

Abstract
While the fundamental trade-off in leaf traits related to carbon capture as described by the leaf economics spectrum is well-established among plant species, the relationship of the leaf economics spectrum to stem hydraulics is much less known. Since carbon capture and transpiration are coupled, a close connection between leaf traits and stem hydraulics should be expected. We thus asked whether xylem traits that describe drought tolerance and vulnerability to cavitation are linked to particular leaf traits. We assessed xylem vulnerability, using the pressure sleeve technique, and anatomical xylem characteristics in 39 subtropical tree species grown under common garden conditions in the BEF-China experiment and tested for correlations with traits related to the leaf economics spectrum as well as to stomatal control, including maximum stomatal conductance, vapor pressure deficit at maximum stomatal conductance and vapor pressure deficit at which stomatal conductance is down-regulated. Our results revealed that specific xylem hydraulic conductivity and cavitation resistance were closely linked to traits represented in the leaf economic spectrum, in particular to leaf nitrogen concentration, as well as to log leaf area and leaf carbon to nitrogen ratio but not to any parameter of stomatal conductance. The study highlights the potential use of well-known leaf traits from the leaf economics spectrum to predict plant species' drought resistance.
Introduction
The worldwide leaf economics spectrum (LES) represents an important framework of trade-offs between key functional leaf traits [1]. It describes different strategies of carbon capture among vascular land plants, from that of short-lived leaves with high photosynthetic capacity per leaf mass, to long-lived leaves with low mass-based carbon assimilation rates. Mass-based photosynthetic capacity is positively related to mass-based leaf nitrogen concentration (LNC) and to specific leaf area [2] and is negatively related to leaf life span [1]. Many studies have confirmed the global validity of these trade-off patterns [3]–[5], and Osnas et al. [6] recently demonstrated that such relationships between traits in the LES result from relationships to leaf area and from normalizing area-proportional traits by leaf mass. Kröber & Bruelheide [7] have demonstrated that there are additional dimensions to plants' functional traits that are orthogonal to the LES. They found parameters of stomatal regulation (derived from stomatal conductance - vapor pressure deficit relationships), stomatal density and stomatal size to be independent from the LES.
So far, reported relationships between traits of the LES and those of plant organs other than leaves, such as wood or roots, are equivocal. For example, Baraloto et al. [8] found the main axes in leaf and wood traits to be decoupled, while Freschet et al. [9] provided evidence for a tight relationship between the main dimensions of the leaf, stem and root economics spectra. However, from a 'whole plant' perspective, tight relationships would be expected for those leaf and wood traits that determine a plant's tolerated minimum water potential, because the plant's water status links a multitude of physiological processes [10]. Choat et al. [11] and Poorter et al. [12] reported that leaves with high specific leaf area (SLA) were linked to stems with low wood density. Similar to wood density, hydraulic xylem properties would also be expected to be correlated across roots, stems and leaves. In particular, a high photosynthetic capacity of leaves, as expressed by high SLA, should be associated with high xylem hydraulic conductivity to facilitate sufficient water supply required for high stomatal conductance. Accordingly, in a study on ten tropical tree species in Panama, Sack and Frole [13] reported that leaf hydraulic resistance was strongly linked to leaf venation and mesophyll structure. The relationship between hydraulic conductivity and leaf venation was also confirmed across 43 species worldwide [14]. However, Sack et al. [15] argued that leaf hydraulic conductance might be mechanistically independent from the LES, but might be linked statistically as both hydraulic conductance and LES traits affect mass-based photosynthesis.
Besides being hydraulically efficient, another required feature of the vascular plumbing network is drought resistance. In this regard, species with stress-resistant leaves, as indicated by low SLA values, should be expected to have stress-resistant wood. Wood stress resistance is reflected in high wood density, which is thought to confer a higher tolerance from shade, wind, herbivores and drought [16]. In particular, drought resistance determined by measuring xylem vulnerability to cavitation should be correlated between leaves and wood, because cavitation is a persistent hazard under drought stress and affects leaves and wood [17]. Sustaining low water potentials requires high cavitation resistance of conduits, as derived from xylem vulnerability curves [18]. These curves allow quantifying the specific xylem hydraulic conductivity of the xylem (KS) and the xylem pressure at which 50% loss of the maximum specific xylem hydraulic conductivity occurs (Ψ50). Ψ50 is mainly determined by pit size and structure [19],[20]. However, low water potentials are transmitted throughout the whole plant, from the point where the water-pathway ends and the regulation of the water flow takes place to the xylem, where cavitation occurs. Thus, Ψ50 should be reflected in functional leaf traits and parameters of stomatal regulation.
Tree species that are able to endure severe drought periods have characteristic leaves. The leaves are tough and have a high leaf dry matter content (LDMC), allowing them to sustain low water potentials [21],[22]. Such species should likewise be characterized by low Ψ50 values. SLA is inversely related to LDMC, in that it decreases with drought resistance [23],[24], and it would be expected to scale negatively with Ψ50. In addition, cavitation vulnerability should also be related to stomatal regulation, because cavitation-sensitive and -insensitive species would be expected to close their stomata at low and high vapor pressure deficits, respectively [25]–[27]. Such parameters of stomatal closure have recently been provided by Kröber & Bruelheide [7] for the same 39 species used also in the current study. The authors measured daily courses of stomatal conductance (gs) with porometry in the same plots as in the present study, and modeled the species-specific gs ∼ vapor pressure deficit (VPD) relationships. They found that mean gs can be predicted from leaf traits that reflect the LES, with a positive relationship to LNC and a negative relationship to leaf carbon to nitrogen ratio. In contrast, the maximum of the gs ∼ VPD curve was unrelated to traits of the LES and increased with leaf carbon concentration (LCC) and vein length. The VPD at which gs was down-regulated, characterized by the point of inflection of the gs ∼ VPD curve at high VPD, was lower for species with higher stomatal density and lower leaf carbon concentration. In addition to leaf trait measurements, we use these parameters of stomatal control from Kröber & Bruelheide [7] to predict xylem hydraulics.
The objective of our study was to quantify hydraulic conductivity and Ψ50 from xylem vulnerability curves, making use of the common garden situation of the BEF-China experiment. Comparing 39 broad-leaved tree species, we hypothesized that (1) leaf traits describing the leaf economics spectrum are related to specific xylem hydraulic conductivity and cavitation resistance. Accordingly, we expected that (2) evergreen species characterized by low SLA and high LDMC are more resistant to cavitation, i.e. have lower Ψ50 values than deciduous species, Finally, (3) we tested the hypothesis that parameters of stomatal regulation, such as maximum stomatal conductance, the vapor pressure deficit (VPD) at maximum stomatal conductance and VPD at which stomatal conductance is down-regulated, are related to high xylem hydraulic conductivity.
Materials and Methods
Study Site
The study was conducted in the BEF-China project, which is a biodiversity-ecosystem functioning experiment based in Jiangxi Province, southeast China (http://www.bef-china.de; 29.08–29.11 N, 117.90–117.93 E). The climate at the experimental site is subtropical with moderately cold and dry winters and warm summers. Based on data of meteorological stations established at the sites, mean annual temperature was 17.4°C and mean annual precipitation was 1635 mm (Fig. 1). Across an area of 38 ha, 219,000 trees were planted at different levels of species richness [29]. The diversity gradient spans from monoculture to two, four, eight, 16 and up to 24 species per plot. The 39 tree species included in the study (see Table 1) are representative of the local natural broadleaved subtropical forest community [30],[31], and the trees assessed had already reached an age of four or five years at the time when our study was carried out. Using young even-aged trees in a common garden situation allowed for controlling for confounding factors, such as different ontological stage, but also allowed to sample leaves and branches at a standardized height above ground. No specific permissions were required for these locations and activities. The field studies did involve neither endangered nor protected species.
Figure 1. Climate diagram according to Walter & Lieth [28] of Xingangshan, the location of the experimental sites.
