

Abstract
The reproductive biology encompassing phenology, floral biology, pollination and breeding systems, of Butea monosperma, a beautiful tree of the Indian subcontinent, was investigated in a protected dry, deciduous forest located in New Delhi. Phenological studies indicated that although the species shows a regular flowering season, all trees do not flower every year. Flowers are typically papilionaceous; the stigma is wet papillate and the style is hollow. The flowers show characteristics of bird pollination being large and bright orange‐red in colour with copious amounts of nectar, and exhibiting diurnal anthesis. Although the flowers are frequented by as many as seven species of birds belonging to six families, only one species, the purple sunbird (Nectarinia asiatica), is the effective pollinator. The flowers are also pollinated by the three‐striped squirrel (Funambulus tristiatus). Unlike other flower visitors, these two pollinators forage the nectar from the open side of the keel (legitimate path) during which pollen grains are deposited on their body parts. After the first visit of a sunbird or a squirrel, virgin flowers showed pollen load on the stigma and developed into fruits. B. monosperma shows a weak form of self‐incompatibility. Fruit set following manual self‐pollination (5·25 %) was comparable with open‐pollination (approx. 5 %) but was significantly lower than manual cross‐pollination (22·51 %). This indicates that there is a high degree of geitonogamous pollination in this species, which may lead to a weakening of self‐incompatibility as a means of reproductive assurance. The results are analysed in the light of prevailing discussions on specialized vs. generalized pollination systems.
Key words: Breeding system, Butea monosperma (Lam.) Taub., floral biology, ornithophily, squirrel‐pollination, tree legume
INTRODUCTION
The reproductive biology of flowering plants is important for determining barriers to seed and fruit set, for conservation, and for understanding pollination and breeding systems that regulate the genetic structure of populations. Detailed studies on reproductive biology of Indian plant species, especially trees, are limited. Out of nearly 2500 tree species reported from India, 180 are leguminous (Dr M. Sanjappa, Director, Botanical Survey of India, pers. comm.) being predominantly pollinated by bees. There are very few species that are pollinated by birds.
According to a survey on ornithophilous species in India, 93 are regularly visited by 58 species of birds (Subramanya and Radhamani, 1993). Over 80 % of these plant species are frequented by more than one species of bird and, similarly, over 80 % of the bird species visit more than one plant species. Thus, a generalized relationship exists between plant species and bird pollinators (Subramanya and Radhamani, 1993; Waser et al., 1996). In the majority of ornithophilous species, no attempts have been made to establish the obligate necessity of birds for pollination and to distinguish pollinators from floral visitors. More intensive studies are needed on Indian species for a better understanding of bird‐pollination systems.
Butea monosperma (Lam.) Taub (Fabaceae) is a medium‐sized tree of the Indian sub‐continent. It is an important species that yields several non‐timber forest products, such as a water‐soluble dye, lac resin, fodder and leaves for platters (Anon., 1988). The plant has beautiful flowers adorning leafless canopies during early summer and is aptly described as ‘the flame of the forest’. The flower colour has been variously described as scarlet, red and orange‐red (Anon., 1983, 1988, 1994) but, according to the Royal Horticultural Society colour chart (Anon., 1946), is most accurately termed Indian‐orange. Although rare, yellow and white flowering trees have also been reported (Sanjappa, 1987). Except for a report by Ali (1932) based on observations that several bird species visit the flowers, there have been no systematic studies on the reproductive biology of B. monosperma implicating bird pollination. The tree is largely propagated by seeds. Fruit set is very low and each fruit bears only one seed (hence the specific epithet monosperma).
In recent years, there has been a discussion on generalization vs. specialization in pollination systems (Waser et al., 1996; Johnson and Steiner, 2000). In B. monosperma, as many as 12 bird species have been reported to visit flowers (Subramanya and Radhamani, 1993). These features fit into the generalized relationship between flowers and bird pollinators. However, unlike many of the species pollinated by passerine birds in which flowers are open‐type and have unprotected nectar, B. monosperma bears flowers in which the nectar is concealed within the keel and requires a special foraging effort by the vector. Thus, a specialized relationship between the structural organization of the flower and the pollinator is implicated.
The present work on B. monosperma covers (a) the structural and temporal details of flowers to understand their relationship with pollination and fruit set, (b) pollination biology, (c) the breeding system and (d) fruit and seed biology. As the flowers are visited by several bird species, studies on pollination biology were aimed specifically at distinguishing floral visitors from pollinators and studying the involvement of any other vector in the pollination of this species.
MATERIALS AND METHODS
Study site
The study site is located (between 28°40′–28°35′N and 77°10′–77°30′E) in the Central Ridge Forest (CRF), representing the northern part of the Aravalli range in New Delhi. There are about 200 wild and sparsely distributed Butea monosperma trees in the CRF. Other trees that occur within the study site include Acacia leucophloea, A. modesta, Balanites roxburghii, Cordia dichotoma, Erythrina indica, Ehretia laevis, Prosopis juliflora, Wrightia tinctoria and Zizyphus mauritiana (Maheshwari, 1963).
Phenology and floral biology
Thirty trees were marked at various locations within the CRF for phenological studies. Of these, ten (marked B1–B10) were selected for detailed studies on floral biology, breeding system and pollination ecology.
