

Abstract
CONTENTS
| INTRODUCTION | 45 |
|---|---|
| PROGRESS | 46 |
| Improved systematic representation (species and families) | 46 |
| (i) First estimates for species | 46 |
| (ii) First estimates for families | 47 |
| PROBLEMS | 48 |
| Geographical representation and distribution | 48 |
| Plant life form | 48 |
| Obsolescence time bomb | 49 |
| Errors and inexactitudes | 49 |
| Genome size, ‘complete’ genome sequencing, and, the euchromatic genome | 50 |
| The completely sequenced genome | 50 |
| Weeding out erroneous data | 52 |
| What is the smallest reliable C-value for an angiosperm? | 52 |
| What is the minimum C-value for a free-living angiosperm and other free-living organisms? | 53 |
| PROSPECTS FOR THE NEXT TEN YEARS | 54 |
| Holistic genomics | 55 |
| LITERATURE CITED | 56 |
| APPENDIX | 59 |
| Notes to the Appendix | 59 |
| Original references for DNA values | 89 |
• Background The nuclear DNA amount in an unreplicated haploid chromosome complement (1C-value) is a key diversity character with many uses. Angiosperm C-values have been listed for reference purposes since 1976, and pooled in an electronic database since 1997 (http://www.kew.org/cval/homepage). Such lists are cited frequently and provide data for many comparative studies. The last compilation was published in 2000, so a further supplementary list is timely to monitor progress against targets set at the first plant genome size workshop in 1997 and to facilitate new goal setting.
• Scope The present work lists DNA C-values for 804 species including first values for 628 species from 88 original sources, not included in any previous compilation, plus additional values for 176 species included in a previous compilation.
• Conclusions 1998–2002 saw striking progress in our knowledge of angiosperm C-values. At least 1700 first values for species were measured (the most in any five-year period) and familial representation rose from 30 % to 50 %. The loss of many densitometers used to measure DNA C-values proved less serious than feared, owing to the development of relatively inexpensive flow cytometers and computer-based image analysis systems. New uses of the term genome (e.g. in ‘complete’ genome sequencing) can cause confusion. The Arabidopsis Genome Initiative C-value for Arabidopsis thaliana (125 Mb) was a gross underestimate, and an exact C-value based on genome sequencing alone is unlikely to be obtained soon for any angiosperm. Lack of this expected benchmark poses a quandary as to what to use as the basal calibration standard for angiosperms. The next decade offers exciting prospects for angiosperm genome size research. The database (http://www.kew.org/cval/homepage) should become sufficiently representative of the global flora to answer most questions without needing new estimations. DNA amount variation will remain a key interest as an integrated strand of holistic genomics.
Keywords: Angiosperm DNA amounts, DNA C-values, nuclear genome size, plant DNA C-values database
INTRODUCTION
It has been possible to estimate the amount of DNA in plant nuclei for over 50 years, and since the key role of DNA in biology was discovered in 1953, such research has increased in each successive decade. Work on plants has played a leading part in research to describe and understand the origin, extent and effects of variation in the DNA amount in the unreplicated haploid nuclear chromosome complement (defined by Swift, 1950, as the 1C-value) of different taxa. Indeed, angiosperms are probably the most intensively studied major taxonomic ‘group’ of organisms, with published C-values for over 4100 species.
Early research to address questions such as possible relationships between DNA C-value and the rate of cell development (e.g. Van't Hof, 1965) usually required work to estimate C-values for most of the taxa concerned, as these were unavailable. Later, as taxa with ‘known’ C-values increased, it was possible to use such data in new comparisons (supplemented by further first estimates made for sample taxa). However, it was often difficult to know whether a C-value existed for a particular taxon, and if so, where to find it. Such estimates were widely scattered in the literature or even unpublished. Small lists of nuclear DNA amounts were published in reviews and research papers, but the first large list of DNA amounts for angiosperms, compiled primarily as a reference source was published in 1976. This contained data for over 750 species from 54 original sources (Bennett and Smith, 1976), and noted an intention to publish supplementary lists for reference purposes at intervals. Five such lists, together giving pooled data for over 2900 species from 323 original sources, have followed (Bennett et al., 1982, 2000; Bennett and Smith, 1991; Bennett and Leitch, 1995, 1997). Data from the first five publications were pooled in an electronic form – the Angiosperm DNA C-values database, which went live in April 1997. This was updated as release 3.1 and incorporated, with databases for gymnosperms, pteridophytes and bryophytes, into the Plant DNA C-values database (release 1.0) in 2001.
These data are clearly much used, as the published lists have been cited over 1400 times, including over 700 times since 1997, whilst the electronic database has received over 50000 hits. Recently they have provided the large samples of data needed for many diverse comparative studies, such as testing for possible relationships between nuclear DNA amount and risk of extinction (Vinogradov, 2003), ecological factors in California (Knight and Ackerly, 2002), lead pollution in Slovenia (B. Vilhar, University of Ljubljana, Slovenia, pers. comm.); ploidy level (Leitch and Bennett, 2004), and land plant evolution (Leitch et al., 2005).
Given their ongoing use as reference sources, publication of a sixth supplementary list of angiosperm C-values is timely, if not overdue. The present work lists DNA C-values for 804 species from 88 original sources, including first estimates for 628 species not included in any previous compilation, plus additional estimates for 176 species already included in one or more previous compilation. Data in the Appendix table were prepared for analysis at the second Plant Genome Size Discussion Meeting in September 2003, so it is fitting that they are included in this special supplement. Whilst they represent most of the new C-value data published or estimated in 2000–2002, we are already aware of a further large sample estimated but unpublished either by late 2002, or subsequently. Thus, despite its large size, the present list will soon be followed by a seventh supplement.
Appendix.
Chromosome number, ploidy level, life-cycle type, and nuclear DNA content in 804 angiosperm species (the superscript letters refer to notes preceding this table)
| DNA amount |
||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Entry numberg |
Species |
Voucher |
Family |
Higher group# |
2n‡ |
Ploidylevel (x) |
Life cycletype§ |
1C (Mbps) |
1C (pg) |
2C (pg) |
4C (pg) |
Original ref.a |
Present amount† |
Standard species*b1 |
Method†† |
|||
| 1a | Acacia dealbata Link. | No | Leguminosae | E | 26 | 2 | P | 853 | 0·9 | 1·7 | 3·5 | 441bb | O | J | FC:PI | |||
| 2 | Acacia dealbata Link. | No | Leguminosae | E | 39° | 3 | P | —t | —t | 2·5 | 5·1 | 441bb | O | J | FC:PI | |||
| 3 | Acacia dealbata Link. | No | Leguminosae | E | 52 | 4 | P | 1,671 | 1·7 | 3·4 | 6·8 | 441bb | O | J | FC:PI | |||
| 4a | Acacia mangium Willd. | No | Leguminosae | E | 26 | 2 | P | 637 | 0·7 | 1·3 | 2·6 | 441bb | O | J | FC:PI | |||
| 5 | Acridocarpus natalitius A. Juss. | No | Malpighiaceae | E | c. 216 | 24 | P | 1,490 | 1·5 | 3·0 | 6·1 | 379 | O | J | Fe | |||
| 6 | Adenanthera microsperma Teijsm & Binn. | No | Leguminosae | E | —n | —p | P | 681 | 0·7 | 1·4 | 2·8 | 454 | O | Bc | Fe | |||
| 7 | Adenanthera pavonina L. | No | Leguminosae | E | 26° | —p | P | 666 | 0·7 | 1·4 | 2·7 | 454 | O | Bc | Fe | |||
| 8 | Adina cordifolia (Roxb.) Hook. f. | No | Rubiaceae | E | 22° | 2 | P | 816 | 0·8 | 1·7 | 3·3 | 454 | O | Bc | Fe | |||
| 9 | Aeonium haworthii Webb & Berth. | No | Crassulaceae | E | 72° | 4 or 8 | P | 760 | 0·8 | 1·6 | 3·1 | 378 | O | J | Fe | |||
| 10a | Aesculus hippocastanum L. | No | Sapindaceae | E | 40 | 2 | P | 588 | 0·6 | 1·2 | 2·4 | 465 | O | Lycopers.c | FC:PI | |||
| 11b | Agave americana L. | —m | Asparagaceae | M | 120 | 4 | P | 7,791 | 8·0 | 15·9 | 31·8 | 384aa | O | Homof | FC:PI | |||
| 12d | Agave sisalana Perr. | —m | Asparagaceae | M | 150 | 5 | P | —t | —t | 20·0 | 40·0 | 384aa | O | Homof | FC:PI | |||
| 13 | Agave stricta Salm. | —m | Asparagaceae | M | 60 | 2 | P | 3,822 | 3·9 | 7·8 | 15·6 | 384aa | O | Homof | FC:PI | |||
| 14 | Agrostis palustris Huds. | No | Gramineae | M | 28 | 4 | P | 2,769 | 2·8 | 5·7 | 11·3 | 417 | O | Gallusf | FC:PI | |||
| 15 | Ailanthus grandis Prain | No | Simaroubaceae | E | 64° | —p | P | 2,134 | 2·2 | 4·4 | 8·7 | 454 | O | Bc | Fe | |||
| 16a | Albuca pendula B.Mathew | No | Asparagaceae | M | 16 | 2 | P | 2,967 | 3·0 | 6·1 | 12·1 | 465 | O | G | Fe | |||
| 16b | Albuca pendula B.Mathew | No | Asparagaceae | M | 14 | 2 | P | 3,033 | 3·1 | 6·2 | 12·4 | 465 | O | G | Fe | |||
| 17k | Allium cepa L. | No | Alliaceaek | M | 16° | 2 | P | 16,415 | 16·8 | 33·5 | 67·0 | 457bm | O | Bd | Fe | |||
| 18 | Allocasuarina verticillata (Lam.) L.Johnson | No | Casuarinaceae | E | 20-28° | 2 | P | 931 | 1·0 | 1·9 | 3·8 | 452 | O | Petuniae | FC:PI | |||
| 19 | Alocasia cucullata (Lour) Schott | No | Araceae | M | 98 | —p | AP | 8,200 | 8·4 | 16·7 | 33·5 | 411 | O | Bc | Fe | |||
| 20 | Alocasia hilobeauty Host. | No | Araceae | M | 32 | —p | A | 3,680 | 3·8 | 7·5 | 15·0 | 411 | O | Bc | Fe | |||
| 21 | Aloe albiflora Guillaumin | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,337 | 15·7 | 31·3 | 62·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 22 | Aloe alooides (Bolus) Druten | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,083 | 13·4 | 26·7 | 53·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 23 | Aloe antandroi (Decary) H.Perrier | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,199 | 17·6 | 35·1 | 70·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 24 | Aloe arborescens (yellow flowers) Mill.i | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,671 | 14·0 | 27·9 | 55·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 25a | Aloe aristata Haw. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,729 | 16·1 | 32·1 | 64·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 25b | Aloe aristata var. parvifolia Baker Haw. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,023 | 16·4 | 32·7 | 65·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 26 | Aloe bakeri Scott-Elliot | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,925 | 16·3 | 32·5 | 65·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 27 | Aloe barberae Dyer | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,043 | 15·4 | 30·7 | 61·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 28 | Aloe bellatula Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,268 | 16·6 | 33·2 | 66·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 29 | Aloe boiteaui Guillaumin | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,023 | 16·4 | 32·7 | 65·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 30 | Aloe bowiea Schult. & Schult.f. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,268 | 16·6 | 33·2 | 66·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 31 | Aloe brevifolia Mill. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,553 | 14·9 | 29·7 | 59·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 32b | Aloe cameronii Hemsl. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,052 | 17·4 | 34·8 | 69·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 33 | Aloe capitata Baker | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,386 | 15·7 | 31·4 | 62·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 34 | Aloe chabaudii Schonland | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,934 | 18·3 | 36·6 | 73·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 35 | Aloe ciliaris var. tidmarshii Schonland Haw. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 10,535 | 10·8 | 21·5 | 43·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 36 | Aloe ciliaris Haw. | No | Xanthorrhoeaceaek | M | 35° | 5 | P | —t | —t | 53·3 | 106·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 37 | Aloe ciliaris Haw. | No | Xanthorrhoeaceaek | M | 42° | 6 | P | 30,723 | 31·4 | 62·7 | 125·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 38 | Aloe comptonii Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,426 | 13·7 | 27·4 | 54·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 39a | Aloe cryptopoda Baker | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,161 | 14·5 | 28·9 | 57·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 39b | Aloe cryptopoda “Wickensii” Baker | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,357 | 14·7 | 29·3 | 58·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 40 | Aloe dawei Berger | No | Xanthorrhoeaceaek | M | 28° | 4 | P | 35,231 | 36·0 | 71·9 | 143·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 41a | Aloe descoingsii Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,974 | 16·3 | 32·6 | 65·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 41b | Aloe descoingsii Reynolds ssp. augustina Lavranos | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,219 | 16·6 | 33·1 | 66·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 42a | Aloe dichotoma Masson var. ramosissima (Pillans) Glen & D.S.Hardy | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,005 | 12·3 | 24·5 | 49·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 42b | Aloe dichotoma Masson | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,103 | 12·4 | 24·7 | 49·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 43 | Aloe dinteri A.Berger | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,366 | 16·7 | 33·4 | 66·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 44b | Aloe distans Haw. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,622 | 13·9 | 27·8 | 55·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 45 | Aloe dorotheae A.Berger | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,288 | 15·6 | 31·2 | 62·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 46 | Aloe elegans Tod. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,346 | 17·7 | 35·4 | 70·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 47 | Aloe erinacea D.S.Hardy | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,103 | 12·4 | 24·7 | 49·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 48 | Aloe ferox Mill. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,896 | 15·2 | 30·4 | 60·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 49 | Aloe fleurentiniorum Lavranos & L.E.Newton | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 18,179 | 18·6 | 37·1 | 74·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 50 | Aloe gariepensis (?) Pillans | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,729 | 16·1 | 32·1 | 64·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 51 | Aloe glauca Mill. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,680 | 16·0 | 32·0 | 64·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 52 | Aloe globuligemma Pole-Evans | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,611 | 17·0 | 33·9 | 67·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 53 | Aloe haemanthifolia A.Berger & Marloth | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 7,938 | 8·1 | 16·2 | 32·4 | 440 | O | Agave sp.ab | FC:PI | |||
| DNA amount |
||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Entry numberg |
Species |
Voucher |
Family |
Higher group# |
2n‡ |
Ploidylevel (x) |
Life cycletype§ |
1C (Mbps) |
1C (pg) |
2C (pg) |
4C (pg) |
Original ref.a |
Present amount† |
Standard species*b1 |
Method†† |
|||
| 54 | Aloe haworthioides Baker | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,749 | 15·1 | 30·1 | 60·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 55 | Aloe hereroensis Engl. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 18,130 | 18·5 | 37·0 | 74·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 56 | Aloe humilis (small form) (L.) Mill.i | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,562 | 16·9 | 33·8 | 67·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 57 | Aloe jacksonii Reynolds | No | Xanthorrhoeaceaek | M | 28° | 4 | P | 32,487 | 33·2 | 66·3 | 132·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 58 | Aloe jucunda Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,591 | 18·0 | 35·9 | 71·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 59b | Aloe juvenna Brandham & Carter | No | Xanthorrhoeaceaek | M | 28° | 4 | P | 34,790 | 35·5 | 71·0 | 142·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 60 | Aloe krapohliana var. dumoulinii Lavranos | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,346 | 17·7 | 35·4 | 70·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 61 | Aloe linearifolia A.Berger | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,936 | 13·2 | 26·4 | 52·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 62 | Aloe lomatophylloides Balf.f. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,248 | 17·6 | 35·2 | 70·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 63 | Aloe longistyla Baker | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,582 | 15·9 | 31·8 | 63·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 64 | Aloe macrosiphon Bak. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,934 | 18·3 | 36·6 | 73·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 65 | Aloe maculata Allionii | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 18,620 | 19·0 | 38·0 | 76·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 66a | Aloe marlothii A.Berger “Spectabilis” | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,435 | 15·8 | 31·5 | 63·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 66b | Aloe marlothii A.Berger var. bicolor Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,631 | 16·0 | 31·9 | 63·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 67b | Aloe mcloughlinii Christian | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,219 | 16·6 | 33·1 | 66·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 68 | Aloe melanacantha A.Berger | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,299 | 12·6 | 25·1 | 50·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 69 | Aloe microstigma Salm-Dyck | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,092 | 15·4 | 30·8 | 61·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 70 | Aloe mitriformis Mill. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,475 | 13·8 | 27·5 | 55·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 71b | Aloe ngobitensis Reynolds | No | Xanthorrhoeaceaek | M | 28° | 4 | P | 28,420 | 29·0 | 58·0 | 116·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 72 | Aloe occidentalis (H.Perrier) L.E.Newton & G.D.Rowley | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 20,286 | 20·7 | 41·4 | 82·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 73 | Aloe parvula A.Berger | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,562 | 16·9 | 33·8 | 67·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 74 | Aloe pearsonii Schonland | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,348 | 12·6 | 25·2 | 50·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 75b | Aloe peckii Bally & Verdoorn | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,444 | 17·8 | 35·6 | 71·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 76 | Aloe peglerae Schonland | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,729 | 16·1 | 32·1 | 64·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 77 | Aloe petricola Pole-Evans | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,092 | 15·4 | 30·8 | 61·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 78 | Aloe pillansii L.Guthrie | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 12,593 | 12·9 | 25·7 | 51·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 79 | Aloe plicatilis (L.) Mill. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 8,624 | 8·8 | 17·6 | 35·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 80 | Aloe pluridens Haworth | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,161 | 14·5 | 28·9 | 57·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 81 | Aloe polyphylla Schonland | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,377 | 13·7 | 27·3 | 54·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 82 | Aloe prinslooi Verdoorn & Hardy | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,444 | 17·8 | 35·6 | 71·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 83 | Aloe prostrata (H.Perrier) L.E.Newton & G.D.Rowley | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 20,139 | 20·6 | 41·1 | 82·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 84 | Aloe rauhii Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,337 | 15·7 | 31·3 | 62·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 85 | Aloe richardsiae Reynolds | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 21,756 | 22·2 | 44·4 | 88·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 86 | Aloe rivierei Lavranos & L.E. Newton | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,562 | 16·9 | 33·8 | 67·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 87 | Aloe secundiflora Engl. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,591 | 18·0 | 35·9 | 71·8 | 440 | O | Agave sp.ab | FC:PI | |||
