMed] [Google Scholar]
- Brown D. C.; Iadarola M. J.; Perkowski S. Z.; Erin H.; Shofer F.; Laszlo K. J.; Olah Z.; Mannes A. J. (2005) Physiologic and antinociceptive effects of intrathecal resiniferatoxin in a canine bone cancer model. Anesthesiology 103, 1052–1059. [DOI] [PubMed] [Google Scholar]
- Iadarola M. J.; Mannes A. J. (2011) The vanilloid agonist resiniferatoxin for interventional-based pain control. Curr. Top. Med. Chem. 11, 2171–2179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang B. H.; Piao Z. G.; Kim Y. B.; Lee C. H.; Lee J. K.; Park K.; Kim J. S.; Oh S. B. (2003) (2003) Activation of vanilloid receptor 1 (VR1) by eugenol. J. Dent. Res. 82, 781–785. [DOI] [PubMed] [Google Scholar]
- Behrendt H. J.; Germann T.; Gillen C.; Hatt H.; Jostock R. (2004) Characterization of the mouse cold-menthol receptor TRPM8 and vanilloid receptor type-1 VR1 using a fluorometric imaging plate reader (FLIPR) assay. Br. J. Pharmacol. 141, 737–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNamara F. N.; Randall A.; Gunthorpe M. J. (2005) Effects of piperine, the pungent component of black pepper, at the human vanilloid receptor (TRPV1). Br. J. Pharmacol. 144, 781–790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhardwaj R. K.; Glaeser H.; Becquemont L.; Klotz U.; Gupta S. K.; Fromm M. F. (2002) Piperine, a major constituent of black pepper, inhibits human P-glycoprotein and CYP3A4. J. Pharmacol. Exp. Ther. 302, 645–650. [DOI] [PubMed] [Google Scholar]
- Bisogno T.; Hanus L.; De Petrocellis L.; Tchilibon S.; Ponde D. E.; Brandi I.; Moriello A. S.; Davis J. B.; Mechoulam R.; Di Marzo V. (2001) Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 134, 845–852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu H.; Blair N. T.; Clapham D. E. (2005) Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism. J. Neurosci. 25, 8924–8937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearce L. V.; Petukhov P. A.; Szabo T.; Kedei N.; Bizik F.; Kozikowski A. P.; Blumberg P. M. (2004) Evodiamine functions as an agonist for the vanilloid receptor TRPV1. Org. Biomol. Chem. 2, 2281–2286. [DOI] [PubMed] [Google Scholar]
- Morera E.; De Petrocellis L.; Morera L.; Moriello A. S.; Nalli M.; Di Marzo V.; Ortar G. (2012) Synthesis and biological evaluation of [6]-gingerol analogues as transient receptor potential channel TRPV1 and TRPA1 modulators. Bioorg. Med. Chem. Lett. 22, 1674–1677. [DOI] [PubMed] [Google Scholar]
- Xu H.; Delling M.; Jun J. C.; Clapham D. E. (2006) Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels. Nat. Neurosci. 9, 628–635. [DOI] [PubMed] [Google Scholar]
- Andre E.; Campi B.; Trevisani M.; Ferreira J.; Malheiros A.; Yunes R. A.; Calixto J. B.; Geppetti P. (2006) Pharmacological characterisation of the plant sesquiterpenes polygodial and drimanial as vanilloid receptor agonists. Biochem. Pharmacol. 71, 1248–1254. [DOI] [PubMed] [Google Scholar]
- Iwasaki Y.; Tanabe M.; Kayama Y.; Abe M.; Kashio M.; Koizumi K.; Okumura Y.; Morimitsu Y.; Tominaga M.; Ozawa Y.; Watanabe T. (2009) Miogadial and miogatrial with alpha,beta-unsaturated 1,4-dialdehyde moieties—novel and potent TRPA1 agonists. Life Sci. 85, 60–69. [DOI] [PubMed] [Google Scholar]
- Lübbert M.; Kyereme J.; Schöbel N.; Beltrán L.; Wetzel C. H.; Hatt H. (2013) Transient receptor potential channels encode volatile chemicals sensed by rat trigeminal ganglion neurons. PLoS One 8, e77998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trevisan G.; Rossato M. F.; Walker C. I.; Klafke J. Z.; Rosa F.; Oliveira S. M.; Tonello R.; Guerra G. P.; Boligon A. A.; Zanon R. B.; Athayde M. L.; Ferreira J. (2012) Identification of the plant steroid alpha-spinasterol as a novel transient receptor potential vanilloid 1 antagonist with antinociceptive properties. J. Pharmacol. Exp. Ther. 343, 258–269. [DOI] [PubMed] [Google Scholar]
- Birnbaumer L.; Yildirim E.; Abramowitz J. (2003) A comparison of the genes coding for canonical TRP channels and their M, V and P relatives. Cell Calcium 33, 419–432. [DOI] [PubMed] [Google Scholar]