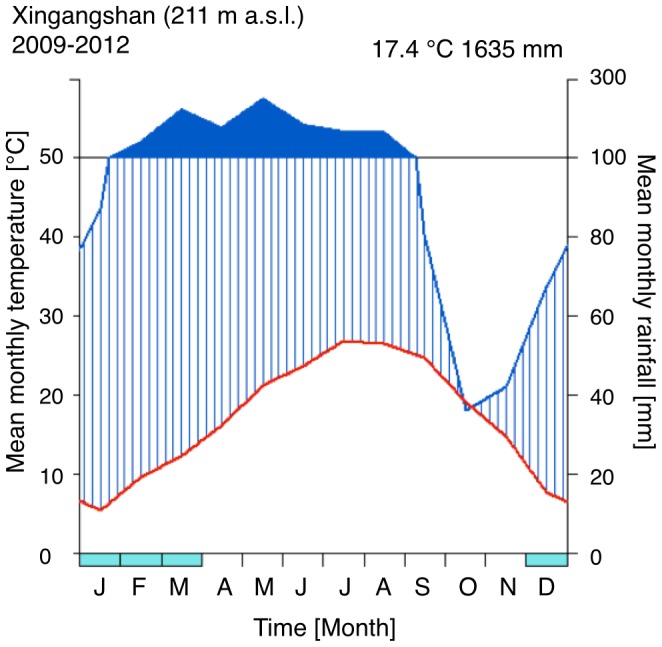
Elevation: 211 m above sea level. Observation period was March 2009 to October 2012. Mean annual temperature was 17.4°C and total annual precipitation was 1635 mm. Monthly precipitation below 100 mm is scaled 2∶1 with mean monthly temperature (vertically hatched) and above 100 mm 15∶1. Turquoise bars below the x-axis show the months where frosts can occur (when absolute monthly minimums are equal or lower than 0°C). Climate data were recorded by a meteorological station established at the very center of the experimental site (Kühn, unpublished).
Table 1. Tree species planted in the BEF-China experiment and included in this study.
| Species name | Family | Abbreviation | Leaf habit |
| Acer davidii Franch. | Aceraceae | Ad | d |
| Ailanthus altissima (Miller) Swingle | Simaroubaceae | Aa | d |
| Alniphyllum fortunei (Hemsl.) Makino | Styracaceae | Af | d |
| Betula luminifera Winkl. | Betulaceae | Bl | d |
| Castanopsis eyrei (Champion ex Bentham) Tutcher | Fagaceae | Ce | e |
| Castanopsis fargesii Franch. | Fagaceae | Cf | e |
| Castanea henryi (Skan) Rehd. et Wils. | Fagaceae | Ch | d |
| Castanopsis sclerophylla (Lindley & Paxton) Schottky | Fagaceae | Cs | e |
| Celtis biondii Pamp. | Cannabaceae | Cb | d |
| Choerospondias axillaris (Roxb.) Burtt et Hill | Anacardiaceae | Ca | d |
| Cinnamomum camphora (Linn.) Presl | Lauraceae | Cc | e |
| Cyclobalanopsis glauca (Thunberg) Oersted | Fagaceae | Cg | e |
| Cyclobalanopsis myrsinifolia (Blume) Oersted | Fagaceae | Cm | e |
| Daphniphyllum oldhamii (Hemsl.) Rosenthal | Daphniphyllaceae | Do | e |
| Diospyros japonica Siebold & Zuccarini | Ebenaceae | Dj | d |
| Elaeocarpus chinensis (Gardn. et Chanp.) Hook. f. ex Benth. | Elaeocarpaceae | Ec | e |
| Elaeocarpus glabripetalus Merr. | Elaeocarpaceae | Eg | e |
| Elaeocarpus japonicus Sieb. et Zucc. | Elaeocarpaceae | Ej | e |
| Idesia polycarpa Maxim. | Flacourtiaceae | Ip | d |
| Koelreuteria bipinnata Franch. | Sapindaceae | Kb | d |
| Liquidambar formosana Hance | Altingiaceae | Lf | d |
| Lithocarpus glaber (Thunb.) Nakai | Fagaceae | Lg | e |
| Machilus grijsii Hance | Lauraceae | Mg | e |
| Machilus leptophylla Hand.-Mazz. | Lauraceae | Ml | e |
| Machilus thunbergii Sieb. et Zucc. | Lauraceae | Mt | e |
| Manglietia fordiana (Oliver) HuY.W.Law | Magnoliaceae | Manf | e |
| Melia azedarach Linn. | Meliaceae | Ma | d |
| Meliosma flexuosa Blume | Sabiaceae | Mf | d |
| Nyssa sinensis Oliver | Nyssaceae | Ns | d |
| Phoebe bournei (Hemsl.) Yen C. Yang, | Lauraceae | Pb | e |
| Quercus acutissima Carruthers | Fagaceae | Qa | d |
| Quercus fabri Hance | Fagaceae | Qf | d |
| Quercus phillyreoides A. Gray | Fagaceae | Qp | e |
| Quercus serrata Murray | Fagaceae | Qs | d |
| Rhus chinensis Mill. | Anacardiaceae | Rc | d |
| Sapindus saponaria Linn. | Sapindaceae | Sd | d |
| Triadica cochinchinensis Loureiro | Euphorbiaceae | Tc | d |
| Triadica sebifera (L.) Small | Euphorbiaceae | Ts | d |
| Schima superba Gardn. et Champ. | Theaceae | Schs | e |
Species names are in accordance with nomenclature in The Flora of China (http://flora.huh.harvard.edu/china). d = deciduous, e = evergreen.
Tree Species and Vulnerability Curves
We randomly chose three individuals per species in the high-diversity plots, with one individual per species being sampled per plot. This enabled us to minimize time between sample cutting and lab procession of samples because different species grew in close proximity to each other. Xylem conductivity and vulnerability and leaf stomatal conductance (see below) were measured on the same plots, but not explicitly on the same individuals. The sampling and measurements on xylem hydraulics were conducted in August - October 2012. This period was characterized by monthly mean temperatures of about 20°C and a monthly precipitation of 40 mm (Fig. 1), which involved dry spells of several weeks, typically resulting in midday depressions of stomatal conductance. Samples were always taken in the early morning hours between 6 and 8 am, when relative humidity was still high (70–95% Rh) and temperatures were around 20°C. Measurements of leaf water potentials were made in spring 2012, using a PMS M1000 Scholander pressure chamber. These data showed that water potentials were well above −2 MPa, and for many species>−1 MPa. A twig with no leaves, buds or branches, around 15 cm in length and 5–15 mm diameter was cut and immediately immersed in water. We are aware that maximum vessel length of some of the species might be larger than 15 cm [32], which would result in overestimating specific xylem hydraulic conductivity (KS) and the absolute value of the xylem pressure at which 50% loss of the maximum specific xylem hydraulic conductivity occurs (Ψ50). However, it has been shown that extreme vessel lengths are very rare [33]. In any case, obtaining non-ramified twigs longer than 15 cm would have been impossible in most species. After transportation to the lab, the stem pieces were then placed into a double-ended pressure sleeve (PMS M1000 Scholander pressure chamber) in the laboratory following established protocols [18],[34] (Fig. 2). Xylem vulnerability was measured within at maximum four hours after cutting. Increasing the air pressure in the cavitation chamber was used to simulate increasingly negative xylem sap pressures [35]. Before the measurements were taken, each twig segment was treated for one hour with perfusion solution pressurized at 0.15 MPa in order to flush out air from older embolism events and any potential air entry into the xylem during the cutting and handling of samples. We used 10 mM citric acid perfusion solution, using filtered and demineralized water to prevent any blockages caused by microorganisms.
Figure 2. Simplified overview of the xylem hydraulics measurement installation.

A) Pressure chamber with the stem segment, B) tank with the perfusion solution, C) flush tank, D) three-way stopcock, E) electronic scale, F) nitrogen pressure cylinder, and G) pressure gauge.
