

Abstract
Musical behaviours such as dancing, singing and music production, which require the ability to entrain to a rhythmic beat, encourage high levels of interpersonal coordination. Such coordination has been associated with increased group cohesion and social bonding between group members. Previously, we demonstrated that this association influences even the social behaviour of 14-month-old infants. Infants were significantly more likely to display helpfulness towards an adult experimenter following synchronous bouncing compared with asynchronous bouncing to music. The present experiment was designed to determine whether interpersonal synchrony acts as a cue for 14-month-olds to direct their prosocial behaviours to specific individuals with whom they have experienced synchronous movement, or whether it acts as a social prime, increasing prosocial behaviour in general. Consistent with the previous results, infants were significantly more likely to help an experimenter following synchronous versus asynchronous movement with this person. Furthermore, this manipulation did not affect infant's behaviour towards a neutral stranger, who was not involved in any movement experience. This indicates that synchronous bouncing acts as a social cue for directing prosociality. These results have implications for how musical engagement and rhythmic synchrony affect social behaviour very early in development.
Keywords: interpersonal synchrony, infancy, social development, music, rhythm
1. Introduction
Musical engagement is an important social experience throughout our lives [1,2]. Even during childhood, engaging in musical activities often occurs in a social context. Infants are rocked to sleep to their mother's lullaby, and preschoolers chant nursery rhymes with schoolmates while jumping rope [3,4]. Across cultures, musical engagement such as singing, dancing or playing musical instruments is almost always present at celebrations and religious ceremonies where in-group social affiliation is emphasized [1,5].
When we engage in musical activities with others, our movements become temporally aligned with our group members as each individual entrains to the underlying beat of the music [6]. This type of interpersonal synchrony encourages prosocial behaviour among those involved [7–14], even in infants [13]. However, it is still not clear whether interpersonal synchrony encourages prosocial behaviour indiscriminately or whether it is specific to those with whom temporal synchrony has been previously established. The assumptions of the social cohesion model of musical behaviour suggest that interpersonal synchrony should act as a cue to specifically direct prosociality towards individuals with whom the interpersonal synchrony was experienced [2]. Alternatively, it may act as a social prime [15,16], which would enhance prosociality even towards individuals with whom interpersonal synchrony was not experienced. This study investigates this distinction in 14-month-old infants, the youngest age group in which an effect of synchronous movement on prosocial behaviour has been measured to date [13].
(a). Auditory–motor interactions and rhythmic entrainment
Rhythmic entrainment relies on our ability to extract the underlying beat from a piece of music. Even neonates seem capable of perceiving the beat [17] and people become quite adept at this task by adulthood [6,18,19]. Coordinating movements to an auditory beat requires the ability to predict the onset times of future beats in order for the motor system to plan and execute movements at the appropriate times. Infants cannot control their movements well enough to synchronize to a beat, but at 5 months of age, infants engage in more rhythmic movements when listening to music or drumbeats compared with when listening to speech [20]. Rhythmic entrainment improves over childhood [18] such that adults are able to accurately tap along to a beat when it is in a musically relevant tempo (approx. 300–1800 ms inter-onset intervals) [19,21,22].
While perceiving auditory rhythms primes the motor system for movement [23–25], the way we move also influences how we perceive the beat. For example, adults are better at detecting when a drumbeat in a sequence occurs earlier or later than expected if they are tapping along as opposed to simply listening [26,27]. Movement can also affect the perception of ambiguous metrical patterns (without physical accents) in which accented beats could be perceived to be on either every second beat (as in a march) or on every third beat (as in a waltz). In seven-month-old infants as well as adults, moving on every second versus on every third beat of such metrically ambiguous patterns encourages participants to perceive illusory auditory accents that are congruent with their movement [28,29]. Perhaps because of this complicated interplay between the auditory and motor systems, humans are one of the few species that can successfully synchronize movements to a musical beat [30,31]. The ability to align movements with the timing of musical beats makes music a powerful tool for facilitating interpersonal synchrony and accompanying social effects.
(b). Interpersonal synchrony encourages prosocial behaviour
A growing body of literature suggests that moving in synchrony with others promotes prosocial behaviour. In musical contexts, for example, individuals instructed to sing or drum together are later more likely to help one another [32] or cooperate when presented with an economic dilemma [7,11]. Additionally, children who incorporate joint music making in their play have a higher propensity to spontaneously help each other and collaboratively solve a task compared with children who engage in non-musical play [9]. These findings support the social cohesion model of musical behaviour, which suggests that musical behaviour is evolutionarily adaptive as it enhances prosociality among group members. Such increased group cohesion would be adaptive for individuals if it maintained high in-group reciprocity [2,33].
Increased prosocial behaviour following interpersonal synchrony has been noted also in non-musical settings. For example, when participants rated the similarity of two experimenters who waved together, the degree of temporal coordination predicted ratings of inter-experimenter similarity [34]. Comparably, experimenter–participant synchrony during a tapping task predicted participants' compassion responses [14] and reported affiliation ratings toward the experimenter [8]. Also, adults have a greater expectation that their group members will cooperate with them after walking in-step versus out-of-step with one another [7]. Interpersonal synchrony may also facilitate aspects of social interaction other than cooperation and trust, such as the refining of motor skills required to react appropriately during a joint action task [12].
Interpersonal rhythmic synchrony is considered to be a social experience even by young infants. In a previous study, we investigated whether the prosocial behaviour of 14-month-old infants could be influenced by interpersonal rhythmic synchrony [13]. Infants were held by an assistant in a child carrier facing forwards and bounced gently to the beat of a song for 2.5 min. At the same time, the experimenter faced the infant and also bounced, either in- or out-of-synchrony with the way the infant was bounced by the assistant. Subsequently, we used instrumental helping tasks to measure infants' willingness to aid the experimenter after she accidently dropped the objects needed to complete an intended goal. The number of objects that the infant handed back to the experimenter indexed the degree of prosociality. Infants were significantly more likely to hand back the objects if they had been bounced synchronously compared to if they had been bounced asynchronously with the experimenter [13]. Thus, the effect of synchrony on prosocial behaviour can be measured early in development.