| 88 | Aloe sinkatana Reynolds (red flowers)i | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 17,542 | 17·9 | 35·8 | 71·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 89 | Aloe sladeniana Pole-Evans | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 15,974 | 16·3 | 32·6 | 65·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 90 | Aloe speciosa Baker | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,112 | 14·4 | 28·8 | 57·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 91 | Aloe spicata L.f. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,259 | 14·6 | 29·1 | 58·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 92 | Aloe striata Haw. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 18,914 | 19·3 | 38·6 | 77·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 93 | Aloe suprafoliata Pole-Evans | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 14,014 | 14·3 | 28·6 | 57·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 94 | Aloe suzannae Decary | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,317 | 16·7 | 33·3 | 66·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 95b | Aloe tenuior Haw. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 10,633 | 10·9 | 21·7 | 43·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 96 | Aloe trichosantha Berger | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 18,179 | 18·6 | 37·1 | 74·2 | 440 | O | Agave sp.ab | FC:PI | |||
| 97 | Aloe vanbalenii Pillans | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 13,965 | 14·3 | 28·5 | 57·0 | 440 | O | Agave sp.ab | FC:PI | |||
| 98a | Aloe variegata L. “Ausana” | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,268 | 16·6 | 33·2 | 66·4 | 440 | O | Agave sp.ab | FC:PI | |||
| 98b | Aloe variegata L. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,562 | 16·9 | 33·8 | 67·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 99 | Aloe vera (L.) Burm.f. | No | Xanthorrhoeaceaek | M | 14° | 2 | P | 16,072 | 16·4 | 32·8 | 65·6 | 440 | O | Agave sp.ab | FC:PI | |||
| 100 | Alstonia macrophylla Wall. ex G.Don. | No | Apocynaceae | E | —n | —p | P | 718 | 0·7 | 1·5 | 2·9 | 454 | O | Bc | Fe | |||
| 101c | Alstroemeria aurea Grahamh | No | Alstroemeriaceae | M | 16 | 2 | P | 24,843 | 25·4 | 50·7 | 101·4 | 436 | O | B | FC:PI | |||
| 101d | Alstroemeria aurea Grahamh | No | Alstroemeriaceae | M | 16 | 2 | P | 27,097 | 27·7 | 55·3 | 110·6 | 436 | O | B | FC:PI | |||
| 102 | Alstroemeria aurea Grahamh | No | Alstroemeriaceae | M | 24 | 3 | P | —t | —t | 80·9 | 161·8 | 436 | O | B | FC:PI | |||
| 103d | Alstroemeria ligtu L. ssp. incarnata L.h | No | Alstroemeriaceae | M | 16 | 2 | P | 34,300 | 35·0 | 70·0 | 140·0 | 436 | O | B | FC:PI | |||
| 103e | Alstroemeria ligtu L. ssp. simsiih | No | Alstroemeriaceae | M | 16 | 2 | P | 31,948 | 32·6 | 65·2 | 130·4 | 436 | O | B | FC:PI | |||
| 103f | Alstroemeria ligtu L. ssp. simsiih | No | Alstroemeriaceae | M | 16 | 2 | P | 38,661 | 39·5 | 78·9 | 157·8 | 436 | O | B | FC:PI | |||
| 103g | Alstroemeria ligtu L. ssp. ligtuh | No | Alstroemeriaceae | M | 16 | 2 | P | 34,006 | 34·7 | 69·4 | 138·8 | 436 | O | B | FC:PI | |||
| 103h | Alstroemeria ligtu L. ssp. ligtuh | No | Alstroemeriaceae | M | 16 | 2 | P | 33,369 | 34·1 | 68·1 | 136·2 | 436 | O | B | FC:PI | |||
| 104c | Alstroemeria magnifica Herb. ssp. magnificah | No | Alstroemeriaceae | M | 16 | 2 | P | 17,885 | 18·3 | 36·5 | 73·0 | 436 | O | B | FC:PI | |||
| 104d | Alstroemeria magnifica Herb. ssp. magnificah | No | Alstroemeriaceae | M | 16 | 2 | P | 20,531 | 21·0 | 41·9 | 83·8 | 436 | O | B | FC:PI | |||
| 105 | Alstroemeria magnifica Herb. ssp. magnificah | No | Alstroemeriaceae | M | 24 | 3 | P | —t | —t | 61·6 | 123·2 | 436 | O | B | FC:PI | |||
| 106 | Amborella trichopoda Baill. | No | Amborellaceae | BA | 26° | —p | P | 870 | 0·9 | 1·8 | 3·6 | 381 | O | K | FC:PI | |||
| 107 | Amoreuxia wrightii A.Gray | No | Cochlospermaceae | E | c.12-14 | 2 | P | 164 | 0·2 | 0·3 | 0·7 | 378 | O | J | Fe | |||
| 108 | Anthemis altissimal | No | Compositaej | E | —n | —p | —q | 7,742 | 7·9 | 15·8 | 31·6 | 457bm | O | Bd | Fe | |||
| 109 | Anthemis montanal | No | Compositaej | E | —n | —p | —q | 8,281 | 8·5 | 16·9 | 33·8 | 457bm | O | Bd | Fe | |||
| 110 | Anthurium grande Host. | No | Araceae | M | 28 | —p | P | 13,252 | 13·5 | 27·0 | 54·1 | 411 | O | Bc | Fe | |||
| 111 | Anthurium tetragonum Schott | No | Araceae | M | 30 | —p | P | 7,485 | 7·6 | 15·3 | 30·6 | 411 | O | Bc | Fe | |||
| 112 | Aphyllanthes monspeliensis L. | No | Asparagaceae | M | c. 32 | —p | P | 635 | 0·6 | 1·3 | 2·6 | 380 | O | J | FC:PI | |||
| 113 | Arabidopsis korshynskyil | No | Cruciferaej | E | —n | —p | —q | 245 | 0·3 | 0·5 | 1·0 | 457bm | O | Bd | Fe | |||
| 114a | Arabidopsis thaliana (L.) Heynh. ecotype Columbia | No | Cruciferae | E | 10 | 2 | A | 157 | 0·2 | 0·3 | 0·6 | 461 | O | Caeno.f | FC:PI | |||
| 114g | Arabidopsis thaliana (L.) Heynh. ecotype Columbia | No | Cruciferae | E | 10 | 2 | A | 164 | 0·2 | 0·3 | 0·7 | 461 | O | Gallusf | FC:PI | |||
| 114h | Arabidopsis thaliana (L.) Heynh. ecotype Columbia | No | Cruciferae | E | 10 | 2 | A | 150 | 0·2 | 0·3 | 0·6 | 461 | O | Dros.f | FC:PI | |||
| 114i | Arabidopsis thaliana (L.) Heynh. | —m | Cruciferae | E | 10 | 2 | A | 125be | 0·1be | 0·3be | 0·5be | 448be | O | — | GS | |||
| 114j | Arabidopsis thaliana (L.) Heynh. ecotype Columbia | No | Cruciferae | E | 10 | 2 | A | 167 | 0·2 | 0·3 | 0·7 | 463 | O | Gallusf | FC:DAPI | |||
| 114k | Arabidopsis thaliana (L.) Heynh. ecotype Columbia | No | Cruciferae | E | 10 | 2 | A | 162 | 0·2 | 0·3 | 0·7 | 463 | O | Gallusf | FC:HO | |||
| 114l | Arabidopsis thaliana (L.) Heynh. ecotype Columbia | No | Cruciferae | E | 10 | 2 | A | 157 | 0·2 | 0·3 | 0·6 | 463 | O | Gallusf | FC:MI | |||
| 114m | Arabidopsis thaliana (L.) Heynh. line Landsberg erecta | No | Cruciferae | E | 10 | 2 | A | 51 | 0·05 | 0·10 | 0·21 | 464 | O | — | RK | |||
| 115a | Arachis duranensis Krapov. & W.C.Gregoryh | No | Leguminosaej | E | 20 | 2 | A | 1,243 | 1·3 | 2·5 | 5·1 | 396 | O | Gc | FC:PI | |||
| 115b | Arachis duranensis Krapov. & W.C.Gregoryh | No | Leguminosaej | E | 20 | 2 | A | 1,324 | 1·4 | 2·7 | 5·4 | 396 | O | Gc | FC:PI | |||
| 115c | Arachis duranensis Krapov. & W.C.Gregoryh | No | Leguminosaej | E | 20 | 2 | A | 1,333 | 1·4 | 2·7 | 5·4 | 396 | O | Gc | Fe | |||
| 116a | Arachis hypogaea L. | No | Leguminosae | E | 40 | 4 | A | 2,813 | 2·9 | 5·7 | 11·5 | 395aj | O | Gc | Fe | |||
| 116b | Arachis hypogaea L. | No | Leguminosae | E | 40 | 4 | A | 2,898 | 3·0 | 5·9 | 11·8 | 395aj | O | Gc | FC:PI | |||
| 116r | Arachis hypogaea L. | No | Leguminosaej | E | 40° | 4 | A | 1,568 | 1·6 | 3·2 | 6·4 | 457bm | O | Bd | Fe | |||
| 117a | Arachis monticola Krapov. & Rigoni | No | Leguminosae | E | 40 | 4 | A | 2,891 | 3·0 | 5·9 | 11·8 | 395aj | O | Gc | Fe | |||
| 117b | Arachis monticola Krapov. & Rigoni | No | Leguminosae | E | 40 | 4 | A | 2,930 | 3·0 | 6·0 | 12·0 | 395aj | O | Gc | FC:PI | |||
| 118 | Archidendron monadelphum (Roxb.) I.C. Neilsen | No | Leguminosae | E | —n | —p | P | 1,470 | 1·5 | 3·0 | 6·0 | 454 | O | Bc | Fe | |||
| 119c | Artemisia absinthium L. | No | Compositaej | E | 18 | 2 | P | 4,175 | 4·3 | 8·5 | 17·0 | 386 | O | G-120d | FC:PI | |||
| 120d | Artemisia annua L. | No | Compositaej | E | 18 | 2 | A | 1,715 | 1·8 | 3·5 | 7·0 | 386 | O | Gc | FC:PI | |||
| 121 | Artemisia barrelieri Besser | No | Compositaej | E | 36 | 4 | P | 6,350 | 6·5 | 13·0 | 25·9 | 386 | O | Gc | FC:PI | |||
| 122 | Artemisia caerulescens L. ssp. gallica (Willd.) K.Persson | No | Compositaej | E | 18 | 2 | P | 3,263 | 3·3 | 6·7 | 13·3 | 386 | O | Gc | FC:PI | |||
| 123 | Artemisia campestris L. | No | Compositaej | E | 18 | 2 | P | 2,876 | 2·9 | 5·9 | 11·7 | 386 | O | Gc | FC:PI | |||
| 124 | Artemisia campestris L. | No | Compositaej | E | 36 | 4 | P | 5,390 | 5·5 | 11·0 | 22·0 | 386 | O | Gc | FC:PI | |||
| 125 | Artemisia cana Pursh. | No | Compositaej | E | 72 | 8 | P | 12,569 | 12·8 | 25·7 | 51·3 | 386 | O | Gc | FC:PI | |||
| 126 | Artemisia chamaemelifolia Vill. | No | Compositaej | E | 18 | 2 | P | 2,960 | 3·0 | 6·0 | 12·1 | 386 | O | Gc | FC:PI | |||
| 127 | Artemisia crithmifolia L. | No | Compositaej | E | 54 | 6 | P | 7,644 | 7·8 | 15·6 | 31·2 | 386 | O | Gc | FC:PI | |||
| 128 | Artemisia dracunculus L. | No | Compositaej | E | 90 | 10 | P | 11,378 | 11·6 | 23·2 | 46·4 | 386 | O | Gc | FC:PI | |||
| 129 | Artemisia fragrans Willd. | No | Compositaej | E | 18 | 2 | P | 2,622 | 2·7 | 5·4 | 10·7 | 386 | O | Gc | FC:PI | |||
| 130 | Artemisia herba-alba Asso ssp. valentina (Lam.) Mascl. | No | Compositaej | E | 18 | 2 | P | 3,219 | 3·3 | 6·6 | 13·1 | 386 | O | Gc | FC:PI | |||
| 131 | Artemisia herba-alba Asso ssp. herba-alba | No | Compositaej | E | 36 | 4 | P | 6,115 | 6·2 | 12·5 | 25·0 | 386 | O | Gc | FC:PI | |||
| 132c | Artemisia judaica L. | No | Compositaej | E | 16 | 2 | P | 5,645 | 5·8 | 11·5 | 23·0 | 386 | O | Gc | FC:PI | |||
| 133 | Artemisia lucentica O.Bolos, Valles & Vigo in O.Bolos & Vigo | No | Compositaej | E | 16 | 2 | P | 3,763 | 3·8 | 7·7 | 15·4 | 386 | O | G-120d | FC:PI | |||
| 134 | Artemisia molinieri Quezel, Barbero & R.Loisel | No | Compositaej | E | 18 | 2 | P | 2,920 | 3·0 | 6·0 | 11·9 | 386 | O | Gc | FC:PI | |||
| 135 | Artemisia monosperma Delile | No | Compositaej | E | 36 | 4 | P | 5,400 | 5·5 | 11·0 | 22·0 | 386 | O | Gc | FC:PI | |||
| 136 | Artemisia splendens Willd. | No | Compositaej | E | 32 | 4 | P | 6,659 | 6·8 | 13·6 | 27·2 | 386 | O | Gc | FC:PI | |||
| 137 | Artemisia thuscula Cav. | No | Compositaej | E | 18 | 2 | P | 5,155 | 5·3 | 10·5 | 21·0 | 386 | O | Gc | FC:PI | |||
| 138 | Artemisia tournefortiana Reichenb. | No | Compositaej | E | 18 | 2 | AB | 3,278 | 3·3 | 6·7 | 13·4 | 386 | O | Gc | FC:PI | |||
| 139 | Artemisia tridentata Nutt. ssp. spiciformis Kartesz & Gandhi | No | Compositaej | E | 18 | 2 | P | 4,008 | 4·1 | 8·2 | 16·4 | 386 | O | G-120d | FC:PI | |||
| 140 | Artemisia umbelliformis Lam. ssp. umbelliformis | No | Compositaej | E | 34 | 4 | P | 6,081 | 6·2 | 12·4 | 24·8 | 386 | O | Gc | FC:PI | |||
| 141b | Artemisia vulgaris L. | No | Compositaej | E | 16 | 2 | P | 2,979 | 3·0 | 6·1 | 12·2 | 386 | O | Gc | FC:PI | |||
| 142 | Artemisia vulgaris L. | No | Compositaej | E | 34 | 4 | P | 4,773 | 4·9 | 9·7 | 19·5 | 386 | O | Gc | FC:PI | |||
| 143b | Arum maculatum L. | No | Araceae | M | 56° | 8 | P | 10,682 | 10·9 | 21·8 | 43·6 | 457bm | O | Bd | Fe | |||
| 144 | Asarum europaeum L. | No | Aristolochiaceae | BA | —n | —p | P | 4,753 | 4·9 | 9·7 | 19·4 | 457bm | O | Bd | Fe | |||
| 145 | Astelia fragrans Colenso | No | Asteliaceae | M | c. 60 | 8 | P | 1,240 | 1·3 | 2·5 | 5·1 | 380 | O | K | FC:PI | |||
| 146 | Atalantia ceylanica (Arn.) Oliv.) | No | Rutaceae | E | 18° | 2 | P | 515 | 0·5 | 1·1 | 2·1 | 426 | O | Gallusf | FC:PI | |||
| 147 | Austrobaileya scandens C.T.White | No | Austrobaileyaceae | BA | 44° | —p | P | 9,327 | 9·5 | 19·0 | 38·1 | 381 | O | G | FC:PI | |||
| 148 | Averrhoa carambola L. | No | Oxalidaceae | E | —n | —p | P | 235 | 0·2 | 0·5 | 1·0 | 454 | O | Bc | Fe | |||
| 149 | Azadirachta indica A.Juss | No | Meliaceae | E | 28° | —p | P | 385 | 0·4 | 0·8 | 1·6 | 454 | O | Bc | Fe | |||
| 150 | Bauhinia hookeri (F.Muell.) Pedley | No | Leguminosae | E | 26° | —p | P | 620 | 0·6 | 1·3 | 2·5 | 454 | O | Bc | Fe | |||
| 151b | Bauhinia purpurea L. | No | Leguminosae | E | 28° | 2 | P | 573 | 0·6 | 1·2 | 2·3 | 454 | O | Bc | Fe | |||
| 152 | Bauhinia tomentosa L. | No | Leguminosae | E | 28° | 2 | P | 613 | 0·6 | 1·3 | 2·5 | 454 | O | Bc | Fe | |||
| 153 | Bellevalia rixii P.Wendalbo | No | Asparagaceae | M | 8 | 2 | P | 9,102 | 9·3 | 18·6 | 37·2 | 465 | O | B | Fe | |||
| 154 | Berberidopsis corallina Hook. f. | No | Berberidopsidaceae | E | c. 42 | 6 | P | 252 | 0·3 | 0·5 | 1·0 | 379 | O | J | Fe | |||
| 155 | Berrya cordifolia (Willd.) Burret | No | Malvaceae | E | —n | —p | P | 549 | 0·6 | 1·1 | 2·2 | 454 | O | Bc | Fe | |||
| 156a | Bixa orellana L. | No | Bixaceae | E | 14 | 2 | P | 191 | 0·2 | 0·4 | 0·8 | 379 | O | J | Fe | |||
| 156b | Bixa orellana L. | No | Bixaceae | E | 14° | 2 | P | 203 | 0·2 | 0·4 | 0·8 | 454 | O | Bc | Fe | |||
| 157 | Blandfordia punicea Sweet. | No | Blandfordiaceae | M | 68 | 4 | P | 7,970 | 8·1 | 16·3 | 32·5 | 380 | O | G | Fe | |||
| 158 | Bombax ceiba L. | No | Malvaceae | E | 92° | —p | P | 1,590 | 1·6 | 3·2 | 6·5 | 454 | O | Bc | Fe | |||
| 159 | Boswellia serrata Roxb. | No | Burseraceae | E | 22° | —p | P | 684 | 0·7 | 1·4 | 2·8 | 454 | O | Bc | Fe | |||
| 160 | Brachychiton discolor F.Muell | No | Malvaceae | E | 40° | —p | P | 1,132 | 1·2 | 2·3 | 4·6 | 454 | O | Bc | Fe | |||
| 161a | Brachypodium distachyon (L.) P.Beauv. | No | Gramineae | M | 10 | 2 | A | 355 | 0·4 | 0·7 | 1·5 | 465 | O | K | FC:PI | |||
| 161b | Brachypodium distachyon (L.) P.Beauv. | —m | Gramineae | M | 10 | 2 | A | 294 | 0·3 | 0·6 | 1·2 | 460bh | O | J | Fe | |||
| 162e | Brassica napus L. | No | Cruciferaej | E | 38° | 4 | AB | 1,568 | 1·6 | 3·2 | 6·4 | 457bm | O | Bd | Fe | |||
| 163b | Bromus arvensis L. | No | Gramineae | M | 14 | 2 | A | 5,699 | 5·8 | 11·6 | 23·3 | 391af | O | H-164b | FC:PI | |||
| 164b | Bromus carinatus Hooker & Arnott cv. Broma | No | Gramineae | M | 56 | 8 | P | 11,241 | 11·5 | 22·9 | 45·9 | 391af | O | Hc | FC:PI | |||
| 165c | Bromus erectus Hudson | No | Gramineae | M | 56 | 8 | P | 12,079 | 12·3 | 24·7 | 49·3 | 391af | O | Hc | FC:PI | |||
| 166c | Bromus hordeaceus L. | No | Gramineae | M | 28 | 4 | A | 11,285 | 11·5 | 23·0 | 46·1 | 391af | O | Hc | FC:PI | |||
| 167b | Bromus inermis Leysser | No | Gramineae | M | 56 | 8 | P | 12,025 | 12·3 | 24·5 | 49·1 | 391af | O | Hc | FC:PI | |||
| 168 | Bromus willdenowii Knuth | No | Gramineae | M | 42 | 6 | P | 6,365 | 6·5 | 13·0 | 26·0 | 391af | O | Hc | FC:PI | |||
| 169 | Buchloe dactyloides (Nutt.) Engelm. | No | Gramineae | M | 40 | 4 | P | 779 | 0·8 | 1·6 | 3·2 | 417 | O | Gallusf | FC:PI | |||
| 170 | Buddleja globosa Hope | No | Buddlejaceae | E | 38 | 2 | P | 840 | 0·9 | 1·7 | 3·4 | 378 | O | J | Fe | |||
| 171 | Bulbine alooides Willd. | No | Xanthorrhoeaceae | M | 14 | 2 | P | 10,601 | 10·8 | 21·6 | 43·3 | 465 | O | B | Fe | |||
| 172 | Bulbine fallax Poelln. | No | Xanthorrhoeaceae | M | 14 | 2 | P | 11,201 | 11·4 | 22·9 | 45·7 | 465 | O | B | Fe | |||
| 173 | Bulbine lagopus (Thunb.) N.E.Brown | No | Xanthorrhoeaceae | M | —n | —p | P | 7,938 | 8·1 | 16·2 | 32·4 | 465 | O | B | Fe | |||
| 174 | Bulbine praemorsa Spreng. | No | Xanthorrhoeaceae | M | 14 | 2 | P | 12,213 | 12·5 | 24·9 | 49·9 | 465 | O | B | Fe | |||
| 175a | Bunias erucago L. | No | Cruciferae | E | 14 | 2 | A | 2,029 | 2·1 | 4·1 | 8·3 | 393 | O | Gc | Fe | |||
| 175b | Bunias erucago L. | No | Cruciferae | E | 14 | 2 | A | 2,136 | 2·2 | 4·4 | 8·7 | 393 | O | Glycinee | FC:PI | |||
| 176a | Bunias orientalis L. | No | Cruciferae | E | 14 | 2 | P | 2,538 | 2·6 | 5·2 | 10·4 | 393 | O | Gc | Fe | |||
| 176b | Bunias orientalis L. | No | Cruciferae | E | 14 | 2 | P | 2,636 | 2·7 | 5·4 | 10·8 | 393 | O | Glycinee | FC:PI | |||
| 177 | Buxus papillosa C.K. Schneid. | No | Buxaceae | E | —n | —p | P | 1,389 | 1·4 | 2·8 | 5·7 | 454 | O | Bc | Fe | |||
| 178 | Buxus sempervirensl | No | Buxaceae | E | 28 | 2 or 4 | P | 794 | 0·8 | 1·6 | 3·2 | 380 | O | K | FC:PI | |||
| 179 | Byblis liniflora Salisb. | No | Byblidaceae | E | 32 | 2 | A | 870 | 0·9 | 1·8 | 3·6 | 378 | O | J | Fe | |||
| 180 | Cajanus albicans (Wight. & Am.) Maesen | No | Leguminosae | E | 22 | 2 | A | 1,259 | 1·3 | 2·6 | 5·1 | 443bc | O | Bc | Fe | |||
| 181 | Cajanus mollis (Benth.) Maesen | No | Leguminosae | E | 22 | 2 | B | 804 | 0·8 | 1·6 | 3·3 | 443bc | O | Bc | Fe | |||
| 182 | Cajanus sericeus (Benth. ex Bak.) Maesen | No | Leguminosae | E | 22 | 2 | P | 1,414 | 1·4 | 2·9 | 5·8 | 443bc | O | Bc | Fe | |||
| 183 | Caladium bicolor Vent. var. red polka | No | Araceae | M | 32 | —p | A | 5,407 | 5·5 | 11·0 | 22·1 | 411 | O | Bc | Fe | |||
| 184 | Caladium bicolor Vent. var. red polka large | No | Araceae | M | 66 | —p | A | 9,927 | 10·1 | 20·3 | 40·5 | 411 | O | Bc | Fe | |||
| 185 | Calceolaria acutifolia Witasek | No | Scrophulariaceae | E | —n | —p | —q | 1,348 | 1·4 | 2·8 | 5·5 | 465 | O | J | Fe | |||
| 186 | Calceolaria gracilisl | No | Scrophulariaceae | E | —n | —p | —q | 1,335 | 1·4 | 2·7 | 5·5 | 465 | O | J | Fe | |||
| 187 | Calibrachoa calycina (Sendtn.) Wijsman | No | Solanaceae | E | 18 | 2 | P | 1,504 | 1·5 | 3·1 | 6·1 | 387ae | O | Gallus-398p | FC:PI | |||
| 188 | Calibrachoa dusenii (R.E.Fr.) Stehmann & Semir. | No | Solanaceae | E | 18 | 2 | P | 1,401 | 1·4 | 2·9 | 5·7 | 387ae | O | Gallus-398p | FC:PI | |||
| 189 | Calibrachoa eglandulata Stehmann & Semir. | No | Solanaceae | E | 18° | 2 | P | 1,411 | 1·4 | 2·9 | 5·8 | 387ae | O | Gallus-398p | FC:PI | |||
| 190 | Calibrachoa elegans (Miers) Stehmann & Semir. | No | Solanaceae | E | 18 | 2 | P | 1,563 | 1·6 | 3·2 | 6·4 | 387ae | O | Gallus-398p | FC:PI | |||
| 191 | Calibrachoa ericaefolia (R.E.Fr.) Wijsman | No | Solanaceae | E | 18 | 2 | P | 1,436 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 192 | Calibrachoa heterophylla (Sendtn.) Wijsman | No | Solanaceae | E | 18° | 2 | P | 1,455 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 193 | Calibrachoa linearis (Hook.) Wijsman | No | Solanaceae | E | 18 | 2 | P | 1,485 | 1·5 | 3·0 | 6·1 | 387ae | O | Gallus-398p | FC:PI | |||
| 194 | Calibrachoa linoides (Sendtn.) Wijsman | No | Solanaceae | E | 18 | 2 | P | 1,397 | 1·4 | 2·9 | 5·7 | 387ae | O | Gallus-398p | FC:PI | |||
| 195 | Calibrachoa macrodactylon (L.B.Sm. & Downs) Wijsman | No | Solanaceae | E | 18 | 2 | P | 1,480 | 1·5 | 3·0 | 6·0 | 387ae | O | Gallus-398p | FC:PI | |||
| 196 | Calibrachoa micrantha (R.E.Fr.) Stehmann & Semir. | No | Solanaceae | E | 18 | 2 | P | 1,411 | 1·4 | 2·9 | 5·8 | 387ae | O | Gallus-398p | FC:PI | |||
| 197 | Calibrachoa parviflora (Juss.) Wijsman | No | Solanaceae | E | 18 | 2 | A | 936 | 1·0 | 1·9 | 3·8 | 387ae | O | Gallus-398p | FC:PI | |||
| 198 | Calibrachoa pygmaea (R.E.Fr.) Wijsman | No | Solanaceae | E | 18 | 2 | A | 764 | 0·8 | 1·6 | 3·1 | 387ae | O | Gallus-398p | FC:PI | |||
| 199 | Calibrachoa rupestris (Dusen) Wijsman | No | Solanaceae | E | 18° | 2 | P | 1,597 | 1·6 | 3·3 | 6·5 | 387ae | O | Gallus-398p | FC:PI | |||
| 200 | Calibrachoa selloviana (Sendtn.) Wijsman | No | Solanaceae | E | 18 | 2 | P | 1,455 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 201 | Calibrachoa sendtneriana (R.E.Fr.) Stehmann & Semir. | No | Solanaceae | E | 18° | 2 | P | 1,450 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 202 | Calibrachoa serrulata (L.B.Sm. & Downs) Stehmann & Semir. | No | Solanaceae | E | 18 | 2 | P | 1,446 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 203 | Calibrachoa spathulata (L.B.Sm. & Downs) Stehmann & Semir. | No | Solanaceae | E | 18 | 2 | P | 1,416 | 1·4 | 2·9 | 5·8 | 387ae | O | Gallus-398p | FC:PI | |||
| 204 | Calibrachoa thymifolia (A. St.-Hil.) Stehmann & Semir. | No | Solanaceae | E | 18 | 2 | P | 1,485 | 1·5 | 3·0 | 6·1 | 387ae | O | Gallus-398p | FC:PI | |||
| 205 | Callistemon citrinus (Curtis) Skeels | No | Myrtaceae | E | 22° | —p | P | 1,014 | 1·0 | 2·1 | 4·1 | 454 | O | Bc | Fe | |||
| 206 | Callistemon rigidus R.Br. | No | Myrtaceae | E | —n | —p | P | 1,526 | 1·6 | 3·1 | 6·2 | 454 | O | Bc | Fe | |||
| 207 | Camellia sinensis Kuntze | No | Theaceae | E | 30 | 2 | P | 3,824 | 3·9 | 7·8 | 15·6 | 379 | O | G | Fe | |||
| 208 | Canna indical | No | Cannaceae | M | 18 | 2 | P | 706 | 0·7 | 1·4 | 2·9 | 379 | O | J | Fe | |||
| 209a | Cannabis sativa L. (female) | No | Cannabaceae | E | 20 | 2 | A | 818.am | 0·8am | 1·7am | 3·3am | 414 | O | Arab.e | FC:DAPI | |||
| 209b | Cannabis sativa L. (male) | No | Cannabaceae | E | 20 | 2 | A | 843.am | 0·9am | 1·7am | 3·4am | 414 | O | Arab.e | FC:DAPI | |||
| 210 | Canotia holacantha Torr. | No | Celastraceae | E | 30 | 2 | P | 181 | 0·2 | 0·4 | 0·7 | 378 | O | J | Fe | |||
| 211b | Capsella bursa-pastoris (L.) Medic. | No | Cruciferaej | E | 32° | 4 | A | 686 | 0·7 | 1·4 | 2·8 | 457bm | O | Bd | Fe | |||
| 212h | Capsicum annuum L. cv. Doux Long des Landes | No | Solanaceae | E | 24 | 2 | —q | 3,734 | 3·8 | 7·6 | 15·2 | 434 | O | Gc | FC:PI | |||
| 213c | Capsicum baccatum L. ssp. pendulum | No | Solanaceae | E | 24 | 2 | —q | 4,111 | 4·2 | 8·4 | 16·8 | 434ay | O | G-212h | FC:PI | |||
| 213d | Capsicum baccatum L. ssp. baccatum | No | Solanaceae | E | 24 | 2 | —q | 4,131 | 4·2 | 8·4 | 16·9 | 434ay | O | G-212h | FC:PI | |||
| 214 | Capsicum cardenasii Heiser & Smith | No | Solanaceae | E | 24° | 2 | —q | 4,395 | 4·5 | 9·0 | 17·9 | 434 | O | G-212h | FC:PI | |||
| 215 | Capsicum chacoense A.T.Hunz. | No | Solanaceae | E | 24° | 2 | —q | 3,753 | 3·8 | 7·7 | 15·3 | 434 | O | G-212h | FC:PI | |||
| 216b | Capsicum chinense Jacq. | No | Solanaceae | E | 24 | 2 | —q | 3,940 | 4·0 | 8·0 | 16·1 | 434ay | O | G-212h | FC:PI | |||
| 217b | Capsicum eximium A.T.Hunz. | No | Solanaceae | E | 24° | 2 | —q | 4,263 | 4·4 | 8·7 | 17·4 | 434ay | O | G-212h | FC:PI | |||
| 218b | Capsicum frutescens L. | No | Solanaceae | E | 24° | 2 | —q | 3,891 | 4·0 | 7·9 | 15·9 | 434ay | O | G-212h | FC:PI | |||
| 219 | Capsicum praetermissum Heiser & Smith | No | Solanaceae | E | 24° | 2 | —q | 4,474 | 4·6 | 9·1 | 18·3 | 434 | O | G-212h | FC:PI | |||
| 220b | Capsicum pubescens R. & P. | No | Solanaceae | E | 24° | 2 | —q | 4,763 | 4·9 | 9·7 | 19·4 | 434ay | O | G-212h | FC:PI | |||
| 221 | Capsicum tovarii Eshbaugh, Smith & Nickrent | No | Solanaceae | E | 24° | 2 | —q | 3,886 | 4·0 | 7·9 | 15·9 | 434 | O | G-212h | FC:PI | |||