Specific xylem hydraulic conductivity (KS) was measured as the mass of flow-through perfusion solution through the piece of wood per unit of time and per cross-sectional area of the twig. The measurements were started at ambient air pressure and repeated as pressure was increased in increments of 0.5 MPa up to 7 MPa, or to the pressure at which no more perfusion solution flow was encountered.
Measurements of Stomatal Conductance
Data on stomatal conductance were taken from [7]. For the measurements of stomatal conductance the authors had randomly selected twelve high-diversity plots (with 16 or 24 species). Eleven to 23 individuals of all species were measured per plot. In total, 3,290 measurements of stomatal conductance were made in August to October 2010, May/June 2011 and August to October 2011. Each of the 39 species was represented between at least three and at maximum 13 individuals, resulting in 218 individuals in total. This non-balanced sampling design resulted from measuring daily course of stomatal conductance, which required walking time to be minimal. The repeated measurements were taken on the same leaf, which was fully developed, undamaged and fully exposed to the sun. Daily courses of stomatal conductance were produced for all species in every plot. Stomatal conductance was recorded with an SC1 porometer (Decagon). Air temperature and relative humidity was measured simultaneously using a T2 thermo-hygrometer (Trotec). Vapor pressure deficit (VPD) was calculated following the August-Roche Magnus formula. The daily courses of all different individuals from all different daily courses were then aggregated to one gs ∼ VPD relationship which included all data for one species. Mean and maximum stomatal conductance (gsmax) could then be estimated per species. The species-specific gs ∼ VPD relationships were modeled by regressing the logits of gs/gsmax to VPD and the quadratic term of VPD using a generalized linear model with a binomial error distribution. The parameters of the model allowed calculating the maximum stomatal conductance and the VPD at which the modeled stomatal conductance was maximal.
Trait Measurements
A total of 34 leaf and wood traits were assessed to analyze possible relationships with Ψ50 and KS (see Table 2). To accomplish this, four total sets of samples were taken: 1) A set of five individuals with five leaves being sampled per individual for the traditional leaf traits, such as absolute area per leaf, leaf fresh-weight, leaf dry-weight, leaf nitrogen concentration (LNC) and leaf carbon concentration (LCC). The data were used to calculate specific leaf area (SLA), leaf dry matter content (LDMC) and carbon to nitrogen ratio (CN). We also determined leaf habit (deciduous/evergreen), leaf pinnation (pinnate or simple), leaf margin (entire or serrate) and recorded the presence or absence of extrafloral nectaries. 2) Another 30 leaves were sampled from three individuals per species to determine leaf tensile strength as a measure of leaf toughness. Leaf tensile strength was measured with a tearing apparatus modified after Hendry [36]. 3) The same leaves on which stomatal conductance was measured (see above), were taken to analyze the stomatal related traits. Stomatal traits were analyzed after Gerlach [37], with stomata being counted on a minimum area of 50,000 µm2 on three leaves from three individuals per species. Stomata were counted on nail polish impressions made on leaf samples, which had been stored in 70% ethanol. Length and width of three stomata per replicate were measured, and stomatal density was expressed as stomatal number per area. The analysis was performed with a light-optical microscope (Zeiss Axioskop 2 plus) and using the Axio Vision (Version 3.0) software. 4) A sample was taken from each twig used in the cavitation sensitivity analysis for further xylem anatomical investigation. Twig sections were prepared for light microscopic inspection and, from an area of 4.4 mm2 per sample, every xylem vessel was analyzed. To determine xylem traits, we made use of XylemDetector that was implemented as part of the open-source package MiToBo (http://www.informatik.uni-halle.de/mitobo), an extension of the Java image processing software ImageJ. We measured the mean lumen area of conducting vessels (MEANAREA) and the mean roundness of conducting vessels (MEANROUND), which is a measure of how close the vessel shape is to a perfect circle, and ranges from 0 to 1. MEANROUND was calculated as:
where a is the area and p the perimeter of the lumen.
Table 2. List of the leaf traits measured across the 39 tree species.
| Code | Trait | Analytical technique | Type | Units/categories |
| Ψ50 | Loss of 50% initial conductivity | Pressure Chamber | Continuous | MPa |
| KS | Maximum conductivity | Pressure Chamber | Continuous | kg m−1 s−1 MPa−1 |
| b | Parameter b (Sigmoid Regression) | Pressure Chamber | Continuous | nondimensional |
| CONMEAN | Average stomatal condutance | Porometer | Continuous | mmol m2 s−1 |
| CONMAX | Maximum stomatal condutance | Porometer | Continuous | mmol m2 s−1 |
| VPDMAX | VPD at CONMAX | Porometer, Hygrometer, Thermometer | Continuous | hPa |
| CONMAXFIT | Fitted Max. stomatal conductance | Porometer, Hygrometer, Thermometer | Continuous | nondimensional |
| VPDMAXFIT | VPD at CONMAXFIT | Porometer, Hygrometer, Thermometer | Continuous | hPa |
| VPDPOI | VPD at point of inflection of fitted stomatal conductance | Porometer, Hygrometer, Thermometer | Continuous | hPa |
| SLA | Specific leaf area | Scanner, Balance | Continuous | m2 kg−1 |
| LOG10LA | Decadic log (Leaf Area) | Scanner | Continuous | mm2 |
| LDMC | Leaf dry matter content | Balance | Continuous | mg g−1 |
| LT | Leaf toughness | Leaf toughness device | Continuous | N mm−1 |
| LEAFHABIT | Leaf habit | Literature | Binary | (0) evergreen; (1) deciduous |
| LNC | Leaf nitrogen concentration | CN Analyzer | Continuous | mg g−1 |
| LCC | Leaf carbon concentration | CN Analyzer | Continuous | mg g−1 |
| CN | Carbon-nitrogen ratio | CN Analyzer | Continuous | ratio |
| CA | Leaf calcium concentration | AAS Analyzer | Continuous | mg g−1 |
| K | Leaf potassium concentration | AAS Analyzer | Continuous | mg g−1 |
| MG | Leaf magnesium concentration | AAS Analyzer | Continuous | mg g−1 |
| LEAFPIN | Leaf pinnation | Field Observation | Binary | (0) pinnate; (1) simple |
| LEAFMAR | Leaf margin | Field Observation | Binary | (0) dentate; (1) entire |
| EXTRAFLO | Extrafloral nectaries | Field Observation | Binary | (0) no; (1) yes |
| STODENS | Stomata density | Microscopy | Continuous | 1 mm−2 |
| STOSIZE | Stomata size | Microscopy | Continuous | µm2 |
| STOIND | Stomata index | Microscopy | Continuous | ratio |
| DIAMVEIN1 | Diameter veins 1st order | Scanner | Continuous | cm |
| DIAMVEIN2 | Diameter veins 2nd order | Scanner | Continuous | cm |
| VEINDENS | Length of veins per unit leaf area | Scanner | Continuous | cm cm−2 |
| WPOT | Water potential | Pressure Chamber | Continuous | MPa |
| WOODDENS | Wood density | Balance | Continuous | g cm3 |
| MEANAREA | Mean area of conducting vessels | Microscopy | Continuous | µm2 |
| MEANROUND | Mean roundness of conducting vessels | Microscopy | Continuous | nondimensional |
| DHYD | Hydraulically weighted diameter of conducting vessels | Microscopy | Continuous | µm |
All traits were assessed on the individuals planted in the experiment. The table includes the trait abbreviations (Code) used throughout the text.
Following Sperry [38], we calculated hydraulically weighted conduit diameter (DHYD) from the lumen area data according to:
where r are the circle radii calculated from the lumen areas.