Phenological events (bud break, flowering, fruiting, shedding of leaves, fresh leaf emergence, fruit dispersal and seed germination) were recorded over a period of two flowering seasons (1996 and 1997). To obtain the above information, observations were made every day in the morning hours during the flowering period (February–April). Subsequent events, such as fruit maturation and fruit dispersal, were noted and recorded once a week.
The average numbers of flower borne on an inflorescence were recorded from a set of randomly tagged flowering branches (n = 30). Five developmental stages (S1–S5) were recognized on the basis of morphometry, time of anther dehiscence and period of stigma receptivity. The details are presented in Table 1 and the specific stages used for emasculation and manual pollinations are specified. For estimating the average pollen grain numbers produced in a flower, mature but undehisced anthers (n = 20) were squashed in two or three drops of 25 % glycerol (v/v) + 1 drop of 1 % acetocarmine. Pollen grain viability at the time of anther dehiscence and later stages was estimated using the fluorescein diacetate (FDA) test (Heslop‐Harrison and Heslop‐Harrison, 1970). The presence of starch or lipids as a reserve material in the pollen grains was determined using I2KI and Sudan III+IV (Dafni, 1992), respectively.
Table 1.
Developmental stages of flowers of Butea monosperma identified for pollination studies
| Stage | Days to (–)/after (+) anthesis | Flower length (cm) | Colour and external features | Internal features |
| S1 | –1* | 4·5–5·4 | Wing petals not expanded | Anthers not dehisced, nectar absent |
| S2 | –1[+6–9 h] | 4·5–5·4 | Wing petals not expanded | Anthers dehisced, nectar absent |
| S3 | 0** | 5·0–6·0 | Anthesis beginning; expansion of wing petals and bending of the vexillum | Stigma receptive, nectar present, natural pollination occurs |
| S4 | 0 [+2 h]† | 5·0–6·0 | Flower fully opened | Stigma receptive, nectar present, natural pollination occurs |
| S5 | +2 | 5·0–6·0 | Petals and sepals start shrivelling, petal colour faded | Ovary swollen |
* Floral stage selected for emasculation.
† Stages for manual pollinations.
On the stigmatic surface, non‐specific esterases were localized using α‐naphthyl acetate (Mattsson et al., 1974) and phosphatases using α‐naphthyl acid phosphate (Scandlios, 1969) as substrates. Stigma receptivity and temporal details of post‐pollination events were recorded using the method described by Shivanna and Rangaswamy (1992). Pollen germination and pollen tube growth were examined using the aniline blue fluorescence method (Shivanna and Rangaswamy, 1992).
Structural details of the stigma and style were investigated using resin‐embedded sections (3 and 4 µm thick) (Fedder and O’Brien, 1968). The cuticle was localized using auramine O (Heslop‐Harrison, 1977), proteins using Coomassie brilliant blue R (Fisher, 1968), insoluble polysaccharides using PAS reagent (McGukin and Mackenzie, 1958), pectins using alcian blue (Heslop‐Harrison, 1979) and lipids using auramine O (Heslop‐Harrison, 1977). Ovule number was determined by clearing the fresh pistils (n = 20) with NaOH (1 n) and staining with 1 % acetocarmine.
Pollination
Nectar production was estimated every hour between 0500 and 1800 h. Flowers at the S2 stage (n = 25) were randomly selected on five trees and bagged (butter paper bags, 10 × 5 cm) 1 d before initiation of nectar production. On the day of anthesis, nectar was collected by gently inserting a calibrated micropipette through the opening of the keel. Flowers were re‐bagged after each collection.
Temporal activities of the floral visitors were recorded between 0430 and 1900 h over a 1‐week period in each season, using a pair of field binoculars. Each floral visitor was carefully observed for the time spent on each flower, and its mode of foraging noted. A distinction was made between the legitimate (those which foraged through the open margin of the keel petals) and the destructive (those which consumed nectar either by making a hole in the calyx or by damaging the floral parts) foragers. Pollination efficiency was assessed by exposing virgin flowers to a single visit by a vector and studying pollen load on the stigma and also by fruit production (Schemske and Horovitz, 1984). To study pollen load, stigmas from flowers visited by legitimate foragers (n = 10 each for sunbird and squirrel) were carefully excised in the field, placed on damp cotton in a screw‐capped vial (Tandon et al., 2001) and brought to the laboratory. A liquid fixative was deliberately not used in order to prevent dislodging of pollen grains. The stigma was carefully mounted in a drop of auramine O (a fluorescent dye for pollen exine) and observed under a fluorescence microscope. Pollen grains that were in contact with the stigma, as well those dislodged from the stigma, were counted. To determine pollination efficiency, a set of 48 freshly opened flowers which were legitimately foraged by squirrels (n = 30) and sunbirds (n = 18) were bagged and tagged to monitor fruit set.
Breeding system
On the basis of stigma receptivity, floral stages S3 and S4 were used for carrying out manual pollinations in the field. As fruit set under open‐pollination in this species is very low (approx. 5 %), the number of pollinations for each treatment had to be increased substantially to get a clear picture. To prevent autogamy, flowers were emasculated at the S1 stage using a fine pair of forceps and then bagged. The bags were opened on the day of anthesis and the flowers manually pollinated by gently brushing the receptive stigma with freshly dehisced anthers after which the flowers were re‐bagged. To effect cross‐pollination on emasculated and bagged flowers (n = 604), pollen grains were used from different trees and for geitonogamous self‐pollinations (n = 495) pollen grains were applied from other flowers of the same tree. To investigate any possible occurrence of apomixis, flowers at the S1 stage (n = 200) were emasculated and bagged without pollination. Spontaneous autogamy (n = 138) was tested by bagging unpollinated and unemasculated flowers.