Various theories attempt to explain why interpersonal synchrony promotes prosocial behaviour. One idea is that non-musical temporal coordination of movement is linked with the establishment of a social unit. When a person's movements are temporally aligned with those of another, the perception of self and other might closely overlap, interfering with the ability to differentiate between goals involving the self and the other [8,35]. Expanding this hypothesis is the argument that the main function of synchrony involves increasing perceived similarity or entitativity [14,36]. A second idea is that prosocial behaviour resulting from synchronous activities arises from greater mutual attentiveness [10,37]. For example, dance partners who coordinate their movements to the same song are more likely to remember visual details about each other afterwards [10]. In this case, increased cooperation following synchrony may simply be a product of increased communication among the participants [11]. A third idea is that the association between synchrony and reward may explain the link between increased cooperativeness and synchrony. Brain activity in the caudate, a region associated with reward, increases when participants are asked to engage in coordinated activities [32]. These hypotheses are not mutually exclusive, and a number of factors may drive the prosocial effects of interpersonal synchrony.
However, all of these hypotheses rest on the assumption that interpersonal synchrony generates feelings of prosociality that are directed towards the group members with whom synchrony was experienced. An alternative explanation is that interpersonal synchrony primes prosocial behaviour in general. In other words, individuals may be more likely to behave prosocially following interpersonally synchronous movement, even towards a person with whom they have not experienced synchronous movements. Such social priming using visual or linguistic cues alters the general social behaviour of both adults and young infants [15,38]. Additionally, mimicry (which is similar to synchrony) enhances general as well as specifically directed prosociality in adults [39] and infants [16], therefore working as a social prime rather than a social cue. If this alternative hypothesis were supported, it would imply that interpersonal synchrony works more as a social prime than a social cue, and does not lead to differentiation between in-group and out-group individuals. One recent study investigating this question using groups of adult participants did not find differences in helpfulness following synchronous movement between group and non-group members [40]. However, ‘prosociality’ was measured as the amount of time spent helping an experimenter by completing a questionnaire, which may not be a sensitive measure. Indeed, with this measure, the difference in prosocial behaviour following synchronous and asynchronous movement experience was not significant [40].
(c). Purpose
The goal of the present experiment was to investigate whether interpersonal synchrony acts as a cue or a prime for the prosocial behaviour of 14-month-old infants. Instrumental helping behaviours can be reliably measured in a laboratory setting with infants as young as 14 months, because they can typically walk independently to retrieve dropped items, and they understand joint collaborative activities and goal-directed behaviours [41–43]. If interpersonal synchrony acts as a cue for 14-month-olds, then we expected infants to selectively direct their prosocial behaviours only towards the person with whom they experienced interpersonal synchrony, and not towards a neutral stranger who is present throughout the experiment but has no involvement in the movement manipulation. According to the assumptions of the social cohesion model of musical behaviour, we hypothesized that the boost in helpfulness by the 14-month-olds following interpersonal synchrony would be specifically directed towards the person with whom they moved synchronously, thereby supporting the social cue hypothesis. Alternatively, if interpersonal synchrony primes infants to be generally prosocial, then we expected participants to display prosocial behaviour towards not only the person with whom they experienced interpersonal synchrony but also an uninvolved neutral stranger.
To investigate this question, 14-month-old infants participated in two phases of the experiment. The interpersonal movement phase involved exposing infants to either synchronous or asynchronous interpersonal bouncing based on random between-subject assignment. This was followed by the prosocial test phase, during which the infants' prosocial behaviours were measured. Three researchers were involved: (i) the assistant, who held and bounced the infant in an infant carrier during the interpersonal movement phase; (ii) the experimenter, who faced the infant and bounced either in- or out-of-synchrony with the infant during the interpersonal movement phase, and then later performed half of the prosocial test phase tasks; and (iii) the neutral stranger, who sat quietly in the corner reading a book during the interpersonal movement phase, and then later performed half of the prosocial test phase tasks. The roles of experimenter and neutral stranger were counterbalanced between two female researchers. One of these women wore a blue t-shirt while the other wore a yellow t-shirt to help the infant remember their identities.
2. Material and methods
(a). Participants
Thirty 14-month-old infants (15 girls and 15 boys; M age = 14.5 months; s.d. = 0.3 months) were recruited from the Developmental Studies Database at McMaster University. These infants were raised in English-speaking homes and were able to walk unassisted. Ten additional infants who participated in the experiment were excluded due to excessive fussiness.
(b). Stimuli and apparatus
Each infant heard a 140-s musical instrument digital interface version of the Beatle's Twist and Shout. The song was played at 100 beats per minute (BPM) by a Macintosh computer (OSX) through a Denon amplifier (PMA-480R) connected to an audiological loudspeaker (GSI) 6.5 ft. away from the right side of the infant (same stimulus as in Cirelli et al. [13]). Both the infant and the assistant holding the infant heard this song stimulus over the loudspeaker. Through Denon AH-D501 headphones, the experimenter (who bounced either in- or out-of synchrony while facing the infant) listened to the ‘bounce instruction track’ containing woodblock sounds placed at every second downbeat, overlaid on pink noise. The assistant and infant always heard the song track at 100 BPM. In the synchronous condition, the experimenter also heard the bounce instruction track at 100 BPM, whereas in the asynchronous condition the experimenter heard the bounce instruction track at 140 BPM. Even though the bouncing of the assistant and experimenter lined up every 6 s, for the most part they were not synchronized. This was confirmed in movement analyses from Wii data (see below).