Statistical Analyses
We plotted vulnerability curves that show the flow rates of perfusion solution through stem segments as a function of water potential [35]. A sigmoid, three-parameter regression was applied to the vulnerability data [39],[40]:
where KS is the specific hydraulic conductivity of the xylem [kg m−1 s−1 MPa−1], Ψ is the xylem pressure at which water flow was measured [MPa], a is the original maximum specific xylem hydraulic conductivity, b is the slope of the regression and Ψ50 is the xylem pressure at which 50% loss of the original maximum specific xylem hydraulic conductivity occurs. Fig. 3 shows an example of how the sigmoid model was fitted to predict the loss of specific xylem hydraulic conductivity (KS) from water potential (please refer to the Figure S1 and Table S1 for raw data on xylem vulnerability). We made use of the FactoMineR package in R (http://cran.r-project.org/web/packages/FactoMineR/index.html) to correlate the physiological parameters to the species' traits in a PCA. KS, Ψ50 and the slope b of the KS ∼ Ψ relationship were tested for differences between deciduous and evergreen leaf habit by analysis of variance (ANOVA) and for bivariate relationships to leaf traits by linear regression models. As these tests performed multiple testing, they run the risk of error inflation and cannot be used to infer statistical significances. These tests were exploratory and had the purpose to identify possible candidate predictors and to show the direction of their effects. To further investigate the emerging significant relationships, we rerun all significant linear regressions by additionally including the interaction with leaf habit. For all statistics, R software version 3.0.2 was used.
Figure 3. Conductivity rates as a function of decreasing water potential, taking Castanea henryi as an example.
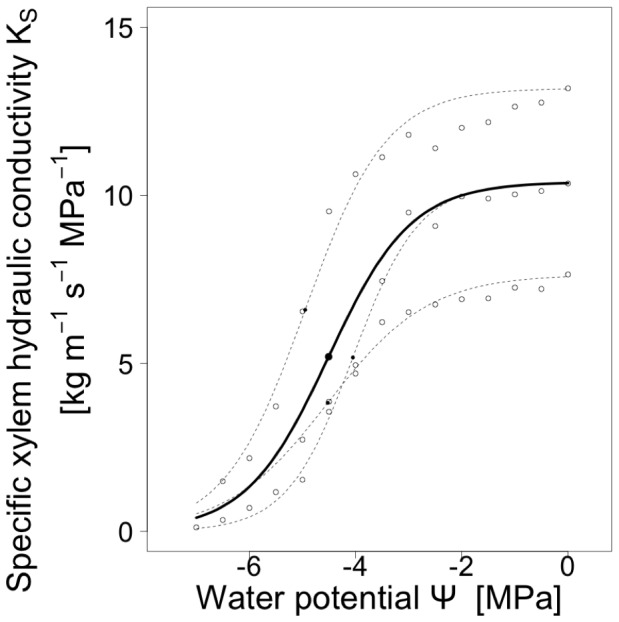
Outline dots show the measured values of three independent vulnerability curves. Dotted lines show the single regression functions obtained from the measured value per vulnerability curve, obtained from a 3-parametric sigmoid function. The bold line shows the regression lines constructed from the means of the parameters from the three single vulnerability curves. Filled dots represent Ψ50 values.
Results
Fig. 4 shows the vulnerability curves for all 39 species included in the study. Values of Ψ50 ranged between −1.08 MPa and −6.6 MPa for Celtis biondii and Lithocarpus glaber, respectively, with an overall mean of −3.78 MPa (SD = 1.48). Specific xylem hydraulic conductivity (KS) was highest in Melia azedarach, Triadica sebifera and Castanea henryi (17.52, 11.01 and 10.40 kg m−1 s−1 MPa−1, respectively) and lowest in Machilus grijsii (0.036 kg m−1 s−1 MPa−1), with a overall mean of 2.44 kg m−1 s−1 MPa−1 (SD = 3.31). KS and Ψ50 were not correlated across all species (p = 0.512). Evergreen species had significantly lower values of maximum hydraulic conductivity and lower Ψ50 values than deciduous species (Fig. 5).
Figure 4. Plots of modeled specific xylem hydraulic conductivity KS versus water potentials for all 39 species included in the study.

A) and B) show the same data at different scale of KS. Deciduous species are shown in red, evergreen species shown in black. For species abbreviations see Table 1. For details of calculation of regression lines, see Fig. 2 and Methods.
Figure 5. Boxplots characterizing the species set by leaf habit.

A) xylem pressure at 50% loss of the maximum specific xylem hydraulic conductivity (Ψ50) and B) specific xylem hydraulic conductivity of the xylem (KS) as a function of leaf habit. d = deciduous, e = evergreen. The probability values were derived from an ANOVA.
Fig. 6 shows the principal components analysis (PCA) of all 34 leaf traits, including the parameters of stomatal control and xylem vulnerability for all 39 study species (Fig. 6 a, b). The species mean values of all traits are provided in Table S1. The first three PCA axes explained 43.3% of the total variance, with eigenvalues of 7.03, 4.2 and 3.48, respectively. While evergreen species tended to score higher on the first PCA axes than deciduous species, there was a large overlap between the two leaf habits (Fig.6 c, d). Positive scores on the first PCA axis reflected both decreasing xylem vulnerability and increasing values of traits of the leaf economics spectrum, such as leaf nitrogen concentration (LNC) and specific leaf area (SLA), as well as evergreen leaf habit and the logarithm of the area of a single leaf (Log10LA), while leaf toughness (LEAFT) and leaf carbon to nitrogen ratio (CN) showed negative loadings. Parameters of stomatal control were correlated with the second PCA axis, with positive loadings being recorded for stomatal index (STOIND), stomatal density (STOMDENS) and wood density (WOODDENS), and negative ones for the point of inflection of the gs ∼ VPD curve (VPDPOI) and maximum stomatal conductance (CONMAX).
Figure 6. Principal component analysis (PCA) biplots showing the relationships between the mean values of all chemical and morphological leaf traits (black arrows), all parameters of stomatal regulation (green arrows) and all parameters of cavitation sensitivity (blue arrows).

A) and C) PCA axes 1 and 2, B) and D) PCA axes 1 and 3. A) and B) loadings of the different traits. C) and D) species scores in the PCA shown separately by leaf habit. Deciduous species are shown in red and evergreen species in black. See Table 1 for abbreviations of species names. Eigenvalues: axis 1 = 7.03, axis 2 = 4.2, axis 3 = 3.48, with cumulative proportion of explained inertia 20.1%, 33.0% and 43.3%, respectively. See Table 2 for abbreviations of trait names.
We found xylem vulnerability to be significantly related to numerous traits (Table 3, Fig. 7 & 8). Ψ50 was positively correlated with leaf area, LNC, hydraulically weighted diameter and leaf magnesium concentration, while it was only marginally significantly related to leaf calcium concentration, and negatively related to leaf toughness and carbon to nitrogen ratio. A similar pattern was found for maximum hydraulic conductivity (KS), which showed a positive relationship to leaf area (Log10LA), leaf nitrogen concentration (LNC) and two morphological wood traits (i.e. the mean area of conducting vessels (MEANAREA), and the hydraulically weighted diameter) and a negative correlation with leaf carbon to nitrogen ratio (CN). The regression parameter b was not related to any of the traits studied. The regression equations from the significant linear models are shown in Table 4. There were no significant correlations between Ψ50 or KS to any parameter of stomatal control.