In all manual pollinations, the bags were removed after 48 h to monitor flower retention. The index of self‐incompatibility (ISI) was expressed as the ratio of the number of fruits formed through manual self‐pollination to those formed through manual cross‐pollination (Zapata and Arroyo, 1978) at the end of 18–20 d. By randomly tagging the flowering branches (n = 100) on 25 trees in the population, the total number of flower buds borne and the number of fruits that developed were computed.
Fruit dispersal distance was computed by marking five isolated trees at different locations within the population. A red dot (4–5 mm diameter) was placed on the mature fruit (n = 50 each, on five separate marked individuals), close to the pedicel, when they were still attached to the tree. The distance travelled by fruits after their dispersal was measured from the base of the trunk of respective trees. Fruit densities at various distances from the tree were measured by laying down quadrats. Student’s t‐test (Sokal and Rohlf, 1969) was performed to establish the significance of the difference between the fruit set through manual cross‐ and self‐pollination on marked trees.
RESULTS
Phenology
Trees marked for phenological observations (n = 30) showed variation with respect to leaf fall and flowering. Twenty‐seven trees flowered in the first year but only 18 did so in the second. Trees marked for manual pollination flowered in both years. The trees that flowered began to shed their foliage in December and were completely bare by January. However, those that failed to flower retained their leaves. Occasionally, trees that flowered retained a scanty amount of foliage, especially on the lower branches.
The inception of inflorescence primordia in the leaf axils occurred at the end of February. The flower buds were black and velvety, and commenced opening during the first/second week of March. Peak flowering time in both years was the first week in April, trees remaining in bloom for 6–8 weeks. Fruiting commenced from the last week of March to the first week of April, the fruits reaching maturity by the end of May and dispersing in mid‐June. Seeds were not liberated from the fruits. Leaf primordia appeared in April/May and leaves attained their maximum size by May/June.
At the study site, B. monosperma shared its flowering period with that of other trees such as Ehretia laevis (Ehretiaceae) and Bauhinia variegata (Caesalpiniaceae). The declining flowering phase of B. monosperma coincided with the peak flowering phase of Erythrina indica (Fabaceae), another nectariferous tree with bright scarlet flowers that attract birds.
Floral biology
The inflorescence of B. monosperma is a profusely branched, fascicled, paniculate raceme, 20·97 ± 5·21 cm (n = 30) in length. On average, an inflorescence developed 166·21 ± 53·31 (n = 30) flowers that opened in acropetal order. Anthesis (flower bud opening) occurred between 0900 and 1030 h.
The flowers have a typical papilionaceous corolla (see Fig. 2A). The petals are bright Indian‐orange, silvery tomentose outside and glossy inside. During anthesis, the standard petal (vexillum) curves backward after the expansion of the wing petals. Each flower lasts 1–2 d. A gamosepalous cup‐like, green and fleshy calyx bulges out behind the standard petal. Of the ten stamens, nine are united and form a long staminal tube with two nostril‐like openings at the base. The distal end bears nine very short filaments, each terminating in a bilobed anther. The free stamen (vexillary stamen) is invariably shorter than the united stamens and is aligned with the bulged part of the calyx. Nectar is secreted by five nectaries located at the base of the ovary and accumulates in the calyx cup.

Fig. 2. A, A pair of freshly opened flowers showing erect keel, fully expanded lateral wings and the standard petal. The stamens and the pistil are enveloped in the keel. B, A female purple sunbird (solid arrow) withdrawing nectar from the opening of the keel (unfilled arrow). C, A three‐striped squirrel foraging freshly opened flowers. Pollen can be seen as yellow powder (small arrows) on the snout and the head (inset) of the squirrel. D, Squirrel pollinating a flower. Note that the head of the squirrel is in contact with the stigma (long arrow), effecting pollination.
Pollen grains are trizonocolporate and the exine is tectate‐reticulate (Fig. 1H). The average number of pollen grains in a flower was 52775·49 ± 1698·03 (n = 20). Whereas each anther of the nine united stamens contained 5382 ± 340·76 pollen grains, the anther from the free stamen bore a slightly lower number (4334·83 ± 329·22). About 37·3 % of pollen grains were sterile. They were shrivelled, lacked cytoplasmic contents and did not fluoresce when stained with FDA. Fertile and viable grains were 28 ± 1·36 µm in diameter, fluoresced brightly with FDA and contained starch as reserve material (Fig. 1I). Each ovary bore 5·43 ± 0·6 (n = 30) ovules; and the pollen to ovule ratio was 9719 : 1.