(c). Procedure
(i). Phase 1: interpersonal movement phase
When the infant and parent arrived at the laboratory, the assistant interacted with the infant and exposed him or her to the objects that would later be used during the prosocial test phase. The experimenter obtained parental consent and the parent completed a demographics questionnaire as well as three scales (activity, approach and smiling) of the Infant Behaviour Questionnaire (IBQ) [44]. The neutral stranger waited in the sound-attenuating chamber (Industrial Acoustics Co.) where the experiment took place. Thus, neither the experimenter nor the neutral stranger interacted directly with the infant prior to the start of the experiment.
Once the infant was brought into the sound-attenuating chamber, the interpersonal movement phase began. The parent placed their child, facing forwards, in a child carrier worn by the assistant (Infantino Flip 2012 Infantino LLC). The parent then sat in a chair behind the assistant and listened to masking music over headphones. The experimenter stood 4.5 ft. from the assistant, directly facing her and the infant. Both the experimenter and the assistant bounced in a standing position. The neutral stranger did not interact with anyone during this phase, sitting quietly behind the experimenter within the line of sight of the infant and reading a book (see figure 1 for a picture of this set-up). The assistant bounced up and down at the knees (keeping her feet on the floor), reaching the lowest part of her trajectory at the time of every second downbeat in the song played over loudspeakers. If the infant was randomly assigned to the synchronous bouncing condition, the experimenter bounced in a similar manner to the 100 BPM bounce instruction track, and the infant and experimenter bounced synchronously. If the infant was randomly assigned to the asynchronous condition, the experimenter bounced to the incongruent 140 BPM bounce instruction track, and the infant and experimenter bounced asynchronously at different tempos. In the asynchronous movement condition, the assistant holding the infant was instructed to avert her gaze so that the movement of the experimenter did not influence her ability to bounce to the underlying beats in the song played over loudspeakers [45]. The interpersonal movement phase began when the song files started playing and ended when they stopped and was therefore 140 s in duration.
Figure 1.

Experimental set-up for the interpersonal movement phase. The assistant holds the infant facing forwards towards the experimenter, while the neutral stranger sits within the line of the infant's sight, reading silently. (Online version in colour.)
The assistant and the experimenter both wore Nintendo Wii remotes at their waist while they bounced together, so that their vertical motion over time could be recorded with the internal accelerometers using WiiDataCapture_v2.1 (University of Jyväskylä, Toiviainen & Burger [46]). These remotes are often used to measure movement to music, either in individual or group settings (e.g. [47–49]). Cirelli et al. [13] also used this equipment to verify that the bouncing was indeed appropriately synchronous and asynchronous depending on the condition. If two people are bouncing in synchrony with one another, their vertical accelerations over time should be highly correlated (they will be accelerating downwards at a similar rate, changing directions at the same time, accelerating upwards at a similar rate, etc.), whereas if they are bouncing asynchronously, their vertical accelerations over time should not be highly correlated.
(ii). Phase 2: prosocial test phase
During the prosocial test phase, the experimenter and neutral stranger each performed two different instrumental helping tasks and one sharing task with the infant (therefore, four helping tasks in total were required). Half of the infants interacted with the experimenter first and then with the neutral stranger, and half interacted first with the neutral stranger. The order of the tasks themselves was held constant: the clothespin task, paper ball task and sharing task were followed by the marker task, block task and sharing task. Each of the instrumental helping tasks were comprised of three trials during which the experimenter or neutral stranger pretended to accidently drop the object that was required to complete the task. During these tasks, the infant could move freely around the soundbooth and was placed on the floor in the main space by their parent before the tasks began. The infant had a 30 s trial window during which he or she had the opportunity to assist the experimenter by picking up the object and handing it back. During the first 10 s of the trial, the experimenter or neutral stranger reached for the object, eye gaze focused on the object. For the next 10 s, the experimenter or neutral stranger alternated gaze between the infant and object. For the final 10 s, the experimenter or neutral stranger vocalized about the object (i.e. ‘My marker!’ or ‘My clothespin!’). The trial terminated either when the infant handed the object back or when 30 s had elapsed. These tasks were based on the instrumental helping tasks developed by Warneken & Tomasello [42,43].
(1) Clothespin task. In this task, the experimenter or neutral stranger showed the infant a dishcloth, and then clipped one corner of the dishcloth to a clothesline with a plastic clothespin. The first trial was initiated when she attempted to clip up the next corner of the dishcloth, but fumbled and dropped the clothespin to the floor. If the infant handed the clothespin back, the experimenter used it to successfully clip up that corner. If the infant did not hand the clothespin back before the end of the 30 s trial, a new clothespin was used to successfully clip up that corner before the next trial began.
(2) Paper ball task. The experimenter or neutral stranger placed a translucent plastic bucket on a 2.5-ft high table, and held a jar containing six paper balls. She stood behind the table and successfully tossed one of the paper balls into the bucket, cheered and showed the infant. The trial began when she attempted to toss the next paper ball into the bucket, and overshot. If the infant handed the paper ball back, she successfully tossed it in the bucket. If the infant did not hand the paper ball back, she took a new paper ball from the jar and successfully tossed it into the bucket before initiating the next trial.
(3) Marker task. The experimenter or neutral stranger placed a sheet of white paper and four coloured markers on the same table. Kneeling behind the table, the experimenter began to draw a flower with the markers, showing the infant her progress along the way. A trial was initiated when she ‘accidently’ bumped one of the capped markers off the table. If the infant handed the marker back, she used this marker to continue the picture. If the infant did not hand the marker back, she picked up a new marker and used it to continue the picture before initiating the next trial.
(4) Block task. The experimenter or neutral stranger placed two soft baby blocks on the same table beside each other. Standing behind the table, she then grabbed a third block and stacked it on top of this base. The trial was initiated when she attempted to place another block on top of this block, but fumbled the block and dropped it to the ground in front of the table. If the infant handed back the block, she successfully stacked it on the others. If the infant did not hand back the block, a new block was successfully stacked on the others before the next trial was initiated.