Table 3. Correlation matrix of hydraulic xylem characteristics with numerical leaf traits and parameters of stomatal control.
| Ψ50 | KS | b | ||||
| r | p | r | p | r | p | |
| CONMEAN | 0.12 | 0.49 | 0.13 | 0.42 | −0.12 | 0.49 |
| CONMAX | 0.02 | 0.92 | 0.02 | 0.91 | −0.06 | 0.73 |
| VPDMAX | −0.04 | 0.81 | 0.15 | 0.36 | 0.14 | 0.39 |
| CONMAXFIT | 0.14 | 0.41 | 0.26 | 0.12 | −0.11 | 0.50 |
| VPDMAXFIT | −0.13 | 0.43 | 0.18 | 0.26 | −0.01 | 0.96 |
| VPDPOI | 0.13 | 0.44 | 0.13 | 0.44 | −0.10 | 0.53 |
| SLA | 0.27 | 0.09 | 0.05 | 0.78 | −0.17 | 0.30 |
| LOG10LA | 0.34 | 0.03 | 0.10 | 0.04 | −0.20 | 0.22 |
| LDMC | −0.02 | 0.91 | −0.02 | 0.91 | −0.03 | 0.85 |
| LEAFT | −0.47 | 0.00 | −0.23 | 0.16 | 0.02 | 0.90 |
| LNC | 0.43 | 0.01 | 0.59 | 0.00 | −0.08 | 0.62 |
| LCC | −0.23 | 0.17 | −0.12 | 0.49 | 0.05 | 0.76 |
| CN | −0.43 | 0.01 | −0.48 | 0.00 | 0.05 | 0.75 |
| CA | 0.32 | 0.05 | 0.09 | 0.58 | −0.16 | 0.35 |
| K | 0.16 | 0.33 | 0.05 | 0.75 | −0.13 | 0.44 |
| MG | 0.33 | 0.04 | 0.02 | 0.91 | −0.24 | 0.15 |
| STOMDENS | 0.09 | 0.58 | 0.07 | 0.65 | −0.08 | 0.64 |
| STOMSIZE | 0.06 | 0.74 | −0.06 | 0.71 | −0.01 | 0.96 |
| STOIND | 0.11 | 0.52 | 0.01 | 0.69 | −0.09 | 0.59 |
| DIAMVEIN1 | 0.20 | 0.23 | −0.09 | 0.59 | −0.23 | 0.17 |
| DIAMVEIN2 | 0.17 | 0.33 | 0.01 | 0.93 | −0.14 | 0.42 |
| VEINLENGTH | −0.15 | 0.37 | −0.09 | 0.61 | 0.19 | 0.24 |
| WPOT | 0.09 | 0.57 | −0.04 | 0.79 | 0.01 | 0.95 |
| WOODDENS | −0.06 | 0.73 | 0.06 | 0.71 | −0.13 | 0.42 |
| MEANAREA | 0.24 | 0.15 | 0.85 | 0.00 | −0.11 | 0.52 |
| MEANROUND | 0.04 | 0.82 | −0.01 | 0.94 | −0.02 | 0.92 |
| DHYD | 0.39 | 0.01 | 0.72 | 0.00 | −0.20 | 0.23 |
The correlations were calculated from linear model analyses between functional leaf traits and the extracted physiological parameters. Significant relationships are shown in bold letters. Ψ50 = xylem pressure at which 50% loss of the original maximum specific xylem hydraulic conductivity occurred, KS = specific hydraulic conductivity of the xylem, b = slope of the three-parametric sigmoid model of the vulnerability curve, CONMEAN = mean gs; CONMAX = mean gs max; VPDMax = VPD at gs max; CONMAXFIT = modeled gs max; VPDMaxfit = VPD at modeled gs max, and VPDPoi = VPD at the point of inflexion of the gs ∼ VPD curve, MEANAREA = the mean area of conducting vessels, MEANROUND = the mean roundness of conducting vessels, and DHYD the hydraulically weighted diameter of conducting vessels.
Figure 7. Ψ50 as a function of A) leaf area (p = 0.0315, r = 0.34), B) leaf toughness (p<0.0003, r = −0.47), C) leaf nitrogen concentration (p<0.0075, r = 0.43), D) leaf carbon to nitrogen ratio (p<0.0078, r = −0.43), E) leaf magnesium concentration (p<0.042, r = 0.33) and F) hydraulically weighted conduit diameter (p<0.01, r = 0.39).

Filled black dots represent species of evergreen leaf habit; empty dots represent species of deciduous leaf habit.
Figure 8. KS as a function of A) leaf area (p<0.044, r = 0.1), B) leaf nitrogen concentration (p<0.00001, r = 0.59), C) leaf carbon to nitrogen ratio (p<0.0019, r = 0.48), D) mean area of conducting vessels (p<0.0001, r = 0.85), and E) hydraulically weighted diameter of conducting vessels (p<0.0001, r = 0.72).

Filled black dots represent species of evergreen leaf habit and empty dots represent species of deciduous leaf habit. In B) and C) KS also showed significant interactions with leaf habits. Dotted lines represent species of evergreen and broken lines represent species of deciduous leaf habit. B) leaf habit p = 0.061, interaction leaf nitrogen concentration and leaf habit p = 0.049, C) leaf habit p = 0.026, interaction leaf carbon to nitrogen ratio and leaf habit p = 0.031.
Table 4. Regression equations for all significant correlations of Ψ50 and KS to the functional traits.
| Xylem parameter | Functional trait | Equation |
| Ψ50 | LOG10LA | y = 1.0090x−7.3717 |
| Ψ50 | LEAFT | y = −3.2808x−2.3454 |
| Ψ50 | LNC | y = 1.9556x−7.1038 |
| Ψ50 | CN | y = −0.12210x−0.25799 |
| Ψ50 | MG | y = 0.2763x−5.0792 |
| Ψ50 | DHYD | y = 0.0136x−5.005 |
| KS | LOG10LA | y = 2.0699x−4.8375 |
| KS | LNC | y = 6.003x−7.734 |
| KS | CN | y = −0.31132x+11.44974 |
| KS | MEANAREA | y = 0.0011481x−0.7089940 |
| KS | DHYD | y = 0.0569x−2.611 |
The correlations were calculated from linear model analyses. Ψ50 = xylem pressure at which 50% loss of the original maximum specific xylem hydraulic conductivity occurred, KS = specific hydraulic conductivity of the xylem, b = slope of the three-parametric sigmoid model of the vulnerability curve, CONMEAN = mean gs; CONMAX = mean gs max; VPDMax = VPD at gs max; CONMAXFIT = modeled gs max; VPDMaxfit = VPD at modeled gs max, and VPDPoi = VPD at the point of inflexion of the gs ∼ VPD curve, MEANAREA = the mean area of conducting vessels, and DHYD the hydraulically weighted diameter of conducting vessels.
Including leaf habit in the significant models resulted in two models with significant interactions with leaf habit, i.e. for leaf nitrogen concentration (LNC) and the leaf carbon to nitrogen ratio (CN). While with increasing CN and decreasing LNC, KS increased in evergreen species, deciduous species showed a decrease (Fig. 8 a, b).
Discussion
We found a large variation in specific xylem hydraulic conductivity and xylem vulnerability among our study species, which points to different complementary strategies of the species in the same subtropical forest community. Thus, our results conform to the findings of Böhnke et al. [41], who revealed a high and temporally constant level of functional diversity in the course of succession in these forests. In particular, variation in cavitation resistance could offer an explanation for species coexistence in the same community [42]. Our values on specific xylem hydraulic conductivity with a range of KS between 0.036 and 17.52 kg m−1 s−1 MPa−1 and xylem vulnerability to cavitation Ψ50 between −1.08 and −6.6 MPa cover a large part of the total range recorded for such variables in other studies [43]. For example, Cavender-Bares et al. [44] described a range of maximum specific xylem hydraulic conductivity of between 1.75 and 5 kg m−1 s−1 MPa−1 for 17 oak species, while Maherali et al. [45] reported a mean of 1.36 kg m−1 s−1 MPa−1 of maximum specific xylem hydraulic conductivity (n species = 87) and a mean of −3.15 MPa for Ψ50 values (n species = 167). Slightly higher Ψ50 values of −1.2 up to −2.76 MPa were encountered for eight tree species from a tropical dry forest [46]. However, we have to consider that extreme values of KS of more than 10 kg m−1 s−1 MPa−1 might be methodological artifacts, caused by some open vessels in these samples. However, the three species with extreme KS also ranked high in values of the predictor traits for KS, indicating that the relative rank in KS in these species might be realistic. As vessel length of the species was not measured, and thus, in some species, specific xylem hydraulic conductivity might have been overestimated, comparisons to other studies should be done with caution. However, as Melcher et al. [33] pointed out, such an overestimation is probably not severe, as long vessels are also very rare.