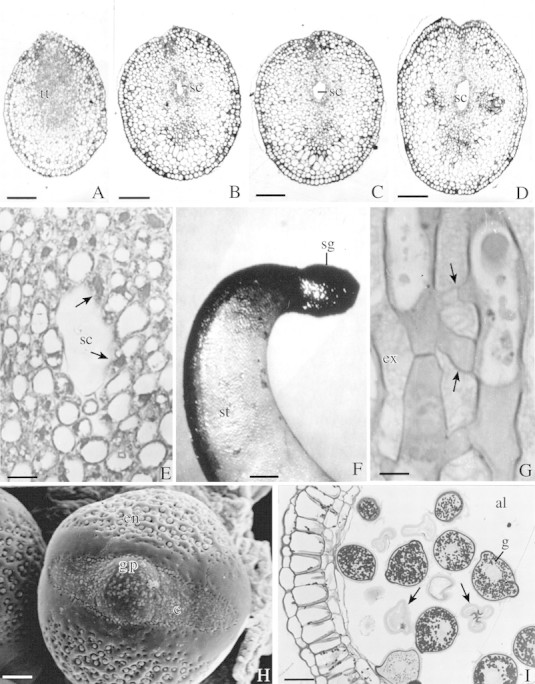
Fig. 1. A–D, Transverse sections of a style at different levels: just below the stigma (A), about 2 mm below the stigma (B), middle (C) and lower portion of the style. The style is solid just below the stigma; the stylar canal appears a few millimetres below the stigma and merges with the terminal part of the ovarian cavity. Bar = 125 µm. E, Enlarged view of transverse section of a style below the solid portion showing a canal formed as a result of lysis (arrows) of a few cells of the transmitting tissue. Bar = 65 µm. F, A receptive stigma with intense activity for non‐specific esterases. Bar = 1 mm. G, Longitudinal section of a portion of a stigma showing the septate inter‐papillar connections (arrows) and stigmatic exudate in the interstices between the papillae. Bar = 60 µm. H, Scanning electron micrograph of a tri‐zono‐colporate pollen grain presenting a prominent colpus with a germ pore. The exine is reticulate. Bar = 5 µm. I, Transverse section of a mature anther lobe stained with PAS reagent. Note fertile starchy pollen grains and collapsed sterile pollen grains (arrows). Bar = 65 µm. Abbreviations: al, anther locule; c, colpus; en, exine; ex, exudate, g, starch grains; gp, germ pore; sc, stylar canal; sg, stigma; st, style; tt, transmitting tissue.
Pistils were 5·19 ± 0·13 cm long at maturity and adaxially curved along the staminal tube. A globose, papillate and wet stigma is located near the tip of the keel, facing the standard petal. Papillae are unicellular. Young papillae are covered with a pellicle–cuticle layer and contain starch grains. At the time of anthesis the stigmatic exudate accumulates in the interstices of the papillae (Fig. 1G). The neighbouring stigmatic papillae are connected to each other through septate inter‐papillar connections (Fig. 1G). Each papilla is vacuolated and has a large nucleus.
The style subjacent to the stigma is solid (Fig. 1A). In transverse section it is 15–20 cells thick and made up of cortical cells with a core of transmitting tissue (Fig. 1A). Sections cut a few millimetres below the stigma showed a narrow stylar canal formed by the disintegration of a few transmitting tissue cells (Fig. 1B and E). The stylar canal gradually expands towards the base of the style and eventually joins the ovarian cavity (Fig. 1B–D).
Floral rewards and foragers
A profuse amount of nectar constitutes the primary floral reward for the visitors. Nectar is absent at the time of anther dehiscence but accumulates by the time the stigma becomes receptive. The quantity of nectar was measured on the day of flower opening at hourly intervals between 0500 and 1800 h. On average, the total amount of nectar collected during this period in each flower was 73 ± 18·31 µl. Maximum nectar production occurred between 0900 and 1100 h.
Freshly opened flowers are brightly coloured and attract a large number of visitors (Table 2). Maximum visitation occurred between 0900 and 1100 h. Rose‐ringed parakeets, purple sunbirds, white‐eyes and three‐striped squirrels were the most frequent visitors (Table 2).
Table 2.
Visitors to flowers of Butea monosperma
| Flower visitor | No. of visits in a day (n = 3 d) | Flower handling time (s) (n = 20) | Food and mode of foraging | No. of pollen grains deposited by legitimate visitors | Fruit set after visits by legitimate visitors (%) |
| Birds | |||||
| House crow (Corvus splendens) | 4 ± 2 | Does not forage | Does not forage, only perches | NR | NR |
| Indian tree pie (Dendrocitta vagbunda) | 3·66 ± 2·08 | Does not forage | Does not forage, only perches | NR | NR |
| Purple sunbird (Nectarinia asiatica) | 12·33 ± 2·51 | 3·72 ± 0·83 | Nectar consumed by legitimate foraging | 22·6 ± 10·79 (n = 10) | 5·55 (n = 18) |
| Rose‐ringed parakeet (Psittacula krameri) | 23 ± 9·53 | 9·75 ± 3·61 | Small fresh pistils removed from young flower buds | NR | NR |
| Red‐vented bulbul (Pycnonotus cafer) | 6·66 ± 1·15 | 3·30 ± 1·06 | Nectar consumed by making a hole in the calyx | NR | NR |
| Jungle babbler (Turdoides striatus) | 2·33 ± 1·52 | Does not forage | Does not forage | NR | NR |
| White eye (Zosterops palpebrosa) | 9·6 ± 5·5 | 2·0 ± 0·11 | Nectar consumed by making a hole in the calyx | NR | NR |
| Rodent | |||||
| Three‐striped squirrel (Funambulus tristiatus) | 7·6 ± 1·52 | 4·77 ± 1·84 | Nectar consumed by legitimate foraging | 44·1 ± 10·32(n = 10) | 6·66(n = 30) |
| Bee | |||||
| Giant Asian honeybee (Apis dorsata) | NR | 3·06 ± 1·18 | Left over nectar consumed from fresh holes made by other birds. | NR | NR |
NR, not recorded.