(5) Sharing task. The sharing task was based on the request phase of the sharing task used by Schmidt & Sommerville [50]. During each of the sharing tasks (one with the experimenter and one with the neutral stranger), the assistant reentered the sound booth holding a bucket containing two plastic animal toys. She placed the bucket in front of the infant. If the infant did not immediately pick up one of the toys, she attempted to place one in their hand. Once the infant had at least one of the toys in hand, the experimenter or neutral stranger, sitting on the ground in front of the infant, reached out her hand, palm up, towards the infant asking ‘Can I have one? Can I have one please?’ She repeated these questions for 30 s, or until the infant handed her a toy, at which point she looked at her hand and exclaimed pleasantly ‘Alright!’.
(6) Data coding. Two mounted video cameras (a Canon PowerShot SD1000 and a Samsung 65X Intelli-zoom) were used to capture the infant and experimenter behaviour throughout the two phases. Two adult raters blind to the conditions used this video footage to code infant prosocial behaviour. During each trial of the instrumental helping tasks, infants were given one point if they handed back the object. If they attempted but failed to hand the object back, or handed it back after the 30-s trial window had elapsed, they received 0.5 points. For each instrumental helping task, the per trial points were used to assign a total helping score out of 3 to each infant. Inter-rater reliability was extremely high, r = 0.997, p < 0.001. Raters calculated a total score for experimenter-directed helping out of six by adding the total helping scores, each out of three, for each of the two instrumental helping tasks performed by this person. Raters calculated a total score for neutral stranger-directed helping in a similar fashion. For each of these conditions (helping towards the experimenter versus the neutral stranger), the percent helping rate was then calculated (score(task 1) + score(task 2))/2 tasks × 3 trials × 100%. For the sharing tasks, the raters recorded whether or not the infant shared within the 30-s trial window.
3. Results
One infant (a female in the synchrony condition) was excluded from the analysis owing to missing data (parents did not complete the required IBQ). Using a z-score cut-off of ±2, an additional infant (a male in the asynchrony condition) was removed from the analysis as an outlier.
(a). Helping
Consistent with the hypothesis that increased helpfulness is directed toward the person with whom synchronous movement was experienced, infants who experienced synchronous movement were significantly more likely to help the experimenter (mean = 61.3%, s.e.m. = 9.4%) than the neutral stranger (mean = 38.6%, s.e.m. = 8.9%), t13 = 2.39, p < 0.05. Helping rates in the asynchronous condition were low and not significantly different toward the experimenter (mean = 17.4%, s.e.m. = 5.9%) and the neutral stranger (mean = 26.8%, s.e.m. = 9.4%), t13 = −0.93, p = 0.37.
Significant Pearson correlations were found between infants' rates of helping the neutral stranger and parent ratings (as per the IBQ) for ‘smiling’ (r = 0.68, p < 0.01) and ‘approach’ (r = −0.45, p < 0.05). Specifically, infants who were rated by parents as having a more positive disposition and being less likely to shy away from novelty were more likely to display helpfulness towards the neutral stranger. These correlations were not significant experimenter-directed helping rates (smiling: r = 0.30, p = 0.12; approach: r = −0.27, p = 0.16).
An analysis of covariance was conducted with condition (synchronous, asynchronous) as a between-subjects factor and person (experimenter, neutral stranger) as a within-subjects factor, with parent IBQ ratings of ‘smiling’ and ‘approach’ as covariates in the analysis. Results revealed a significant interaction between movement condition and researcher role, F1,24 = 8.23, p < 0.01, 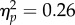 . As seen in figure 2, infants in the synchronous condition were significantly more likely to display helpfulness towards the experimenter than infants from the asynchronous condition (mean = 61.3%, s.e.m. = 9.4%; mean = 17.4%, s.e.m. = 5.9%, respectively), t21.8 = 3.94, p < 0.001, replicating the results of Cirelli et al. [13]. However, the difference between synchronous and asynchronous conditions was not significant in the case of the neutral stranger, t26 = 0.92, p = 0.37. Together, these results show that infants in the synchronous condition were more helpful towards the experimenter than infants in the asynchronous condition, but that this helpfulness did not generalize to the neutral stranger (figure 2).
. As seen in figure 2, infants in the synchronous condition were significantly more likely to display helpfulness towards the experimenter than infants from the asynchronous condition (mean = 61.3%, s.e.m. = 9.4%; mean = 17.4%, s.e.m. = 5.9%, respectively), t21.8 = 3.94, p < 0.001, replicating the results of Cirelli et al. [13]. However, the difference between synchronous and asynchronous conditions was not significant in the case of the neutral stranger, t26 = 0.92, p = 0.37. Together, these results show that infants in the synchronous condition were more helpful towards the experimenter than infants in the asynchronous condition, but that this helpfulness did not generalize to the neutral stranger (figure 2).
Figure 2.
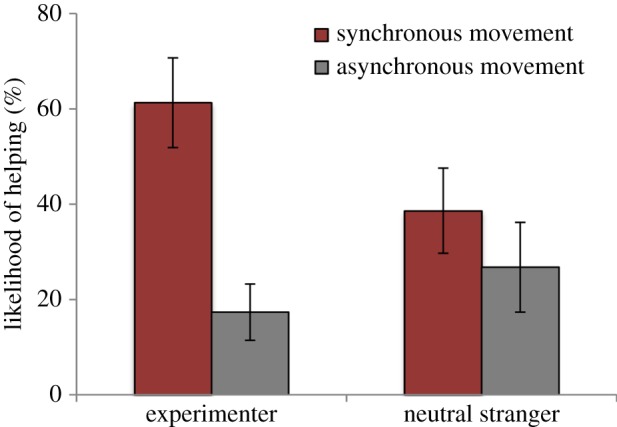
Infant helpfulness towards the experimenter and the neutral stranger by infants in the synchronous compared with the asynchronous condition. Error bars represent standard error of the mean. (Online version in colour.)