Important leaf traits such as leaf nitrogen concentration (LNC) and leaf area (Log10LA) were highly correlated to Ψ50 and KS. Thus, we can fully confirm our first hypothesis that leaf traits describing the leaf economics spectrum are related to specific xylem hydraulic conductivity and cavitation resistance. Interestingly, there were no significant correlations for some of the traits of the leaf economics spectrum, such as SLA and LDMC. Thus, leaf thickness and water content of leaves seem to have less importance for hydraulic characteristics than the leaves' absolute size and protein content. This contrasts with the findings reported by Willson et al. [47], who described a significant relationship between SLA and Ψ50 for the genus Juniperus. Thus, comparative studies confined to certain taxonomic levels, such as congeneric comparisons, might arrive at different conclusions than studies covering a wider range of taxa. Alternatively, SLA may have a differing level of importance with regard to the physiology of gymnosperms and angiosperms. Interestingly, our study did also not support a link between KS and leaf vein density, which was predicted by the flux trait network suggested by Sack et al. [15]. However, there was only equivocal evidence for a significant relationship between KS and leaf vein density in their reviewed studies [15].
Our results of a strong relationship of Ψ50 and KS to leaf area, conform to those encountered for eight southern African tree species of a seasonally dry tropical forest by Vinya et al. [48], except that they reported a link to leaf area only for KS, but not for Ψ50. Our findings of a relationship of Ψ50 to Mg concentration and a marginal one to Ca concentration might indicate that the non-vein and non-sclerenchyma mesophyll density in the leaf are more relevant for cavitation resistance than overall leaf tissue density. As a central component of chlorophyll, Mg concentration is directly related to photosynthetic capacity, and thus, might capture this proportion of actively assimilating tissue in the leaf. In addition, as a cofactor of many enzymatic processes, Mg can be considered an indicator for the plant's nutrition status [49].
The absence of any direct relationship between SLA or LDMC with Ψ50 or KS which is in accordance with Sack et al. [50], in combination with the large overlap in Ψ50 and KS detected between deciduous and evergreen species, is not a conclusive result. It appears that evergreen and deciduous subtropical forest species form two ends of a gradient from cavitation resistance to cavitation avoidance, respectively. This view is supported by a recent study by Fu et al. [17], who investigated the relationship of stem hydraulics and leaf phenology in Asian tropical dry forest species. In particular, they found a negative relationship between leaf life span and KS but no significant relationship between leaf life span and Ψ50. In accordance with our results, Maherali et al., Choat et al. and Chen et al. [27],[45],[51] also reported significant differences in Ψ50 and KS between the different leaf habit groups. Such differences in hydraulic characteristics also translate to higher growth rates, as shown by Fan et al. [52] for 40 Asian tropical trees. However, some other studies failed to detect any differences, such as that of Markesteijn et al. [42], who attributed the substantial differences they encountered in Ψ50 and KS to shade tolerance. They also argued that the distinction between pioneer vs. shade-tolerant species predicts hydraulic properties better than leaf habit, because there are considerable overlaps in strategies along the gradient of leaf longevity. As Givnish [53] pointed out, evergreen leaves can be advantageous under a wide range of ecological conditions, and the relationship of leaf habit with Ψ50 and KS may therefore strongly depend on the system considered. We can also confirm our second hypothesis that evergreen species characterized by low SLA and high LDMC show lower Ψ50 and KS values than deciduous species. Interestingly, we found leaf habit to significantly influence the relationships of KS to leaf nitrogen concentration and carbon to nitrogen ratio, which might be explained by differences in basic leaf constructing principles. Deciduous species tend to invest high amounts of nitrogen to maximize photosynthetic assimilation per leaf mass, whereas in evergreen species, the focus is on increased leaf lifespan, which is reflected in higher leaf carbon concentration [54]. Furthermore, deciduous species show a more conservative stomatal control to avoid embolism, whereas evergreen species tend to have more cavitation-resistant vessels [7,25] [7],[25]–[27],[55]. The underlying reason is that evergreen species are mostly diffuse-porous, which also explains the strong impact of the hydraulically weighted conduit diameter and mean area of conducting vessels on KS, which is also well-known from the literature [19].
Contrary to expectations, this study did not find a significant link between xylem hydraulic conductivity and parameters of stomatal regulation. Neither the maximum stomatal conductance, the vapor pressure deficit at maximum stomatal conductance nor the vapor pressure deficit at which stomatal conductance is down-regulated was related to any parameter of the xylem vulnerability curves. As such, our third hypothesis has to be rejected, which implies that the ability of a very precise and fast stomatal regulation versus a retarded and inert stomatal regulation does not translate into cavitation resistance. Additional insights into the relationship between KS and leaf stomatal regulation might be gained by calculating leaf-specific xylem hydraulic conductivity KL, which would directly refer to the capacity of the vascular system of a stem to supply the water to that stem [56]. However, our results confirm those of Brodribb et al. [46], who found no correlation between Ψ50 and the leaf water potential at stomatal closure in eight tropical, dry forest trees. They concluded that xylem cavitation and stomatal closure are linked through complex indirect regulatory mechanisms and argue that this potential linkage is considerably flexible, especially with regard to different leaf phenology strategies, and that there may be carry-over effects of preceding embolism events on stomatal control. A further explanation of a lacking relationship between xylem vulnerability and stomatal regulation may be the different scale at which stomatal regulation is considered. At the level of whole trees, Litvak et al. [55] found a strong linear relationship between the sensitivity of tree-level sap flow to VPD and Ψ50 both within diffuse- and ring-porous species, which was not encountered for leaf-level transpiration rates. The authors argue that the tree-level transpiration sensitivity, in addition to stomatal regulation, also directly responds to drought-induced embolisms.
Several studies showed a trade-off between high hydraulic conductivity and cavitation resistance [57]–[60]. In contrast to these studies, we found Ψ50 and KS to be unrelated. According to our current understanding of the causes of xylem embolism under drought conditions, there may indeed be no mechanistic link between these two hydraulic characteristics. As such, diameter and length of vessels may differ autonomously from pit structure and size [19]. While KS is mainly driven by vessel diameter, Ψ50 depends on pit size and structure [19],[20]. The pit area hypothesis states that cavitation resistance is linked to the total area of inter-vessel pits per vessel [61],[62]. Thus, the risk of an embolism expanding between vessels rises with the maximum size of the pit membrane pore, which in turn is dependent on the associated pit membrane area per vessel. This was demonstrated by Hacke et al. [57], who reported a strong negative link between xylem vulnerability and pit membrane area per vessel, resulting in small pits potentially increasing hydraulic resistance and decreasing KS. However, pit size may be of minor importance to KS compared to that of vessel diameter, and the relationship of hydraulic conductivity and cavitation resistance might depend on the specific ecosystem considered. Tyree et al. [59] distinguished between frost- and drought-induced cavitation. In their meta-analysis, the trade-off between hydraulic conductivity and cavitation resistance was mainly related to frost-induced cavitation events. Although frosts occur in the Chinese subtropics, they are neither very strong, nor long-lasting [63]. Thus, cavitation in the forests of our study area will mainly be brought about by drought events, which may result in far fewer, or insignificant, trade-offs.
Since all our individuals have the same age, our species set provides a high comparability usually not found in comparative studies. We expect that some of our response variables will change with tree age, such as specific xylem hydraulic conductivity [64],[65]. In addition, future comparisons should take the sustained leaf area into account, as whole-tree leaf-specific hydraulic conductance (KL) is known to decrease with tree age [66],[67].