- Peier A. M.; Reeve A. J.; Andersson D. A.; Moqrich A.; Earley T. J.; Hergarden A. C.; Story G. M.; Colley S.; Hogenesch J. B.; McIntyre P.; Bevan S.; Patapoutian A. (2002) A heat-sensitive TRP channel expressed in keratinocytes. Science 296, 2046–2049. [DOI] [PubMed] [Google Scholar]
- Moqrich A.; Hwang S. W.; Earley T. J.; Petrus M. J.; Murray A. N.; Spencer K. S.; Andahazy M.; Story G. M.; Patapoutian A. (2005) Impaired thermosensation in mice lacking TRPV3, a heat and camphor sensor in the skin. Science 307, 1468–1472. [DOI] [PubMed] [Google Scholar]
- Xu H.; Ramsey I. S.; Kotecha S. A.; Moran M. M.; Chong J. A.; Lawson D.; Ge P.; Lilly J.; Silos-Santiago I.; Xie Y.; DiStefano P. S.; Curtis R.; Clapham D. E. (2002) TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 418, 181–186. [DOI] [PubMed] [Google Scholar]
- Smith G. D.; Gunthorpe M. J.; Kelsell R. E.; Hayes P. D.; Reilly P.; Facer P.; Wright J. E.; Jerman J. C.; Walhin J. P.; Ooi L.; Egerton J.; Charles K. J.; Smart D.; Randall A. D.; Anand P.; Davis J. B. (2002) TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 418, 186–190. [DOI] [PubMed] [Google Scholar]
- Gopinath P.; Wan E.; Holdcroft A.; Facer P.; Davis J. B.; Smith G. D.; Bountra C.; Anand P. (2005) Increased capsaicin receptor TRPV1 in skin nerve fibres and related vanilloid receptors TRPV3 and TRPV4 in keratinocytes in human breast pain. BMC Women’s Health 5, 2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Facer P.; Casula M. A.; Smith G. D.; Benham C. D.; Chessell I. P.; Bountra C.; Sinisi M.; Birch R.; Anand P. (2007) Differential expression of the capsaicin receptor TRPV1 and related novel receptors TRPV3, TRPV4 and TRPM8 in normal human tissues and changes in traumatic and diabetic neuropathy. BMC Neurol. 7, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang S. M.; Lee H.; Chung M. K.; Park U.; Yu Y. Y.; Bradshaw H. B.; Coulombe P. A.; Walker J. M.; Caterina M. J. (2008) Overexpressed transient receptor potential vanilloid 3 ion channels in skin keratinocytes modulate pain sensitivity via prostaglandin E2. J. Neurosci. 28, 13727–13737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammarstrom S.; Hamberg M.; Samuelsson B.; Duell E. A.; Stawiski M.; Voorhees J. J. (1975) Increased concentrations of nonesterified arachidonic acid, 12L-hydroxy-5,8,10,14-eicosatetraenoic acid, prostaglandin E2, and prostaglandin F2alpha in epidermis of psoriasis. Proc. Natl. Acad. Sci. U.S.A. 72, 5130–5134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brash A. R. (2001) Arachidonic acid as a bioactive molecule. J. Clin. Invest. 107, 1339–1345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandadi S.; Sokabe T.; Shibasaki K.; Katanosaka K.; Mizuno A.; Moqrich A.; Patapoutian A.; Fukumi-Tominaga T.; Mizumura K.; Tominaga M. (2009) TRPV3 in keratinocytes transmits temperature information to sensory neurons via ATP. Pfluegers Arch. 458, 1093–2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao R.; Tang J.; Wang C.; Colton C. K.; Tian J.; Zhu M. X. (2008) Calcium plays a central role in the sensitization of TRPV3 channel to repetitive stimulations. J. Biol. Chem. 283, 6162–6174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imura K. (2007) Influence of TRPV3 mutation on hair growth cycle in mice. Biochem. Biophys. Res. Commun. 363, 479–483. [DOI] [PubMed] [Google Scholar]
- Asakawa M.; Yoshioka T.; Matsutani T.; Hikita I.; Suzuki M.; Oshima I.; Tsukahara K.; Arimura A.; Horikawa T.; Hirasawa T.; Sakata T. (2006) Association of a mutation in TRPV3 with defective hair growth in rodents. J. Invest. Dermatol. 126, 2664–2672. [DOI] [PubMed] [Google Scholar]
- Macpherson L. J.; Hwang S. W.; Miyamoto T.; Dubin A. E.; Patapoutian A.; Story G. M. (2006) More than cool: promiscuous relationships of menthol and other sensory compounds. Mol. Cell Neurosci. 32, 335–343. [DOI] [PubMed] [Google Scholar]