| 222a | Cardamine amara L. | No | Cruciferae | E | 16 | 2 | P | 238 | 0·2 | 0·5 | 1·0 | 465 | O | Lycopers.c | FC:PI | |||
| 223 | Castanospermum australe A.Cunn. & C.Fraser | No | Leguminosae | E | —n | —p | P | 554 | 0·6 | 1·1 | 2·3 | 454 | O | Bc | Fe | |||
| 224 | Casuarina glauca Sieb. ex Spring. | No | Casuarinaceae | E | 18° | 2 | P | 343 | 0·4 | 0·7 | 1·4 | 452 | O | Petuniae | FC:PI | |||
| 225 | Catunaregam spinosa (Thunb.) Trivengadum | No | Rubiaceae | E | 22° | —p | P | 343 | 0·4 | 0·7 | 1·4 | 454 | O | Bc | Fe | |||
| 226b | Centaurea scabiosal | No | Compositae | E | —n | —p | P | 1,254 | 1·3 | 2·6 | 5·1 | 465 | O | J | Fe | |||
| 227 | Cephalotus follicularis Labill. | No | Cephalotaceae | E | 20° | 2 | P | 625 | 0·6 | 1·3 | 2·6 | 378 | O | J | Fe | |||
| 228a | Cerastium alpinum L. | No | Caryophyllaceae | E | 72 | 4 | P | 1,813 | 1·9 | 3·7 | 7·4 | 427 | O | Cerastiume | FC:DAPI | |||
| 228b | Cerastium alpinum L. | No | Caryophyllaceae | E | 72 | 4 | P | 1,970 | 2·0 | 4·0 | 8·0 | 427 | O | Bc | Fe | |||
| 229 | Cerastium arcticum Lange s. str. | No | Caryophyllaceae | E | 108 | 6 | P | 3,126 | 3·2 | 6·4 | 12·8 | 427 | O | Cerastiume | FC:DAPI | |||
| 230 | Cerastium arvense L. ssp. glandulosum (Kit.) Soo | No | Caryophyllaceae | E | 36 | 2au | P | 666 | 0·7 | 1·4 | 2·7 | 427 | O | Cerastiume | FC:DAPI | |||
| 231 | Cerastium arvense L. ssp. arvense | No | Caryophyllaceae | E | 72 | 4 | P | 1,274 | 1·3 | 2·6 | 5·2 | 427 | O | Cerastiume | FC:DAPI | |||
| 232a | Cerastium banaticum (Rochel) Heuff. | No | Caryophyllaceae | E | 36 | 2au | P | 1,470 | 1·5 | 3·0 | 6·0 | 427 | O | Bc | Fe | |||
| 232b | Cerastium banaticum (Rochel) Heuff. | No | Caryophyllaceae | E | 36 | 2au | P | 1,529 | 1·6 | 3·1 | 6·2 | 427 | O | Cerastiume | FC:DAPI | |||
| 233 | Cerastium carinthiacum Vest | No | Caryophyllaceae | E | 36 | 2au | P | 1,480 | 1·5 | 3·0 | 6·0 | 427 | O | Cerastiume | FC:DAPI | |||
| 234a | Cerastium eriophorum Kit. in Schult. | No | Caryophyllaceae | E | 36 | 2au | P | 1,264 | 1·3 | 2·6 | 5·2 | 427 | O | Bc | Fe | |||
| 234b | Cerastium eriophorum Kit. in Schult. | No | Caryophyllaceae | E | 36 | 2au | P | 1,274 | 1·3 | 2·6 | 5·2 | 427 | O | Nicot.e | FC:DAPI | |||
| 235b | Cerastium fontanum Baumg. | No | Caryophyllaceae | E | 144 | 8 | P | 3,469 | 3·5 | 7·1 | 14·2 | 427 | O | Cerastiume | FC:DAPI | |||
| 236a | Cerastium latifolium L. | No | Caryophyllaceae | E | 36 | 2au | P | 1,421 | 1·5 | 2·9 | 5·8 | 427 | O | Bc | Fe | |||
| 236b | Cerastium latifolium L. | No | Caryophyllaceae | E | 36 | 2au | P | 1,470 | 1·5 | 3·0 | 6·0 | 427 | O | Cerastiume | FC:DAPI | |||
| 237a | Cerastium transsylvanicum Schur ex Griseb. & Schenk | No | Caryophyllaceae | E | 108 | 6 | P | 3,038 | 3·1 | 6·2 | 12·4 | 427 | O | Cerastiume | FC:DAPI | |||
| 237b | Cerastium transsylvanicum Schur ex Griseb. & Schenk | No | Caryophyllaceae | E | 108 | 6 | P | 3,048 | 3·1 | 6·2 | 12·4 | 427 | O | Bc | Fe | |||
| 238 | Ceratophyllum demersum L. | No | Ceratophyllaceae | BA | c.70 | 6 | P | 674 | 0·7 | 1·4 | 2·8 | 381 | O | J | Fe | |||
| 239 | Chenopodium album L.h | No | Amaranthaceae | E | 18° | 2 | A | 750 | 0·8 | 1·5 | 3·1 | 455 | O | Bc | Fe | |||
| 240 | Chenopodium album L.h | No | Amaranthaceae | E | 36° | 4 | A | 1,597 | 1·6 | 3·3 | 6·5 | 455 | O | Bc | Fe | |||
| 241b | Chenopodium album L.h | No | Amaranthaceae | E | 54° | 6 | A | 2,423 | 2·5 | 4·9 | 9·9 | 455 | O | Bc | Fe | |||
| 242 | Chenopodium berlandieri (Saff.) Wilson & Heiser ssp. nuttalliaeh | No | Amaranthaceae | E | 36° | 4 | A | 1,446 | 1·5 | 3·0 | 5·9 | 455 | O | Bc | Fe | |||
| 243 | Chenopodium bushianum Aellen | No | Amaranthaceae | E | 36° | 4 | A | 1,558 | 1·6 | 3·2 | 6·4 | 455 | O | Bc | Fe | |||
| 244 | Chenopodium ficifolium Sm. | No | Amaranthaceae | E | 18° | 2 | A | 649 | 0·7 | 1·3 | 2·7 | 455 | O | Bc | Fe | |||
| 245 | Chenopodium giganteum D.Don | No | Amaranthaceae | E | 54° | 6 | A | 2,151 | 2·2 | 4·4 | 8·8 | 455 | O | Bc | Fe | |||
| 246 | Chenopodium murale L. | No | Amaranthaceae | E | 18° | 2 | A | 610 | 0·6 | 1·2 | 2·5 | 455 | O | Bc | Fe | |||
| 247 | Chenopodium opulifolium Schrad. ex Koch & Ziz | No | Amaranthaceae | E | 36° | 4 | A | 1,303 | 1·3 | 2·7 | 5·3 | 455 | O | Bc | Fe | |||
| 248b | Chenopodium pallidicaule Aellen | No | Amaranthaceae | E | 18° | 2 | A | 617 | 0·6 | 1·3 | 2·5 | 455 | O | Bc | Fe | |||
| 249b | Chenopodium quinoa Willd.h | No | Amaranthaceae | E | 36° | 4 | A | 1,585 | 1·6 | 3·2 | 6·5 | 455 | O | Bc | Fe | |||
| 250 | Chenopodium ugandae (Aell.) Aell. | No | Amaranthaceae | E | 32° | —p | A | 1,401 | 1·4 | 2·9 | 5·7 | 455 | O | Bc | Fe | |||
| 251 | Chenopodium vulvaria L. | No | Amaranthaceae | E | 18° | 2 | A | 622 | 0·6 | 1·3 | 2·5 | 455 | O | Bc | Fe | |||
| 252 | Chloranthus spicatus Mak. | No | Chloranthaceae | BA | 30 | —p | P | 3,526 | 3·6 | 7·2 | 14·4 | 381 | O | G | FC:PI | |||
| 253 | Chorisia speciosa St. Hill | No | Malvaceae | E | 86° | —p | P | 845 | 0·9 | 1·7 | 3·5 | 454 | O | Bc | Fe | |||
| 254 | Cicca acida (L.) Merr. | No | Euphorbiaceae | E | —n | —p | P | 938 | 1·0 | 1·9 | 3·8 | 454 | O | Bc | Fe | |||
| 255 | Cicer songaricum Steph. ex DC. | No | Leguminosae | E | 16 | 2 | P | 1,328 | 1·4 | 2·7 | 5·4 | 435 | O | Bc | Fe | |||
| 256 | Cienfuegosia tripartita H.B.K. Gurke | —m | Malvaceae | E | 20 | 2 | —q | 931 | 1·0 | 1·9 | 3·8 | 444bd | O | Gb2 | FC:PI | |||
| 257 | Cienfuegosia yucatanensis Millspaugh | —m | Malvaceae | E | 22 | 2 | —q | 980 | 1·0 | 2·0 | 4·0 | 444bd | O | Gb2 | FC:PI | |||
| 258 | Cistus albanicus E.F.Warburg ex Heywood | No | Cistaceae | E | 18° | 2 | P | 2,617 | 2·7 | 5·3 | 10·7 | 404 | O | Raphanuse | FC:PI | |||
| 259 | Cistus albidus L. | No | Cistaceae | E | 18° | 2 | P | 2,342 | 2·4 | 4·8 | 9·6 | 404 | O | Raphanuse | FC:PI | |||
| 260 | Cistus clusii Dunal | No | Cistaceae | E | 18° | 2 | P | 2,587 | 2·6 | 5·3 | 10·6 | 404 | O | Raphanuse | FC:PI | |||
| 261 | Cistus creticus L. | No | Cistaceae | E | 18° | 2 | P | 2,127 | 2·2 | 4·3 | 8·7 | 404 | O | Raphanuse | FC:PI | |||
| 262 | Cistus crispus L. | No | Cistaceae | E | 18° | 2 | P | 1,921 | 2·0 | 3·9 | 7·8 | 404 | O | Raphanuse | FC:PI | |||
| 263 | Cistus heterophyllus Desf. ssp. carthaginensis (Pau) Crespo & Mateo | No | Cistaceae | E | 18° | 2 | P | 2,362 | 2·4 | 4·8 | 9·6 | 404 | O | Raphanuse | FC:PI | |||
| 264 | Cistus ladanifer L. | No | Cistaceae | E | 18° | 2 | P | 2,181 | 2·2 | 4·5 | 8·9 | 404 | O | Raphanuse | FC:PI | |||
| 265 | Cistus laurifolius L. | No | Cistaceae | E | 18° | 2 | P | 2,185 | 2·2 | 4·5 | 8·9 | 404 | O | Raphanuse | FC:PI | |||
| 266 | Cistus libanotis L. | No | Cistaceae | E | 18° | 2 | P | 2,827 | 2·9 | 5·8 | 11·5 | 404 | O | Raphanuse | FC:PI | |||
| 267 | Cistus monspeliensis L. | No | Cistaceae | E | 18° | 2 | P | 2,881 | 2·9 | 5·9 | 11·8 | 404 | O | Raphanuse | FC:PI | |||
| 268 | Cistus osbeckiaefolius Webb ex Pitard & Proust | No | Cistaceae | E | 18° | 2 | P | 2,024 | 2·1 | 4·1 | 8·3 | 404 | O | Raphanuse | FC:PI | |||
| 269 | Cistus parviflorus Lam. | No | Cistaceae | E | 18° | 2 | P | 2,430 | 2·5 | 5·0 | 9·9 | 404 | O | Raphanuse | FC:PI | |||
| 270 | Cistus populifolius L. | No | Cistaceae | E | 18° | 2 | P | 2,102 | 2·1 | 4·3 | 8·6 | 404 | O | Raphanuse | FC:PI | |||
| 271 | Cistus psilosepalus Sweet | No | Cistaceae | E | 18° | 2 | P | 2,558 | 2·6 | 5·2 | 10·4 | 404 | O | Raphanuse | FC:PI | |||
| 272 | Cistus salviifolius L. | No | Cistaceae | E | 18° | 2 | P | 2,332 | 2·4 | 4·8 | 9·5 | 404 | O | Raphanuse | FC:PI | |||
| 273 | Cistus symphytifolius Lam. | No | Cistaceae | E | 18° | 2 | P | 2,406 | 2·5 | 4·9 | 9·8 | 404 | O | Raphanuse | FC:PI | |||
| 274b | Citrus aurantium L. | No | Rutaceae | E | 18° | 2 | P | 431 | 0·4 | 0·9 | 1·8 | 426 | O | Gallusf | FC:PI | |||
| 275b | Citrus grandis (L.) Osbeck | No | Rutaceae | E | 18° | 2 | P | 377 | 0·4 | 0·8 | 1·5 | 426 | O | Gallusf | FC:PI | |||
| 276b | Citrus limon (L.) Burm. f.i | No | Rutaceae | E | 18° | 2 | P | 392 | 0·4 | 0·8 | 1·6 | 426 | O | Gallusf | FC:PI | |||
| 277 | Citrus limonia Osbeck cv. Brome Rangpur | No | Rutaceae | E | 18° | 2 | P | 402 | 0·4 | 0·8 | 1·6 | 426 | O | Gallusf | FC:PI | |||
| 278b | Citrus paradisi Macfad. | No | Rutaceae | E | 18° | 2 | P | 392 | 0·4 | 0·8 | 1·6 | 426 | O | Gallusf | FC:PI | |||
| 279 | Citrus reshni Hort. ex Tanaka | No | Rutaceae | E | 18° | 2 | P | 402 | 0·4 | 0·8 | 1·6 | 426 | O | Gallusf | FC:PI | |||
| 280e | Citrus sinensis (L.) Osbeck cv. Sargoins Grosse Rondeh | No | Rutaceae | E | 18° | 2 | P | 372 | 0·4 | 0·8 | 1·5 | 426 | O | Gallusf | FC:PI | |||
| 280f | Citrus sinensis (L.) Osbeck cv. Pineappleh | No | Rutaceae | E | 18° | 2 | P | 417 | 0·4 | 0·9 | 1·7 | 426 | O | Gallusf | FC:PI | |||
| 280g | Citrus sinensis (L.) Osbeck | No | Rutaceae | E | 18° | 2 | P | 588 | 0·6 | 1·2 | 2·4 | 457bm | O | Bd | Fe | |||
| 281 | Citrus volkameriana Ten. & Pasq. | No | Rutaceae | E | 18° | 2 | P | 387 | 0·4 | 0·8 | 1·6 | 426 | O | Gallusf | FC:PI | |||
| 282 | Coccoloba diversifolia Jacq. | No | Polygonaceae | E | —n | —p | P | 1,127 | 1·2 | 2·3 | 4·6 | 454 | O | Bc | Fe | |||
| 283h | Coffea arabica L. | —m | Rubiaceae | E | 44 | 4 | P | 1,279 | 1·3 | 2·6 | 5·2 | 424 | O | Kc | FC:PI | |||
| 283i | Coffea arabica L. | No | Rubiaceae | E | 44° | 4 | P | 1,122 | 1·1 | 2·3 | 4·6 | 454 | O | Bc | Fe | |||
| 284c | Coffea brevipes Hiern. | —m | Rubiaceae | E | 22 | 2 | P | 760 | 0·8 | 1·6 | 3·1 | 424 | O | Kc | FC:PI | |||
| 285d | Coffea canephora Pierre. ex Froehn. | —m | Rubiaceae | E | 22 | 2 | P | 755 | 0·8 | 1·5 | 3·1 | 424 | O | Kc | FC:PI | |||
| 286c | Coffea congensis Froehn. | —m | Rubiaceae | E | 22 | 2 | P | 794 | 0·8 | 1·6 | 3·2 | 424 | O | Kc | FC:PI | |||
| 287d | Coffea eugenioides S.Moore | —m | Rubiaceae | E | 22 | 2 | P | 681 | 0·7 | 1·4 | 2·8 | 424 | O | Kc | FC:PI | |||
| 288c | Coffea humilis A.Cheval. | —m | Rubiaceae | E | 22 | 2 | P | 872 | 0·9 | 1·8 | 3·6 | 424 | O | Kc | FC:PI | |||
| 289d | Coffea liberica L. ssp. dewevrei Wild & Dur. Hiern | No | Rubiaceae | E | 22 | 2 | P | 703 | 0·7 | 1·4 | 2·9 | 401 | O | Petuniae | FC:PI | |||
| 289e | Coffea liberica L. | —m | Rubiaceae | E | 22 | 2 | P | 823 | 0·8 | 1·7 | 3·4 | 424 | O | Kc | FC:PI | |||
| 290d | Coffea pseudozanguebariae D.M.Bridson | —m | Rubiaceae | E | 22 | 2 | P | 534 | 0·5 | 1·1 | 2·2 | 424 | O | Kc | FC:PI | |||
| 291c | Coffea racemosal | —m | Rubiaceae | E | 22 | 2 | P | 466 | 0·5 | 1·0 | 1·9 | 424 | O | Kc | FC:PI | |||
| 292c | Coffea sessiliflora D.M.Bridson | —m | Rubiaceae | E | 22 | 2 | P | 510 | 0·5 | 1·0 | 2·1 | 424 | O | Kc | FC:PI | |||
| 293 | Coffea sp. F. Bridsony | —m | Rubiaceae | E | 22 | 2 | P | 652 | 0·7 | 1·3 | 2·7 | 424 | O | Kc | FC:PI | |||
| 294 | Coffea sp. Moloundouy | —m | Rubiaceae | E | 22 | 2 | P | 828 | 0·8 | 1·7 | 3·4 | 424 | O | Kc | FC:PI | |||
| 295c | Coffea stenophylla G.Don. | —m | Rubiaceae | E | 22 | 2 | P | 662 | 0·7 | 1·4 | 2·7 | 424 | O | Kc | FC:PI | |||
| 296b | Colocasia antiquorum Schott var. 1i | No | Araceae | M | 32 | —p | AP | 4,951 | 5·1 | 10·1 | 20·2 | 411 | O | Bc | Fe | |||
| 297 | Commiphora mossambicensis Engl. | No | Burseraceae | E | 26 | 2 | P | 613 | 0·6 | 1·3 | 2·5 | 379 | O | J | Fe | |||
| 298 | Coriaria myrtifolia L. | No | Coriariaceae | E | c. 72 | 8 | P | 326 | 0·3 | 0·7 | 1·3 | 378 | O | J | Fe | |||
| 299 | Cosmos atrosanguineusl | No | Compositae | E | 48 | —p | P | 7,191 | 7·3 | 14·7 | 29·4 | 465 | O | F | Fe | |||
| 300c | Crepis biennis L.h | No | Compositae | E | c. 40 | 10 | B | 7,448 | 7·6 | 15·2 | 30·4 | 394ah | O | Gc | CIA | |||
| 300d | Crepis biennis L.h | No | Compositae | E | c. 40 | 10 | B | 7,928 | 8·1 | 16·2 | 32·4 | 394ah | O | Gc | CIA | |||
| 300e | Crepis biennis L.h | No | Compositae | E | c. 40 | 10 | B | 8,173 | 8·3 | 16·7 | 33·4 | 394ah | O | Gc | Fe | |||
| 300f | Crepis biennis L.h | No | Compositae | E | c. 40 | 10 | B | 8,555 | 8·7 | 17·5 | 34·9 | 394ah | O | Gc | Fe | |||
| 301a | Crepis bithynica var. pirinica Acht.i | No | Compositae | E | 10 | 2 | P | 3,156 | 3·2 | 6·4 | 12·9 | 394 | O | Gc | Fe | |||
| 301b | Crepis bithynica var. bithynica Boiss.i | No | Compositae | E | 10 | 2 | P | 3,244 | 3·3 | 6·6 | 13·2 | 394 | O | Gc | Fe | |||
| 302h | Crepis capillaris (L.) Wallr. | No | Compositaej | E | 6° | 2 | A | 2,597 | 2·7 | 5·3 | 10·6 | 457bm | O | Bd | Fe | |||
| 303a | Crepis conyzaefolia (Gouan) A.Kerneri | No | Compositae | E | 8 | 2 | P | 5,400 | 5·5 | 11·0 | 22·0 | 394 | O | Gc | Fe | |||
| 303b | Crepis conyzaefolia (Gouan) A.Kerneri | No | Compositae | E | 8 | 2 | P | 5,576 | 5·7 | 11·4 | 22·8 | 394 | O | Ec | FC:PI | |||
| 304a | Crepis paludosa (L.) Moenchi | No | Compositae | E | 12 | 2 | P | 4,077 | 4·2 | 8·3 | 16·6 | 394 | O | Gc | Fe | |||
| 304b | Crepis paludosa (L.) Moenchi | No | Compositae | E | 12 | 2 | P | 4,361 | 4·5 | 8·9 | 17·8 | 394 | O | Ec | FC:PI | |||
| 305b | Crepis pulchra L.h | No | Compositae | E | 8 | 2 | A | 4,459 | 4·6 | 9·1 | 18·2 | 394 | O | Gc | CIA | |||
| 305c | Crepis pulchra L.h | No | Compositae | E | 8 | 2 | A | 5,449 | 5·6 | 11·1 | 22·2 | 394ai | O | Gc | CIA | |||
| 305d | Crepis pulchra L.h | No | Compositae | E | 8 | 2 | A | 4,851 | 5·0 | 9·9 | 19·8 | 394 | O | Gc | Fe | |||
| 305e | Crepis pulchra L.h | No | Compositae | E | 8 | 2 | A | 5,400 | 5·5 | 11·0 | 22·0 | 394ai | O | Gc | Fe | |||
| 305f | Crepis pulchra L.h | No | Compositae | E | 8 | 2 | A | 5,380 | 5·5 | 11·0 | 22·0 | 394 | O | Ec | FC:PI | |||
| 305g | Crepis pulchra L.h | No | Compositae | E | 8 | 2 | A | 5,988 | 6·1 | 12·2 | 24·4 | 394 | O | Ec | FC:PI | |||
| 306a | Crepis sancta (L.) Babc.h | No | Compositae | E | 10 | 2 | A | 2,048 | 2·1 | 4·2 | 8·4 | 394ah | O | Gc | Fe | |||
| 306b | Crepis sancta (L.) Babc.h | No | Compositae | E | 10 | 2 | A | 2,176 | 2·2 | 4·4 | 8·9 | 394ah | O | Gc | CIA | |||
| 307 | Crepis schachtii Babc.i | No | Compositae | E | 10 | 2 | P | 2,764 | 2·8 | 5·6 | 11·3 | 394 | O | Gc | Fe | |||
| 308b | Crepis setosa Haller f.i | No | Compositae | E | 8 | 2 | A | 1,695 | 1·7 | 3·5 | 6·9 | 394 | O | Gc | CIA | |||
| 309a | Crepis viscidula Froel.i | No | Compositae | E | 12 | 2 | P | 4,283 | 4·4 | 8·7 | 17·5 | 394 | O | Gc | CIA | |||
| 309b | Crepis viscidula Froel.i | No | Compositae | E | 12 | 2 | P | 4,782 | 4·9 | 9·8 | 19·5 | 394 | O | Ec | FC:PI | |||
| 310b | Crepis zacintha (L.) Babc.i | No | Compositae | E | 6 | 2 | A | 1,058 | 1·1 | 2·2 | 4·3 | 394 | O | Gc | CIA | |||
| 311 | Crocus biflorus Mill. | No | Iridaceae | M | 8ao | 2 | P | 4,253 | 4·3 | 8·7 | 17·4 | 419 | O | Homof | FC:PI | |||
| 312 | Crocus cartwrightianus Herb.i | No | Iridaceae | M | 16 | 2 | P | 3,886 | 4·0 | 7·9 | 15·9 | 419 | O | Homof | FC:PI | |||
| 313 | Crocus etruscus Parl. | No | Iridaceae | M | 8 | 2 | P | 3,553 | 3·6 | 7·3 | 14·5 | 419 | O | Homof | FC:PI | |||
| 314 | Crocus sativus L.i | No | Iridaceae | M | 24 | 3 | P | —t | —t | 11·8 | 23·6 | 419 | O | Homof | FC:PI | |||
| 315 | Crocus thomasii Ten. | No | Iridaceae | M | 16 | 2 | P | 4,258 | 4·3 | 8·7 | 17·4 | 419 | O | Homof | FC:PI | |||
| 316 | Cyclamen hederifoliuml | No | Primulaceae | E | —n | —p | P | 2,923 | 3·0 | 6·0 | 11·9 | 465 | O | G | FC:PI | |||
| 317a | Cyclamen trochopteranthum O.Schwarzh | No | Primulaceae | E | 30 | —p | P | 10,373 | 10·6 | 21·2 | 42·3 | 465 | O | G | Fe | |||
| 317b | Cyclamen trochopteranthum O.Schwarzh | No | Primulaceae | E | 30 | —p | P | 13,458 | 13·7 | 27·5 | 54·9 | 465 | O | G | Fe | |||
| 318b | Cynodon dactylon (L.) Pers. | No | Gramineae | M | 36 | 4 | P | 956 | 1·0 | 2·0 | 3·9 | 417 | O | Gallusf | FC:PI | |||
| 318c | Cynodon dactylon (L.) Pers. var. dactylon | No | Gramineae | M | 36 | 4 | P | 1,103 | 1·1 | 2·3 | 4·5 | 438ba | O | Susf | FC:PI | |||
| 319 | Cynodon dactylon (L.) Pers. var. dactylon | No | Gramineae | M | 54 | 6 | P | 1,436 | 1·5 | 2·9 | 5·9 | 438ba | O | Ictal.f | FC:PI | |||
| 320a | Cynodon transvaalensis Burtt-Davy | No | Gramineae | M | 18 | 2 | P | 505 | 0·5 | 1·0 | 2·1 | 417 | O | Gallusf | FC:PI | |||
| 320b | Cynodon transvaalensis Burtt-Davy | No | Gramineae | M | 18 | 2 | P | 544 | 0·6 | 1·1 | 2·2 | 438ba | O | Ictal.f | FC:PI | |||
| 321 | Dalbergia horrida Dennst. | No | Leguminosae | E | 20 | 2 | P | 1,928 | 2·0 | 3·9 | 7·9 | 445 | O | G | Fe | |||
| 322a | Dalbergia lanceolaria Lin. f. | No | Leguminosae | E | 20 | 2 | P | 1,431 | 1·5 | 2·9 | 5·8 | 445 | O | G | Fe | |||
| 323a | Dalbergia latifolia Roxb. | No | Leguminosae | E | 20 | 2 | P | 1,681 | 1·7 | 3·4 | 6·9 | 445 | O | G | Fe | |||
| 324 | Dalbergia malabarica Prain. | No | Leguminosae | E | 20 | 2 | P | 1,847 | 1·9 | 3·8 | 7·5 | 445 | O | G | Fe | |||
| 325 | Dalbergia melanoxylon Guill & Perr. | No | Leguminosae | E | 20 | 2 | P | 1,806 | 1·8 | 3·7 | 7·4 | 445 | O | G | Fe | |||
| 326 | Dalbergia paniculata Roxb. | No | Leguminosae | E | 20 | 2 | P | 1,551 | 1·6 | 3·2 | 6·3 | 445 | O | G | Fe | |||
| 327 | Dalbergia rubiginosa Roxb. | No | Leguminosae | E | 20 | 2 | P | 1,803 | 1·8 | 3·7 | 7·4 | 445 | O | G | Fe | |||
| 328 | Dalbergia sissoides Grah. | No | Leguminosae | E | 20 | 2 | P | 1,764 | 1·8 | 3·6 | 7·2 | 445 | O | G | Fe | |||
| 329a | Dalbergia sissoo Roxb. ex DC. | No | Leguminosae | E | 20 | 2 | P | 1,585 | 1·6 | 3·2 | 6·5 | 445 | O | G | Fe | |||
| 329b | Dalbergia sissoo Roxb. ex DC. | No | Leguminosae | E | 20° | 2 | P | 691 | 0·7 | 1·4 | 2·8 | 454 | O | Bc | Fe | |||
| 330 | Dalbergia volubilis Roxb. | No | Leguminosae | E | 20 | 2 | P | 1,901 | 1·9 | 3·9 | 7·8 | 445 | O | G | Fe | |||
| 331 | Damasonium alisma Mill. | No | Alismataceae | M | —n | —p | A | 23,143 | 23·6 | 47·2 | 94·5 | 465 | O | B | FC:PI | |||
| 332 | Dasypogon hookeri Drumm. | No | Dasypogonaceae | M | 14 | 2 | P | 426 | 0·4 | 0·9 | 1·7 | 380 | O | J | FC:PI | |||
| 333 | Dasypyrum hordeaceum (Cosson & Durieu) Candargy | No | Gramineae | M | 28 | 4 | P | 10,288 | 10·5 | 21·0 | 42·0 | 431 | C | —aw | Fe | |||
| 334d | Dasypyrum villosum (=Haynaldia villosa) (L.) P. Candargyh | No | Gramineae | M | 14 | 2 | A | 10,408 | 10·6 | 21·2 | 42·5 | 430 | O | Cc | Fe | |||
| 334e | Dasypyrum villosum (=Haynaldia villosa) (L.) P. Candargyh | No | Gramineae | M | 14 | 2 | A | 6,262 | 6·4 | 12·8 | 25·6 | 430 | O | Cc | Fe | |||
| 335b | Decaisnea fargesii Franch. | No | Lardizabalaceaek | E | 40° | 2 | P | 2,450 | 2·5 | 5·0 | 10·0 | 457bm | O | Bd | Fe | |||
| 336 | Deutzia prunifolia Rehder | No | Hydrangeaceae | E | 52 | 4 | P | 1,835 | 1·9 | 3·7 | 7·5 | 378 | O | J | Fe | |||
| 337 | Dictamnus albus L. | No | Rutaceae | E | 36° | —p | P | 3,381 | 3·5 | 6·9 | 13·8 | 457bm | O | Bd | Fe | |||
| 338 | Dieffenbachia picta Schott | No | Araceae | M | 36 | —p | P | 12,083 | 12·3 | 24·7 | 49·3 | 411 | O | Bc | Fe | |||
| 339 | Diospyros discolor Willd. | No | Ebenaceae | E | 30° | —p | P | 1,174 | 1·2 | 2·4 | 4·8 | 454 | O | Bc | Fe | |||
| 340 | Diospyros malabarica Kost. | No | Ebenaceae | E | 30° | —p | P | 1,436 | 1·5 | 2·9 | 5·9 | 454 | O | Bc | Fe | |||
| 341 | Dissotis canescens Hook. f. | No | Melastomataceae | E | c. 28-32 | —p | P | 181 | 0·2 | 0·4 | 0·7 | 378 | O | J | Fe | |||
| 342a | Doritis pulcherrima Lindl. | No | Orchidaceae | M | 38 | 2 | P | 6,610 | 6·7 | 13·5 | 27·0 | 447 | O | Gb2 | FC:PI | |||
| 343 | Doryanthes palmeri W. Hill ex Benth. | No | Doryanthaceae | M | 48 | —p | P | 3,239 | 3·3 | 6·6 | 13·2 | 380 | O | G | FC:PI | |||
| 344 | Drimys vickeriana A.C. Smith | No | Winteraceae | BA | —n | —p | P | 1,105 | 1·1 | 2·3 | 4·5 | 381 | O | K | FC:PI | |||
| 345 | Drypetes roxburghii Wall. | No | Putranjavaceaek | E | 42° | —p | P | 1,002 | 1·0 | 2·0 | 4·1 | 449bf | O | Bc | Fe | |||
| 346 | Ehretia laevis (Rottler ex G.Don) Roxb. | No | Boraginaceae | E | —n | —p | P | 3,533 | 3·6 | 7·2 | 14·4 | 454 | O | Bc | Fe | |||
| 347 | Eremochloa ophiuroides (Munro) Hack. | No | Gramineae | M | 18 | 2 | P | 813 | 0·8 | 1·7 | 3·3 | 417 | O | Gallusf | FC:PI | |||
| 348 | Eriocaulon aquaticuml | No | Eriocaulaceae | M | 32 | 4 | P | 4,101 | 4·2 | 8·4 | 16·7 | 380 | O | G | FC:PI | |||
| 349 | Escallonia rubral | No | Escalloniaceae | E | 24 | 2 | P | 414 | 0·4 | 0·8 | 1·7 | 380 | O | J | Fe | |||
| 350 | Eucnide grandiflora Rose | No | Loasaceae | E | c. 38-40 | 2 or 6 | P | 588 | 0·6 | 1·2 | 2·4 | 378 | O | J | Fe | |||
| 351 | Eucommia ulmoides Oliver | No | Eucommiaceae | E | 34 | 2 | P | 725 | 0·7 | 1·5 | 3·0 | 379 | O | G | Fe | |||
| 352a | Fagus sylvatica L. var. tortuosa Pepin Willk. | No | Fagaceae | E | 24° | 2 | P | 544 | 0·6 | 1·1 | 2·2 | 433 | O | Petuniae | FC:PI | |||
| 352b | Fagus sylvatica L. | No | Fagaceae | E | 24° | 2 | P | 544 | 0·6 | 1·1 | 2·2 | 433 | O | Petuniae | FC:PI | |||
| 352c | Fagus sylvatica L. var. purpurea Ait. | No | Fagaceae | E | 24° | 2 | P | 549 | 0·6 | 1·1 | 2·2 | 433 | O | Petuniae | FC:PI | |||
| 352d | Fagus sylvatica L. var. pendula Lodd. | No | Fagaceae | E | 24° | 2 | P | 554 | 0·6 | 1·1 | 2·3 | 433 | O | Petuniae | FC:PI | |||
| 353d | Festuca arundinacea Schreb. | No | Gramineae | M | 42 | 6 | P | 7,639 | 7·8 | 15·6 | 31·2 | 417 | O | Nicot.e | FC:PI | |||
| 354 | Festuca longifolia Thuill. | No | Gramineae | M | 42 | 6 | P | 6,223 | 6·4 | 12·7 | 25·4 | 417 | O | Fc | FC:PI | |||
| 355 | Firmiana colorata (Roxb.) R.Br. | No | Malvaceae | E | 40° | —p | P | 1,615 | 1·6 | 3·3 | 6·6 | 454 | O | Bc | Fe | |||
| 356 | Flagellaria guineensis Schum. | No | Flagellariaceae | M | 38° | 2 | P | 880 | 0·9 | 1·8 | 3·6 | 380 | O | K | FC:PI | |||
| 357 | Flemingia bracteata Wight | No | Leguminosae | E | 22 | 2 | P | 1,570 | 1·6 | 3·2 | 6·4 | 443bc | O | Bc | Fe | |||
| 358 | Fortunella hindsii Swing. | No | Rutaceae | E | 36° | 4 | P | 622 | 0·6 | 1·3 | 2·5 | 426 | O | Gallusf | FC:PI | |||
| 359 | Fouquieria splendens Engelm. | No | Fouquieriaceae | E | 24 | 4 | P | 519 | 0·5 | 1·1 | 2·1 | 378 | O | J | Fe | |||