Conclusion
For the studied subtropical forest community, we demonstrated a clear link of KS and Ψ50 with functional traits, and particularly with leaf nitrogen concentration, log leaf area and leaf carbon to nitrogen ratio. Thus, easily measured leaf traits from the LES have the potential to predict plant species' drought resistance. However, current knowledge on xylem vulnerability and traits from other ecosystems do not allow generalizing from these results. In addition, our finding of an absence of any relationship between parameters of stomatal control raises the question whether stomatal control as characteristics that are an independent axis of the LES might be related to an axis of xylem characteristics that are independent of specific xylem hydraulic conductivity and xylem vulnerability.
Supporting Information
Raw data for the vulnerability curves of the 39 study species analyzed. Filled dots represent measured data, empty dots show estimated Ψ50 values and the broken lines represent the fitted models of xylem vulnerability. For species abbreviations see Table 1.
(PNG)
{kind=link}
Trait raw data for the 39 study species analyzed. For trait codes and full species names, please see Tables 1 and 2.
(TXT)
Acknowledgments
We are very grateful for the help of Xuefei Yang, Sabine Both, Lin Chen and Xiaojuan Liu in coordinating the fieldwork for the BEF-China experiment. We also thank the whole BEF-China research group for their general support. In addition, we would like to thank Ricardo Schöps, Ricarda Pohl and Michael Staab for their support in the lab. Special thanks are due to Tim Langhammer and Stefan Posch for providing data on the xylem structure and to Peter Kühn for providing climate data.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files. Requests can also be directed to Wenzel Kröber, wenzel.kroeber@botanik.uni-halle.de.
Funding Statement
The authors gratefully acknowledge funding by the German Research Foundation (DFG FOR 891/1 and 891/2) with a grant to H.B. (DFG BR 1698/9-2) and the Sino-German Centre for Research Promotion in Beijing (GZ 524, 592, 698, 699, 785, 970 and 1020) as well as a grant to S.Z. of the National Natural Science Foundation of China (No. 31170457). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, et al. (2004) The worldwide leaf economics spectrum. Nature 428:821–827. [DOI] [PubMed] [Google Scholar]
- 2. Reich PB, Ellsworth DS, Walters MB, Vose JM, Gresham C, et al. (1999) Generality of leaf trait relationships: A test across six biomes. Ecology 80:1955–1969. [Google Scholar]
- 3. Shipley B, Lechowicz MJ, Wright I, Reich PB (2006) Fundamental trade-offs generating the worldwide leaf economics spectrum. Ecology 87:535–541. [DOI] [PubMed] [Google Scholar]
- 4. Cornelissen JHC, Wright IJ, Reich PB, Falster DS, Garnier E, et al. (2005) Assessing the generality of global leaf trait relationships. New Phytol 166:485–496. [DOI] [PubMed] [Google Scholar]
- 5. Santiago LS (2007) Extending the leaf economics spectrum to decomposition: evidence from a tropical forest. Ecology 88:1126–1131. [DOI] [PubMed] [Google Scholar]
- 6. Osnas JLD, Lichstein JW, Reich PB, Pacala SW (2013) Global leaf trait relationships: mass, area, and the leaf economics spectrum. Science 340:741–744. [DOI] [PubMed] [Google Scholar]
- 7. Kröber W, Bruelheide H (2014) Transpiration and stomatal control: A cross-species study of leaf traits in 39 evergreen and deciduous broadleaved subtropical tree species. Trees 28:901–914. [Google Scholar]
- 8. Baraloto C, Paine C, Poorter L, Beauchene J, Bonal D, et al. (2010) Decoupled leaf and stem economics in rain forest trees. Ecol Letters 13:1338–1347. [DOI] [PubMed] [Google Scholar]
- 9. Freschet GT, Cornelissen JHC, van Logtestijn RSP, Aerts R (2010) Evidence of the “plant economics spectrum” in a subarctic flora. J Ecol 98:362–373. [Google Scholar]
- 10.Lambers H, Chapin F III, Pons T (2008) Plant Physiological Ecology. 2nd ed. Berlin: Springer. 623 pp.
- 11. Sperry J, Choat B, Jansen S, Brodribb TJ, Cochard H, et al. (2012) Global convergence in the vulnerability of forests to drought. Nature 491:752–755. [DOI] [PubMed] [Google Scholar]
- 12. Poorter L, Mcdonald I, Alarcón A, Fichtler E, Licona J-C, et al. (2010) The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytol 185:481–492. [DOI] [PubMed] [Google Scholar]
- 13. Sack L, Frole K (2006) Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology 87:483–491. [DOI] [PubMed] [Google Scholar]
- 14. Brodribb TJ, Feild TS, Jordan GJ (2007) Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiol 144:1890–1898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Sack L, Scoffoni C, John GP, Poorter H, Mason CM, et al. (2013) How do leaf veins influence the worldwide leaf economic spectrum? Review and synthesis. J Exp Bot 64:4053–4080. [DOI] [PubMed] [Google Scholar]
- 16. Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, et al. (2009) Towards a worldwide wood economics spectrum. Ecol Letters 12:351–366. [DOI] [PubMed] [Google Scholar]
- 17. Fu P-L, Jiang Y-J, Wang A-Y, Brodribb TJ, Zhang J-L, et al. (2012) Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest. Ann Bot-London 110:189–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Sperry J, Donelly J, Tyree MT (1988) A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ 11:35–40. [Google Scholar]
- 19. Hacke U, Sperry J (2000) Functional and ecological xylem anatomy. PPEES 4:97–115. [Google Scholar]
- 20.Tyree MT, Zimmermann MH (2002) Xylem Structure and the Ascent of Sap. 2nd ed. Berlin: Springer. 302 pp.
- 21. Kursar TA, Engelbrecht BM, Burke A, Tyree MT, EI Omari B, et al. (2009) Tolerance to low leaf water status of tropical tree seedlings is related to drought performance and distribution. Funct Ecol 23:93–102. [Google Scholar]
- 22. Engelbrecht BM, Kursar TA (2003) Comparative drought-resistance of seedlings of 28 species of co-occurring tropical woody plants. Oecologia 136:383–393. [DOI] [PubMed] [Google Scholar]
- 23. Fonseca CR, Overton JM, Collins B, Westoby M (2000) Shifts in trait-combinations along rainfall and phosphorus gradients. J Ecol 88:964–977. [Google Scholar]
- 24. Skarpe C (1996) Plant functional types and climate in a southern African savanna. J Veg Sci 7:397–404. [Google Scholar]
- 25. Brodribb TJ, Jordan GJ (2008) Internal coordination between hydraulics and stomatal control in leaves. Plant Cell Environ 31:1557–1564. [DOI] [PubMed] [Google Scholar]
- 26. Cochard H, Bréda N, Granier A (1996) Whole tree hydraulic conductance and water loss regulation in Quercus during drought: evidence for stomatal control of embolism? Ann For Sci 53:197–206. [Google Scholar]
- 27. Choat B, Ball MC, Luly JG, Holtum JAM (2005) Hydraulic architecture of deciduous and evergreen dry rainforest tree species from north-eastern Australia. Trees 19:305–311. [Google Scholar]
- 28.Walter H, Lieth H (1967) Klima-Diagramm Weltatlas. Jena: Gustav Fischer Verlag. 250 pp.