(b). Sharing
Two independent sample t-tests were used to investigate the effect of interpersonal synchrony on experimenter- and neutral-stranger-directed sharing. Infants from the synchronous and asynchronous conditions were not significantly different in their sharing rates towards either the experimenter (t26 = −0.82, p = 0.42) or the neutral stranger (t26 = −0.82, p = 0.42). Overall sharing rates were quite high (25 of the 28 infants in the analysis shared with at least one of the researchers), suggesting that a ceiling effect may have reduced the ability to detect differences between conditions.
(c). Experimenter/assistant movement synchrony analysis
Wii remote data were available for 19 of the 28 sessions. With these data, correlations between the assistant and the experimenter's vertical acceleration over time during the interpersonal movement phase were calculated as a measure of interpersonal synchrony (for rationale, see §2c(i)). A 30-s long window of data was selected for 30 s into the interpersonal movement phase. This analysis revealed that the vertical accelerations of the experimenter and assistant were significantly and strongly correlated in the synchrony conditions (mean r = 0.75, s.e.m. = 0.04) and were weak and non-significantly correlated in the asynchrony conditions (mean r = 0.01, s.e.m. = 0.003), as expected. An independent-samples t-test was used to verify that the assistant and experimenter's vertical acceleration was significantly more correlated in the synchronous conditions than in the asynchronous conditions, t17 = 15.50, p < 0.001.
(d). Experimenter consistency
As two researchers took turns playing the role of experimenter and neutral stranger, we wanted to ensure that one researcher was not driving the synchrony effect. An ANOVA with condition (synchronous, asynchronous) and researcher role (Researcher A playing the experimenter, Researcher B playing the experimenter) as two between-subjects factors revealed that, as predicted, there was no main effect of researcher role, F1,24 = 0.42, p = 0.84, or interaction between the two variables, F1,24 = 0.27, p = 0.61. This validates the assumption that it was not a single researcher driving the effect of synchrony on experimenter-directed helping.
(e). Post hoc video rating
Adult raters naive to the hypotheses of the experiment (n = 8) watched eight pairs of videos. Each video displayed the experimenter or neutral stranger's behaviours during one trial of the prosocial test phase (the second trial of the second task). Videos were paired so that one researcher's behaviour with an infant from the synchronous condition and an infant from the asynchronous condition were always compared. The videos were also matched so that (i) the two videos always showed the same researcher, (ii) the researcher always played the same role (experimenter or neutral stranger) and (iii) both infants either helped or did not help. After watching both videos from one of the eight pairs, raters responded to the question: ‘Which baby does the experimenter seem to prefer?’ Supporting our assumption that the researchers behaved consistently between conditions (synchronous and asynchronous), for both researchers, the raters showed no evidence that the researcher preferred the infants in either the synchronous or asynchronous condition, t7 = 0.11, p = 0.91 for Researcher A and t7 = −1.80, p = 0.11 for Researcher B.
4. Discussion
The results support the hypothesis that interpersonal rhythmic synchrony acts as a social cue, as opposed to a social prime. Infants who were bounced synchronously with the experimenter were more likely to help that experimenter than a neutral stranger with whom they did not experience synchronous movement. Furthermore, infants in the synchronous bouncing condition were significantly more likely to help the experimenter than infants in the asynchronous bouncing condition, but this was not the case for helping directed toward the neutral stranger. Interestingly, socially relevant personality measures from the parent report IBQ (smiling and approach) correlated significantly with helping directed at the neutral stranger but not helping directed at the experimenter, which was driven instead by the experimental manipulation. This suggests that experiencing synchrony with an adult overrides infants' intrinsic tendencies to be more or less helpful in general. The results indicate that interpersonal rhythmic synchrony affects the affiliation between those directly involved in the movement experience. This supports assumptions of the social cohesion model of musical behaviour by suggesting that in-group affiliation is enhanced by interpersonal synchrony, which is facilitated by mutual musical engagement.
These findings appear to be in conflict with those of Reddish et al. [40], who concluded from their results that interpersonal synchrony may lead to generalized prosociality. However, Reddish et al. [40] tested adults, whereas infants were tested in this study, so it is possible that the effect is person-specific early in development but becomes generalized by adulthood. On the other hand, the conflicting conclusions may be due to other factors such as differences in the sensitivity of the measures of prosocial behaviour across the two studies. This study used several tasks, all of which had previously been validated with infants [42,43]. Redding et al. [40] used a single measure, the time participants spent filling out forms for the experimenter, which might not have optimally captured participants' prosocial feelings. A further difference between the studies is whether the synchrony was experienced between dyads or within a group. It is possible that prosocial outcomes directed at individuals are strongest when the synchrony is experienced as a dyad. In any case, further studies with adults should be conducted in order to clarify these discrepancies.
In this study, only instrumental helping and not sharing behaviours were influenced by the experimental manipulations. This might be due to a ceiling effect for the sharing measure. However, there is evidence that helping and sharing represent different types of social engagement and do not necessarily correlate with one another [51,52]. One clear difference between sharing and instrumental helping is in the goal-directed nature of such behaviours. While sharing typically targets the understanding of even distribution of goods, instrumental helping targets the understanding of goal-directed behaviours [51]. Perhaps the joint activity inherent in interpersonal synchrony specifically encourages prosocial behaviours that rely on the perception of joint action, as would be expected in instrumental helping behaviours. In addition, this sharing measure may have been inappropriate for the age group in question. In the first year after birth, infants tend to share resources indiscriminately. By 2 years of age, children become more selective about whom they share with and start sharing at a cost to themselves [53]. It is possible that our ceiling effect represents this indiscriminate sharing behaviour that is typical of this younger age group [54]. The differential effects of synchrony on sharing and instrumental helping measures in this study highlight the fact that the way prosociality is measured must be clearly defined in studies of interpersonal synchrony.