| 360 | Fragaria x ananassa cv. Redcoat Duch. | —m | Rosaceae | E | 56 | 8 | P | 598 | 0·6 | 1·2 | 2·4 | 442 | O | Gallusf | FC:M | |||
| 361a | Gagea lutea (L.) Ker Gawl. | No | Liliaceae | M | 72 | 6 | P | 19,355 | 19·8 | 39·5 | 79·0 | 413 | O | Bc | Fe | |||
| 361b | Gagea lutea (L.) Ker Gawl. | No | Liliaceae | M | 72 | 6 | P | 19,825 | 20·2 | 40·5 | 80·9 | 413 | O | Bc | FC:EB | |||
| 362 | Gardenia resiniflua Hiern | No | Rubiaceae | E | —n | —p | P | 1,269 | 1·3 | 2·6 | 5·2 | 454 | O | Bc | Fe | |||
| 363 | Garrya fremontii Torr. | No | Garryaceae | E | c. 20 | 2 | P | 1,490 | 1·5 | 3·0 | 6·1 | 380 | O | Lycopers.c | FC:PI | |||
| 364j | Glycine max (L.) Merr. strain T215h | No | Leguminosae | E | 40° | 2 | A | 1,161 | 1·2 | 2·4 | 4·7 | 423as | O | Fc | FC:PI | |||
| 364k | Glycine max (L.) Merr. strain PI423·894h | No | Leguminosae | E | 40° | 2 | A | 1,215 | 1·2 | 2·5 | 5·0 | 423as | O | Fc | FC:PI | |||
| 364l | Glycine max (L.) Merr.h | —m | Leguminosae | E | 40° | 2 | A | 1,250 | 1·3 | 2·6 | 5·1 | 432ax | C | Glycinee | FC:PI | |||
| 364m | Glycine max (L.) Merr.h | —m | Leguminosae | E | 40° | 2 | A | 1,401 | 1·4 | 2·9 | 5·7 | 432ax | C | Glycinee | FC:PI | |||
| 365 | Goodenia mimuloides S.Moore | No | Goodeniaceae | E | 16 | 2 | A | 507 | 0·5 | 1·0 | 2·1 | 379 | O | G | Fe | |||
| 366 | Gossypioides herbaceum L. | —m | Malvaceae | E | 26° | 2 | —q | 1,813 | 1·9 | 3·7 | 7·4 | 444bd | O | Gb2 | FC:PI | |||
| 367 | Gossypioides raimondii Ulbrich | —m | Malvaceae | E | 26° | 2 | —q | 980 | 1·0 | 2·0 | 4·0 | 444bd | O | Gb2 | FC:PI | |||
| 368 | Gunnera manicata Linden | No | Gunneraceae | E | 34 | 2 | P | 7,286 | 7·4 | 14·9 | 29·7 | 379 | O | F | Fe | |||
| 369 | Gymnostoma deplancheana (Miq.) L.Johnson | No | Moraceaek | E | 16° | 2 | P | 368 | 0·4 | 0·8 | 1·5 | 452 | O | Petuniae | FC:PI | |||
| 370 | Haldina cordifolia (Roxb.) Ridsdale | No | Rubiaceae | E | 44° | 4 | P | 1,296 | 1·3 | 2·6 | 5·3 | 454 | O | Bc | Fe | |||
| 371 | Hampea appendiculata (J. Donnell-Smith) Standley | —m | Malvaceae | E | 26° | 2 | —q | 2,891 | 3·0 | 5·9 | 11·8 | 444bd | O | Gb2 | FC:PI | |||
| 372 | Hanguana malayana Merrill | No | Hanguanaceae | M | c. 170 | —p | P | 1,612 | 1·6 | 3·3 | 6·6 | 380 | O | F | Fe | |||
| 373a | Hedera canariensis Willd. | No | Araliaceae | E | 48° | 2 | P | 1,372 | 1·4 | 2·8 | 5·6 | 429 | O | Glycinee | FC:PI | |||
| 373b | Hedera canariensis Willd. | No | Araliaceae | E | 48° | 2 | P | 1,509 | 1·5 | 3·1 | 6·2 | 429 | O | Gc | CIA | |||
| 374a | Hedera colchica C.Koch. | No | Araliaceae | E | 192 | 8 | P | 5,341 | 5·5 | 10·9 | 21·8 | 429 | O | Glycinee | FC:PI | |||
| 374b | Hedera colchica C.Koch. | No | Araliaceae | E | 192 | 8 | P | 5,586 | 5·7 | 11·4 | 22·8 | 429 | O | Gc | CIA | |||
| 375e | Hedera helix L. | No | Araliaceae | E | 48 | 2 | P | 1,460 | 1·5 | 3·0 | 6·0 | 429av | O | Gc | CIA | |||
| 375f | Hedera helix L. | No | Araliaceae | E | 48 | 2 | P | 1,372 | 1·4 | 2·8 | 5·6 | 429av | O | Glycinee | FC:PI | |||
| 375g | Hedera helix L. f. arborescens C.K. Schneider | No | Araliaceae | E | 48 | 2 | P | 1,509 | 1·5 | 3·1 | 6·2 | 429 | O | Gc | CIA | |||
| 375h | Hedera helix L. f. arborescens C.K. Schneider | No | Araliaceae | E | 48 | 2 | P | 1,382 | 1·4 | 2·8 | 5·6 | 429 | O | Glycinee | FC:PI | |||
| 376r | Helianthus annuus L. | No | Compositae | E | 34 | 2 | A | 3,577 | 3·7 | 7·3 | 14·6 | 403 | O | G | FC:PI | |||
| 377 | Heliconia rostrata Ruiz & Pav. | No | Heliconiaceae | M | 24 | 2 | P | 441 | 0·5 | 0·9 | 1·8 | 379 | O | J | Fe | |||
| 378 | Helleborus argutifolius Viv. | No | Ranunculaceae | E | 32 | 2 | P | 9,261 | 9·5 | 18·9 | 37·8 | 383z | O | Agavee | FC:PI | |||
| 379a | Helleborus atrorubens Waldst. & Kit. ‘Cupreus’i | No | Ranunculaceae | E | 32 | 2 | P | 14,504 | 14·8 | 29·6 | 59·2 | 383z | O | Agavee | FC:PI | |||
| 379b | Helleborus atrorubens Waldst. & Kit.i | No | Ranunculaceae | E | 32 | 2 | P | 15,092 | 15·4 | 30·8 | 61·6 | 383z | O | Agavee | FC:PI | |||
| 380 | Helleborus cyclophyllus (A.Br.) Boiss. | No | Ranunculaceae | E | 32 | 2 | P | 14,651 | 15·0 | 29·9 | 59·8 | 383z | O | Agavee | FC:PI | |||
| 381 | Helleborus dumetorum Waldst. & Kit. | No | Ranunculaceae | E | 32 | 2 | P | 15,876 | 16·2 | 32·4 | 64·8 | 383z | O | Agavee | FC:PI | |||
| 382a | Helleborus foetidus L. ‘Wester Flisk’ | No | Ranunculaceae | E | 32 | 2 | P | 11,417 | 11·7 | 23·3 | 46·6 | 383z | O | Agavee | FC:PI | |||
| 382b | Helleborus foetidus L.i | No | Ranunculaceae | E | 32 | 2 | P | 11,466 | 11·7 | 23·4 | 46·8 | 383z | O | Agavee | FC:PI | |||
| 383 | Helleborus lividus Aiton | No | Ranunculaceae | E | 32 | 2 | P | 9,310 | 9·5 | 19·0 | 38·0 | 383z | O | Agavee | FC:PI | |||
| 384a | Helleborus multifidus Vis. ssp. hercegovinus (Martinis) B.Mathew | No | Ranunculaceae | E | 32 | 2 | P | 14,504 | 14·8 | 29·6 | 59·2 | 383z | O | Agavee | FC:PI | |||
| 384b | Helleborus multifidus Vis. ssp. istriacus (Schiffner) Merxm & Podl. | No | Ranunculaceae | E | 32 | 2 | P | 14,749 | 15·1 | 30·1 | 60·2 | 383z | O | Agavee | FC:PI | |||
| 384c | Helleborus multifidus Vis. ssp. multifidus | No | Ranunculaceae | E | 32 | 2 | P | 14,798 | 15·1 | 30·2 | 60·4 | 383z | O | Agavee | FC:PI | |||
| 384d | Helleborus multifidus Vis. ssp. bocconei siculus | No | Ranunculaceae | E | 32 | 2 | P | 15,043 | 15·4 | 30·7 | 61·4 | 383z | O | Agavee | FC:PI | |||
| 384e | Helleborus multifidus Vis.ssp. bocconei (Tenore) B.Mathew | No | Ranunculaceae | E | 32 | 2 | P | 15,092 | 15·4 | 30·8 | 61·6 | 383z | O | Agavee | FC:PI | |||
| 385a | Helleborus niger L. (double flower)i | No | Ranunculaceae | E | 32 | 2 | P | 13,720 | 14·0 | 28·0 | 56·0 | 383z | O | Agavee | FC:PI | |||
| 385b | Helleborus niger L.i | No | Ranunculaceae | E | 32 | 2 | P | 13,867 | 14·2 | 28·3 | 56·6 | 383z | O | Agavee | FC:PI | |||
| 385c | Helleborus niger L. ssp. macranthus (Freyn) Schiffneri | No | Ranunculaceae | E | 32 | 2 | P | 14,406 | 14·7 | 29·4 | 58·8 | 383z | O | Agavee | FC:PI | |||
| 386 | Helleborus odorus Waldst. & Kit.i | No | Ranunculaceae | E | 32 | 2 | P | 15,043 | 15·4 | 30·7 | 61·4 | 383z | O | Agavee | FC:PI | |||
| 387a | Helleborus orientalis Lamarck ssp. orientalisi | No | Ranunculaceae | E | 32 | 2 | P | 14,553 | 14·9 | 29·7 | 59·4 | 383z | O | Agavee | FC:PI | |||
| 387b | Helleborus orientalis Lamarcki | No | Ranunculaceae | E | 32 | 2 | P | 14,725 | 15·0 | 30·1 | 60·1 | 383z | O | Agavee | FC:PI | |||
| 387c | Helleborus orientalis Lamarck ssp. guttatus (A.Br. & Sauer) B.Mathewi | No | Ranunculaceae | E | 32 | 2 | P | 14,749 | 15·1 | 30·1 | 60·2 | 383z | O | Agavee | FC:PI | |||
| 387d | Helleborus orientalis Lamarck ssp. abchasicus (A.Br.) B.Mathewi | No | Ranunculaceae | E | 32 | 2 | P | 14,798 | 15·1 | 30·2 | 60·4 | 383z | O | Agavee | FC:PI | |||
| 387e | Helleborus orientalis Lamarck ‘Kochii’i | No | Ranunculaceae | E | 32 | 2 | P | 14,994 | 15·3 | 30·6 | 61·2 | 383z | O | Agavee | FC:PI | |||
| 388 | Helleborus purpurascens Waldst. & Kit. | No | Ranunculaceae | E | 32 | 2 | P | 14,945 | 15·3 | 30·5 | 61·0 | 383z | O | Agavee | FC:PI | |||
| 389 | Helleborus thibetanus Franchet | No | Ranunculaceae | E | 32 | 2 | P | 17,493 | 17·9 | 35·7 | 71·4 | 383z | O | Agavee | FC:PI | |||
| 390a | Helleborus torquatusArcher Hind ‘Dido’ (double flowers)i | No | Ranunculaceae | E | 32 | 2 | P | 14,602 | 14·9 | 29·8 | 59·6 | 383z | O | Agavee | FC:PI | |||
| 390b | Helleborus torquatus Archer Hindi | No | Ranunculaceae | E | 32 | 2 | P | 14,749 | 15·1 | 30·1 | 60·2 | 383z | O | Agavee | FC:PI | |||
| 390c | Helleborus torquatus Archer Hind ‘Croaticus’i | No | Ranunculaceae | E | 32 | 2 | P | 14,700 | 15·0 | 30·0 | 60·0 | 383z | O | Agavee | FC:PI | |||
| 391 | Helleborus vesicarius Aucher | No | Ranunculaceae | E | 32 | 2 | P | 13,867 | 14·2 | 28·3 | 56·6 | 383z | O | Agavee | FC:PI | |||
| 392a | Helleborus viridis L. ssp. viridis | No | Ranunculaceae | E | 32 | 2 | P | 14,896 | 15·2 | 30·4 | 60·8 | 383z | O | Agavee | FC:PI | |||
| 392b | Helleborus viridis L. ssp. occidentalis (Reut.) Schiffner | No | Ranunculaceae | E | 32 | 2 | P | 15,092 | 15·4 | 30·8 | 61·6 | 383z | O | Agavee | FC:PI | |||
| 393 | Hernandia nymphaeifolia (C.Presl.) Kubitzki | No | Hernandiaceae | BA | —n | —p | P | 2,340 | 2·4 | 4·8 | 9·6 | 454 | O | Bc | Fe | |||
| 394 | Herniaria glabra Linn. | No | Caryophyllaceae | E | 18 | 2 | AP | 515 | 0·5 | 1·1 | 2·1 | 465 | O | J | Fe | |||
| 395b | Hieracium aurantiacum L. | No | Compositaej | E | 36° | 4 | P | 3,626 | 3·7 | 7·4 | 14·8 | 457bm | O | Bd | Fe | |||
| 396 | Holoptelea integrifolia Planch. | No | Ulmaceae | E | 28° | —p | P | 666 | 0·7 | 1·4 | 2·7 | 454 | O | Bc | Fe | |||
| 397 | Homalomena rubescens Kunth | No | Araceae | M | 34 | —p | AP | 8,955 | 9·1 | 18·3 | 36·6 | 411 | O | Bc | Fe | |||
| 398p | Hordeum vulgare L. cv. New Golden | No | Gramineae | M | 14° | 2 | A | 5,096 | 5·2 | 10·4 | 20·8 | 387ae | O | Gallusf | FC:PI | |||
| 399 | Hosta capitata (Koidzumi) Nakai | No | Asparagaceaek | M | 60 | 2 | P | 9,457 | 9·7 | 19·3 | 38·6 | 384aa | O | Agave sp.ab | FC:PI | |||
| 400 | Hosta clausa var. normalis F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 9,408 | 9·6 | 19·2 | 38·4 | 384aa | O | Agave sp.ab | FC:PI | |||
| 401 | Hosta clausa Nakai var. clausa | No | Asparagaceaek | M | 90 | 3 | P | —t | —t | 28·5 | 57·0 | 384aa | O | Agave sp.ab | FC:PI | |||
| 402 | Hosta gracillima F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 10,829 | 11·1 | 22·1 | 44·2 | 384aa | O | Agave sp.ab | FC:PI | |||
| 403 | Hosta hypoleuca Murata | No | Asparagaceaek | M | 60 | 2 | P | 12,495 | 12·8 | 25·5 | 51·0 | 384aa | O | Agave sp.ab | FC:PI | |||
| 404 | Hosta jonesii M.Chung | No | Asparagaceaek | M | 60 | 2 | P | 8,575 | 8·8 | 17·5 | 35·0 | 384aa | O | Agave sp.ab | FC:PI | |||
| 405 | Hosta kikutii F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 11,172 | 11·4 | 22·8 | 45·6 | 384aa | O | Agave sp.ab | FC:PI | |||
| 406 | Hosta kiyosumiensis F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 11,907 | 12·2 | 24·3 | 48·6 | 384aa | O | Agave sp.ab | FC:PI | |||
| 407 | Hosta longipes var. longipes Matsumura | No | Asparagaceaek | M | 60 | 2 | P | 12,740 | 13·0 | 26·0 | 52·0 | 384aa | O | Agave sp.ab | FC:PI | |||
| 408 | Hosta longissima Honda | No | Asparagaceaek | M | 60 | 2 | P | 9,604 | 9·8 | 19·6 | 39·2 | 384aa | O | Agave sp.ab | FC:PI | |||
| 409 | Hosta minor Nakai ‘Gosan’ | No | Asparagaceaek | M | 60 | 2 | P | 8,428 | 8·6 | 17·2 | 34·4 | 384aa | O | Agave sp.ab | FC:PI | |||
| 410 | Hosta plantaginea (Lamarck) Ascherson | No | Asparagaceaek | M | 60 | 2 | P | 12,103 | 12·4 | 24·7 | 49·4 | 384aa | O | Agave sp.ab | FC:PI | |||
| 411 | Hosta pulchella N.Fujita | No | Asparagaceaek | M | 60 | 2 | P | 10,633 | 10·9 | 21·7 | 43·4 | 384aa | O | Agave sp.ab | FC:PI | |||
| 412 | Hosta pycnophylla F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 10,878 | 11·1 | 22·2 | 44·4 | 384aa | O | Agave sp.ab | FC:PI | |||
| 413b | Hosta rectifolia Nakai | No | Asparagaceaek | M | 60 | 2 | P | 10,437 | 10·7 | 21·3 | 42·6 | 384aa | O | Agave sp.ab | FC:PI | |||
| 414 | Hosta rupifraga Nakai | No | Asparagaceaek | M | 60 | 2 | P | 12,985 | 13·3 | 26·5 | 53·0 | 384aa | O | Agave sp.ab | FC:PI | |||
| 415 | Hosta shikokiana N.Fujita | No | Asparagaceaek | M | 60 | 2 | P | 11,221 | 11·5 | 22·9 | 45·8 | 384aa | O | Agave sp.ab | FC:PI | |||
| 416 | Hosta sieboldiana var. sieboldiana (Hooker) Engler | No | Asparagaceaek | M | 60 | 2 | P | 11,564 | 11·8 | 23·6 | 47·2 | 384aa | O | Agave sp.ab | FC:PI | |||
| 417 | Hosta sieboldii P.O.(Paxton) Ingram | No | Asparagaceaek | M | 60 | 2 | P | 11,025 | 11·3 | 22·5 | 45·0 | 384aa | O | Agave sp.ab | FC:PI | |||
| 418 | Hosta tibae F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 8,624 | 8·8 | 17·6 | 35·2 | 384aa | O | Agave sp.ab | FC:PI | |||
| 419 | Hosta tsushimensis N.Fujita | No | Asparagaceaek | M | 60 | 2 | P | 8,477 | 8·7 | 17·3 | 34·6 | 384aa | O | Agave sp.ab | FC:PI | |||
| 420 | Hosta ventricosa Stearn | No | Asparagaceaek | M | 120 | 4 | P | 19,208 | 19·6 | 39·2 | 78·4 | 384aa | O | Agave sp.ab | FC:PI | |||
| 421 | Hosta venusta F.Maekawa | No | Asparagaceaek | M | 60 | 2 | P | 8,477 | 8·7 | 17·3 | 34·6 | 384aa | O | Agave sp.ab | FC:PI | |||
| 422 | Hosta yingeri S.B.Jones | No | Asparagaceaek | M | 60 | 2 | P | 9,359 | 9·6 | 19·1 | 38·2 | 384aa | O | Agave sp.ab | FC:PI | |||
| 423a | Hydrangea anomala D.Don ssp. petiolaris Sieb. & Zucc. | No | Hydrangeaceae | E | 36 | 2 | P | 1,328 | 1·4 | 2·7 | 5·4 | 397 | O | Gc | FC:EB | |||
| 423b | Hydrangea anomala D.Don ssp. anomala McClint. | No | Hydrangeaceae | E | 36 | 2 | P | 1,534 | 1·6 | 3·1 | 6·3 | 397 | O | Gc | FC:EB | |||
| 424 | Hydrangea arborescens L. | No | Hydrangeaceae | E | 36 | 2 | P | 1,132 | 1·2 | 2·3 | 4·6 | 397 | O | Gc | FC:EB | |||
| 425a | Hydrangea aspera Don. ssp. robusta McClint. (=H. longipes Franch.) | No | Hydrangeaceae | E | 34 | 2 | P | 1,480 | 1·5 | 3·0 | 6·0 | 397 | O | Gc | FC:EB | |||
| 425b | Hydrangea aspera Don. ssp. sargentiana (Rehder) McClint. | No | Hydrangeaceae | E | 34 | 2 | P | 1,529 | 1·6 | 3·1 | 6·2 | 397 | O | Gc | FC:EB | |||
| 425c | Hydrangea aspera Don. ssp. strigosa McClint. | No | Hydrangeaceae | E | 34 | 2 | P | 1,700 | 1·7 | 3·5 | 6·9 | 397 | O | Gc | FC:EB | |||
| 425d | Hydrangea aspera Don. ssp. aspera McClint. | No | Hydrangeaceae | E | 36 | 2 | P | 2,323 | 2·4 | 4·7 | 9·5 | 397 | O | Gc | FC:EB | |||
| 426 | Hydrangea heteromalla D.Don | No | Hydrangeaceae | E | 36 | 2 | P | 1,446 | 1·5 | 3·0 | 5·9 | 397 | O | Gc | FC:EB | |||
| 427 | Hydrangea involucrata Sieb. | No | Hydrangeaceae | E | 30 | 2 | P | 2,450 | 2·5 | 5·0 | 10·0 | 397 | O | Gc | FC:EB | |||
| 428a | Hydrangea macrophylla (Thunb.) Ser. ssp. serrata (Thunb.) Makino | No | Hydrangeaceae | E | 36 | 2 | P | 1,887 | 1·9 | 3·9 | 7·7 | 397 | O | Gc | FC:EB | |||
| 428b | Hydrangea macrophylla (Thunb.) Ser. ssp. macrophylla McClint. | No | Hydrangeaceae | E | 36 | 2 | P | 2,107 | 2·2 | 4·3 | 8·6 | 397 | O | Gc | FC:EB | |||
| 429 | Hydrangea paniculata Sieb. | No | Hydrangeaceae | E | 36 | 2 | P | 1,847 | 1·9 | 3·8 | 7·5 | 397 | O | Gc | FC:EB | |||
| 430 | Hydrangea quercifolia Bartr. | No | Hydrangeaceae | E | 36 | 2 | P | 956 | 1·0 | 2·0 | 3·9 | 397 | O | Gc | FC:EB | |||
| 431a | Hydrangea scandens (L.f.) ssp. scandens McClint. | No | Hydrangeaceae | E | 36 | 2 | P | 1,803 | 1·8 | 3·7 | 7·4 | 397 | O | Gc | FC:EB | |||
| 431b | Hydrangea scandens (L.f.) ssp. luikinensis (Nakai) McClint. | No | Hydrangeaceae | E | 36 | 2 | P | 1,872 | 1·9 | 3·8 | 7·6 | 397 | O | Gc | FC:EB | |||
| 432 | Hydrangea seemannii Riley | No | Hydrangeaceae | E | 36 | 2 | P | 1,024 | 1·0 | 2·1 | 4·2 | 397 | O | Gc | FC:EB | |||
| 433 | Hypericum hirsutuml | No | Hypericaceae | E | —n | —p | P | 147 | 0·2 | 0·3 | 0·6 | 465 | O | J | Fe | |||
| 434 | Inga dulcis (Roxb.) Willd. | No | Leguminosae | E | —n | —p | P | 402 | 0·4 | 0·8 | 1·6 | 454 | O | Bc | Fe | |||
| 435a | Iris stenophylla Hausskn. ex Bakerh | No | Iridaceae | M | 24 | 2 | P | 8,359 | 8·5 | 17·1 | 34·1 | 465 | O | B | Fe | |||
| 435b | Iris stenophylla Hausskn. ex Bakerh | No | Iridaceae | M | 26 | 2 | P | 10,743 | 11·0 | 21·9 | 43·9 | 465 | O | B | Fe | |||
| 435c | Iris stenophylla Hausskn. ex Bakerh | No | Iridaceae | M | 26 | 2 | P | 11,439 | 11·7 | 23·3 | 46·7 | 465 | O | B | Fe | |||
| 436 | Ixiolirion ledebourii Fisch. & Mey. | No | Ixioliriaceae | M | c. 24 | 2 | P | 995 | 1·0 | 2·0 | 4·1 | 380 | O | J | Fe | |||
| 437 | Ixora arborea Roxb. ex Sm. | No | Rubiaceae | E | —n | —p | P | 1,365 | 1·4 | 2·8 | 5·6 | 454 | O | Bc | Fe | |||
| 438 | Jacquinia aristata Jacq. | No | Theophrastaceae | E | 38 | 2 | P | 593 | 0·6 | 1·2 | 2·4 | 378 | O | J | Fe | |||
| 439 | Kentranthus ruber Druce | No | Valerianaceae | E | 32 | 4 | P | 407 | 0·4 | 0·8 | 1·7 | 379 | O | G | Fe | |||
| 440 | Khaya senegalensis (Desr.) A.Juss. | No | Meliaceae | E | —n | —p | P | 853 | 0·9 | 1·7 | 3·5 | 454 | O | Bc | Fe | |||
| 441 | Kigelia africana (Lam.) Benth. | No | Bignoniaceae | E | —n | —p | P | 1,700 | 1·7 | 3·5 | 6·9 | 454 | O | Bc | Fe | |||
| 442 | Kirkia acuminata Oliver | No | Kirkiaceae | E | c. 30 | 2 | P | 319 | 0·3 | 0·7 | 1·3 | 378 | O | J | Fe | |||
| 443 | Lagerstroemia tomentosa C.Presl. Presl. | No | Lythraceae | E | —n | —p | P | 965 | 1·0 | 2·0 | 3·9 | 454 | O | Bc | Fe | |||
| 444 | Lantana camara L. | No | Verbenaceae | E | 22° | —p | P | 2,697 | 2·8 | 5·5 | 11·0 | 454 | O | Bc | Fe | |||
| 445 | Lapageria rosea Ruiz & Pav. | No | Philesiaceae | M | 30 + 1B | 2 | P | 6,644 | 6·8 | 13·6 | 27·1 | 380 | O | G | Fe | |||
| 446b | Lathyrus amphicarpos L. | —m | Leguminosae | E | 14 | 2 | A | 5,123 | 5·2 | 10·5 | 20·9 | 412 | O | —v | Fe | |||
| 447e | Lathyrus annuus L. | —m | Leguminosae | E | 14 | 2 | A | 6,429 | 6·6 | 13·1 | 26·2 | 412 | O | —v | Fe | |||
| 447f | Lathyrus annuus L. | —m | Leguminosae | E | 14 | 2 | A | 6,282 | 6·4 | 12·8 | 25·6 | 418 | O | Gc | FC:PI | |||
| 448j | Lathyrus aphaca L. | —m | Leguminosae | E | 14 | 2 | A | 4,528 | 4·6 | 9·2 | 18·5 | 418 | O | C & Ec | FC:PI | |||
| 449b | Lathyrus chloranthus Boiss. | —m | Leguminosae | E | 14 | 2 | A | 5,944 | 6·1 | 12·1 | 24·3 | 412 | O | —v | Fe | |||
| 450f | Lathyrus cicera L. | —m | Leguminosae | E | 14 | 2 | A | 5,194 | 5·3 | 10·6 | 21·2 | 418 | O | Gc | FC:PI | |||
| 451e | Lathyrus clymenum L. | —m | Leguminosae | E | 14 | 2 | A | 4,297 | 4·4 | 8·8 | 17·5 | 418 | O | C & Ec | FC:PI | |||
| 451f | Lathyrus clymenum L. | —m | Leguminosae | E | 14 | 2 | A | 6,399 | 6·5 | 13·1 | 26·1 | 412 | O | —v | Fe | |||
| 452 | Lathyrus gmelinii Fritsch | —m | Leguminosae | E | 14 | 2 | P | 8,791 | 9·0 | 17·9 | 35·9 | 412 | O | —v | Fe | |||
| 453b | Lathyrus grandiflorus Sibth. & Sm. | —m | Leguminosae | E | 14 | 2 | P | 8,724 | 8·9 | 17·8 | 35·6 | 412 | O | —v | Fe | |||
| 454b | Lathyrus heterophyllus L. | —m | Leguminosae | E | 14 | 2 | P | 8,639 | 8·8 | 17·6 | 35·3 | 412 | O | —v | Fe | |||
| 455 | Lathyrus laevigatus (Waldst. & Kit.) | —m | Leguminosae | E | 14 | 2 | P | 11,008 | 11·2 | 22·5 | 44·9 | 412 | O | —v | Fe | |||
| 456c | Lathyrus maritimus Bigelow | —m | Leguminosae | E | 14 | 2 | P | 6,777 | 6·9 | 13·8 | 27·7 | 412 | O | —v | Fe | |||
| 457e | Lathyrus nissolia L. | —m | Leguminosae | E | 14 | 2 | A | 4,853 | 5·0 | 9·9 | 19·8 | 412 | O | —v | Fe | |||
| 458e | Lathyrus ochrus (L.) DC | —m | Leguminosae | E | 14 | 2 | A | 4,542 | 4·6 | 9·3 | 18·5 | 418 | O | C & Ec | FC:PI | |||
| 459i | Lathyrus odoratus L. | No | Leguminosaej | E | 14° | 2 | A | 8,134 | 8·3 | 16·6 | 33·2 | 457bm | O | Bd | Fe | |||
| 460f | Lathyrus sativus L. | —m | Leguminosae | E | 14 | 2 | A | 6,551 | 6·7 | 13·4 | 26·7 | 418 | O | Gc | FC:PI | |||
| 460g | Lathyrus sativus L. | —m | Leguminosae | E | 14 | 2 | A | 6,899 | 7·0 | 14·1 | 28·2 | 412 | O | —v | Fe | |||
| 461f | Lathyrus sylvestris L. | —m | Leguminosae | E | 14 | 2 | P | 10,104 | 10·3 | 20·6 | 41·2 | 412 | O | —v | Fe | |||
| 462g | Lathyrus tingitanus L. | —m | Leguminosae | E | 14 | 2 | A | 7,693 | 7·9 | 15·7 | 31·4 | 418 | O | Gc | FC:PI | |||
| 462h | Lathyrus tingitanus L. | —m | Leguminosae | E | 14 | 2 | A | 7,691 | 7·8 | 15·7 | 31·4 | 412 | O | —v | Fe | |||
| 465 | Lawsonia inermis L. | No | Lythraceae | E | c. 30-34 | 4 | P | 333 | 0·3 | 0·7 | 1·4 | 378 | O | J | Fe | |||
| 464 | Lebronnecia kokioides Fosberg | —m | Malvaceae | E | 26 | 2 | —q | 1,764 | 1·8 | 3·6 | 7·2 | 444bd | O | Gb2 | FC:PI | |||
| 465 | Lemna minor L. | No | Araceae | M | 126 | 6 | P | 1,426 | 1·5 | 2·9 | 5·8 | 400 | O | G | Fe | |||
| 466 | Leucaena collinsii Britton & Rose | No | Leguminosae | E | 52, ?56° | 2 | P | 529 | 0·5 | 1·1 | 2·2 | 425 | O | —at | FC:EB | |||
| 467b | Leucaena confertiflora S.Zarate | No | Leguminosae | E | 112° | 4 | P | 828 | 0·8 | 1·7 | 3·4 | 425 | O | —at | FC:EB | |||
| 468 | Leucaena cuspidata Standley | No | Leguminosae | E | —n | —p | P | 686 | 0·7 | 1·4 | 2·8 | 425 | O | —at | FC:EB | |||
| 469 | Leucaena diversifolia (Schltdl. Benth. | No | Leguminosae | E | 104 | 4 | P | 1,328 | 1·4 | 2·7 | 5·4 | 425 | O | —at | FC:EB | |||
| 470c | Leucaena esculenta (Sesse & Moc. ex DC) Benth. | No | Leguminosae | E | 52 (?56, ?112) | —p | P | 706 | 0·7 | 1·4 | 2·9 | 425 | O | —at | FC:EB | |||
| 471 | Leucaena greggii S.Watson | No | Leguminosae | E | 56° | 2 | P | 887 | 0·9 | 1·8 | 3·6 | 425 | O | —at | FC:EB | |||
| 472 | Leucaena involucrata S.Zarate | No | Leguminosae | E | —n | —p | P | 1,122 | 1·1 | 2·3 | 4·6 | 425 | O | —at | FC:EB | |||
| 473c | Leucaena lanceolata S.Watson | No | Leguminosae | E | 52 | 2 | P | 706 | 0·7 | 1·4 | 2·9 | 425 | O | —at | FC:EB | |||
| 474 | Leucaena lempirana C.E.Hughes | No | Leguminosae | E | —n | —p | P | 426 | 0·4 | 0·9 | 1·7 | 425 | O | —at | FC:EB | |||
| 475b | Leucaena leucocephala (Lam.) De Wit. | No | Leguminosae | E | 104 | 4 | P | 1,455 | 1·5 | 3·0 | 5·9 | 425 | O | —at | FC:EB | |||
| 476 | Leucaena macrophylla Benth. | No | Leguminosae | E | ?52° | —p | P | 299 | 0·3 | 0·6 | 1·2 | 425 | O | —at | FC:EB | |||
| 477 | Leucaena magnifica (C.E.Hughes) C.E.Hughes | No | Leguminosae | E | ?52° | —p | P | 500 | 0·5 | 1·0 | 2·0 | 425 | O | —at | FC:EB | |||
| 478 | Leucaena matudae (S.Zarate) C.E.Hughes | No | Leguminosae | E | ?52° | —p | P | 519 | 0·5 | 1·1 | 2·1 | 425 | O | —at | FC:EB | |||
| 479 | Leucaena multicapitula Schery | No | Leguminosae | E | ?52° | —p | P | 470 | 0·5 | 1·0 | 1·9 | 425 | O | —at | FC:EB | |||
| 480 | Leucaena pallida Britton & Rose | No | Leguminosae | E | 104 (?110/112)° | 4 | P | 774 | 0·8 | 1·6 | 3·2 | 425 | O | —at | FC:EB | |||
| 481 | Leucaena pueblana Britton & Rose | No | Leguminosae | E | —n | —p | P | 490 | 0·5 | 1·0 | 2·0 | 425 | O | —at | FC:EB | |||