As nectar is located deep in the keel, penetration and active foraging becomes necessary to access it. Although many birds visit the flowers for nectar, purple sunbirds are the only ones that forage from the legitimate position between the standard and the keel (Fig. 2B). Short‐billed birds, such as white‐eye and red‐vented bulbul, do not forage for nectar from the legitimate side (i.e. the keel), preferring to pierce through the bulged side of the calyx cup to consume the nectar. Three‐striped squirrels also foraged the flower from the legitimate position.
Bees were the earliest visitors to be noted. However, the thick standard petal and the massive keel petals prevented their access to pollen or nectar. The bees could forage nectar only after a hole had been made in the calyx by birds, such as the white‐eye.
Pollination
The arrangement of the anthers is such that it maximizes pollen presentation when nectar is consumed from the opening of the keel. Stamens and anthers are placed at different heights in the staminal tube. Pollen presentation occurs when legitimate visitors press down the keel (carina). As it is lowered, the stiff pistil and the staminal tube emerge through the tip of the keel and rub against the body parts of the legitimate foragers. Smearing of pollen is facilitated by the fanning out of anthers borne at the ends of the filaments. Pollination occurs when the stigma touches the pollen‐covered crown of the pollinating sunbird or snout and head of a squirrel (Fig. 2C–D). Freshly opened flowers (n = 20) visited for the first time by squirrels (n = 10) or sunbirds (n = 10) resulted in successful deposition of pollen on the stigma. A stigma following the first visit of a squirrel received a higher number of pollen grains (44·1 ± 10·32), than one visited by a sunbird (22·69 ± 10·79). Flowers legitimately foraged by squirrels (n = 30) and sunbirds (n = 18) resulted in 6·66 % and 5·55 % fruit set, respectively (Table 2), which is comparable with fruit set under open‐pollination (approx. 5 %).
Breeding system
The flowers are protandrous. Anthers dehisce between 0830 and 1500 h 1 d before the mature flower buds open (anthesis). The stigma is receptive between 0700 and 1400 h on the day of anthesis and is covered by a large amount of exudate. The exudate stains intensely for non‐specific esterases (Fig. 1F), phosphatases and lipids.
Pollen viability tests were carried out at different times from anther dehiscence to 48 h after anthesis. Around 63 % of pollen grains were viable at the time of anther dehiscence. Viability declined to around 45 % after 24 h and to 30 % after 48 h at room temperature. Pollen grains collected 30–60 min after anther dehiscence were used for manual pollinations.
All flowers (n = 200) emasculated and bagged to ascertain the occurrence of apomixis abscised within 48 h of bagging. Flowers bagged to check for spontaneous autogamy (n = 138) also dropped at different times during the 48‐h period. The percentage fruit set through manual cross‐pollinations was significantly higher (22·51 %) than through manual self‐pollinations (5·25 %) (P < 0·05, t‐value = 8·2, d.f. = 18) (Table 3). Ten trees (B1–B10) used for manual pollinations showed variation in the indices of their compatibility. Some trees (B3, B4 and B5) did not set fruit through self‐pollination, whereas others set approx. 5 % of fruits. Fruit set of cross‐pollinated flowers on all trees was much higher (18–27 %) than that of self‐pollinated flowers. Although there are four or five ovules in each ovary, successful pollination invariably leads to the formation of only one seed in a fruit. Only the ovule located at either the first or the second position towards the stylar end matures into a seed. The remaining ovules abort.
Table 3.
Percentage fruit set in controlled manual pollinations
| Type of pollination/treatment | No. of flowers treated | Fruit set (%) |
| Apomixis* | 200 | 0 |
| Spontaneous autogamy | 138 | 0 |
| Geitonogmay | 495 | 5·25† |
| Xenogmay | 604 | 22·51† |
* Emasculated and bagged, without pollinations.
† The difference in fruit set by cross‐ and self‐pollinations was significant (P < 0·05, t‐value = 8·2, d.f. = 18).
Fruit dispersal and natural recruitment
The fruit of B. monosperma is a single‐seeded samara (Augspurger, 1989) and disperses as a diaspore. Fruits were seen to drop in and around the vicinity of the tree on which they were produced and did not travel much distance from it. Observations on isolated trees (n = 5) at different locations within the population showed that the farthest distance to which fruits dispersed was 10·3 ± 2 m from the base of the trunk, which was only 2 ± 1·5 m outside the canopy area. Maximum fruit density (5·9 pods m–2) was recorded between 4 and 7 m from the tree trunk. Natural germination occurs during the monsoon season (July–August).
Prior to seed germination, the pericarp of a dispersed fruit opens to form a small slit near the stylar end. As the slit is shorter than the length of the seed, the seed is not liberated from the fruit. Seed germination occurs in situ. The radicle emerges first, followed by elongation of the cotyledonary petiole. The plumule emerges from the envelopment of the cotyledonary petiole. The starchy cotyledons remain inside the pericarp. This type of germination has been described as Helciopsis by de Vogel (1980). The first pair of leaves is simple. In the first two years after germination, the shoot dies back during summers (April–June) and a fresh shoot is formed from the underground part of a 1‐year‐old seedling during the rainy season.