It is interesting to note that the movement experience by the infants was passively generated. The infants were not moving to the music on their own; rather, the assistant was passively bouncing them. In older age groups, the effect of synchrony on prosociality has been measured when participants engage in self-generated active movements that align temporally with the movement of others [7–12]. In the present experiment and in our previous investigation [13], the effect was measured in 14-month-olds who experienced passive movement while being held in a carrier and bounced by an adult. Passive movements have been shown to influence beat and meter perception in adults and infants [28,55], so it would be interesting to investigate in future research if passive movements can encourage prosociality in adulthood as they do in infancy.
The present results are also of interest when considering the differences between the social implications of mimicry and interpersonal synchrony. While mimicry and synchrony are similar concepts and both can drive prosociality [8,39], they are different phenomena. Unlike in synchrony, in mimicry actions are not synchronous in time, but follow each other [38,56,57]. In addition, while synchrony is quite often conscious and intentional, mimicry encourages prosocial effects only if it is undetected by the mimicked individual, and most often happens unconsciously [12,14,58]. Specifically relevant to the above findings, 18-month-old infants who are mimicked by an experimenter are later more likely to display helpfulness towards either that experimenter or a neutral experimenter [16]. In light of these findings, it could be argued that while mimicry acts as an unconscious social prime, interpersonal synchrony is instead an overt and intentional cue for self-other similarity.
One question for future research concerns the generalizability of interpersonal synchrony as a social cue. The present results suggest that interpersonal synchrony does not lead to generalized prosociality with strangers, and is therefore not simply a social prime. However, as a social cue providing information about a specific individual, interpersonal synchrony might still encourage a more restricted kind of generalization. More specifically, the present study investigated the role of synchrony towards an experimenter and a neutral stranger, individuals who did not interact with each other. If the neutral stranger was not completely neutral, but was instead perceived to be a member of the same group as the bouncing experimenter, prosociality may generalize; infants bounced in synchrony with an experimenter may be more helpful to other members of the experimenter's group, even if they did not bounce with them. The social cohesion model of musical behaviour speaks about group behaviour, and as such, the social cue provided by interpersonal synchrony might also be a cue containing information about a person's group. This is an interesting question for future research.
5. Conclusion
The present findings replicate the previous report that interpersonal synchrony increases prosocial behaviour in infants [13]. They additionally support the hypothesis that interpersonal synchrony acts as a cue to direct prosocial behaviour toward individuals rather than as a prime for generalized prosocial behaviour. Future research could benefit from a focus on the role of music in synchronized movements and on the development of methodologies for determining whether experiencing synchronous movement affects social behaviour in infants younger than 14 months of age.
Supplementary Material
Acknowledgements
L.K.C. was the primary researcher and L.J.T. the senior researcher but all authors contributed to the ideas, analyses and writing of the manuscript. L.K.C. and S.J.W. tested the participants, along with Christina Spinelli who we would like to formally acknowledge and thank. We also thank Fiona Manning for comments on an earlier draft.
Ethics statement
Consent was obtained from parents, as per the McMaster Research Ethics Board (MREB) guidelines.
Data accessibility
The dataset associated with this paper is available in the electronic supplementary material.
Funding statement
This research was funded by a grant from the Natural Sciences and Engineering Research Council of Canada to L.J.T. (197033-2009) and by the Social Sciences and Humanities Research Council to L.K.C.
References
- 1.Dissanayake E. 2006. Ritual and ritualization: musical means of conveying and shaping emotion in humans and other animals. In Music and manipulation: on the social uses and social control of music (eds Brown S, Voglsten U.), pp. 31–56. New York, NY: Berghahn Books. [Google Scholar]
- 2.Brown S. 2000. Evolutionary models of music: from sexual selection to group selection. In Perspectives in ethology, volume 14: evolution, culture and behaviour (eds Tonneau S, Thompson NS.), pp. 231–281. New York, NY: Kluwer Academic Publishers. [Google Scholar]
- 3.Trainor LJ, Hannon EE. 2012. Musical development. In The psychology of music (ed Deutsch D.), pp. 423–498. London, UK: Elsevier Inc. [Google Scholar]
- 4.Trehub S, Trainor L. 1998. Singing to infants: lullabies and play songs. Adv. Infancy Res. 12, 43–77. [Google Scholar]
- 5.Freeman WJ. 2000. A neurobiological role of music in social bonding. In The origins of music (eds Wallin N, Merkur B, Brown S.), pp. 1–13. Cambridge, MA: MIT Press. [Google Scholar]