| 482 | Leucaena pulverulenta (Schltdl.) Benth. | No | Leguminosae | E | 56 | 2 | P | 686 | 0·7 | 1·4 | 2·8 | 425 | O | —at | FC:EB | |||
| 483 | Leucaena retusa Benth. | No | Leguminosae | E | 56° | 2 | P | 764 | 0·8 | 1·6 | 3·1 | 425 | O | —at | FC:EB | |||
| 484 | Leucaena salvadorensis Standley ex Britton & Rose | No | Leguminosae | E | ?56° | —p | P | 887 | 0·9 | 1·8 | 3·6 | 425 | O | —at | FC:EB | |||
| 485 | Leucaena shannonii J.D.Smith | No | Leguminosae | E | 52° | 2 | P | 691 | 0·7 | 1·4 | 2·8 | 425 | O | —at | FC:EB | |||
| 486 | Leucaena trichandra (Zucc.) Urban | No | Leguminosae | E | 52 (?56)° | 2 | P | 764 | 0·8 | 1·6 | 3·1 | 425 | O | —at | FC:EB | |||
| 487 | Leucaena trichodes (Jacq.) Benth. | No | Leguminosae | E | 52° | 2 | P | 539 | 0·6 | 1·1 | 2·2 | 425 | O | —at | FC:EB | |||
| 488 | Limnanthes douglasii R. Br. | No | Limnanthaceae | E | 10 | 2 | A | 1,362 | 1·4 | 2·8 | 5·6 | 379 | O | J | Fe | |||
| 489 | Litsea glutinosa (Lour.) C.B. Robinson | No | Lauraceae | BA | 48° | —p | P | 2,766 | 2·8 | 5·6 | 11·3 | 454 | O | Bc | Fe | |||
| 490c | Lolium perenne L. | No | Gramineae | M | 14 | 2 | P | 2,773 | 2·8 | 5·7 | 11·3 | 417 | O | Gallusf | FC:PI | |||
| 491 | Loranthus europaeusl | No | Loranthaceaek | E | —n | —p | —q | 8,085 | 8·3 | 16·5 | 33·0 | 457bm | O | Bd | Fe | |||
| 492b | Lupinus angustifolius L. | No | Leguminosae | E | 38, 42, 44 | —p | A | 752.an | 0·8an | 1·5an | 3·1an | 416 | C | —an | FC:PI | |||
| 493 | Lupinus atlanticus Gladst. | No | Leguminosae | E | 38 | —p | A | 1,458.an | 1·5an | 3·0an | 6·0an | 416 | C | —an | FC:PI | |||
| 494 | Lupinus cosentinii Guss. | No | Leguminosae | E | 32 | —p | A | 1,126.an | 1·1an | 2·3an | 4·6an | 416 | C | —an | FC:PI | |||
| 495 | Lupinus digitatus Forssk. | No | Leguminosae | E | 36 | —p | A | 1,286.an | 1·3an | 2·6an | 5·3an | 416 | C | —an | FC:PI | |||
| 496 | Lupinus micranthus Guss. | No | Leguminosae | E | 52 | —p | A | 461.an | 0·5an | 0·9an | 1·9an | 416 | C | —an | FC:PI | |||
| 497 | Lupinus palaestinus Boiss. | No | Leguminosae | E | 42 | —p | A | 1,201.an | 1·2an | 2·5an | 4·9an | 416 | C | —an | FC:PI | |||
| 498c | Lupinus pilosus Murr. | No | Leguminosae | E | 42 | —p | A | 1,201.an | 1·2an | 2·5an | 4·9an | 416 | C | —an | FC:PI | |||
| 499c | Luzula campestris (L.) DC. | No | Juncaceae | M | 12 | —u | P | 1,443 | 1·5 | 2·9 | 5·9 | 420 | O | Bc | Fe | |||
| 500c | Luzula elegans Guthnick | No | Juncaceae | M | 6 | —u | P | 1,512 | 1·5 | 3·1 | 6·2 | 420 | O | Bc | Fe | |||
| 501d | Luzula luzuloides (Lam.) Dandy & Wilmott | No | Juncaceae | M | 12 | —u | P | 1,722.ap | 1·8ap | 3·5ap | 7·0ap | 420 | O | Bc | Fe | |||
| 502d | Luzula nivea Lam. & DC. | No | Juncaceae | M | 12 | —u | P | 1,482.ap | 1·5ap | 3·0ap | 6·1ap | 420 | O | Bc | Fe | |||
| 503c | Luzula pedemontana Boiss & Reut. | No | Juncaceae | M | 30 | —u | P | 1,717 | 1·8 | 3·5 | 7·0 | 420 | O | Bc | Fe | |||
| 504c | Luzula pediformis DC. | No | Juncaceae | M | 12 | —u | P | 1,583 | 1·6 | 3·2 | 6·5 | 420 | O | Bc | Fe | |||
| 505 | Luzula spicata DC. | No | Juncaceae | M | 24 | —u | P | 1,904 | 1·9 | 3·9 | 7·8 | 420 | O | Bc | Fe | |||
| 506 | Luzula sudetica DC. | No | Juncaceae | M | 48 | —u | P | 1,686 | 1·7 | 3·4 | 6·9 | 420 | O | Bc | Fe | |||
| 507k | Lycopersicon esculentum Mill. cv. Gardener's Delight | No | Solanaceae | E | —n | —p | A | 980 | 1·0 | 2·0 | 4·0 | 382 | O | Hc | FC:PI | |||
| 508b | Malva sylvestris L. | —m | Malvaceae | E | 42° | 6 | P | 1,470 | 1·5 | 3·0 | 6·0 | 444bd | O | Gb2 | FC:PI | |||
| 509b | Mangifera indica L. | No | Anacardiaceae | E | 40° | 4 | P | 882 | 0·9 | 1·8 | 3·6 | 454 | O | Bc | Fe | |||
| 510 | Matthiola incanal | No | Cruciferaej | E | —n | —p | —q | 2,597 | 2·7 | 5·3 | 10·6 | 457bm | O | Bd | Fe | |||
| 511a | Melaleuca leucadendra L. | No | Myrtaceae | E | 22° | —p | P | 1,110 | 1·1 | 2·3 | 4·5 | 449bf | O | Bc | Fe | |||
| 512 | Melampyrum arvense Linn. | No | Orobanchaceae | E | —n | —p | P | 8,073 | 8·2 | 16·5 | 33·0 | 465 | O | G | FC:PI | |||
| 513 | Melia azedarach L. | No | Meliaceae | E | 28° | —p | P | 421 | 0·4 | 0·9 | 1·7 | 454 | O | Bc | Fe | |||
| 514 | Melianthus major L. | No | Melianthaceae | E | 36, 38° | 2 or 4 | P | 627 | 0·6 | 1·3 | 2·6 | 378 | O | J | Fe | |||
| 515 | Mentha longifolial | No | Labiatae | E | 24 | 2 | P | 385 | 0·4 | 0·8 | 1·6 | 465 | O | J | Fe | |||
| 516 | Merrilliodendron megacarpum (Hemsl.) Sleum. | No | Icacinaceae | E | 30 | 2 | P | 1,071 | 1·1 | 2·2 | 4·4 | 380 | O | K | FC:PI | |||
| 517 | Mimusops elengi L. | No | Sapotaceae | E | 24° | —p | P | 274 | 0·3 | 0·6 | 1·1 | 454 | O | Bc | Fe | |||
| 518a | Monstera deliciosa Liebm. | No | Araceae | M | 50 | —p | P | 9,384 | 9·6 | 19·2 | 38·3 | 411 | O | Bc | Fe | |||
| 519 | Monstera obliqua Miq. | No | Araceae | M | 44 | —p | P | 8,822 | 9·0 | 18·0 | 36·0 | 411 | O | Bc | Fe | |||
| 520 | Montinia caryophyllacea Thunb. | No | Montiniaceae | E | 24 | 2 | P | 554 | 0·6 | 1·1 | 2·3 | 380 | O | J | Fe | |||
| 521d | Musa acuminata Colla ssp. banksii | —m | Musaceae | M | 22 | 2 | P | 600 | 0·6 | 1·2 | 2·5 | 402 | O | Glycinee | FC:PI | |||
| 521e | Musa acuminata Colla ssp. siamea | —m | Musaceae | M | 22 | 2 | P | 618 | 0·6 | 1·3 | 2·5 | 402 | O | Glycinee | FC:PI | |||
| 521f | Musa acuminata Colla ssp. banksiih | No | Musaceae | M | 22° | 2 | P | 588 | 0·6 | 1·2 | 2·4 | 410 | O | Petuniae | FC:EB | |||
| 521g | Musa acuminata Colla ssp. malaccensis Accession Selangorh | No | Musaceae | M | 22° | 2 | P | 598 | 0·6 | 1·2 | 2·4 | 410 | O | Petuniae | FC:EB | |||
| 521h | Musa acuminata Colla ssp. banksiih | No | Musaceae | M | 22° | 2 | P | 637 | 0·7 | 1·3 | 2·6 | 410 | O | Petuniae | FC:EB | |||
| 521i | Musa acuminata Colla ssp. siamea Accession Siamh | No | Musaceae | M | 22 | 2 | P | 652 | 0·7 | 1·3 | 2·7 | 410 | O | Petuniae | FC:EB | |||
| 521j | Musa acuminata Colla ssp. truncata | —m | Musaceae | M | 22 | 2 | P | 626 | 0·6 | 1·3 | 2·6 | 421aq | O | Glycinee | FC:PI | |||
| 521k | Musa acuminata Colla genotype PPC | —m | Musaceae | M | 22 | 2 | P | 610 | 0·6 | 1·2 | 2·5 | 421aq | O | Glycinee | FC:PI | |||
| 522c | Musa balbisiana Colla | —m | Musaceae | M | 22 | 2 | P | 556 | 0·6 | 1·1 | 2·3 | 421 | O | Glycinee | FC:PI | |||
| 522d | Musa balbisiana Colla | —m | Musaceae | M | 22 | 2 | P | 549 | 0·6 | 1·1 | 2·2 | 402 | O | Glycinee | FC:PI | |||
| 522e | Musa balbisiana Colla Accession Tani (1120) | No | Musaceae | M | 22° | 2 | P | 568 | 0·6 | 1·2 | 2·3 | 410 | O | Petuniae | FC:EB | |||
| 523 | Musa ornata Roxb. | No | Musaceae | M | 22° | 2 | P | 603 | 0·6 | 1·2 | 2·5 | 410 | O | Petuniae | FC:EB | |||
| 524 | Musa violascensl | —m | Musaceae | M | 20 | 2 | P | 691 | 0·7 | 1·4 | 2·8 | 421 | O | Glycinee | FC:PI | |||
| 525 | Muscari adilii M.B. Guner & H. Duman | No | Asparagaceae | M | 18 | 2 | P | 2,550 | 2·6 | 5·2 | 10·4 | 465 | O | G | Fe | |||
| 526 | Muscari mcbeathianuml | No | Asparagaceae | M | 18 | 2 | P | 2,989 | 3·1 | 6·1 | 12·2 | 465 | O | B | Fe | |||
| 527 | Myoporum mauritianum A. DC. | No | Myoporaceae | E | 72 | 8 | P | 1,904 | 1·9 | 3·9 | 7·8 | 379 | O | G | Fe | |||
| 528 | Myrica gale Linn. | No | Myricaceae | E | 16, 48, 80, 96° | —p | P | 409 | 0·4 | 0·8 | 1·7 | 379 | O | J | Fe | |||
| 529 | Myrsine africana L. | No | Myrsinaceae | E | 46 | —p | P | 1,205 | 1·2 | 2·5 | 4·9 | 380 | O | J | Fe | |||
| 530 | Narthecium ossifragum Huds. | No | Nartheciaceae | M | 26° | 2 | P | 404 | 0·4 | 0·8 | 1·7 | 380 | O | J | FC:PI | |||
| 531 | Navarretia squarrosa Hook. & Arn. | No | Polemoniaceae | E | 18 | 2 | A | 1,289 | 1·3 | 2·6 | 5·3 | 378 | O | J | Fe | |||
| 532 | Nelumbo nucifera Gaertn. | No | Nelumbonaceae | E | 16 | 2 | P | 238 | 0·2 | 0·5 | 1·0 | 379 | O | J | Fe | |||
| 533 | Nemophila menziesii Hook. & Arn. | No | Boraginaceae | E | 18 | 2 | A | 1,220 | 1·2 | 2·5 | 5·0 | 465 | O | J | Fe | |||
| 534 | Nepenthes pervillei Blume | No | Nepenthaceae | E | —n | —p | P | 274 | 0·3 | 0·6 | 1·1 | 378 | O | J | Fe | |||
| 535k | Nicotiana tabacum L. | No | Solanaceae | E | 48° | 4 | A | 7,546 | 7·7 | 15·4 | 30·8 | 457bm | O | Bd | Fe | |||
| 536 | Nyctanthes arbor-tristis L. | No | Oleaceae | E | 44° | —p | P | 1,203 | 1·2 | 2·5 | 4·9 | 454 | O | Bc | Fe | |||
| 537 | Odontites luteal | No | Orobanchaceae | E | —n | —p | P | 554 | 0·6 | 1·1 | 2·3 | 465 | O | J | Fe | |||
| 538 | Odontites verna Dum. | No | Orobanchaceae | E | c. 18 | 2 | P | 559 | 0·6 | 1·1 | 2·3 | 465 | O | J | Fe | |||
| 539 | Odontostomum hartwegii Torr. | No | Techophilaeaceae | M | 20 | 2 | P | 2,506 | 2·6 | 5·1 | 10·2 | 380 | O | G | FC:PI | |||
| 540 | Oenothera ammophila Focke | No | Onagraceae | E | 14 | 2 | BP | 1,142 | 1·2 | 2·3 | 4·7 | 439 | O | Sorghume | FC:PI | |||
| 541 | Oenothera biennis L. | No | Onagraceae | E | 14 | 2 | BP | 1,196 | 1·2 | 2·4 | 4·9 | 439 | O | Sorghume | FC:PI | |||
| 542 | Olea africana Mill. | No | Oleaceae | E | 46 | 2 | P | 1,548 | 1·6 | 3·2 | 6·3 | 462 | O | Sorghume | Fe | |||
| 543 | Olea cuspidata Wall. | No | Oleaceae | E | 46 | 2 | P | 2,029 | 2·1 | 4·1 | 8·3 | 462 | O | Sorghume | Fe | |||
| 544a | Olea europaea L. cv. Dolce Agogiah | No | Oleaceae | E | 46 | 2 | P | 1,911 | 2·0 | 3·9 | 7·8 | 462 | O | Sorghume | Fe | |||
| 544b | Olea europaea L. cv. Pendolinoh | No | Oleaceae | E | 46 | 2 | P | 2,283 | 2·3 | 4·7 | 9·3 | 462 | O | Sorghume | Fe | |||
| 545 | Olea ferruginea Royale | No | Oleaceae | E | 46 | 2 | P | 1,813 | 1·9 | 3·7 | 7·4 | 462 | O | Sorghume | Fe | |||
| 546 | Olea indica Klein | No | Oleaceae | E | 46 | 2 | P | 1,646 | 1·7 | 3·4 | 6·7 | 462 | O | Sorghume | Fe | |||
| 547r | Oryza sativa L. ssp. indica | No | Gramineae | M | 24 | 2 | A | 466 | 0·5 | 1·0 | 1·9 | 450bg | O | — | GS | |||
| 547s | Oryza sativa L. ssp. indica cv. IR8 | —m | Gramineae | M | 24 | 2 | A | 440 | 0·4 | 0·9 | 1·8 | 389 | O | Arab.e | FC:PI | |||
| 547t | Oryza sativa L. ssp. japonica cv. Nipponbare | —m | Gramineae | M | 24 | 2 | A | 401 | 0·4 | 0·8 | 1·6 | 389 | O | Arab.e | FC:PI | |||
| 547u | Oryza sativa L. ssp. japonica cv. Nipponbare | No | Gramineae | M | 24 | 2 | A | 420 | 0·4 | 0·9 | 1·7 | 451bh | O | — | GS | |||
| 548 | Oxalis boliviana Britton | No | Oxalidaceae | E | —n | —p | P | 469 | 0·5 | 1·0 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 549 | Oxalis coralleoides R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 521 | 0·5 | 1·1 | 2·1 | 456bk | O | Gallusf | FC:PI | |||
| 550 | Oxalis cuzcensis R.Knuth | No | Oxalidaceae | E | 16° | 2 | AP | 575 | 0·6 | 1·2 | 2·3 | 456bk | O | Gallusf | FC:PI | |||
| 551 | Oxalis herrerae R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 454 | 0·5 | 0·9 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 552 | Oxalis humbertii R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 510 | 0·5 | 1·0 | 2·1 | 456bk | O | Gallusf | FC:PI | |||
| 553 | Oxalis lotoides Kunth | No | Oxalidaceae | E | 16° | 2 | P | 432 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 554a | Oxalis lucumayensis R.Knuth ssp. lucumayensis | No | Oxalidaceae | E | 16° | 2 | P | 455 | 0·5 | 0·9 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 554b | Oxalis lucumayensis R.Knuth ssp. subiens Lourteig | No | Oxalidaceae | E | 16° | 2 | P | 483 | 0·5 | 1·0 | 2·0 | 456bk | O | Gallusf | FC:PI | |||
| 555 | Oxalis marcapatensis R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 440 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 556 | Oxalis medicaginea Kunth | No | Oxalidaceae | E | 16° | 2 | P | 417 | 0·4 | 0·9 | 1·7 | 456bk | O | Gallusf | FC:PI | |||
| 557 | Oxalis megalorrhiza Jacquin | No | Oxalidaceae | E | 14 or 18° | 2 | P | 399 | 0·4 | 0·8 | 1·6 | 456bk | O | Gallusf | FC:PI | |||
| 558 | Oxalis mollis Kunth | No | Oxalidaceae | E | 16° | 2 | P | 437 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 559 | Oxalis oulophora Lourteig | No | Oxalidaceae | E | 16° | 2 | P | 433 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 560 | Oxalis paucartambensis R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 470 | 0·5 | 1·0 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 561a | Oxalis peduncularis Kunthh | No | Oxalidaceae | E | 16° | 2 | P | 454 | 0·5 | 0·9 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 561b | Oxalis peduncularis Kunth var. pilosah | No | Oxalidaceae | E | 16° | 2 | P | 570 | 0·6 | 1·2 | 2·3 | 456bk | O | Gallusf | FC:PI | |||
| 562 | Oxalis petrophila R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 482 | 0·5 | 1·0 | 2·0 | 456bk | O | Gallusf | FC:PI | |||
| 563 | Oxalis phaeotricha Diels | No | Oxalidaceae | E | 32° | 4 | P | 820 | 0·8 | 1·7 | 3·3 | 456bk | O | Gallusf | FC:PI | |||
| 564 | Oxalis picchensis R.Knuth | No | Oxalidaceae | E | 32° | 4 | P | 821 | 0·8 | 1·7 | 3·4 | 456bk | O | Gallusf | FC:PI | |||
| 565a | Oxalis ptychoclada Diels | No | Oxalidaceae | E | 16° | 2 | P | 434 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 565b | Oxalis ptychoclada Diels var. trichocarpa Lourteig | No | Oxalidaceae | E | 16° | 2 | P | 466 | 0·5 | 1·0 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 566 | Oxalis san-miguelii R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 428 | 0·4 | 0·9 | 1·7 | 456bk | O | Gallusf | FC:PI | |||
| 567 | Oxalis sp. cfr. melilotoides Zuccariniy | No | Oxalidaceae | E | 16° | 2 | P | 456 | 0·5 | 0·9 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 568 | Oxalis sp. cfr. teneriensis R. Knuthy | No | Oxalidaceae | E | 16° | 2 | P | 473 | 0·5 | 1·0 | 1·9 | 456bk | O | Gallusf | FC:PI | |||
| 569a | Oxalis spiralis R. & P. ex G.Donh | No | Oxalidaceae | E | 16° | 2 | AP | 520 | 0·5 | 1·1 | 2·1 | 456bk | O | Gallusf | FC:PI | |||
| 569b | Oxalis spiralis R. & P. ex G.Donh | No | Oxalidaceae | E | 16° | 2 | AP | 656 | 0·7 | 1·3 | 2·7 | 456bk | O | Gallusf | FC:PI | |||
| 570 | Oxalis tabaconasensis R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 515 | 0·5 | 1·1 | 2·1 | 456bk | O | Gallusf | FC:PI | |||
| 571 | Oxalis tuberosa Molina | No | Oxalidaceae | E | 64° | 8 | P | 1,722 | 1·8 | 3·5 | 7·0 | 456bl | O | Gallusf | FC:PI | |||
| 572 | Oxalis unduavensis (Rusby) R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 505 | 0·5 | 1·0 | 2·1 | 456bk | O | Gallusf | FC:PI | |||
| 573 | Oxalis urubambensis R.Knuth | No | Oxalidaceae | E | 16° | 2 | P | 431 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 574 | Oxalis vulcanicola Donn. Sm. | No | Oxalidaceae | E | 16° | 2 | P | 434 | 0·4 | 0·9 | 1·8 | 456bk | O | Gallusf | FC:PI | |||
| 575a | Paeonia caucasica (Schipcz.) Schipcz.h | —m | Paeoniaceae | E | 10 | 2 | P | 15,592 | 15·9 | 31·8 | 63·6 | 459 | C | —bq | Fe | |||
| 575b | Paeonia caucasica (Schipcz.) Schipcz.h | —m | Paeoniaceae | E | 10 | 2 | P | 16,087 | 16·4 | 32·8 | 65·7 | 459 | C | —bq | Fe | |||
| 576a | Paeonia daurica Andr.h | —m | Paeoniaceae | E | 10 | 2 | P | 11,804 | 12·0 | 24·1 | 48·2 | 459 | C | —bq | Fe | |||
| 576b | Paeonia daurica Andr.h | —m | Paeoniaceae | E | 10 | 2 | P | 12,970 | 13·2 | 26·5 | 52·9 | 459 | C | —bq | Fe | |||
| 577a | Paeonia lagodechiana Kem.-Nath.h | —m | Paeoniaceae | E | 10 | 2 | P | 11,981 | 12·2 | 24·5 | 48·9 | 459 | C | —bq | Fe | |||
| 577b | Paeonia lagodechiana Kem.-Nath.h | —m | Paeoniaceae | E | 10 | 2 | P | 14,009 | 14·3 | 28·6 | 57·2 | 459 | C | —bq | Fe | |||
| 578a | Paeonia macrophylla (Albov) Lomak.h | —m | Paeoniaceae | E | 20 | 4 | P | 29,449 | 30·1 | 60·1 | 120·2 | 459 | C | —bq | Fe | |||
| 578b | Paeonia macrophylla (Albov) Lomak.h | —m | Paeoniaceae | E | 20 | 4 | P | 30,086 | 30·7 | 61·4 | 122·8 | 459 | C | —bq | Fe | |||
| 579b | Paeonia mlokosewitschi Lomak.h | —m | Paeoniaceae | E | 10 | 2 | P | 16,790 | 17·1 | 34·3 | 68·5 | 459 | C | —bq | Fe | |||
| 579c | Paeonia mlokosewitschi Lomak.h | —m | Paeoniaceae | E | 10 | 2 | P | 17,571 | 17·9 | 35·9 | 71·7 | 459 | C | —bq | Fe | |||
| 580b | Paeonia officinalis L.h | —m | Paeoniaceae | E | 20 | 4 | P | 25,995 | 26·5 | 53·1 | 106·1 | 459 | C | —bq | Fe | |||
| 581a | Paeonia ruprechtiana Kem.-Nath.h | —m | Paeoniaceae | E | 10 | 2 | P | 15,224 | 15·5 | 31·1 | 62·1 | 459 | C | —bq | Fe | |||
| 581b | Paeonia ruprechtiana Kem.-Nath.h | —m | Paeoniaceae | E | 10 | 2 | P | 17,331 | 17·7 | 35·4 | 70·7 | 459 | C | —bq | Fe | |||
| 582a | Paeonia steveniana Kem.-Nath.h | —m | Paeoniaceae | E | 20 | 4 | P | 27,955 | 28·5 | 57·1 | 114·1 | 459 | C | —bq | Fe | |||
| 582b | Paeonia steveniana Kem.-Nath.h | —m | Paeoniaceae | E | 20 | 4 | P | 29,939 | 30·6 | 61·1 | 122·2 | 459 | C | —bq | Fe | |||
| 583b | Paeonia tenuifolia L.h | —m | Paeoniaceae | E | 10 | 2 | P | 7,700 | 7·9 | 15·7 | 31·4 | 459 | C | —bq | Fe | |||
| 583c | Paeonia tenuifolia L.h | —m | Paeoniaceae | E | 10 | 2 | P | 11,598 | 11·8 | 23·7 | 47·3 | 459 | C | —bq | Fe | |||
| 584a | Paeonia tomentosa (Lomak.) N.Buschh | —m | Paeoniaceae | E | 20 | 4 | P | 25,260 | 25·8 | 51·6 | 103·1 | 459 | C | —bq | Fe | |||
| 584b | Paeonia tomentosa (Lomak.) N.Buschh | —m | Paeoniaceae | E | 20 | 4 | P | 27,391 | 28·0 | 55·9 | 111·8 | 459 | C | —bq | Fe | |||
| 585a | Paeonia wittmanniana Hartwiss ex Lindl.h | —m | Paeoniaceae | E | 20 | 4 | P | 27,514 | 28·1 | 56·2 | 112·3 | 459 | C | —bq | Fe | |||
| 585b | Paeonia wittmanniana Hartwiss ex Lindl.h | —m | Paeoniaceae | E | 20 | 4 | P | 31,042 | 31·7 | 63·4 | 126·7 | 459 | C | —bq | Fe | |||
| 586d | Papaver rhoeas L. | No | Papaveraceae | E | 14° | 2 | A | 2,548 | 2·6 | 5·2 | 10·4 | 457bm | O | Bd | Fe | |||
| 587 | Parmentiera cereifera Seem. | No | Bignoniaceae | E | —n | —p | P | 647 | 0·7 | 1·3 | 2·6 | 454 | O | Bc | Fe | |||
| 588 | Paspalum notatum Flugge. | No | Gramineae | M | 20 | 2 | P | 706 | 0·7 | 1·4 | 2·9 | 417 | O | Gallusf | FC:PI | |||
| 589 | Peltophorum pterocarpum (DC.) Baker ex K.Heyne | No | Leguminosae | E | 26° | —p | P | 777 | 0·8 | 1·6 | 3·2 | 454 | O | Bc | Fe | |||
| 590a | Petroselinum crispum cv. Champion Moss Curledl | No | Umbelliferae | E | —n | —p | P | 2,205 | 2·3 | 4·5 | 9·0 | 382 | O | Lycopers.c | FC:PI | |||
| Petunia – Some taxa once included in Petunia are now included in Calibrachoa (see footnote ae) | ||||||||||||||||||
| 591 | Petunia alpicola L.B.Sm. & Downs | No | Solanaceae | E | 18 | 2 | P | 1,450 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 592 | Petunia altiplana T.Ando & Hashim. | No | Solanaceae | E | 14 | 2 | P | 1,274 | 1·3 | 2·6 | 5·2 | 387ae | O | Gallus-398p | FC:PI | |||
| 593b | Petunia axillaris (Lam.) Britton, Sterns & Poggenb. ssp. axillaris | No | Solanaceae | E | 14 | 2 | P | 1,436 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 593c | Petunia axillaris (Lam.) Britton, Sterns & Poggenb. ssp. subandina | No | Solanaceae | E | 14° | 2 | P | 1,465 | 1·5 | 3·0 | 6·0 | 387ae | O | Gallus-398p | FC:PI | |||
| 593d | Petunia axillaris (Lam.) Britton, Sterns & Poggenb. ssp. parodii | No | Solanaceae | E | 14 | 2 | P | 1,470 | 1·5 | 3·0 | 6·0 | 387ae | O | Gallus-398p | FC:PI | |||
| 594 | Petunia bajeensis T.Ando & Hashim. | No | Solanaceae | E | 14° | 2 | P | 1,450 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 595 | Petunia bonjardinensis T.Ando & Hashim. | No | Solanaceae | E | 14 | 2 | P | 1,421 | 1·5 | 2·9 | 5·8 | 387ae | O | Gallus-398p | FC:PI | |||
| 596 | Petunia exserta Stehmann. | No | Solanaceae | E | 14° | 2 | P | 1,539 | 1·6 | 3·1 | 6·3 | 387ae | O | Gallus-398p | FC:PI | |||
| 597 | Petunia guarapuavensis T.Ando & Hashim. | No | Solanaceae | E | 14 | 2 | P | 1,499 | 1·5 | 3·1 | 6·1 | 387ae | O | Gallus-398p | FC:PI | |||
| 598 | Petunia helianthemoides Sendtn. | No | Solanaceae | E | 18 | 2 | P | 1,436 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 599e | Petunia hybrida Vilm. cv. Pearl Sky Blue | No | Solanaceae | E | 14° | 2 | P | 1,441 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 600a | Petunia integrifolia (Hook.) Schinz & Thell. ssp. inflata (R.E.Fr.) | No | Solanaceae | E | 14 | 2 | A | 1,333 | 1·4 | 2·7 | 5·4 | 387ae | O | Gallus-398p | FC:PI | |||
| 600b | Petunia integrifolia (Hook.) Schinz & Thell. ssp. integrifolia var. integrifolia | No | Solanaceae | E | 14 | 2 | P | 1,436 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 600c | Petunia integrifolia (Hook.) Schinz & Thell. ssp. integrifolia var. depauperata (R.E.Fr.) | No | Solanaceae | E | 14 | 2 | P | 1,490 | 1·5 | 3·0 | 6·1 | 387ae | O | Gallus-398p | FC:PI | |||
| 601 | Petunia interior T.Ando & Hashim. | No | Solanaceae | E | 14 | 2 | P | 1,455 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 602 | Petunia kleinii L.B.Sm. & Downs | No | Solanaceae | E | 18 | 2 | P | 1,436 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 603 | Petunia littoralis L.B. Sm. & Downs | No | Solanaceae | E | 14 | 2 | P | 1,455 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 604 | Petunia mantiqueirensis T.Ando & Hashim. | No | Solanaceae | E | 14° | 2 | P | 1,524 | 1·6 | 3·1 | 6·2 | 387ae | O | Gallus-398p | FC:PI | |||
| 605 | Petunia occidentalis R.E.Fr. | No | Solanaceae | E | 14 | 2 | A | 1,362 | 1·4 | 2·8 | 5·6 | 387ae | O | Gallus-398p | FC:PI | |||
| 606 | Petunia pubescens (Spreng.) R.E.Fr. | No | Solanaceae | E | 18 | 2 | P | 1,446 | 1·5 | 3·0 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 607 | Petunia reitzii L.B.Sm. & Downs. | No | Solanaceae | E | 14 | 2 | P | 1,406 | 1·4 | 2·9 | 5·7 | 387ae | O | Gallus-398p | FC:PI | |||
| 608 | Petunia riograndensis T.Ando & Hashim. | No | Solanaceae | E | 14° | 2 | P | 1,460 | 1·5 | 3·0 | 6·0 | 387ae | O | Gallus-398p | FC:PI | |||
| 609 | Petunia saxicola L.B.Sm. & Downs | No | Solanaceae | E | 14 | 2 | P | 1,411 | 1·4 | 2·9 | 5·8 | 387ae | O | Gallus-398p | FC:PI | |||
| 610 | Petunia scheideana L.B.Sm. & Downs | No | Solanaceae | E | 14 | 2 | P | 1,436 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 611 | Petunia variabilisR.E.Fr. | No | Solanaceae | E | 18 | 2 | P | 1,441 | 1·5 | 2·9 | 5·9 | 387ae | O | Gallus-398p | FC:PI | |||