DISCUSSION
This paper reports the first comprehensive study of the reproductive biology of Butea monosperma. Like many other tropical trees, B. monosperma has a regular flowering season. However, all trees did not flower during the two years devoted to the phenological study. Out of 30 trees, three failed to flower in the first year and 12 in the second. The structural organization of the flower, including that of the stigma and the style, of B. monosperma is comparable with that of other papilionoid legumes so far reported (Shivanna and Owens, 1989) indicating that these features are conserved.
Pollination system
The flowers of B. monosperma show many features, such as a bright reddish‐orange colour, abundant nectar protected by the keel, and diurnal anthesis, characteristic of bird pollination (Faegri and van der Pijl, 1979). Present studies on pollination biology have revealed an unusual pollination guild in B. monosperma. As indicated by floral morphology, flowers are frequented by as many as seven bird species belonging to six families. Of these, only one species, Nectarinia asiatica, forages from the legitimate position and brings about effective pollination. All other bird species pierce their bills through the calyx to reach the nectar. As they do not come into direct contact with the anthers and the stigma, they have no role in pollination. Although honeybees were seen around flowers, they were observed to forage the nectar only from the holes made in the calyx by non‐pollinating birds. The flowers are too large, and the stigma and anthers remain out of reach, for the bees to bring about pollination. As suggested by Johnson and Steiner (2000), the results of the present study show clearly that the large number of pollinating species frequenting a plant species is not a satisfactory method to measure the degree of specialization; the active pollinators comprise only a very small fraction of the total number of visitors. The findings reported here have established that B. monosperma demonstrates a specialized bird‐pollination syndrome (Johnson and Steiner, 2000).
Sunbirds constitute one of the most important groups of bird pollinators and are reported to frequent up to 58 species of flowering plants in India (Subramanya and Radhamani, 1993). Interestingly, an analysis of the ratio of beak length to body size of all visitors to B. monosperma flowers showed that sunbirds had a much higher ratio (0·210) compared with all other visitors (0·125–0·074) (Subramanya and Radhamani, 1993). Elongated, gently decurved beaks seem to facilitate sunbirds to harvest the deep‐seated nectar from flowers in a large number of plant species.
Squirrels foraged from the legitimate position, the stigma and anthers come into contact with their snouts and heads. Furthermore, after the first visit by purple sunbirds or squirrels, pollen was deposited on the stigmas of visited flowers, which subsequently developed into fruits. These results were comparable to those obtained by open‐pollination, confirming that these two foragers are indeed the pollinators. Nectar is their reward. Nectar production is confined to the period during which the stigma is receptive. Visits by both purple sunbirds and squirrels were correlated with the availability of nectar.
No unique morphological features that would facilitate squirrel pollination were identified in B. monosperma flowers. However, many features of ornithophily, such as large, robust flowers (that could withstand the foraging activity of squirrels), abundant nectar, anthers and stigma positions, and diurnal opening of the flowers, are ideal for squirrel foraging and pollination. Squirrels are generally territorial and restrict their movement to one or a few neighbouring trees. Their intensive search for food could have led them to explore a vast number of very conspicuous flowers which opportunistically would have brought them into contact with nectar. Due to easy floral access and the availability of abundant nectar, visits would have stabilized without requiring any change in floral features. Thus, flowers of B. monosperma conform to the classical bird pollination syndrome but are flexible enough to accommodate opportunistic pollinators belonging to an entirely different group of animals (Johnson and Steiner, 2000). Pollination by squirrels has been reported in Grevillea robusta (McCann, 1933) and in Ganua sp. (Yumoto et al., 1996), although these taxa do not show any special features for squirrel pollination. In Ganua, the flowers are open and it is suggested that the fleshy perianth with attached stamens serves as a reward for the squirrels.
The pollination system in B. monosperma involving birds and squirrels is an unusual pollination guild. In another papilionoid legume, Erythrina sp. (Bruneau, 1997), there are two modes of pollination, one by hummingbirds and the other by passerine birds. These pollination modes essentially require structural changes in the flower’s morphology. In hummingbird‐pollinated Erythrina sp., the standard petal is conduplicately folded to form a pseudo‐tube, whereas in passerine‐pollinated flowers the standard petal is open so that nectar is visible and accessible. However, in Erythrina crista‐galli, Galetto et al. (2000) noticed that, in addition to the birds, bees also pollinated the flowers, indicating an intermediate step from entomophily (typical of the tribe Phaseoleae) to ornithophily (typical of Erythrina). In B. monosperma, two different groups of vectors are able to act as effective pollinators without any change being necessary in the floral features. Bird‐ (ornithophily) and squirrel‐pollination (therophily) in this species is thus an atypical pollinator guild in the tribe Phaseoleae.