- 6.Repp B. 2006. Musical synchronization. In Music, motor control, and the brain (eds Altenmüller E, Wiesendanger W, Kesselring J.), pp. 55–76. Oxford, UK: Oxford University Press. [Google Scholar]
- 7.Wiltermuth SS, Heath C. 2009. Synchrony and cooperation. Psychol. Sci. 20, 1–5. ( 10.1111/j.1467-9280.2008.02253.x) [DOI] [PubMed] [Google Scholar]
- 8.Hove MJ, Risen JL. 2009. It's all in the timing : interpersonal synchrony increases affiliation. Soc. Cogn. 27, 949–960. ( 10.1521/soco.2009.27.6.949) [DOI] [Google Scholar]
- 9.Kirschner S, Tomasello M. 2010. Joint music making promotes prosocial behavior in 4-year-old children. Evol. Hum. Behav. 31, 354–364. ( 10.1016/j.evolhumbehav.2010.04.004) [DOI] [Google Scholar]
- 10.Woolhouse M, Tidhar D. 2010. Group dancing leads to increased person-perception. In Proc. 11th Int. Conf. Music Percept. Cogn. 23–27 August 2010, Seattle, WA (eds SM Demorest, SJ Morrison, PS Campbell), pp. 605–608. [Google Scholar]
- 11.Anshel A, Kippler D. 1988. The influence of group singing on trust and cooperation. J. Music Ther. 25, 145–155. ( 10.1093/jmt/25.3.145) [DOI] [Google Scholar]
- 12.Valdesolo P, Ouyang J, DeSteno D. 2010. The rhythm of joint action: synchrony promotes cooperative ability. J. Exp. Soc. Psychol. 46, 693–695. ( 10.1016/j.jesp.2010.03.004) [DOI] [Google Scholar]
- 13.Cirelli LK, Einarson KM, Trainor LJ. In press Interpersonal synchrony increases prosocial behavior in infants. Dev. Sci. ( 10.1111/desc.12193) [DOI] [PubMed]
- 14.Valdesolo P, Desteno D. 2011. Synchrony and the social tuning of compassion. Emotion 11, 262–266. ( 10.1037/a0021302) [DOI] [PubMed] [Google Scholar]
- 15.Over H, Carpenter M. 2009. Eighteen-month-old infants show increased helping following priming with affiliation. Psychol. Sci. 20, 1189–1193. ( 10.1111/j.1467-9280.2009.02419.x) [DOI] [PubMed] [Google Scholar]
- 16.Carpenter M, Uebel J, Tomasello M. 2013. Being mimicked increases prosocial behavior in 18-month-old infants. Child. Dev. 84, 1–8. ( 10.1111/cdev.12083) [DOI] [PubMed] [Google Scholar]
- 17.Winkler I, Háden GP, Ladinig O, Sziller I, Honing H. 2009. Newborn infants detect the beat in music. Proc. Natl Acad. Sci. USA 106, 2468–2471. ( 10.1073/pnas.0809035106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Drake C, Jones MR, Baruch C. 2000. The development of rhythmic attending in auditory sequences: attunement, referent period, focal attending. Cognition 77, 251–288. ( 10.1016/S0010-0277(00)00106-2) [DOI] [PubMed] [Google Scholar]
- 19.Mates J, Müller U, Radil T, Pöppel E. 1994. Temporal integration in sensorimotor synchronization. J. Cogn. Neurosci. 6, 332–340. ( 10.1162/jocn.1994.6.4.332) [DOI] [PubMed] [Google Scholar]
- 20.Zentner M, Eerola T. 2010. Rhythmic engagement with music in infancy. Proc. Natl Acad. Sci. USA 107, 5768–5773. ( 10.1073/pnas.1000121107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fraisse P. 1982. Rhythm and tempo. In The psychology of music (ed. Deutsch D.), pp. 149–180. New York, NY: Academic Press. [Google Scholar]
- 22.Drake C, Botte MC. 1993. Tempo sensitivity in auditory sequences: evidence for a multiple-look model. Percept. Psychophys. 54, 277–286. ( 10.3758/BF03205262) [DOI] [PubMed] [Google Scholar]
- 23.Grahn JA, Brett M. 2007. Rhythm and beat perception in motor areas of the brain. J. Cogn. Neurosci. 19, 893–906. ( 10.1162/jocn.2007.19.5.893) [DOI] [PubMed] [Google Scholar]
- 24.Fujioka T, Trainor LJ, Large EW, Ross B. 2012. Internalized timing of isochronous sounds is represented in neuromagnetic β oscillations. J. Neurosci. 32, 1791–1802. ( 10.1523/JNEUROSCI.4107-11.2012) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zatorre RJ, Chen JL, Penhune VB. 2007. When the brain plays music: auditory-motor interactions in music perception and production. Nat. Rev. Neurosci. 8, 547–558. ( 10.1038/nrn2152) [DOI] [PubMed] [Google Scholar]
- 26.Manning F, Schutz M. 2013. ‘Moving to the beat’ improves timing perception. Psychon. Bull. Rev. 20, 1133–1139. ( 10.3758/s13423-013-0439-7) [DOI] [PubMed] [Google Scholar]
- 27.Butler B, Trainor LJ. 2011. Examining the role of training and movement on rhythm perception in disc jockeys using EEG and behavioural thresholds. Presented at the Conf. Soc. Music Perception and Production, Rochester, NY, USA,11 August 2011. [Google Scholar]
- 28.Phillips-Silver J, Trainor LJ. 2005. Feeling the beat: movement influences infant rhythm perception. Science 308, 1430 ( 10.1126/science.1110922) [DOI] [PubMed] [Google Scholar]
- 29.Phillips-Silver J, Trainor LJ. 2007. Hearing what the body feels: auditory encoding of rhythmic movement. Cognition 105, 533–546. ( 10.1016/j.cognition.2006.11.006) [DOI] [PubMed] [Google Scholar]
- 30.Schachner A, Brady TF, Pepperberg IM, Hauser MD. 2009. Spontaneous motor entrainment to music in multiple vocal mimicking species. Curr. Biol. 19, 831–836. ( 10.1016/j.cub.2009.03.061) [DOI] [PubMed] [Google Scholar]
- 31.Patel AD, Iversen JR, Bregman MR, Schulz I. 2009. Experimental evidence for synchronization to a musical beat in a nonhuman animal. Curr. Biol. 19, 827–830. ( 10.1016/j.cub.2009.03.038) [DOI] [PubMed] [Google Scholar]
- 32.Kokal I, Engel A, Kirschner S, Keysers C. 2011. Synchronized drumming enhances activity in the caudate and facilitates prosocial commitment—if the rhythm comes easily. PLoS ONE 6, e27272 ( 10.1371/journal.pone.0027272) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gintis H, Bowles S, Boyd R, Fehr E. 2003. Explaining altruistic behavior in humans. Evol. Hum. Behav. 24, 153–172. ( 10.1016/S1090-5138(02)00157-5) [DOI] [Google Scholar]