| 612 | Phalaenopsis amboinensis J. J. Smith | No | Orchidaceae | M | 38 | 2 | P | 7,036 | 7·2 | 14·4 | 28·7 | 447 | O | Gb2 | FC:PI | |||
| 613 | Phalaenopsis aphrodite Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 1,372 | 1·4 | 2·8 | 5·6 | 447 | O | Gb2 | FC:PI | |||
| 614 | Phalaenopsis bellina (Rchb. f.) Cristenson | No | Orchidaceae | M | 38 | 2 | P | 7,365 | 7·5 | 15·0 | 30·1 | 447 | O | Gb2 | FC:PI | |||
| 615 | Phalaenopsis cornu-cervi (Breda) Bl & Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 3,156 | 3·2 | 6·4 | 12·9 | 447 | O | Gb2 | FC:PI | |||
| 616a | Phalaenopsis equestris (Schauer) Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 1,651 | 1·7 | 3·4 | 6·7 | 447 | O | Gb2 | FC:PI | |||
| 617 | Phalaenopsis fasciata Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 3,214 | 3·3 | 6·6 | 13·1 | 447 | O | Gb2 | FC:PI | |||
| 618 | Phalaenopsis gigantea J.J.Smith | No | Orchidaceae | M | 38 | 2 | P | 2,587 | 2·6 | 5·3 | 10·6 | 447 | O | Gb2 | FC:PI | |||
| 619a | Phalaenopsis lueddemanniana Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 3,180 | 3·2 | 6·5 | 13·0 | 447 | O | Gb2 | FC:PI | |||
| 620 | Phalaenopsis mannii Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 6,615 | 6·8 | 13·5 | 27·0 | 447 | O | Gb2 | FC:PI | |||
| 621 | Phalaenopsis mariae Burb. ex Warn. & Wms. | No | Orchidaceae | M | 38 | 2 | P | 3,175 | 3·2 | 6·5 | 13·0 | 447 | O | Gb2 | FC:PI | |||
| 622 | Phalaenopsis micholitzii Rolfe | No | Orchidaceae | M | 38 | 2 | P | 3,180 | 3·2 | 6·5 | 13·0 | 447 | O | Gb2 | FC:PI | |||
| 623 | Phalaenopsis modesta J.J.Smith | No | Orchidaceae | M | 38 | 2 | P | 2,524 | 2·6 | 5·2 | 10·3 | 447 | O | Gb2 | FC:PI | |||
| 624 | Phalaenopsis parishii Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 8,139 | 8·3 | 16·6 | 33·2 | 447 | O | Gb2 | FC:PI | |||
| 625 | Phalaenopsis pulchra (Rchb.f.) Sweet | No | Orchidaceae | M | 38 | 2 | P | 3,121 | 3·2 | 6·4 | 12·7 | 447 | O | Gb2 | FC:PI | |||
| 626 | Phalaenopsis sanderiana Rchb.f.i | No | Orchidaceae | M | 38 | 2 | P | 1,372 | 1·4 | 2·7 | 5·6 | 447 | O | Gb2 | FC:PI | |||
| 627 | Phalaenopsis stuartiana Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 1,534 | 1·6 | 3·1 | 6·3 | 447 | O | Gb2 | FC:PI | |||
| 628 | Phalaenopsis sumatrana Korth. & Rchb.f. | No | Orchidaceae | M | 38 | 2 | P | 3,244 | 3·3 | 6·6 | 13·2 | 447 | O | Gb2 | FC:PI | |||
| 629 | Phalaenopsis venosa Shim & Fowl. | No | Orchidaceae | M | 38 | 2 | P | 4,665 | 4·8 | 9·5 | 19·0 | 447 | O | Gb2 | FC:PI | |||
| 630c | Phaseolus acutifolius var. latifolius G.Freeman | —m | Leguminosae | E | 22° | 2 | A | 794 | 0·8 | 1·6 | 3·2 | 390 | O | Petuniae | FC:DAPI | |||
| 630d | Phaseolus acutifolius var. tenuifolius (Wood & Standl) A.Gray | —m | Leguminosae | E | 22° | 2 | A | 799 | 0·8 | 1·6 | 3·3 | 390 | O | Petuniae | FC:DAPI | |||
| 630e | Phaseolus acutifolius var. acutifolius A.Gray | —m | Leguminosae | E | 22° | 2 | A | 862 | 0·9 | 1·8 | 3·5 | 390 | O | Petuniae | FC:DAPI | |||
| 631 | Phaseolus angustissimus A.Gray | —m | Leguminosae | E | 22° | 2 | —q | 647 | 0·7 | 1·3 | 2·6 | 390 | O | Petuniae | FC:DAPI | |||
| 632e | Phaseolus coccineus L. | —m | Leguminosae | E | 22° | 2 | P | 784 | 0·8 | 1·6 | 3·2 | 390 | O | Petuniae | FC:DAPI | |||
| 632f | Phaseolus coccineus L. ssp. Purpurascens | —m | Leguminosae | E | 22° | 2 | P | 794 | 0·8 | 1·6 | 3·2 | 390 | O | Petuniae | FC:DAPI | |||
| 632g | Phaseolus coccineus L. ssp. coccineus cv. Hammond's Dwarf Scarleth | —m | Leguminosae | E | 22° | 2 | P | 794 | 0·8 | 1·6 | 3·2 | 390 | O | Petuniae | FC:DAPI | |||
| 632h | Phaseolus coccineus L. ssp. coccineus cv. Preisgewinnerh | —m | Leguminosae | E | 22° | 2 | P | 809 | 0·8 | 1·7 | 3·3 | 390 | O | Petuniae | FC:DAPI | |||
| 632i | Phaseolus coccineus L. | No | Leguminosaej | E | 22° | 2 | P | 1,715 | 1·8 | 3·5 | 7·0 | 457bm | O | Bd | Fe | |||
| 633b | Phaseolus filiformis Benth. | —m | Leguminosae | E | 22° | 2 | P | 691 | 0·7 | 1·4 | 2·8 | 390 | O | Petuniae | FC:DAPI | |||
| 634b | Phaseolus glabellus Piper | —m | Leguminosae | E | 22° | 2 | P | 1,024 | 1·0 | 2·1 | 4·2 | 390 | O | Petuniae | FC:DAPI | |||
| 635 | Phaseolus grayanus Wood. & Standl | —m | Leguminosae | E | 22° | 2 | —q | 931 | 1·0 | 1·9 | 3·8 | 390 | O | Petuniae | FC:DAPI | |||
| 636b | Phaseolus hintonii Delgado | —m | Leguminosae | E | 22° | 2 | P | 715 | 0·7 | 1·5 | 2·9 | 390 | O | Petuniae | FC:DAPI | |||
| 637 | Phaseolus leptostachys var. leptostachys Benth. | —m | Leguminosae | E | 22° | 2 | —q | 613 | 0·6 | 1·3 | 2·5 | 390 | O | Petuniae | FC:DAPI | |||
| 638d | Phaseolus lunatus L. var. lunatus cv. Early Thorogreenh | —m | Leguminosae | E | 22° | 2 | P | 691 | 0·7 | 1·4 | 2·8 | 390 | O | Petuniae | FC:DAPI | |||
| 638e | Phaseolus lunatus L. var. silvester Baudet | —m | Leguminosae | E | 22° | 2 | P | 696 | 0·7 | 1·4 | 2·8 | 390 | O | Petuniae | FC:DAPI | |||
| 638f | Phaseolus lunatus L. var. lunatus cv. Henderson Bushh | —m | Leguminosae | E | 22° | 2 | P | 701 | 0·7 | 1·4 | 2·9 | 390 | O | Petuniae | FC:DAPI | |||
| 639b | Phaseolus marechalli Delgado | —m | Leguminosae | E | 22° | 2 | P | 784 | 0·8 | 1·6 | 3·2 | 390 | O | Petuniae | FC:DAPI | |||
| 640 | Phaseolus micranthus Hook. & Arn. | —m | Leguminosae | E | 22° | 2 | —q | 588 | 0·6 | 1·2 | 2·4 | 390 | O | Petuniae | FC:DAPI | |||
| 641 | Phaseolus microcarpus Mart. | —m | Leguminosae | E | 22° | 2 | —q | 505 | 0·5 | 1·0 | 2·1 | 390 | O | Petuniae | FC:DAPI | |||
| 642b | Phaseolus neglectus Herm. | —m | Leguminosae | E | 22° | 2 | —q | 941 | 1·0 | 1·9 | 3·8 | 390 | O | Petuniae | FC:DAPI | |||
| 643 | Phaseolus parviflorus G.Freytag | —m | Leguminosae | E | 22° | 2 | —q | 637 | 0·7 | 1·3 | 2·6 | 390 | O | Petuniae | FC:DAPI | |||
| 644b | Phaseolus pluriflorus Marechal | —m | Leguminosae | E | 22° | 2 | —q | 1,068 | 1·1 | 2·2 | 4·4 | 390 | O | Petuniae | FC:DAPI | |||
| 645b | Phaseolus polyanthus Greenm. | —m | Leguminosae | E | 22° | 2 | P | 799 | 0·8 | 1·6 | 3·3 | 390 | O | Petuniae | FC:DAPI | |||
| 646g | Phaseolus vulgaris L. cv. Kentucky Wonderh | —m | Leguminosae | E | 22° | 2 | A | 686 | 0·7 | 1·4 | 2·8 | 390 | O | Petuniae | FC:DAPI | |||
| 646h | Phaseolus vulgaris L. var. aborigineus (Burk.) Baudet | —m | Leguminosae | E | 22° | 2 | A | 720 | 0·7 | 1·5 | 2·9 | 390 | O | Petuniae | FC:DAPI | |||
| 646i | Phaseolus vulgaris L. var. mexicanus | —m | Leguminosae | E | 22° | 2 | A | 735 | 0·8 | 1·5 | 3·0 | 390 | O | Petuniae | FC:DAPI | |||
| 646j | Phaseolus vulgaris L. cv. Sanilac | —m | Leguminosae | E | 22° | 2 | A | 750 | 0·8 | 1·5 | 3·1 | 390 | O | Petuniae | FC:DAPI | |||
| 646k | Phaseolus vulgaris L. | No | Leguminosaej | E | 22° | 2 | A | 1,666.bn | 1·7bn | 3·4bn | 6·8bn | 457bm | O | Bd | Fe | |||
| 647b | Phaseolus xanthotrichus Piper var. xanthotrichus | —m | Leguminosae | E | 22° | 2 | P | 662 | 0·7 | 1·4 | 2·7 | 390 | O | Petuniae | FC:DAPI | |||
| 647c | Phaseolus xanthotrichus Piper | —m | Leguminosae | E | 22° | 2 | P | 848 | 0·9 | 1·7 | 3·5 | 390 | O | Petuniae | FC:DAPI | |||
| 648 | Philodendron erubescens C.Koch & Bousche | No | Araceae | M | 42 | —p | P | 5,174 | 5·3 | 10·6 | 21·1 | 411 | O | Bc | Fe | |||
| 649 | Philodendron selloum C.Koch | No | Araceae | M | 36 | —p | P | 4,895 | 5·0 | 10·0 | 20·0 | 411 | O | Bc | Fe | |||
| 650 | Philodendron squamiferum Poepp. & Endl. | No | Araceae | M | 30 | —p | P | 4,557 | 4·7 | 9·3 | 18·6 | 411 | O | Bc | Fe | |||
| 651 | Phormium tenaxl | No | Hemerocallidaceae | M | 32 | 2 | P | 740 | 0·8 | 1·5 | 3·0 | 379 | O | J | Fe | |||
| 652 | Pinguicula primuliflora C.E.Wood & Godfrey | No | Lentibulariaceae | E | 22 | 2 | P | 669 | 0·7 | 1·4 | 2·7 | 378 | O | J | Fe | |||
| 653 | Piptocalyx moorei Oliver | No | Trimeniaceae | BA | 16° | 2 | P | 4,001 | 4·1 | 8·2 | 16·3 | 381 | O | G | FC:PI | |||
| 654b | Pistia stratiotes L. | No | Araceaek | M | 28 | 2 | P | 250 | 0·3 | 0·5 | 1·0 | 400 | O | G | Fe | |||
| 655c | Pisum abyssinicum A.Braunh | No | Leguminosae | E | 14 | 2 | A | 4,371 | 4·5 | 8·9 | 17·8 | 458bp | Cbp | Gc | FC:EB | |||
| 655d | Pisum abyssinicum A.Braunh | No | Leguminosae | E | 14 | 2 | A | 4,751 | 4·8 | 9·7 | 19·4 | 458bp | Cbp | Gc | FC:EB | |||
| 656c | Pisum elatius Steven ex M.Bieb.h | No | Leguminosae | E | 14 | 2 | A | 4,231 | 4·3 | 8·6 | 17·3 | 458bp | Cbp | Gc | FC:EB | |||
| 656d | Pisum elatius Steven ex M.Bieb.h | No | Leguminosae | E | 14 | 2 | A | 4,978 | 5·1 | 10·2 | 20·3 | 458bp | Cbp | Gc | FC:EB | |||
| 657b | Pisum fulvum Sibth. & Smith | No | Leguminosae | E | 14 | 2 | A | 4,716 | 4·8 | 9·6 | 19·3 | 458bp | Cbp | Gc | FC:EB | |||
| 658c | Pisum humile Boiss & Noëh | No | Leguminosae | E | 14 | 2 | A | 4,258 | 4·3 | 8·7 | 17·4 | 458bp | Cbp | Gc | FC:EB | |||
| 658d | Pisum humile Boiss & Noëh | No | Leguminosae | E | 14 | 2 | A | 4,809 | 4·9 | 9·8 | 19·6 | 458bp | Cbp | Gc | FC:EB | |||
| 659 | Pittosporum tenuifolium Gaertn. | No | Pittosporaceae | E | 24 | 2 | P | 453 | 0·5 | 0·9 | 1·9 | 379 | O | G | Fe | |||
| 660 | Planchonella eerwah (F.M. Bailey) van Royen | No | Sapotaceae | E | c. 24 | 2 | P | 527 | 0·5 | 1·1 | 2·2 | 380 | O | J | Fe | |||
| 661 | Plantago afra L. | No | Plantaginaceae | E | 12 | 2 | A | 1,129 | 1·2 | 2·3 | 4·6 | 388 | O | Bc | Fe | |||
| 662 | Plantago arenaria W. & K. | No | Plantaginaceae | E | 12 | 2 | A | 1,115 | 1·1 | 2·3 | 4·6 | 388 | O | Bc | Fe | |||
| 663 | Plantago coronopus L. | No | Plantaginaceae | E | 10 | 2 | AP | 845 | 0·9 | 1·7 | 3·5 | 388 | O | Bc | Fe | |||
| 664 | Plantago indica L. | No | Plantaginaceae | E | 12 | 2 | —q | 1,088 | 1·1 | 2·2 | 4·4 | 388 | O | Bc | Fe | |||
| 665b | Plantago lagopus L. | No | Plantaginaceae | E | 12 | 2 | —q | 1,046 | 1·1 | 2·1 | 4·3 | 388 | O | Bc | Fe | |||
| 666c | Plantago lanceolata L. | No | Plantaginaceae | E | 12 | 2 | P | 1,299 | 1·3 | 2·7 | 5·3 | 388 | O | Bc | Fe | |||
| 667c | Plantago major L. | No | Plantaginaceae | E | 12 | 2 | P | 867 | 0·9 | 1·8 | 3·5 | 388 | O | Bc | Fe | |||
| 668 | Plantago psyllium L. | No | Plantaginaceae | E | 12 | 2 | —q | 1,142 | 1·2 | 2·3 | 4·7 | 388 | O | Bc | Fe | |||
| 669 | Plantago serraria L. | No | Plantaginaceae | E | 10 | 2 | —q | 882 | 0·9 | 1·8 | 3·6 | 388 | O | Bc | Fe | |||
| 670 | Plantago stepposa K. | No | Plantaginaceae | E | 24 | 4 | —q | 1,593 | 1·6 | 3·3 | 6·5 | 388 | O | Bc | Fe | |||
| 671 | Platanus orientalis L. | No | Platanaceae | E | 42 | 2 | P | 1,274 | 1·3 | 2·6 | 5·2 | 379 | O | J | Fe | |||
| 672 | Poa pratensis L. | No | Gramineae | M | 58-62 | —p | P | 4,155 | 4·2 | 8·5 | 17·0 | 417 | O | Gallusf | FC:PI | |||
| 673 | Poncirus trifoliata (L.) Raf. | No | Rutaceae | E | 18° | 2 | P | 377 | 0·4 | 0·8 | 1·5 | 426 | O | Gallusf | FC:PI | |||
| 674 | Prosopis cineraria (L.) Druce | No | Leguminosae | E | 52° | —p | P | 1,252 | 1·3 | 2·6 | 5·1 | 454 | O | Bc | Fe | |||
| 675 | Protium serratum (Wall. ex Colebr.) | No | Burseraceae | E | —n | —p | P | 924 | 0·9 | 1·9 | 3·8 | 454 | O | Bc | Fe | |||
| 676 | Pterospermum lanceifolium Roxb. | No | Malvaceae | E | 38° | —p | P | 786 | 0·8 | 1·6 | 3·2 | 454 | O | Bc | Fe | |||
| 677 | Pterostyrax psilophylla Diels ex Perkins | No | Styracaceae | E | 24 | 2 | P | 867 | 0·9 | 1·8 | 3·5 | 380 | O | J | Fe | |||
| 678 | Punica granatum L. | No | Sonneratiaceae | E | 16° | 2 | P | 706 | 0·7 | 1·4 | 2·9 | 454 | O | Bc | Fe | |||
| 679 | Reseda luteola L. | No | Resedaceae | E | 26 | 2 | B | 500 | 0·5 | 1·0 | 2·0 | 378 | O | J | Fe | |||
| 680 | Rhapidophora montana Schott | No | Araceae | M | 30 | —p | P | 9,829 | 10·0 | 20·1 | 40·1 | 411 | O | Bc | Fe | |||
| 681 | Rhapidophora peepla Schott | No | Araceae | M | 18 | —p | P | 8,984 | 9·2 | 18·3 | 36·7 | 411 | O | Bc | Fe | |||
| 682 | Rhipogonum papuanum C.T. White | No | Rhipogonaceae | M | 30 | 2 | P | 10,922 | 11·1 | 22·3 | 44·6 | 380 | O | G | Fe | |||
| 683 | Rhodocoma gigantea (Kunth) H.P. Linder | No | Restionaceae | M | —n | —p | P | 728 | 0·7 | 1·5 | 3·0 | 380 | O | J | FC:PI | |||
| 684 | Rhodohypoxis milloides (Baker) Hilliard & B.L. Burtt | No | Hypoxidaceae | M | 24 + 1-2B | 4 | P | 1,394 | 1·4 | 2·8 | 5·7 | 379 | O | J | Fe | |||
| 685c | Rhoeo discolor Hance | No | Commelinaceae | M | 12° | 2 | P | 7,987 | 8·2 | 16·3 | 32·6 | 457bm | O | Bd | Fe | |||
| 686 | Rhoiacarpos capensis A. DC. | No | Santalaceae | E | —n | —p | P | 304 | 0·3 | 0·6 | 1·2 | 379 | O | J | Fe | |||
| 687 | Rhynchosia cyanosperma Benth. Ex Baker | No | Leguminosae | E | 22 | 2 | B | 2,727 | 2·8 | 5·6 | 11·1 | 443bc | O | Bc | Fe | |||
| 688 | Rhynchosia minima (L.) DC. | No | Leguminosae | E | 22 | 2 | P | 1,227 | 1·3 | 2·5 | 5·0 | 443bc | O | Bc | Fe | |||
| 689 | Ribes glutinosuml | No | Grossulariaceae | E | 16 | 2 | P | 534 | 0·5 | 1·1 | 2·2 | 379 | O | J | Fe | |||
| 690 | Roridula gorgonias Planch. | No | Roridulaceae | E | 12 | 2 | P | 186 | 0·2 | 0·4 | 0·8 | 379 | O | G | Fe | |||
| 691c | Ruta graveolens L. | No | Rutaceae | E | —n | —p | P | 735 | 0·8 | 1·5 | 3·0 | 457bm | O | Bd | Fe | |||
| 692 | Salix alba L.h | —m | Salicaceae | E | 76ad | 4ad | P | 809 | 0·8 | 1·7 | 3·3 | 385ac | O | Lycopers.c | FC:PI | |||
| 693 | Salix atrocinerea Brot.h | —m | Salicaceae | E | 76ad | 4ad | P | 804 | 0·8 | 1·6 | 3·3 | 385ac | O | Lycopers.c | FC:PI | |||
| 694b | Salix caprea L.h | —m | Salicaceae | E | 38ad | 2ad | P | 470 | 0·5 | 1·0 | 1·9 | 385ac | O | Lycopers.c | FC:PI | |||
| 695 | Salix cinerea L.h | —m | Salicaceae | E | 76ad | 4ad | P | 828 | 0·8 | 1·7 | 3·4 | 385ac | O | Lycopers.c | FC:PI | |||
| 696 | Salix elaeagnos Scop.h | —m | Salicaceae | E | 38ad | 2ad | P | 417 | 0·4 | 0·9 | 1·7 | 385ac | O | Lycopers.c | FC:PI | |||
| 697 | Salix fragilis L.h | —m | Salicaceae | E | 76ad | 4ad | P | 843 | 0·9 | 1·7 | 3·4 | 385ac | O | Lycopers.c | FC:PI | |||
| 698 | Salix purpurea L.h | —m | Salicaceae | E | 38ad | 2ad | P | 461 | 0·5 | 0·9 | 1·9 | 385ac | O | Lycopers.c | FC:PI | |||
| 699 | Salix pyrenaica Gouanh | —m | Salicaceae | E | 38ad | 2ad | P | 470 | 0·5 | 1·0 | 1·9 | 385ac | O | Lycopers.c | FC:PI | |||
| 700 | Salix triandra L.h | —m | Salicaceae | E | 38ad | 2ad | P | 387 | 0·4 | 0·8 | 1·6 | 385ac | O | Lycopers.c | FC:PI | |||
| 701 | Salix viminalis L.h | —m | Salicaceae | E | 38ad | 2ad | P | 402 | 0·4 | 0·8 | 1·6 | 385ac | O | Lycopers.c | FC:PI | |||
| 702 | Salix viminalis L.h | —m | Salicaceae | E | 76ad | 4ad | P | 794 | 0·8 | 1·6 | 3·2 | 385ac | O | Lycopers.c | FC:PI | |||
| 703a | Sambucus nigra L. | No | Adoxaceaek | E | 36° | 2 | P | 14,945.bo | 15·3bo | 30·5bo | 61·0bo | 457bm | O | Bd | Fe | |||
| 704 | Santalum album L. | No | Santalaceae | E | 20° | —p | P | 282 | 0·3 | 0·6 | 1·2 | 454 | O | Bc | Fe | |||
| 705b | Saxifraga granulata L. ssp. granulata | No | Saxifragaceae | E | 22 | —p | P | 662 | 0·7 | 1·4 | 2·7 | 453 | O | Gc | FC:EB | |||
| 706 | Saxifraga granulata L. | No | Saxifragaceae | E | —bj | —p | P | 1,122 | 1·1 | 2·3 | 4·6 | 453 | O | Gc | FC:EB | |||
| 707a | Saxifraga granulata L. | No | Saxifragaceae | E | 52 | —p | P | 2,332 | 2·4 | 4·8 | 9·5 | 453 | O | Gc | FC:EB | |||
| 707b | Saxifraga granulata L. ssp. fernandesii Redondo & Horjalesi | No | Saxifragaceae | E | 44-56 | —p | P | 1,735 | 1·8 | 3·5 | 7·1 | 453bi | O | Gc | FC:EB | |||
| 708 | Schisandra rubrifloral | No | Schisandraceae | BA | —n | —p | P | 8,938 | 9·1 | 18·2 | 36·5 | 381 | O | G | FC:PI | |||
| 709 | Schleichera oleosa (Lour.) Oken | No | Sapindaceae | E | 32° | —p | P | 1,142 | 1·2 | 2·3 | 4·7 | 454 | O | Bc | Fe | |||
| 710b | Scilla indica (Roxb.) Baker (cytotype II) | No | Asparagaceae | M | 30 | —p | P | 3,504 | 3·6 | 7·2 | 14·3 | 422ar | O | Bc | Fe | |||
| 710c | Scilla indica (Roxb.) Baker (cytotype I)h | No | Asparagaceae | M | 30 | —p | P | 5,701 | 5·8 | 11·6 | 23·3 | 422ar | O | Bc | Fe | |||
| 711 | Scilla nervosa (Burch.) J.P.Jessop | No | Asparagaceae | M | 38 | —p | P | 3,964 | 4·0 | 8·1 | 16·2 | 422 | O | Bc | Fe | |||
| 712f | Scilla siberica Haw. in Andr. | No | Asparagaceae | M | 12 | 2 | P | 30,135 | 30·8 | 61·5 | 123·0 | 422 | O | Bc | Fe | |||
| 713 | Scilla talosii D.Tzanoudakis & Kypriotakis | No | Asparagaceae | M | c. 150 | —p | P | 45,840 | 46·8 | 93·6 | 187·1 | 465 | O | B | Fe | |||
| 714c | Scilla vindobonensis Speta | No | Asparagaceae | M | 18 | 3 | P | —t | —t | 17·9 | 35·7 | 422 | O | Bc | Fe | |||
| 715 | Scindapsus pictus Hassk | No | Araceae | M | 60 | —p | P | 11,517 | 11·8 | 23·5 | 47·0 | 411 | O | Bc | Fe | |||
| 716 | Sedum acre L. | No | Crassulaceae | E | —n | —p | P | 1,225 | 1·3 | 2·5 | 5·0 | 457bm | O | Bd | Fe | |||
| 717 | Sedum album L. | —m | Crassulaceae | E | 34 | 2 | P | 142 | 0·1 | 0·3 | 0·6 | 398 | O | —w | —r | |||
| 718 | Sedum forsterianum Sm. | —m | Crassulaceae | E | 24 | 2 | P | 451 | 0·5 | 0·9 | 1·8 | 398 | O | —w | —r | |||
| 719 | Sedum montanum Song. & Perrier | —m | Crassulaceae | E | 34 | 2 | P | 515 | 0·5 | 1·1 | 2·1 | 398 | O | —w | —r | |||
| 720a | Sedum obtusifolium C.A.Meyer | —m | Crassulaceae | E | 12 | 2 | P | 206 | 0·2 | 0·4 | 0·8 | 398 | O | —w | —r | |||
| 720b | Sedum obtusifolium C.A.Meyer | —m | Crassulaceae | E | 12 | 2 | P | 206 | 0·2 | 0·4 | 0·8 | 399 | O | B-723b | Fe | |||
| 721 | Sedum obtusifolium C.A.Meyer | —m | Crassulaceae | E | 30 | 5 | P | —t | —t | 1·7 | 3·4 | 399 | O | B-723b | Fe | |||
| 722 | Sedum ochroleucum Chaix | —m | Crassulaceae | E | 34 | 2 | P | 446 | 0·5 | 0·9 | 1·8 | 398 | O | —w | —r | |||
| 723a | Sedum rupestre L. ssp. erectum | —m | Crassulaceae | E | 64 | 4 | P | 1,014 | 1·0 | 2·1 | 4·1 | 398 | O | —w | —r | |||
| 723b | Sedum rupestre L. ssp. rupestre | —m | Crassulaceae | E | —n | —p | P | 2,244 | 2·3 | 4·6 | 9·2 | 399 | O | B | Fe | |||
| 724 | Sedum sediforme (Jacq.) Pau | —m | Crassulaceae | E | 32 | 2 | P | 568 | 0·6 | 1·2 | 2·3 | 398 | O | —w | —r | |||
| 725 | Sedum spurium Bieb. | —m | Crassulaceae | E | 28 | 4 | P | 1,735 | 1·8 | 3·5 | 7·1 | 399 | O | B-723b | Fe | |||
| 726 | Sedum spurium Bieb. | —m | Crassulaceae | E | 42 | 6 | P | 2,764 | 2·8 | 5·6 | 11·3 | 399 | O | B-723b | Fe | |||
| 727a | Sedum stellatum L. | —m | Crassulaceae | E | 10 | 2 | P | 289 | 0·3 | 0·6 | 1·2 | 399 | O | B-723b | Fe | |||
| 727b | Sedum stellatum L. | —m | Crassulaceae | E | 10 | 2 | P | 289 | 0·3 | 0·6 | 1·2 | 398 | O | —w | —r | |||
| 728a | Sedum stoloniferum S.G.Gmelin | —m | Crassulaceae | E | 14 | 2 | P | 309 | 0·3 | 0·6 | 1·3 | 399 | O | B-723b | Fe | |||
| 728b | Sedum stoloniferum S.G.Gmelin | —m | Crassulaceae | E | 14 | 2 | P | 309 | 0·3 | 0·6 | 1·3 | 398 | O | —w | —r | |||
| 729 | Senecio viscosus L. | No | Compositaej | E | —n | —p | A | 1,519 | 1·6 | 3·1 | 6·2 | 457bm | O | Bd | Fe | |||
| 730 | Sesamum alatum Thonn. | No | Pedaliaceae | E | 26 | 2 | A | 1,651 | 1·7 | 3·4 | 6·7 | 446 | O | G | Fe | |||
| 731 | Sesamum capense Burm. | No | Pedaliaceae | E | 26 | 2 | A | 1,188 | 1·2 | 2·4 | 4·9 | 446 | O | G | Fe | |||
| 732 | Sesamum indicum L. | No | Pedaliaceae | E | 26 | 2 | A | 951 | 1·0 | 1·9 | 3·9 | 446 | O | G | Fe | |||
| 733 | Sesamum laciniatum Klein. | No | Pedaliaceae | E | 32 | 4 | A | 1,154 | 1·2 | 2·4 | 4·7 | 446 | O | G | Fe | |||
| 734 | Sesamum latifolium Gillett. | No | Pedaliaceae | E | 32 | 4 | A | 933 | 1·0 | 1·9 | 3·8 | 446 | O | G | Fe | |||
| 735 | Sesamum mulayanum Nair. | No | Pedaliaceae | E | 26 | 2 | A | 870 | 0·9 | 1·8 | 3·6 | 446 | O | G | Fe | |||
| 736 | Sesamum occidentale Regel. | No | Pedaliaceae | E | 64 | 8 | A | 1,551 | 1·6 | 3·2 | 6·3 | 446 | O | G | Fe | |||
| 737 | Sesamum radiatum Schumach. | No | Pedaliaceae | E | 64 | 8 | A | 1,306 | 1·3 | 2·7 | 5·3 | 446 | O | G | Fe | |||
| 738 | Sesamum schinzianum Aschers. | No | Pedaliaceae | E | 64 | 8 | A | 1,343 | 1·4 | 2·7 | 5·5 | 446 | O | G | Fe | |||
| 739 | Sesamum triphyllum Welw. ex Aschers. | No | Pedaliaceae | E | c. 26 | 2 | A | 524 | 0·5 | 1·1 | 2·1 | 378 | O | J | Fe | |||
| 740b | Sesleria albicans Kit. ex Schult.h | No | Gramineaej | M | 28° | 4 | P | 4,748 | 4·8 | 9·7 | 19·4 | 428 | O | Homof | FC:PI | |||
| 740c | Sesleria albicans Kit. ex Schult.h | No | Gramineaej | M | 28° | 4 | P | 4,827 | 4·9 | 9·9 | 19·7 | 428 | O | Homof | FC:PI | |||
| 741 | Severinia buxifolia (Poir.) Ten. | No | Rutaceae | E | 18° | 2 | P | 328 | 0·3 | 0·7 | 1·3 | 426 | O | Gallusf | FC:PI | |||
| 742 | Silene chalcedonica L. | No | Caryophyllaceae | E | 24 | 2 | P | 3,229 | 3·3 | 6·6 | 13·2 | 437 | O | Lycopers.c | FC:PI | |||
| 743c | Silene latifolia Poiret (female) | No | Caryophyllaceae | E | 24 | 2 | AP | 2,808 | 2·9 | 5·7 | 11·5 | 437az | O | Lycopers.c | FC:PI | |||
| 743d | Silene latifolia Poiret (male) | No | Caryophyllaceae | E | 24 | 2 | AP | 2,867 | 2·9 | 5·9 | 11·7 | 437az | O | Lycopers.c | FC:PI | |||
| 744 | Silene pendula L. | No | Caryophyllaceae | E | 24 | 2 | A | 1,152 | 1·2 | 2·4 | 4·7 | 437 | O | Hc | FC:PI | |||
| 745 | Silene vulgaris (Moench) Garcke | No | Caryophyllaceae | E | 24 | 2 | P | 1,103 | 1·1 | 2·3 | 4·5 | 437 | O | Hc | FC:PI | |||
| 746a | Simmondsia chinensis (Link) C.K.Schneid. | No | Simmondsiaceae | E | 48-50 | —p | P | 723 | 0·7 | 1·5 | 3·0 | 465 | O | J | Fe | |||
| 747 | Spirodela polyrrhiza (L.) Schleid. | No | Araceaek | M | 80 | 4 | P | 292 | 0·3 | 0·6 | 1·2 | 400 | O | G | Fe | |||
| 748 | Spirodela punctata (G.F.W.Meyer) Thompson | No | Araceae | M | 46 | 2 | P | 363 | 0·4 | 0·7 | 1·5 | 400 | O | G | Fe | |||
| 749 | Stemona tuberosa Lour. | No | Stemonaceae | M | —n | —p | P | 715 | 0·7 | 1·5 | 2·9 | 379 | O | J | Fe | |||
| 750 | Stenotaphrum secundatum (Walt.) Kuntze. | No | Gramineae | M | 18 | 2 | P | 529 | 0·5 | 1·1 | 2·2 | 417 | O | Gallusf | FC:PI | |||
| 751 | Strelitzia nicolai Regel & C. Koch | No | Strelitziaceae | M | 14, 22° | 2 | P | 566 | 0·6 | 1·2 | 2·3 | 379 | O | J | Fe | |||