Breeding system
Among the Leguminosae, arboreal species are reported to maintain a high level of out‐crossing, in contrast to herbaceous species in which autogamy predominates (Arroyo, 1981; Bawa, 1992). However, the breeding system in B. monosperma is not sharply defined. Fruit set following manual self‐pollination was very low (5·21 %). Manual cross‐pollination increased it to 23·32 %, indicating a weak form of self‐incompatibility. The self‐incompatibility index (Zapata and Arroyo, 1978) was 0·19, very close to the arbitrary proposed number of 0·20 required to characterize the species as self‐incompatible. Interestingly, fruit set, even under open‐pollination, was only 5·0 %, comparable with that of self‐pollinated flowers. Since there was no spontaneous autogamy, as indicated by lack of fruit set in unemasculated and bagged flowers, these results indicate that under open‐pollination, pollen load in B. monosperma is largely geitonogamous.
de Jong et al. (1993) stated that geitonogamy is substantial in mass flowering tropical trees. The work reported here on B. monosperma supports this. Availability of large flower numbers on each tree facilitates pollinator visits to more flowers of the same tree in succession. Geitonogamy incurs a fitness cost of reduced pollen export and seed set, particularly in self‐incompatible species (de Jong et al., 1993). In strongly self‐incompatible Polemonium viscosum, deposition of self‐pollen 24 h prior to the deposition of compatible pollen reduced germination of compatible pollen by 32 % and seed set by 40 % (Galen et al., 1989). In B. monosperma, geitonogamy is more prevalent in squirrel‐pollinated flowers, as they have a narrow territorial range and visit mostly flowers on the same tree. Purple sunbirds have a wider territorial range and are likely to import, although infrequently, pollen from other trees. The weak self‐incompatibility in B. monosperma appears to be a feature developed in response to extensive geitonogamous selfing prevalent in the species. Lloyd (1992) also observed that a weak form of self‐incompatibility provides flexibility under which out‐crossing cannot always be guaranteed.
Fruit and seed biology
Several factors may be responsible for the low fruit set in B. monosperma under open‐pollination. Obviously, limitation of compatible pollen is one factor. However, even in manual cross‐pollinations, fruit set was only increased to 23·32 %, indicating that there are additional constraints for fruit set which have not been investigated.
Although there are four or five ovules in each ovary of B. monosperma, only one ovule, usually the one located closest to the style, develops into the seed. In Dalbergia sissoo (Mohan Raju et al., 1996) and Syzigium cumini (Krishnamurthy et al., 1997), in which a single seed is predominantly formed, abortion of other seeds has been reported to be caused by the production of a chemical of indole nature, such as 4‐chloroindoleacetic acid. This aspect has not been investigated in B. monosperma. Augspurger (1989) described B. monosperma fruits as wind‐dispersed. However, our results indicate that wind is not effective in fruit dispersal. Fruits generally fall under the canopy or slightly away from it. As a consequence of fruits travelling, at the most, only a few metres from the trunk of a tree, the resulting population would comprise genetically related neighbouring individuals (see also Levin, 1981). This feature would further increase homozygosity of the population.
ACKNOWLEDGEMENTS
R.T. acknowledges the award of a Junior and a Senior Research Fellowship by the University Grants Commission. H.Y.M. thanks the Indian National Science Academy for financial assistance.
Supplementary Material
Received: 23 April 2003;; Returned for revision: 18 June 2003. Accepted: 28 July 2003; Published electronically: 19 September 2003
References
- AliSA.1932. Flower birds and bird flowers in India. Journal of Bombay Natural History Society 35: 573–605. [Google Scholar]
- Anon.1946.Horticultural colour chart II, Vols I and II. Wilson Colour Ltd and Royal Horticultural Society. [Google Scholar]
- Anon.1983.Troup’s silviculture of Indian trees, Vol. IV, Leguminosae. Delhi: Forest Research Institute and Colleges, Government of India, Controller of Publications. [Google Scholar]
- Anon.1988.Butea monosperma (Lam.) Taub. The wealth of India – raw materials revised, Vol. II: B. New Delhi: Council of Scientific and Industrial Research. [Google Scholar]
- Anon.1994.Indian medicinal plants: a compendium of 500 species, Vol. 1 Madras: Orient Longman Ltd, 314–318. [Google Scholar]
- ArroyoMTK.1981. Breeding systems and pollination biology of Leguminosae. In: Polhill RM, Raven PH, eds. Advances in legume systematics Proceedings of the International Legume Conference, 1978. Royal Botanic Gardens, Kew: Royal Botanic Gardens, 723–769. [Google Scholar]
- AugspurgerCK.1989. Morphology and aerodynamics of wind‐dispersed legumes. In: Stirton CH, Zarucchi JL, eds. Advances in legume biology Proceedings of the Second International Legume Conference, St Louis, Missouri Missouri Botanical Garden 29: 451–466. [Google Scholar]
- BawaKS.1992. Mating systems, genetic differentiation and speciation in tropical rain forest plants. Biotropica 24: 250–255. [Google Scholar]