- 29. Bruelheide H, Nadrowski K, Assmann T, Bauhus J, Both S, et al. (2014) Designing forest biodiversity experiments: general considerations illustrated by a new large experiment in subtropical China. Methods Ecol Evol 5:74–89. [Google Scholar]
- 30. Yang X, Bauhus J, Both S, Fang T, Härdtle W, et al. (2013) Establishment success in a forest biodiversity and ecosystem functioning experiment in subtropical China (BEF-China). Eur J Forest Res 132:593–606. [Google Scholar]
- 31. Bruelheide H, Böhnke M, Both S, Fang T, Assmann T, et al. (2011) Community assembly during secondary forest succession in a Chinese subtropical forest. Ecol Monogr 81:25–41. [Google Scholar]
- 32. Ennajeh M, Simões F, Khemira H, Cochard H (2011) How reliable is the double-ended pressure sleeve technique for assessing xylem vulnerability to cavitation in woody angiosperms? Physiologia Plant 142:205–210. [DOI] [PubMed] [Google Scholar]
- 33. Melcher PJ, Michele Holbrook N, Burns MJ, Zwieniecki MA, Cobb AR, et al. (2012) Measurements of stem xylem hydraulic conductivity in the laboratory and field. Methods Ecol Evol 3:685–694. [Google Scholar]
- 34. Perez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, et al. (2013) New handbook for standardised measurement of plant functional traits worldwide. Aust J Bot 61:167–234. [Google Scholar]
- 35. Cochard H, Badel E, Herbette S, Delzon S, Choat B, et al. (2013) Methods for measuring plant vulnerability to cavitation: a critical review. J Exp Bot 64:4779–4791. [DOI] [PubMed] [Google Scholar]
- 36.Hendry G, Grime J (1993) Methods in comparative Plant Ecology: a laboratory Manual. London: Chapman & Hall. 252 pp.
- 37.Gerlach D (1984) Botanische Mikrotechnik. Stuttgart, New York: Thieme. 289 pp.
- 38. Sperry J, Nichols KL, Sullivan JEM, Eastlack SE (1994) Xylem embolism in ring-porous, diffuse-orous, and coniferous trees of Northern Utah and interior Alaska. Ecology 75:1736. [Google Scholar]
- 39. Vander Willigen C, Sherwin HW, Pammenter NW (2000) Xylem hydraulic characteristics of subtropical trees from contrasting habitats grown under identical environmental conditions. New Phytol 145:51–59. [Google Scholar]
- 40. Domec J, Gartner BL (2001) Cavitation and water storage capacity in bole xylem segments of mature and young Douglas-fir trees. Trees 15:204–214. [Google Scholar]
- 41. Böhnke M, Kröber W, Welk E, Wirth C, Bruelheide H (2013) Maintenance of constant functional diversity during secondary succession of a subtropical forest in China. J Veg Sci 25:897–911. [Google Scholar]
- 42. Markesteijn L, Poorter L, Paz H, Sack L, Bongers F (2011) Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits. Plant Cell Environ 34:137–148. [DOI] [PubMed] [Google Scholar]
- 43. Sperry J, Tyree MT (1989) Vulnerability of xylem to cavitation and embolism. Annu Rev Plant Biol 40:19–36. [Google Scholar]
- 44. Cavender-Bares J, Kitajima K, Bazzaz FA (2004) Multiple trait associations in relation to habitat differentiation among 17 Floridian oak species. Ecol Monogr 74:635–662. [Google Scholar]
- 45. Maherali H, Pockman WT, Jackson RB (2004) Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology 85:2184–2199. [Google Scholar]
- 46. Brodribb TJ, Holbrook NM, Edwards E, Gutiérrez MRVA (2003) Relations between stomatal closure, leaf turgor and xylem vulnerability in eight tropical dry forest trees. Plant Cell Environ 26:443–450. [Google Scholar]
- 47. Willson CJ, Manos PS, Jackson RB (2008) Hydraulic traits are influenced by phylogenetic history in the drought-resistant, invasive genus Juniperus (Cupressaceae). Am J Bot 95:299–314. [DOI] [PubMed] [Google Scholar]
- 48. Vinya R, Malhi Y, Brown N, Fisher JB (2012) Functional coordination between branch hydraulic properties and leaf functional traits in miombo woodlands: implications for water stress management and species habitat preference. Acta Physiol Plant 34:1701–1710. [Google Scholar]
- 49.Bell P (2000) Green Plants: their Origin and Diversity. Cambridge: Cambridge Univ Pr. 349 pp.
- 50. Sack L, Cowan PD, Jaikumar N, Holbrook NM (2003) The “hydrology” of leaves: co-ordination of structure and function in temperate woody species. Plant Cell Environ 26:1343–1356. [Google Scholar]
- 51. Chen J-W, Zhang Q, Cao K-F (2009) Inter-species variation of photosynthetic and xylem hydraulic traits in the deciduous and evergreen Euphorbiaceae tree species from a seasonally tropical forest in south-western China. Ecol Res 24:65–73. [Google Scholar]
- 52. Fan ZX, Zhang S-B, Hao GY, Ferry Slik JW, Cao K-F (2012) Hydraulic conductivity traits predict growth rates and adult stature of 40 Asian tropical tree species better than wood density. J Ecol 100:732–741. [Google Scholar]
- 53. Givnish T (2002) Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox. Silva Fenn 36:703–743. [Google Scholar]
- 54. Aerts R (1995) The advantages of being evergreen. Trends Ecol Evol 10:402–407. [DOI] [PubMed] [Google Scholar]
- 55. Litvak E, McCarthy HR, Pataki DE (2012) Transpiration sensitivity of urban trees in a semi-arid climate is constrained by xylem vulnerability to cavitation. Tree Physiol 32:373–388. [DOI] [PubMed] [Google Scholar]
- 56. Choat B, Sack L, Holbrook NM (2007) Diversity of hydraulic traits in nine Cordia species growing in tropical forests with contrasting precipitation. New Phytol 175:686–698. [DOI] [PubMed] [Google Scholar]
- 57. Hacke UG, Sperry J, Wheeler JK, Castro L (2006) Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol 26:689–701. [DOI] [PubMed] [Google Scholar]
- 58. Markesteijn L, Poorter L, Bongers F, Paz H, Sack L (2011) Hydraulics and life history of tropical dry forest tree species: coordination of species' drought and shade tolerance. New Phytol 191:480–495. [DOI] [PubMed] [Google Scholar]
- 59. Tyree MT, Davis SD, Cochard H (1994) Biophysical perspectives of xylem evolution: is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA J 15:335–360. [Google Scholar]
- 60. Martínez-Vilalta J, Prat E, Oliveras I, Piñol J (2002) Xylem hydraulic properties of roots and stems of nine Mediterranean woody species. Oecologia 133:19–29. [DOI] [PubMed] [Google Scholar]
- 61. Jarbeau JA, Ewers FW, Davis SD (1995) The mechanism of water-stress-induced embolism in two species of chaparral shrubs. Plant Cell Environ 18:189–196. [Google Scholar]
- 62. Hargrave KR, Kolb KJ, Ewers FW, Davis SD (1994) Conduit diameter and drought-induced embolism in Salvia mellifera Greene (Labiatae). New Phytol 126:695–705. [Google Scholar]
- 63.Box E, Peet R, Masuzawa T, Yamada I, Fujiwara K, et al., editors (1995) Vegetation Science in Forestry. Dordrecht: Kluwer Academic Publishers. 664 pp.
- 64. Domec JC, Gartner BL (2003) Relationship between growth rates and xylem hydraulic characteristics in young, mature and old-growth ponderosa pine trees. Plant Cell Environ 26:471–483. [Google Scholar]
- 65. Rosner S, Klein A, Muller U, Karlsson B (2008) Tradeoffs between hydraulic and mechanical stress responses of mature Norway spruce trunk wood. Tree Physiol 28:1179–1188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. McDowell N, Phillips N, Lunch C, Bond B, Ryan M (2002) An investigation of hydraulic limitation and compensation in large, old Douglas-fir trees. Tree Physiol 22:763–774. [DOI] [PubMed] [Google Scholar]
- 67. Delzon S, Sartore M, Burlett R, Dewar R, Lousteau D (2004) Hydraulic responses to height growth in maritime pine trees. Plant Cell Environ 27:1077–1087. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Raw data for the vulnerability curves of the 39 study species analyzed. Filled dots represent measured data, empty dots show estimated Ψ50 values and the broken lines represent the fitted models of xylem vulnerability. For species abbreviations see Table 1.
(PNG)
Trait raw data for the 39 study species analyzed. For trait codes and full species names, please see Tables 1 and 2.
(TXT)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files. Requests can also be directed to Wenzel Kröber, wenzel.kroeber@botanik.uni-halle.de.