| 752 | Streptocarpus cyaneus S. Moore | No | Gesneriaceae | E | c. 30 | 2 or 4 | P | 662 | 0·7 | 1·4 | 2·7 | 378 | O | J | Fe | |||
| 753 | Stylidium adnatum R. Br. | No | Stylidiaceae | E | 30 | 2 | P | 1,495 | 1·5 | 3·1 | 6·1 | 378 | O | J | Fe | |||
| 754 | Stylobasium spathulatum Desf. | No | Surianaceae | E | 30 | 2 | P | 1,284 | 1·3 | 2·6 | 5·2 | 378 | O | J | Fe | |||
| 755 | Syngonium albo-lineatum Bull | No | Araceae | M | 22 | —p | AP | 4,628 | 4·7 | 9·4 | 18·9 | 411 | O | Bc | Fe | |||
| 756 | Syngonium podophyllum Schott | No | Araceae | M | 24 | —p | AP | 4,755 | 4·9 | 9·7 | 19·4 | 411 | O | Bc | Fe | |||
| 757 | Tabebuia argentea (Bureau & K.Schum.) Britton | No | Bignoniaceae | E | —n | —p | P | 782 | 0·8 | 1·6 | 3·2 | 454 | O | Bc | Fe | |||
| 758 | Tamarindus indica L. | No | Leguminosae | E | 26° | —p | P | 821 | 0·8 | 1·7 | 3·4 | 454 | O | Bc | Fe | |||
| 759 | Tecoma stans (L.) Juss. ex Kunth | No | Bignoniaceae | E | 36° | —p | P | 593 | 0·6 | 1·2 | 2·4 | 454 | O | Bc | Fe | |||
| 760 | Thespesia lampas (Cavanilles) Dalzell ex Dalzell & Gibson | —m | Malvaceae | E | 26° | 2 | —q | 1,568 | 1·6 | 3·2 | 6·4 | 444bd | O | Gb2 | FC:PI | |||
| 761a | Thespesia populnea (L.) Solander ex Correa | —m | Malvaceae | E | 26 | 2 | P | 4,018 | 4·1 | 8·2 | 16·4 | 444bd | O | Gb2 | FC:PI | |||
| 761b | Thespesia populnea (L.) Solander ex Correa | No | Malvaceae | E | 26° | 2 | P | 3,011 | 3·1 | 6·1 | 13·3 | 454 | O | Bc | Fe | |||
| 762 | Thespesia thespesioides (R.Brown ex Bentham) Fryxell | —m | Malvaceae | E | 26 | 2 | —q | 1,568 | 1·6 | 3·2 | 6·4 | 444bd | O | Gb2 | FC:PI | |||
| 763 | Triteleia laxa Benth. | No | Asparagaceae | M | 28° | 4 | P | 10,435 | 10·6 | 21·3 | 42·6 | 380 | O | B | FC:PI | |||
| 764 | Trochodendron aralioides Siebold & Zucc. | No | Trochodendraceae | E | 38 | 2 | P | 1,872 | 1·9 | 3·8 | 7·6 | 380 | O | Lycopers.c | FC:PI | |||
| 765 | Typhonium cuspidatum Decne. | No | Araceae | M | 16 | —p | A | 5,064 | 5·2 | 10·3 | 20·7 | 411 | O | Bc | Fe | |||
| 766 | Typhonium trilobatum Schott | No | Araceae | M | 40 | —p | A | 6,453 | 6·6 | 13·2 | 26·3 | 411 | O | Bc | Fe | |||
| 767 | Vicia canescens Lab. | No | Leguminosae | E | 10 | 2 | P | 3,058 | 3·1 | 6·2 | 12·5 | 409 | O | Bc | Fe | |||
| 768a | Vicia cracca L. ssp. tenuifolia | No | Leguminosae | E | 14 | 2 | P | 5,797 | 5·9 | 11·8 | 23·7 | 409 | O | Bc | Fe | |||
| 768b | Vicia cracca L. ssp. cracca | No | Leguminosae | E | 14 | 2 | P | 6,409.al | 6·5al | 13·1al | 26·2al | 409 | O | Bc | Fe | |||
| 769a | Vicia epetiolaris Burk.h | No | Leguminosaej | E | 14 | 2 | A | 4,067 | 4·2 | 8·3 | 16·6 | 415 | O | B | Fe | |||
| 769b | Vicia epetiolaris Burk.h | No | Leguminosaej | E | 14 | 2 | A | 4,533 | 4·6 | 9·3 | 18·5 | 415 | O | B | Fe | |||
| 770b | Vicia eristalioides Maxted | No | Leguminosae | E | 14 | 2 | A | 9,452 | 9·6 | 19·3 | 38·6 | 406 | O | Vicia narb.e | Fe | |||
| 771r | Vicia faba L. | No | Leguminosaej | E | 12° | 2 | A | 12,740 | 13·0 | 26·0 | 52·0 | 457bm | O | Bd | Fe | |||
| 771s | Vicia faba L. var. equina | No | Leguminosae | E | 12 | 2 | A | 14,377 | 14·7 | 29·3 | 58·7 | 408 | O | Cd | Fe | |||
| 771t | Vicia faba L. ‘Futura RZ’ | —m | Leguminosae | E | —n | —p | A | 12,985 | 13·3 | 26·5 | 53·0 | 384aa | O | Homof | FC:PI | |||
| 772a | Vicia galilaea Plitm. & Zoh. | No | Leguminosae | E | 14 | 2 | A | 6,392 | 6·5 | 13·0 | 26·1 | 408 | O | Cd | Fe | |||
| 772b | Vicia galilaea Plitm. & Zoh. | No | Leguminosae | E | 14 | 2 | A | 7,904 | 8·1 | 16·1 | 32·3 | 409 | O | Bc | Fe | |||
| 773d | Vicia graminea Sm.i | No | Leguminosaej | E | 14 | 2 | AB | 4,969 | 5·1 | 10·1 | 20·3 | 415 | O | B | Fe | |||
| 774b | Vicia hyaeniscyamus Mout. | No | Leguminosae | E | 14 | 2 | A | 7,654 | 7·8 | 15·6 | 31·2 | 408 | O | Cd | Fe | |||
| 775e | Vicia hybrida L. | No | Leguminosae | E | 12 | 2 | A | 8,306 | 8·5 | 17·0 | 33·9 | 409 | O | Bc | Fe | |||
| 776e | Vicia hyrcanica Fisch. & Mey. | No | Leguminosae | E | 12 | 2 | A | 7,634 | 7·8 | 15·6 | 31·2 | 409 | O | Bc | Fe | |||
| 777c | Vicia johannis Tamamsch. | No | Leguminosae | E | 14 | 2 | A | 6,145 | 6·3 | 12·5 | 25·1 | 408 | O | Cd | Fe | |||
| 778b | Vicia kalakhensis Khattab, Maxted & Bisby | No | Leguminosae | E | 14 | 2 | A | 10,344 | 10·6 | 21·1 | 42·2 | 406 | O | Vicia narb.e | Fe | |||
| 779 | Vicia macrograminea Burk. | No | Leguminosaej | E | 14 | 2 | BP | 5,821 | 5·9 | 11·9 | 23·8 | 415 | O | B | Fe | |||
| 780d | Vicia melanops Sibth. & Sm. | No | Leguminosae | E | 10 | 2 | A | 6,764.ak | 6·9ak | 13·8ak | 27·6ak | 408 | O | Cd | Fe | |||
| 781 | Vicia nana Vog. | No | Leguminosaej | E | 14 | 2 | A | 4,376 | 4·5 | 8·9 | 17·9 | 415 | O | B | Fe | |||
| 782a | Vicia pampicola Burk.h | No | Leguminosaej | E | 14 | 2 | A | 4,640 | 4·7 | 9·5 | 18·9 | 415 | O | B | Fe | |||
| 782b | Vicia pampicola Burk.h | No | Leguminosaej | E | 14 | 2 | A | 5,223 | 5·3 | 10·7 | 21·3 | 415 | O | B | Fe | |||
| 783d | Vicia peregrina L. | No | Leguminosae | E | 14 | 2 | A | 9,540 | 9·7 | 19·5 | 38·9 | 409 | O | Bc | Fe | |||
| 784d | Vicia pisiformis L.h | No | Leguminosae | E | 12 | 2 | P | 6,223 | 6·4 | 12·7 | 25·4 | 407 | O | Cd | Fe | |||
| 784e | Vicia pisiformis L.h | No | Leguminosae | E | 12 | 2 | P | 8,038 | 8·2 | 16·4 | 32·8 | 407 | O | Cd | Fe | |||
| 785v | Vicia sativa L. line 20·1i | No | Leguminosae | E | 12 | 2 | A | 1,882 | 1·9 | 3·8 | 7·7 | 409 | O | Bc | Fe | |||
| 785w | Vicia sativa L. line 31i | No | Leguminosae | E | 12 | 2 | A | 2,445 | 2·5 | 5·0 | 10·0 | 409 | O | Bc | Fe | |||
| 785x | Vicia sativa L. ssp. amphicarpa | No | Leguminosae | E | 12 | 2 | A | 2,102 | 2·1 | 4·3 | 8·6 | 409 | O | Bc | Fe | |||
| 785y | Vicia sativa L. ssp. nigra var. nigra | No | Leguminosae | E | 12 | 2 | A | 2,337 | 2·4 | 4·8 | 9·5 | 409 | O | Bc | Fe | |||
| 786e | Vicia sepium L. | No | Leguminosae | E | 14 | 2 | P | 4,719 | 4·8 | 9·6 | 19·3 | 409 | O | Bc | Fe | |||
| 787c | Vicia serratifolia Jacq. | No | Leguminosae | E | 14 | 2 | A | 9,700 | 9·9 | 19·8 | 39·6 | 405 | O | Cc | Fe | |||
| 788d | Viscum album L. | No | Loranthaceaek | E | 20° | 2 | P | 52,430 | 53·5 | 107·0 | 214·0 | 457bm | O | Bd | Fe | |||
| 789 | Vitex negundo L. | No | Lamiaceae | E | 34° | —p | P | 1,590 | 1·6 | 3·2 | 6·5 | 454 | O | Bc | Fe | |||
| 790 | Vitex pinnata L. | No | Lamiaceae | E | —n | —p | P | 1,411 | 1·4 | 2·9 | 5·8 | 454 | O | Bc | Fe | |||
| 791 | Voacanga grandifolia (Miq.) Rolfe | No | Apocynaceae | E | —n | —p | P | 358 | 0·4 | 0·7 | 1·5 | 454 | O | Bc | Fe | |||
| 792 | Wolffia arrhiza (L.) Horkel ex Wimmer | No | Araceae | M | 42 | 2 | P | 1,600 | 1·6 | 3·3 | 6·5 | 400 | O | G | Fe | |||
| 793 | Wolffiella oblonga (Phil.) Hegelm. | No | Araceae | M | 42 | 2 | P | 742 | 0·8 | 1·5 | 3·0 | 400 | O | G | Fe | |||
| 794 | Xanthorrhoea preisii Endl. | No | Xanthorrhoeaceae | M | 22 | 2 | P | 1,014 | 1·0 | 2·1 | 4·1 | 380 | O | J | Fe | |||
| 795 | Xanthosoma sagittifolium (L.) Schott | No | Araceae | M | 38 | —p | P | 8,609 | 8·8 | 17·6 | 35·1 | 411 | O | Bc | Fe | |||
| 796 | Xeronema callistemon W.R.B. Oliv. | No | Xeronemataceae | M | 34 | 2 or 4 | P | 3,210 | 3·3 | 6·6 | 13·1 | 380 | O | G | FC:PI | |||
| 797 | Xerophyta humilis Th. Dur. & Schinz. | No | Velloziaceae | M | 48° | 4 or 8 | P | 532 | 0·5 | 1·1 | 2·2 | 378 | O | J | Fe | |||
| 798 | Ximenia americana Linn. | No | Olacaceae | E | 26 | 2 | P | 1,595 | 1·6 | 3·3 | 6·5 | 379 | O | J | Fe | |||
| 799 | Xiphidium caeruleum Aubl. var. caeruleum | No | Haemodoraceae | M | 38° | 2 | P | 767 | 0·8 | 1·6 | 3·1 | 378 | O | J | Fe | |||
| 800 | Xyris gracilis ssp. gracilisl | No | Xyridaceae | M | 26° | 2 | P | 6,867 | 7·0 | 14·0 | 28·0 | 380 | O | B | FC:PI | |||
| 801bx | Zea mays ssp. mays L. line opaque 2h | No | Gramineaej | M | 20 | 4 | A | 3,262 | 3·3 | 6·7 | 13·3 | 392 | O | B | Fe | |||
| 801by | Zea mays ssp. mays L. race Altiplanoh | No | Gramineaej | M | 20 | 4 | A | 2,454 | 2·5 | 5·0 | 10·0 | 392ag | O | B-801bx | Fe | |||
| 801bz | Zea mays ssp. mays L. race Blanco y ocho rayash | No | Gramineaej | M | 20 | 4 | A | 3,311 | 3·4 | 6·8 | 13·5 | 392ag | O | B-801bx | Fe | |||
| 801ca | Zea mays L. | No | Gramineaej | M | 20° | 4 | A | 3,283 | 3·4 | 6·7 | 13·4 | 457bm | O | Bd | Fe | |||
| 802 | Zizyphus glabrata Heyne | No | Rhamnaceae | E | 24° | —p | P | 1,517 | 1·5 | 3·1 | 6·2 | 454 | O | Bc | Fe | |||
| 803 | Zostera marinal | No | Zosteraceae | M | 12° | 2 | P | 309 | 0·3 | 0·6 | 1·3 | 380 | O | J | Fe | |||
| 804 | Zoysia japonica Steud. | No | Gramineae | M | 40 | 4 | P | 421 | 0·4 | 0·9 | 1·7 | 417 | O | Gallusf | FC:PI | |||
Chromosome number.
E, ephemeral; A, annual; B, biennial; P, perennial.
O, original value; C, calibrated value
The standard species used to calibrate the present amount.
Fe, Feulgen microdensitometry; FC, flow cytometry using one of the following fluorochromes: PI, propidium iodide; DAPI, 4′, 6-diamidinophenylindole; EB, ethidium bromide; MI, mithramycin; HO,Hoechst 33258; GS, genome sequencing; CIA, computer image analysis; RK, reassociation kinetics.
E, eudicot; M, monocot; BA, basal angiosperm.
PROGRESS
Research on DNA C-values in angiosperms is unique in having been subject to detailed analyses of its quantity and quality over a long period (Bennett and Leitch, 1995). The importance of identifying gaps in our knowledge concerning this key biodiversity character, of recommending targets for new work to fill them by collaboration of international partners, and of monitoring progress to ensure that any shortfall is recognized, was confirmed by the first plant genome size workshop in 1997 (http://www.kew.org/cval/conference.html#outline, Bennett et al., 2000) and reviewed by participants at the second plant genome size workshop in 2003. Thus, what follows is mainly a summary of the overall progress for angiosperms against key targets set in 1997 for the following quinquennium (1998–2002). However, it also notes meaningful statistics for the data included in the Appendix table, or known to us from personal communications made after the Appendix table was closed.
In 1997 C-values for 2802 species (approximately 1 %) of angiosperm species had been estimated in the previous 40 years. The 1997 workshop concluded that the ideal of a C-value for all taxa was unrealistic, but long-term, estimates for 10–20 % of angiosperms seemed both ultimately achievable and adequate for all conceivable uses provided they were carefully targeted to be representative of the various taxonomic groups, geographical regions, and life forms in the global flora. So the first recommended target was to estimate first C-values for the next 1 % of angiosperm species (i.e. another 2500 species) by 2003. Many saw this goal as aspirational, as achieving it would mean estimating as many C-values in five years as in the past 40. Others thought that new technology (e.g. flow cytometry) would make it easy to achieve.
Improved systematic representation (species and families)
(i) First estimates for species
In September 1997 the Angiosperm DNA C-values database contained data for 2802 species. By September 2003 C-values were listed for 4119 species, including 689 first values for species listed in Bennett et al. (2000) and 628 such values for species in the Appendix table. Progress toward the first target in the five year period (1998–2002) considerably exceeded the average of ∼110 first values for species per annum in the early 1990s. Clearly, the 1997 workshop stimulated an increase in the total output of first C-values for species to its highest level for any five-year period (almost 200 per annum; Fig. 1A). Moreover, the proportion of newly published C-values that were also first estimates for species, which had previously fallen (Bennett et al., 2000), rose as a result of recent targeting and averaged 72·5 % for values published since 1997 (Fig. 1B). Nevertheless, the total number of published first C-values for species (1032) listed since 1997 was only 41 % of the 1997 target of approx. 2500.
Fig. 1.
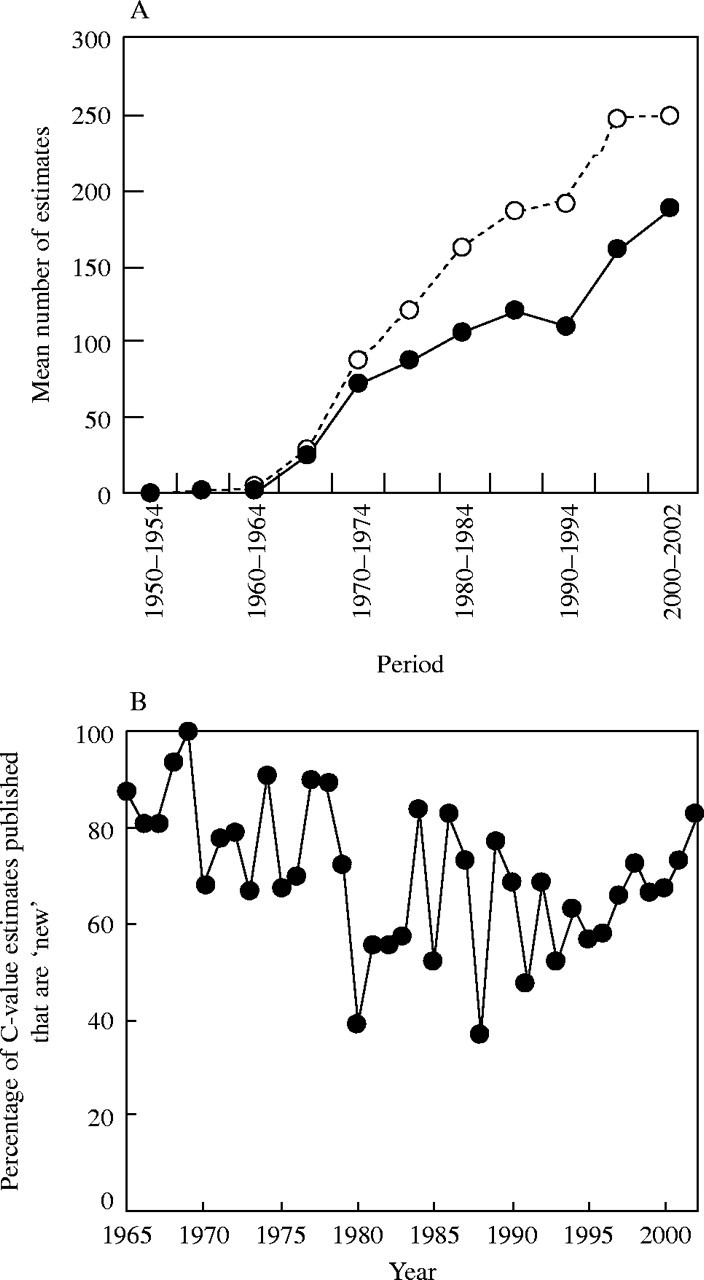
(A) Mean number per year of total (open symbols) and ‘first’ (closed symbols) DNA C-value estimates communicated in ten successive 5-year periods and the 3-year period 2000–2002, between 1950 and 2002. Based on analysis of data listed in the present Appendix table, and the Angiosperm DNA C-values database (release 4.0, January 2003). (B) Percentage of C-value estimates published or communicated during 1965–2002 that are first values for species listed in the present Appendix table and the Angiosperm DNA C-values database (release 4.0, January 2003).
The real total of first C-values for angiosperms estimated after 1996 but unpublished by 2003 was much higher, but is difficult to determine exactly. For example, several hundred values were measured by Ben Zonneveld (pers. comm.) using flow cytometry but not published. Listing for the Appendix table closed in August 2002, ready for the workshop; however, we saw 158 first C-values published by other authors later in 2002, and 22 such values were estimated at RBG, Kew. Adding these data to those listed in our compilations suggests that the total number of first C-values for species estimated in 1997–2002 was probably at least 1700 and hence not less than approx. 66 % of the target set in 1997. Analysis shows that this was achieved by international collaboration involving at least 18 research groups in ten countries. Whilst a target of 2500 was aspirational, it seems attainable as a future five-year goal. However, at the present rate achieving 20 % species representation would take 100 years, so an ultimate goal of 10 % (approx. 25 000 angiosperm species) is more sensible.
(ii) First estimates for families
The 1997 workshop noted that a first C-value was available for only 30 % of angiosperm families recognized at that time. Thus, a second recommended target was ‘To obtain at least one C-value estimate for a species in all angiosperm families’. Monitoring first C-values for species listed in Bennett et al. (2000) showed that progress towards this goal was initially very slow. Indeed, ‘since 1997 first C-values had been listed for 691 angiosperm species, but only 12 (1·7 %) were also first estimates for families’. Work to correct this began at RBG, Kew in 1999. In 2001 two papers reported first C-values for 50 families (Hanson et al., 2001a, b), and 30 more followed, including five basal angiosperm families (Leitch and Hanson, 2002; Hanson et al., 2003), all included in the present Appendix table.
Analysis of listed data for 4119 species shows that a first published value is available for at least 217 of the 457 angiosperm families currently recognized by the Angiosperm Phylogeny Group (APG) (APG II, 2003). Together with first estimates for 11 unlisted families (Hanson, RBG, Kew, pers. comm.; Koce et al., 2003) measured or seen after listing for the Appendix table was closed, the total is 228. Thus, since 1997 (after losses owing to new familial circumscriptions—APG II, 2003; Hanson et al., 2003) first values for at least 85 such families have been measured, so good progress has been made. However, the proportion of families represented rose only from 30 % to 49·9 % (Fig. 2), which is less than one third of the target (100 %) set in 1997. Major factors limiting progress were discussed previously (Hanson et al., 2003). Unlike progress towards the species target, which involved many research groups, movement towards the goal for families since 1997 has depended mainly on work by one institution, as RBG Kew estimated 65 of the 74 (87 %) first values for families listed in the Appendix table.
Fig. 2.

Cumulative percentage of angiosperm families recognized by the Angiosperm Phylogeny Group (APG) (APGII, 2003) with a first C-value represented in the present Appendix table, the Angiosperm DNA C-values database (release 4.0, January 2003), plus eleven known to the present authors in September 2003.
The Plant Genome Size Workshop in 2003 confirmed that global capacity for estimating DNA C-values (determined by available equipment, funding and trained operators) remains very limited. Consequently, any increased focus on targets for other plant groups (e.g. bryophytes, pteridophytes, gymnosperms—reviewed in Leitch and Bennett, 2002b) inevitably reduces progress to improve representation for angiosperm targets, a problem discussed below. However, it should not detract from the highly successful progress to make C-values more representative of the global flora described above.
PROBLEMS
Geographical representation and distribution
We first noted the need to improve geographical representation for angiosperm C-values in Bennett and Leitch (1995). This was confirmed by the 1997 plant genome size workshop, although no specific regional targets were recommended. Perhaps, in consequence, progress in this area has never been monitored in detail, although we have been at pains to advertise the problem in a general way and to provoke action to rectify it in particular regions, such as southern Africa (Leitch and Bennett, 2002a).
There are two critical concerns regarding the geographical distribution of angiosperm C-value work. (i) The first concerns the small number of publications with original C-values by first authors in many regions (Table 1). This reflects a serious imbalance between the geographical distribution of research scientists working on genome size and of taxa whose C-values are unknown. Bennett et al. (2000) noted that ‘Africa remains an unexplored continent’ and that ‘Whereas six out of 377 original sources have first authors with addresses in Africa, still none has an angiosperm C-value estimated in Africa, as all six reported work done in Europe or the USA.’ Analysis of the 88 original sources in the present work shows no improvement, as the number of original sources from Africa (2), China (2), and South America (2) remains low (Table 1). (ii) The second concerns the small number of first C-values by any authors for species endemic to several large geographical regions. With some exceptions, the sample is still dominated by crops and their wild relatives, model species grown for experimental use, and other species growing near laboratories in temperate regions, mainly in Western Europe and North America. Analysis of data in the Appendix table shows that none presented data for other taxa endemic to China, Japan, Brazil, Mexico or Central Africa. Similarly, although island floras are known to be rich in endemics, no original source has reported C-values for any large islands such as Borneo, New Guinea or Madagascar, where 80 % of the 12000 described plant species are endemic (Robinson, 2004).
Table 1.
The number and percentage (in brackets) of original references with first authors from various geographical areas among the total of 465 sources contributing to the present Appendix table and the six lists of angiosperm DNA amounts previously compiled for reference purposes that were pooled in the Angiosperm DNA C-values database (release 4.0, January 2003)
| DNA C-value compilation |
||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Area |
19761 |
19822 |
19913 |
19954 |
19975 |
20006 |
Present Appendix |
Total |
||||||
| Europe | 34 (63·0) | 38 (71·7) | 30 (53·6) | 43 (40·6) | 18 (48·6) | 38 (51·4) | 54 (61·4) | 255 (54·8) | ||||||
| UK | 28 (51·9) | 13 (24·5) | 22 (39·3) | 23 (21·7) | 5 (13·5) | 8 (10·8) | 10 (11·4) | 109 (23·4) | ||||||
| North America | 14 (25·9) | 11 (20·8) | 16 (28·6) | 19 (17·9) | 5 (13·5) | 11 (14·9) | 13 (14·8) | 89 (19·1) | ||||||
| South and Meso America | 0 (0·0) | 0 (0·0) | 3 (5·4) | 9 (8·5) | 1 (2·7) | 6 (8·1) | 2 (2·3) | 21 (4·5) | ||||||
| Africa | 1 (1·9) | 0 (0·0) | 0 (0·0) | 1 (0·9) | 2 (5·4) | 1 (1·4) | 2 (2·3) | 7 (1·5) | ||||||
| Asia | 1 (1·9) | 3 (5·7) | 4 (7·1) | 30 (28·3) | 11 (29·7) | 8 (10·8) | 17 (19·3) | 74 (15·9) | ||||||
| India | 1 (1·9) | 1 (1·9) | 2 (3·6) | 28 (26·4) | 11 (29·7) | 4 (5·4) | 11 (12·5) | 58 (12·5) | ||||||
| China | 0 (0·0) | 0 (0·0) | 0 (0·0) | 0 (0·0) | 0 (0·0) | 0 (0·0) | 2 (2·3) | 2 (0·4) | ||||||
| Australasia | 4 (7·4) | 1 (1·9) | 3 (5·4) | 4 (3·8) | 0 (0·0) | 7 (9·5) | 0 (0·0) | 19 (4·1) | ||||||
| Australia | 4 (7·4) | 1 (1·9) | 3 (5·4) | 1 (0·9) | 0 (0·0) | 1 (1·4) | 0 (0·0) | 10 (2·2) | ||||||
| Total | 54 | 53 | 56 | 106 | 37 | 71 | 88 | 465 (100) | ||||||
Plant life form
There is also a need for the overall sample to represent better the full range of plant types and life forms. We previously identified several associations and life forms as being poorly represented in the database (Bennett and Leitch, 1995), yet taxa from bog, fen, tundra, alpine and desert environments, and halophytic, insectivorous, parasitic, saprophytic and epiphytic species and their associated taxa are all still under-represented.
Solving this problem needs a proactive approach, as recent experience with first C-values for angiosperm families shows. First, a target must be set for each gap. Second, monitoring newly published data against targets must begin. Third, if work on poorly-represented floras or life forms does not increase, then established research groups must re-focus on target material available in existing collections. Unless global capability for estimating plant DNA C-values is significantly increased by new technology, funds or skilled operators, then this change in strategy will reduce progress towards achieving other targets. However, the prime objective remains: to generate a sample representative of the global flora that is able to support most comparative studies. Managers of the limited global capacity for estimating genome size should keep this firmly in mind when targeting taxa for new work.