- Bruneau.A.1997. Evolution and homology of bird pollination syndromes in Erythrina (Leguminoasae). American Journal of Botany 84: 54–71. [Google Scholar]
- DafniA.1992.Pollination ecology: a practical approach. New York: Oxford University Press. [Google Scholar]
- deJongTJ,Waser NM, Klinkhamer PGL.1993. Geitonogamy: the neglected side of selfing. Trends in Ecology and Evolution 8: 321–325. [DOI] [PubMed] [Google Scholar]
- deVogelEF.1980.Seedlings of dicotyledons. Wageningen: Centre for Agricultural Publishing and Documentation. [Google Scholar]
- FaegriK, van der Pijl L.1979.The principles of pollination ecology. Oxford: Pergamon Press. [Google Scholar]
- FedderN, O’BrienTP.1968. Plant microtechnique: some principles and new methods. American Journal of Botany 55: 123–142. [Google Scholar]
- FisherDB.1968. Protein staining of ribboned epon sections for light microscopy. Histochemie 16: 92–96. [DOI] [PubMed] [Google Scholar]
- GalenC, Gregory T, Galloway LF.1989. Cost of self‐pollination in a self‐incompatible plant, Plemonium viscosum American Journal of Botany 76: 1675–1680. [Google Scholar]
- GalettoL, Bernardello G, Isele IC, Vesprini J, Speroni G, Berduc A.2000. Reproductive biology of Erythrina crista‐galli (Fabaceae). Annals of the Missouri Botanical Garden 87: 127–145. [Google Scholar]
- Heslop‐HarrisonJ, Heslop‐Harrison Y.1970. Evaluation of pollen viability by enzymatically‐induced fluorescence; intracellular hydro lysis of fluorescein diacetate. Stain Technology 45: 115–120. [DOI] [PubMed] [Google Scholar]
- Heslop‐HarrisonY.1977. The pollen–stigma interaction: pollen tube penetration in Crocus Annals of Botany 41: 913–922. [Google Scholar]
- Heslop‐HarrisonY.1979. Aspects of structure, cytochemistry and germination of pollen of rye (Secale cereale L.). Annals of Botany 44 (Suppl.): 1–7. [Google Scholar]
- JohnsonSD, Steiner KE.2000. Generalization versus specialization in plant pollination systems. Trends in Ecology and Evolution 15: 140–143. [DOI] [PubMed] [Google Scholar]
- KrishnamurthyKS, Uma Shaanker R, Ganeshaiah KN.1997. Seed abortion in an animal dispersed species, Syzgium cuminii (L.) Skeels (Myrtaceae): the chemical basis. Current Science 73: 869–873. [Google Scholar]
- LevinDA.1981. Dispersal versus gene flow in plants. Annals of the Missouri Botanical Gardens 68: 233–253. [Google Scholar]
- LloydDG.1992. Self‐ and cross‐fertilization in plants. II. The selection of self‐fertilization. Journal of Plant Science 153: 370–380. [Google Scholar]
- McCannC.1933. The flying fox (P. giganteus) and the palm squirrel (F. tristiatus) as agents of pollinization in (Grevillea robusta A. Cunn.) the silky oak. Journal of Bombay Natural History Society 36: 761–764. [Google Scholar]
- McGukinWE, Mackenzie BF.1958. An improved periodic acid fuschin sulphite staining method for evaluation of glycoproteins. Clinical Cytochemistry 4: 476–483. [Google Scholar]
- MaheshwariJK.1963.Flora of Delhi. New Delhi: Council of Scientific and Industrial Research. [Google Scholar]
- MattssonO, Knox RB, Heslop‐Harrison J, Heslop‐Harrison Y.1974. Protein pellicle of stigmatic papillae as a probable recognition site in incompatibility reactions. Nature 247: 298–300. [Google Scholar]
- Mohan RajuB, Uma Shaanker R, Ganeshaiah KN, Bawa KS.1996. Intra‐fruit seed abortion in a wind dispersed tree, Dalbergia sissoo Roxb: proximate mechanisms. Sexual Plant Reproduction 9: 273–278. [Google Scholar]
- SanjappaM.1987. Revision of the genera Butea Roxb. Ex Willd. and Meizotropis Voigt. (Fabaceae). Bulletin of the Botanical Survey of India 29: 199–225. [Google Scholar]
- ScandliosJG.1969. Genetic control of multiple molecular forms of enzymes in plants: a review. Biochemical Genetics 3: 37–79. [Google Scholar]
- SchemskeDW, Horovitz CC.1984. Variation among floral visitors in pollination ability: a precondition for mutualism specialization. Science 255: 519–521. [DOI] [PubMed] [Google Scholar]
- ShivannaKR, Owens SJ.1989. Pollen–pistil interactions (Papilion oideae). In: Stirton CH, Zarucchi JL, eds. Advances in legume biology Proceedings of the Second International Legume Conference, St Louis, Missouri Missouri Botanical Garden 29: 157–182. [Google Scholar]
- ShivannaKR, Rangaswamy NS.1992.Pollen biology: a laboratory manual. Berlin: Springer Verlag. [Google Scholar]
- SokalRE, Rohlf FJ.1969.Biometery: the principles and practice of statistics in biological research. San Francisco: Freeman. [Google Scholar]
- SubramanyaS, Radhamani TR.1993. Pollination by birds and bats. Current Science 65: 201–209. [Google Scholar]
- TandonR, Manohara TN, Nijalingappa BHM, Shivanna KR.2001. Pollination and pollen–pistil interaction in oil palm, Elaeis guineensis. Annals of Botany 87: 831–838. [Google Scholar]
- WaserNM, Chittka L, Price MV.1996. Generalization in pollination systems, and why it matters. Ecology 77: 1043–1060. [Google Scholar]
- YumotoT, Momose K, Nagamasu H.1996. A new pollination syndrome – squirrel pollination in a tropical rain forest in Lambir hills national park. Tropics 9: 147–151. [Google Scholar]
- ZapataTR, Arroyo MTK.1978. Plant reproductive ecology of a secondary deciduous tropical forest in Venezuela. Biotropica 10: 221–230. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}