- Moussaieff A.; Rimmerman N.; Bregman T.; Straiker A.; Felder C. C.; Shoham S.; Kashman Y.; Huang S. M.; Lee H.; Shohami E.; Mackie K.; Caterina M. J.; Walker J. M.; Fride E.; Mechoulam R. (2008) Incensole acetate, an incense component, elicits psychoactivity by activating TRPV3 channels in the brain. FASEB J. 22, 3024–3034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Petrocellis L.; Orlando P.; Moriello A. S.; Aviello G.; Stott C.; Izzo A. A.; Di Marzo V. (2012) Cannabinoid actions at TRPV channels: effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol. 204, 255–266. [DOI] [PubMed] [Google Scholar]
- Strotmann R.; Harteneck C.; Nunnenmacher K.; Schultz G.; Plant T. D. (2000) OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat. Cell Biol. 2, 695–702. [DOI] [PubMed] [Google Scholar]
- Watanabe H.; Davis J. B.; Smart D.; Jerman J. C.; Smith G. D.; Hayes P.; Vriens J.; Cairns W.; Wissenbach U.; Prenen J.; Flockerzi V.; Droogmans G.; Benham C. D.; Nilius B. (2002) Activation of TRPV4 channels (hVRL-2/mTRP12) by phorbol derivatives. J. Biol. Chem. 277, 13569–13577. [DOI] [PubMed] [Google Scholar]
- Suzuki M.; Mizuno A.; Kodaira K.; Imai M. (2003) Impaired pressure sensation in mice lacking TRPV4. J. Biol. Chem. 278, 22664–22668. [DOI] [PubMed] [Google Scholar]
- Nilius B.; Droogmans G.; Wondergem R. (2003) Transient receptor potential channels in endothelium: solving the calcium entry puzzle?. Endothelium 10, 5–15. [DOI] [PubMed] [Google Scholar]
- Kohler R.; Heyken W. T.; Heinau P.; Schubert R.; Si H.; Kacik M.; Busch C.; Grgic I.; Maier T.; Hoyer J. (2006) Evidence for a functional role of endothelial transient receptor potential V4 in shear stress-induced vasodilatation. Arterioscler., Thromb., Vasc. Biol. 26, 1495–15502. [DOI] [PubMed] [Google Scholar]
- Watanabe H.; Vriens J.; Suh S. H.; Benham C. D.; Droogmans G.; Nilius B. (2002) Heat-evoked activation of TRPV4 channels in a HEK293 cell expression system and in native mouse aorta endothelial cells. J. Biol. Chem. 277, 47044–47051. [DOI] [PubMed] [Google Scholar]
- Watanabe H.; Vriens J.; Prenen J.; Droogmans G.; Voets T.; Nilius B. (2003) Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 424, 434–438. [DOI] [PubMed] [Google Scholar]
- Loukin S. H.; Su Z.; Kung C. (2009) Hypotonic shocks activate rat TRPV4 in yeast in the absence of polyunsaturated fatty acids. FEBS Lett. 583, 754–758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao D. S.; Yu S. Q.; Premkumar L. S. (2009) Modulation of transient receptor potential Vanilloid 4-mediated membrane currents and synaptic transmission by protein kinase C. Mol. Pain 5, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews B. D.; Thodeti C. K.; Tytell J. D.; Mammoto A.; Overby D. R.; Ingber D. E. (2010) Ultra-rapid activation of TRPV4 ion channels by mechanical forces applied to cell surface beta1 integrins. Integr. Biol. 2, 435–442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alessandri-Haber N.; Dina O. A.; Joseph E. K.; Reichling D. B.; Levine J. D. (2008) Interaction of transient receptor potential vanilloid 4, integrin, and SRC tyrosine kinase in mechanical hyperalgesia. J. Neurosci. 28, 1046–1057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X.; Alessandri-Haber N.; Levine J. D. (2007) Marked attenuation of inflammatory mediator-induced C-fiber sensitization for mechanical and hypotonic stimuli in TRPV4–/– mice. Mol. Pain 3, 31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alessandri-Haber N.; Yeh J. J.; Boyd A. E.; Parada C. A.; Chen X.; Reichling D. B.; Levine J. D. (2003) Hypotonicity induces TRPV4-mediated nociception in rat. Neuron 39, 497–511. [DOI] [PubMed] [Google Scholar]
- Alessandri-Haber N.; Joseph E.; Dina O. A.; Liedtke W.; Levine J. D. (2005) TRPV4 mediates pain-related behavior induced by mild hypertonic stimuli in the presence of inflammatory mediator. Pain 118, 70–79. [DOI] [PubMed] [Google Scholar]
- Smith P. L.; Maloney K. N.; Pothen R. G.; Clardy J.; Clapham D. E. (2006) Bisandrographolide from Andrographis paniculata activates TRPV4 channels. J. Biol. Chem. 281, 29897–29904. [DOI] [PubMed] [Google Scholar]