- 34.Lakens D. 2010. Movement synchrony and perceived entitativity. J. Exp. Soc. Psychol. 46, 701–708. ( 10.1016/j.jesp.2010.03.015) [DOI] [Google Scholar]
- 35.Georgieff N, Jeannerod M. 1998. Beyond consciousness of external reality: a ‘who’ system for consciousness of action and self-consciousness. Conscious. Cogn. 7, 465–477. ( 10.1006/ccog.1998.0367) [DOI] [PubMed] [Google Scholar]
- 36.Lakens D, Stel M. 2011. If they move in sync, they must feel in sync: movement synchrony leads to attributions of rapport and entitativity. Soc. Cogn. 29, 1–14. ( 10.1521/soco.2011.29.1.1) [DOI] [Google Scholar]
- 37.Macrae CN, Duffy OK, Miles LK, Lawrence J. 2008. A case of hand waving: action synchrony and person perception. Cognition 109, 152–156. ( 10.1016/j.cognition.2008.07.007) [DOI] [PubMed] [Google Scholar]
- 38.Lakin JL, Chartrand TL. 2003. Using nonconscious behavioral mimicry to create affiliation and rapport. Psychol. Sci. 14, 334–339. ( 10.1111/1467-9280.14481) [DOI] [PubMed] [Google Scholar]
- 39.Van Baaren RB, Holland RW, Kawakami K, Knippenberg AV. 2004. Mimicry and prosocial behavior. Psychol. Sci. 15, 71–74. ( 10.1111/j.0963-7214.2004.01501012.x) [DOI] [PubMed] [Google Scholar]
- 40.Reddish P, Bulbulia J, Fischer R. 2013. Does synchrony promote generalized prosociality? Religion Brain Behav. 4, 1–17. ( 10.1080/2153599X.2013.764545) [DOI] [Google Scholar]
- 41.Tomasello M, Carpenter M, Call J, Behne T, Moll H. 2005. Understanding and sharing intentions: The origins of cultural cognition. Behav. Brain Sci. 28, 675–735. ( 10.1017/S0140525X05000129) [DOI] [PubMed] [Google Scholar]
- 42.Warneken F, Tomasello M. 2006. Altruistic helping in human infants and young chimpanzees. Science 311, 1301–1303. ( 10.1126/science.1121448) [DOI] [PubMed] [Google Scholar]
- 43.Warneken F, Tomasello M. 2007. Helping and cooperation at 14 months of Age. Infancy 11, 271–294. ( 10.1111/j.1532-7078.2007.tb00227.x) [DOI] [PubMed] [Google Scholar]
- 44.Rothbart MK. 1981. Measurement of temperament in infancy. Child Dev. 52, 569–578. ( 10.2307/1129176) [DOI] [Google Scholar]
- 45.Lucas G, Clayton M, Leante L. 2011. Inter-group entrainment in Afro-Brazilian congado ritual. Emp. Musicol. Rev. 44, 75–102. [Google Scholar]
- 46.Burger B, Toiviainen P. 2013. Toolbox—a Matlab toolbox for computational analysis of movement data. In Proc. 10th Sound and Music Computing Conference (ed. R Bresin). Stockholm, Sweden: KTH Royal Institute of Technology. [Google Scholar]
- 47.De Bruyn L, Leman M, Demey M, Desmet F, Moelants D. 2008. Measuring and quantifying the impact of social interaction on listeners’ movement to music. In Proc. 5th Int. Symp. Computer Music Modeling and Retrieval: Genesis of Meaning in Sound and Music (eds S Ystad, R Kronland- Martinet, K Jensen), pp. 298–305. Berlin, Germany: Springer. [Google Scholar]
- 48.Phillips-Silver J, Toiviainen P, Gosselin N, Piché O, Nozaradan S, Palmer C, Peretz I. 2011. Born to dance but beat deaf: A new form of congenital amusia. Neuropsychologia 49, 961–969. ( 10.1016/j.neuropsychologia.2011.02.002) [DOI] [PubMed] [Google Scholar]
- 49.Burger B, Toiviainen P. 2013. MoCap Toolbox—a matlab toolbox for computational analysis of movement data. In Proceedings of the 10th Sound and Music Computing Conference (ed. Bresin R.). Stockholm, Sweden: KTH Royal Institute of Technology. [Google Scholar]
- 50.Schmidt MFH, Sommerville JA. 2011. Fairness expectations and altruistic sharing in 15-month-old human infants. PLoS ONE 6, e23223 ( 10.1371/journal.pone.0023223) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dunfield K, Kuhlmeier VA, O'Connell L, Kelley E. 2011. Examining the diversity of prosocial behavior: helping, sharing, and comforting in infancy. Infancy 16, 227–247. ( 10.1111/j.1532-7078.2010.00041.x) [DOI] [PubMed] [Google Scholar]
- 52.Paulus M, Kühn-Popp N, Licata M, Sodian B, Meinhardt J. 2013. Neural correlates of prosocial behavior in infancy: different neurophysiological mechanisms support the emergence of helping and comforting. NeuroImage 66, 522–530. ( 10.1016/j.neuroimage.2012.10.041) [DOI] [PubMed] [Google Scholar]
- 53.Brownell CA, Svetlova M, Nichols S. 2009. To share or not to share: when do toddlers respond to another's needs? Infancy 14, 117–130. ( 10.1080/15250000802569868) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hay DF, Caplan M, Castle J, Stimson CA. 1991. Does sharing become increasingly ‘rational’ in the second year of life? Dev. Sci. 27, 987. [Google Scholar]
- 55.Phillips-Silver J, Trainor LJ. 2008. Vestibular influence on auditory metrical interpretation. Brain Cogn. 67, 94–102. ( 10.1016/j.bandc.2007.11.007) [DOI] [PubMed] [Google Scholar]
- 56.Keller P. 2008. Joint action in music performance. Emerg. Commun. 10, 205–221. [Google Scholar]
- 57.Sebanz N, Knoblich G. 2009. Prediction in joint action: what, when, and where. Top. Cogn. Sci. 1, 353–367. ( 10.1111/j.1756-8765.2009.01024.x) [DOI] [PubMed] [Google Scholar]
- 58.Ashton–James C, van Baaren RB, Chartrand TL, Decety J, Karremans J. 2007. Mimicry and me: the impact of mimicry on self-construal. Soc. Cogn. 25, 518–535. ( 10.1521/soco.2007.25.4.518) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The dataset associated with this paper is available in the electronic supplementary material.