Obsolescence time bomb
Several methods have been used to measure plant DNA C-values, but most values have been estimated by Feulgen microdensitometry (Fe), both overall and since 1997. In 1997 we identified an imminent problem, likely to limit future estimations. This was the failure and non-replacement of densitometers long used by many groups to estimate DNA C-values. Manufacturers were ending their support for such equipment, and users faced difficulty in funding new equipment for this purpose. Moreover, this problem was likely to be most acute in regions where some of the greatest gaps in our knowledge lay. Reviewing the position at the second Plant Genome Size Workshop confirmed that, as predicted, the ‘obsolescence time bomb’ had exploded. By 2003 several laboratories that had long published C-values listed in the Angiosperm DNA C-values database were now unable to estimate C-values by this (e.g. in Mexico, the USA), or any method (e.g. in Argentina). Vickers Instruments no longer supports their M85 microdensitometer, and spare parts for it are unobtainable. A few laboratories, including ours, can still use such machines, but now without servicing and only until they fail catastrophically.
As expected, one response to this problem was the increased use of flow cytometry (FC) to replace Fe. Analysis of data in the Appendix table shows a higher proportion of C-values obtained by FC (58·4 %), mostly since 1997, than noted previously (48·6 %) for data listed in Bennett and Leitch (2000), whilst in Bennett and Leitch (1995, 1997) FC averaged 26·7 %. Several groups have undertaken careful studies to compare DNA estimates made by FC and Fe, to define best practice for FC, or to show that FC can be applied widely to most plants across the full range of known DNA C-values (e.g. see review by Doležel and Bartos, 2005, this volume). Fortunately, the cost of a basic flow cytometer for such work has fallen, and suitable models (e.g. Partec PAII) have recently been set up for this use for approx. £20K (US$30K). If this technology continues to improve, and its costs continue to fall, FC should be more easily available. However, FC easily yields poor data in unskilled hands and by itself does not provide the cytological view of test material(s) that is essential to count chromosome number(s). Its use in some less-developed countries (where the greatest gaps in our knowledge still remain) will depend on training local operators, but such capacity-building may be thwarted by a lack of in-country support by the suppliers of flow cytometers.
A second solution to the problem is a new availability of relatively inexpensive computer-based image analysis (CIA) systems, which can estimate DNA amounts using Feulgen-stained cytological preparations in place of a microdensitometer. Although proprietary hard-wired CIA systems have been available since the 1970s (e.g. Zeiss Quantimet system), they cost much more than microdensitometers, and analysis of the literature shows they have not been used to estimate plant C-values. However, in the 1990s, with advances in computer technology, less expensive systems were developed (e.g. CIRES system) primarily for medical use, and these have also been used to good effect for plant C-value estimations (e.g. Temsch et al., 1998; Greilhuber et al., 2000).
Sadly, the CIRES system that adapted well for this purpose is no longer available, as the software is incompatible with the operating system used on modern PCs. However, computer-literate groups can assemble the kit needed to estimate C-values using CIA, and several software packages written specifically for this purpose are available (Vilhar et al., 2001; Hardie et al., 2002). Hardie et al. (2002) give an excellent review of this technique and practical issues concerned with its use for animal materials, and Vilhar et al. (2001) have compared CIA, Fe and FC, to help define best practice for CIA, demonstrating that CIA can be applied to plants with an approx. 100-fold range of C-values. Vilhar et al. (2001) concluded that ‘DNA image photometry gives accurate and reproducible results, and may be used as an alternative to photometric cytometry in plant nuclear DNA measurements’. CIA can use an existing microscope, costs less than FC to set up, and is easier to service in countries that lack FC manufacturers' support. The field would benefit from development of a standard inexpensive CIA ‘kit’, an agreed best practice CIA technique, and easy access to leading laboratories for training and technology transfer. Given this, CIA could soon become the method of choice for estimating C-values in angiosperms, replacing Fe as a method of choice along with FC, but with the advantage that, unlike FC, it uses microscope slide preparations, allowing users to make cytological observations.
Errors and inexactitudes
Swift (1950) defined the DNA content of an unreplicated haploid complement as its 1C-value (C standing for ‘constant’). Thus, replicated diplophase nuclei have a 4C DNA amount and produce two unreplicated 2C nuclei by mitotic division, and four 1C gametic nuclei after meiosis, irrespective of the organism's ploidy level. This convention applies well to polyploid taxa with diploidized meiotic chromosome pairing such as hexaploid breadwheat, which produce mainly functional, balanced polyhaploid gametes with 1C DNA amounts at meiosis (Rees and Walters, 1965). Consequently, for several previous reference lists, 4C DNA estimates for all taxa were divided by 2 and 4 to generate 2C- and 1C-values respectively (e.g. Bennett et al., 2000). However, a problem with this practice was identified for the few taxa with odd ploidy levels in release 3.1 of the Angiosperm DNA C-values database (namely 45 out of 3493 listed taxa, ∼1·3 %), as the resulting 1C-values are not biologically meaningful. For example, triploids with a 4C amount in their fully replicated metaphase nuclei do regularly produce two 2C nuclei at mitosis, but do not regularly produce four 1C products at meiosis. The authors are grateful to several colleagues who noted this problem and suggested solutions.
This problem has several practical consequences. (i) Regrettably, researchers who use 1C data from the literature or downloaded from the Angiosperm DNA C-values database may have included this error in the samples that they used for comparative analyses. However, this is unlikely to have influenced their conclusions significantly, since the magnitude of the error is relatively small (ranging between −0·25C for a triploid, to +0·25C for a pentaploid—which tend to cancel out), and affects only 1·3 % of all taxa listed. Overall, errors in mean DNA amounts for samples are probably less than 0·5 %. Studies that used data from the 2C or 4C columns for samples of odd-ploid taxa are unaffected by the error. (ii) To ensure that researchers are aware of the problem and do not generate 1C data for taxa with odd ploidy levels in the future, release 4.0 of the Angiosperm DNA C-values database gives 2C- and 4C-values for the 45 out of 3493 entries with odd ploidy levels, plus a warning note in response to any queries for 1C-values. This approach is also followed in the present Appendix table (see footnote t). (iii) This problem also highlights a general need to re-assess definitions of ‘C-value’ and ‘genome size’ in light of recent usage and new theoretical understanding, a topic explored by Greilhuber et al. (2005). Indeed, the above problem shows the need for care when handling data, and the danger of using computer-generated numbers uncritically. It is clearly perilous to ignore basic biology or the literature, as the recent history of genome size, ‘complete’ genome sequencing, and interest in the smallest angiosperm genome clearly shows.
Genome size, ‘complete’ genome sequencing, and, the euchromatic genome
A growing semantic problem concerns different uses of the term ‘genome’ (Greilhuber et al., 2005). As originally defined by Winkler (1920), genome referred to a monoploid chromosome complement. Since a monoploid is defined as ‘having one chromosome set with the basic (x) number of chromosomes’ (Rieger et al., 1991), it followed by definition that any polyploid taxon had three or more genomes. However, an alternative meaning, now in common usage, uses genome as an interchangeable alternative for the 1C-value to refer to the DNA content of an unreplicated gametic nuclear complement, irrespective of ploidy level. Unless the meaning intended is clearly defined on each occasion, this can be confusing, especially when authors use both meanings for a polyploid taxon in the same paper. For example, Devos and Gale (1997) used the term ‘genome’ to refer to both the entire complement of nuclear DNA in a hexaploid wheat nucleus and to the individual A, B and D ‘genomes’.
Further potential for confusion comes from new uses of the term ‘genome’ recently spawned by genome sequencers. These concern the counter-intuitive meaning of a ‘wholly’, ‘completely’ or ‘entirely’ sequenced genome, or of equating ‘genome’ with ‘euchromatic genome’—a confusing concept in which ‘genome’ equals the parts which could be cloned and sequenced, but not the rest (see below). None of these qualitative new uses of genome equates to its quantitative use to mean either a 1C-value, or one monoploid parental genome in a polyploid.
The completely sequenced genome
Since 2000 the scientific and popular press has reported and celebrated the ‘complete’ sequencing of the first insect (Drosophila melanogaster) and plant genome (Arabidopsis thaliana) and the human genome (in 2001). For example, a title in Nature reported: ‘The sequencing of an entire plant genome is now complete.’ Readers could be forgiven for assuming this meant the entire linear sequence of the nuclear DNA had been sequenced and assembled, so that the total size of the nuclear genome in these organisms was now known with certainty, and hence much more accurately than any previous estimate based on other methods subject to various experimental errors. The popular and scientific literature easily gives that impression, and unfortunately that is what many, incorrectly, understood. The truth is otherwise, as a ‘completely sequenced’ genome is a very relative concept. In the same issue of Science where Brenner (2000) wrote ‘We have the complete sequence of the 125-megabase genome of the fruit-fly Drosophila’, Pennisi (2000) noted that ‘the fly sequence still has c. 1000 small gaps’—referring only to the sequenced euchromatin part. But what of the rest? Speaking of heterochromatin, Adams et al. (2000) explained that the ‘genomes of eukaryotes generally contain heterochromatic regions surrounding the centromeres that are intractable to all current sequencing methods’ and that ‘Because of the unclonable repetitive DNA surrounding the centromeres it is highly unlikely that the genomic sequence of chromosomes from eukaryotes such as Drosophila or human will ever be ‘complete’. Moreover, Adams et al. (2000) stated that the unsequenced centric heterochromatin regions comprised ‘one third’ of the approx.180 Mb genome of Drosophila. But how was its size determined? Careful reading revealed that the Mb size of these unsequenced centromeric heterochromatic segments was measured not by any modern molecular method, but by using a ruler on one cell of a plate in a paper by Yamamoto et al. (1990). This important detail is not stated in the main text, but in the legend to fig. 1 in Adams et al. (2000). As Bork and Copley (2001) clearly explain, ‘There are regions, often highly repetitive, that are difficult or impossible to clone (one of the initial steps in a sequencing project) or sequence with current technology.… The extent of these regions varies widely in different species. So, rather than applying a universal gold standard, each sequencing project has made pragmatic decisions as to what constitutes a sufficient level of coverage for a particular genome. For example, as much as one-third of the sequence of the fruitfly Drosophila melanogaster was not stable in the cloning systems used, and so was not sequenced.’
Thus, workers interested in C-values should clearly understand that a ‘completely’, ‘entirely’ or ‘wholly’ sequenced genome is not what those words might imply if taken at face value, and the size given for such a genome may indicate either the amount of DNA sequenced, or the size of that euchromatic genome sequenced plus a best-guess estimate of a lot of unsequenced heterochromatin. Further, it can mean that every type of sequence in an organism has been sequenced, but it need not mean that all copies of all types have been sequenced, or that their copy numbers are known. Without this information total genome size (the DNA C-value) cannot be determined based on genome sequencing (Bennett et al., 2003).
Swift (1953) stated that, ‘in general estimates of the nucleic acids in cells are at present accurate to 10 or 20 %’. Later, Bennett and Smith (1976) concluded that ‘While a few estimates are not accurate even to within 20 %, careful measurements of 4C DNA amounts in species with 0·5–2·0 times that of a standard species are probably accurate to within 5–10 %’. Greilhuber (1998) noted ‘much suspect or demonstrably wrong data have accumulated and continue to be accumulated in the literature’. Sadly, the ‘complete’ genome sequencing of Arabidopsis (Arabidopsis Genome Initiative, 2000), which was expected to provide a new baseline, only added to this phenomenon.
Plant genome size researchers have long recognized the need for an exact calibration standard, whose C-value is not subject to technical errors. Thus, the publication of a precise C-value for the first plant to have its genome completely sequenced was eagerly awaited, as it was expected to provide a baseline, gold-standard reference point, against which all other plants could be compared and expressed. Arabidopsis thaliana ecotype ‘Columbia’ was chosen for complete genome sequencing, partly because its tiny genome should be less costly to sequence than larger genomes in other species.
In 2000 the Arabidopsis Genome Initiative (AGI) published the genome size of Arabidopsis thaliana as 125 Mb, comprising 115·4 Mb in the sequenced regions plus a rough estimate of 10 Mb in unsequenced centromere and ribosomal DNA regions. The accuracy of this estimate was set not by the precision of sequencing and assembling contigs, but by the total inaccuracy in the sizes assumed for the unsequenced gaps (Bennett et al., 2003) and hence was no more accurate than many estimates in the range 150–180 Mb made by other methods. Further analysis showed that the AGI's rough estimate of 10 Mb in the unsequenced gaps was highly inaccurate. Thus, new comparisons using flow cytometry, which co-ran A. thaliana ecotype ‘Columbia’ with three animal species including Caenorhabditis elegans Bristol N2 (whose genome size is accurately established by genome sequencing as just over 100 Mb), gave C-value estimates for A. thaliana in the range 154–162 Mb (with 157 Mb when C. elegans was used as the standard) (Bennett et al., 2003). This value is about 25 % larger than the AGI estimate of 125 Mb which was clearly a gross underestimate, and hence is not the long-awaited first benchmark C-value for a completely sequenced plant genome—giving those words their natural meaning. Other molecular work has confirmed this conclusion (e.g. Hosouchi et al., 2002).
More recently, the draft DNA sequence of the rice (Oryza sativa) genome was published (O. sativa ssp. japonica, Goff et al., 2002; O. sativa ssp. indica, Yu et al., 2002). However, while the estimated genome sizes based on DNA sequencing did not suffer from the serious shortcomings of the Arabidopsis estimate, neither did they fulfil the criteria essential for a new benchmark calibration standard. Yu et al. (2002) gave a new C-value of 466 Mb for O. sativa ssp. indica calculated by adding up the DNA sequencing data for 362 Mb of sequenced scaffolds and 104 Mb of ‘unassembled data’. In contrast Goff et al. (2002) reported the sequencing of DNA which covered a total of 389 809 244 bp of the O. sativa ssp. japonica genome. They stated that this represented 93 % of the 420 Mb rice genome but did not give a reference to the source of 420 Mb. It is therefore unclear whether the C-value of 420 Mb given by Goff et al. (2002) represents a new C-value based on genome sequencing alone. The 1C-value for rice may yet prove to be slightly higher than the values assumed by Goff et al. (2002) and Yu et al. (2002), and approach 490 Mb, equivalent to the 0·5 pg estimated by Bennett and Smith (1991).
Exact C-values based on complete genome sequences would be invaluable (Bennett et al., 2003). The need to complete sequencing gaps in Arabidopsis remains technically difficult, and it is unclear how, when, or if it will be achieved. Genome sequencing becomes more difficult as genome size increases, and experience with Arabidopsis implies that exact C-values are unlikely to be obtained in this way soon for any larger plant genomes, including the established plant C-value standard Oryza sativa.
The current situation poses a quandary for the plant genome size community, who have long paid serious attention to trying to maximize the accuracy and comparability of plant DNA C-values by using agreed calibration standards (both materials and assumed values; e.g. see http://www.rbgkew.org.uk/cval/conference.html#outline, Bennett et al., 2000), while eagerly awaiting the first absolute measurement for a plant obtained by really complete DNA sequencing. Current options include: (i) continue to use the existing small group of plant calibration standards until a plant C-value which meets the required criteria becomes available; (ii) adopt an animal C-value which meets these criteria as the baseline reference for expressing all other plant species values, e.g. Caenorhabditis elegans Bristol N2, whose C-value is known with confidence to within 1 % from genome sequencing to be just above 100 Mb (or roughly 0·1 pg); (iii) adopt a plant value based on direct comparisons with C. elegans, as the base calibration standard for plants, and create a ladder of secondary calibration standards all measured against it in a study replicated between several groups able to use best practice. The C-value for Arabidopsis thaliana (1C = 157 Mb or 0·16 pg), recently measured against C. elegans (Bennett et al., 2003), could be adopted as the basal plant calibration standard. Seed is readily available from stock centres and gives small, easily grown plants. Moreover, the ladder of values for its many endopolyploid nuclei would also provide convenient calibration reference points for higher values up to approx. 2500 Mb or 2·5 pg (i.e. 0·64 − 4C, 1·28 − 8C, and 2·56 − 16C).
Weeding out erroneous data
The value of the database is determined by the accuracy of the data it contains. Ideally, values should be exact, but in reality they are all subject to various technical and other errors, as noted above. This raises questions as to how accurate data are, and what level of error is acceptable in practice, or makes a datum valueless for a particular use or study.
The existence of a database itself is a valuable means of identifying real or potential errors, and hence of improving the accuracy and quality of the whole body of data. For example, where estimates for the same taxon (with the same chromosome number) disagree greatly this suggests an error. Further, where a body of data for a taxon shows close agreement except for one major departure, this identifies the outlier as almost certainly incorrect. For example, in the Appendix table, the 2C-value for diploid Acacia dealbata (1·7 pg) reported by Blakesley et al. (2002) is similar to that reported by Bukhari (1997) of 2C = 1·6 pg (listed in Bennett et al., 2000), but both values differ considerably from the 2C-value of 2·9 pg reported by Mukerjee and Sharma (1993b) (see Notes to the Appendix bb). Another example concerns Brachypodium distachyon. In 1991, the PhD thesis of Shi reported a 1C-value of 0·15 pg, but later Shi et al. (1993) gave its 1C-value as 0·3 pg. To resolve this discrepancy, RBG, Kew obtained some original material studied by Shi and estimated its 1C-value to be 0·36 pg, confirming the larger C-value for this species (see also footnote br). Thus, real errors can be identified with certainty, and potential errors flagged up for users in cautionary footnotes following Appendix tables.
DNA C-values in angiosperms vary approx. 1000-fold (over three orders of magnitude) from approx. 0·1 pg to over 100 pg. It is, therefore, often useful to know whether a species' 1C DNA amount has approximately 0·1, 1, 10 or 100 pg, even if there is still uncertainty regarding whether a species with approximately 1 pg is really closer to 0·8 pg than to 1·2 pg (an error ±20 %). In terms of its predictive value in nucleotypic correlations, such an error still permits useful conclusions to be drawn. The Arabidopsis community laboured long under the misapprehension that its 1C DNA amount was approx. 70 Mb (Leutwiler et al., 1984), and later approx. 100 Mb (Meyerowitz, 1994), when in reality it is much higher (about 157 Mb, Bennett et al., 2003). The level of inaccuracy involved (approx. 50–100 %) was considerable, yet it did not prevent the selection of Arabidopsis as the model plant for first complete genome sequencing, in no small part on the basis of its ‘small genome size’ (NSF, 1990; Somerville and Somerville, 1999). The Convention on Biological Diversity (United Nations Environment Programme, 1992) noted the need to make biodiversity data available, despite imperfections; a view which merits support (Bennett, 1998). Thus, it is better to list available C-value data subject to errors, until improved data with fewer errors become available. The body of data is needed by the scientific community and can clearly already be used to draw important conclusions, to make valuable predictions, and as a basis for necessary planning.
What is the smallest reliable C-value for an angiosperm?
The above examples show how seeing data in the comparative context of the database can help to identify real or potential errors in particular species. It can also facilitate broader enquiries such as ‘what is the smallest reliable C-value for an angiosperm?’. Again, the comparative approach has enabled researchers to be active in identifying potential errors in species with the smallest reported C-values, and to be transparent in correcting mistakes.
Because of error variation, a population of 1C-value estimates for one taxon should vary according to a normal curve, so those in the lower tail are all too low (Fig. 3A). Such values are lost in the frequency histogram for all angiosperm C-value estimates except at its lowest tail where some of the lowest C-values claimed are expected to be too low. This expectation is strongly supported in practice, as shown below. There are 53 1C estimates in the Angiosperm DNA C-values database or the present Appendix with 0·21–0·30 pg, 29 with 0·11–0·20 pg, but only three with 0·10 pg or below (Fig. 3B). Table 2 lists the 24 lowest estimates listed with 0·175 pg or less, but how robust are they?
Fig. 3.
(A) Expected error variation in a large population of DNA C-value estimates for one genotype as underestimates (in the lower tail) and overestimates (in the upper tail) surround more accurate, intermediate, genome size estimates. (B) Histogram showing frequency of C-values for the 85 smallest species in the database or Appendix.
Table 2.
The 24 lowest angiosperm 1C DNA estimates among data listed in the present Appendix table and the Angiosperm DNA C-values database (release 4.0, January 2003)
| Taxon |
1C (pg) |
Original reference |
|---|---|---|
| Arabidopsis thaliana | 0·051 | Francis et al. (1990) |
| Cardamine amara | 0·055 | Band SR (pers. comm. 1984) |
| Arabidopsis thaliana | 0·073 | Leutwiler et al. (1984) |
| Fragaria viridis | 0·108 | Antonius and Ahokas (1996) |
| Rosa wichuriana | 0·125 | Bennett and Smith (1991) |
| Aesculus hippocastanum | 0·125 | Bennett et al. (1982) |
| Arabidopsis thaliana | 0·128 | Arabidopsis Genome Initiative (2000) |
| Sedum album | 0·145 | Hart (1991) |
| Arabidopsis thaliana | 0·150 | Arumaganathan and Earle (1991) |
| Carex nubigera | 0·150 | Nishikawa et al. (1984) |
| Carex paxii | 0·150 | Nishikawa et al. (1984) |
| Epilobium palustre | 0·150 | Band SR (pers. comm. 1984) |
| Hypericum hirsutum | 0·150 | Hanson, Leitch and Bennett (pers. comm. 2002) |
| Thlaspi alpestre | 0·150 | Band SR (pers. comm. 1984) |
| Arabidopsis thaliana | 0·153 | Bennett et al. (2003) |
| Arabidopsis thaliana | 0·160 | Bennett et al. (2003) |
| Arabidopsis thaliana | 0·160 | Galbraith et al. (1991) |
| Arabidopsis thaliana | 0·165 | Galbraith et al. (1991) |
| Arabidopsis thaliana | 0·167 | Krisai and Greilhuber (1997) |
| Arabidopsis thaliana | 0·167 | Bennett et al. (2003) |
| Amoreuxia wrightii | 0·168 | Hanson et al. (2001a) |
| Arabidopsis thaliana | 0·170 | Galbraith et al. (1991) |
| Arabidopsis thaliana | 0·175 | Bennett and Smith (1991) |
| Arabidopsis thaliana | 0·175 | Marie and Brown (1993) |
Thirteen of the 24 estimates in Table 2 are for Arabidopsis thaliana. A comparative approach suggests that some, which featured among the lowest C-values reported for angiosperms, are too low. Thus several C-value estimates made by molecular means in the range 0·05–0·125 pg (Leutwiler et al., 1984; Francis et al., 1990; Arabidopsis Genome Initiative, 2000) are now seen as gross underestimates, while many others in the range 0·15–0·18 pg are shown to span the true value of about 0·16 pg (Bennett et al., 2003).
After discounting the 1C estimate for Arabidopsis thaliana of 0·051 pg by Francis et al. (1990), the next smallest estimate listed is 0·055 pg for Cardamine amara (communicated from S. R. Band in 1984). With only a third the DNA amount of its related crucifer A. thaliana (0·16 pg), it seemed suspiciously low. Cardamine amara seed cannot survive drying, so it is unavailable from seed banks. However, we recently used flow cytometry to compare diploid C. amara collected near Sheffield with several calibration standards including A. thaliana ecotype ‘Columbia’. The 1C-value we obtained was around 0·24 pg (almost five-fold the earlier report). This is in close agreement with independent estimates made elsewhere (e.g. see Bennett and Leitch, 2005; Johnston et al., 2005).
Once the underestimates for Arabidopsis thaliana and Cardamine amara are discounted, few 1C-values of 0·125 pg or below remain for other species. One is the 1C estimate of 0·125 pg for Rosa wichuriana, estimated using callus material from Dr Andy Roberts (Bennett and Smith 1991). This value seemed questionably low in the context of the database, especially as it became clear that culturing may induce stain inhibitors. This concern led to a new collaboration with RBG, Kew using non-callous material, and our doubts were confirmed when it was re-estimated as 1C = 0·575 pg (Yokoya et al., 2000).
Perhaps all estimates below 0·125 pg should be doubted until confirmed. Another candidate was Aesculus hippocastanum whose 1C-value was listed as 0·125 pg (Bennett et al., 1982). This material is rich in tannins and a likely candidate for underestimating its DNA amount (Noirot et al., 2000, 2005). Recent work using flow cytometry at RBG, Kew, showed that the 1C-value of 0·125 pg was clearly an underestimate, as the true value is approx. 0·60 pg (L. Hanson, RBG, Kew, pers. comm.). With 0·125 pg for A. hippocastanum rejected, only one estimate below 0·14 pg remains, namely 0·11 pg for the Green strawberry, Fragaria viridis. Since there is considerable interest in knowing the smallest possible angiosperm genome, checks to establish whether this estimate is robust are now urgently required.
What is the minimum C-value for a free-living angiosperm and other free-living organisms?
Such comparative approaches can also facilitate broader questions such as: ‘what is the minimum genome size in angiosperms and other free-living organisms?’. There is a minimum compendium of nuclear genes essential for the life of any organism. This concept was behind Craig Venter's declared intension to synthesize from scratch a minimal bacterial genome (Check, 2002), and a project for a minimal eukaryote genome may eventually follow. Meanwhile we can only speculate on how small the minimum genome is for an angiosperm, and how closely extant species approach the minimum. It is, of course, below the lowest robust C-value for the group, i.e. less than 0·16 pg established for Arabidopsis thaliana. The presence of six other species with C-values of 0·15–0·169 pg in Table 2 strongly supports this conclusion. The estimate(s) of approx. 0·108 pg for Fragaria viridis may indicate a minimum C-value for extant angiosperms of about 100 Mb, but if so, is it a diploid, or a polyploid with three or more even smaller ancestral genomes?
Whilst the robust 1C-value for A. thaliana is 0·16 pg, this includes >25 % of repeated DNA (Bennett et al., 2003) and analysis of sequenced regions shows that >70 % of coding genes are duplicated (Bowers et al., 2003). Thus, in theory, a minimal genome without duplicated coding genes or repetitive DNA should not exceed approx. 50 Mb. Currently, there is no robust 1C estimate below 0·1 pg for an angiosperm, but if any of the seven species with C-values between 0·14–0·15 pg is a tetraploid, this would indicate a minimum genome size in extant taxa of approx. 75 Mb, or approx. 50 Mb if it is a hexaploid.
The comparative approach is usefully extended to include other groups of organisms. Table 3 shows minimum C-value estimates for multicellular organisms in several widely differing groups obtained by genome sequencing or other methods. Such minima for groups as diverse as nematodes, insects, algae and angiosperms range from 59 to 160 Mb. Thus, the minimum C-value known in extant free-living multicellular higher organisms is around 60 Mb. All may be diploidized paleopolyploids (Wendel, 2000), but except for one early and unconfirmed report of a 1C-value of approx. 39 Mb in a most simple multicellular placozoan animal (Ruthmann and Wenderoth, 1975) there is no evidence for extant diploid multicellular eukaryotic life forms with only 40–50 Mb. This tantalizing possibility will be an interesting driver for new work to find a first angiosperm whose 1C-value is <100 Mb, or a first free-living multicellular plant or animal with a robust 1C-value <50 Mb.
Table 3.
Robust minimum 1C-value estimates for several widely different groups of free-living, multicellular, higher organisms obtained by genome sequencing (*), other best practice techniques, or static cytometry using the fluorochrome DAPI for algae1
| Group |
Species |
Mb |
Original reference |
|||
|---|---|---|---|---|---|---|
| ANIMALS | ||||||
| Nematode | Caenorhabditis elegans | 100* | C. elegans sequencing Consortium (1998) | |||
| Platyhelminthes (flatworms) | Stenostomum brevipharyngium | 59 | Gregory et al. (2000) | |||
| Crustacea | Scapholeberis kingii (water flea) | 157 | Beaton (1988) | |||
| Annelid | Dinophilus gyrociliatus (polychaete worm) | 59 | Soldi et al. (1994) | |||
| Tardigrades (water bears) | Isohypsibius lunulatus | 78 | Redi and Garagna (1987) | |||
| Insect | Peristenus stygicus | 98 | TR Gregory (pers. comm.) | |||
| Arachnid | Tetranychus urticae (spider mite) | 78 | TR Gregory (pers. comm.) | |||
| Urochordates (tunicates) | Oikopleura dioica | 72 | Seo et al. (2001) | |||
| PLANTS | ||||||
| Chlorophyta (green alga) | Caulerpa paspaloides | 88 | Kapraun (2005) | |||
| Rhodophyta (red algae) | Heydrichia wolkerlingii | 69 | Kapraun (2005) | |||
| Phaeophyta (brown algae) | Stilophora rhizodes | 98 | Kapraun (2005) | |||
| Bryophyte | Holomitrium arboreum | 167 | Voglmayr (2000) | |||
| Lycophyte | Selaginella kraussiana | 157 | Obermayer et al. (2002) | |||
| Angiosperm | Arabidopsis thaliana | 157 | Bennett et al. (2003) | |||
The use of the base-specific fluorochrome DAPI for estimating DNA amounts may be less reliable than using intercalating fluorochromes such as propidium iodide (e.g. Doležel et al., 1992).
PROSPECTS FOR THE NEXT TEN YEARS
Apart from better defining the limits of genome size variation, what key developments are targeted, or likely, to occur in angiosperm genome size research in the next decade?
The first concerns the expected progress to increase the total number and representation of angiosperms in the C-values database. As noted above, estimating first values for species reached a historic high during recent years (Fig. 1). At least 1700 such values were added in 1997–2002, and the total number of species' C-value estimates probably reached around 4300. In 2003 the second Plant Genome Size Workshop set a goal of estimating a further 1 % (i.e. approx. 2500 species) in the next five years, and a similar target is likely for the following quinquennium. If so, there is a reasonable prospect that the number of species with a C-value estimate will reach, or significantly exceed 7500 by 2014.
More important than the expected increase in total numbers is the predicted improvement in the spread of new values across taxa, geographical regions and life forms, making the sample more representative of the global angiosperm flora, based on careful targeting to identify and fill knowledge gaps. The next decade should see almost complete representation for families, and a greatly increased representation for genera (especially in monocots), as work focuses increasingly at this taxonomic level. Representation at the generic level is currently approx. 1042 out of an estimated 14000 genera (7·4 %) and is targeted to rise to 10 % by 2009, and might reasonably be expected to approach 15 % within a decade. Moreover, this may approach 100 % for monocots, as they are targeted for holistic genomic studies (including C-values) for the global Monocot Checklist Project (Govaerts, 2004).
Recent experience shows that identifying a gap and setting a target may still not provoke the work needed to fill it. Positive monitoring of trends in published C-value data may also be required to achieve a significant change in research activity (e.g. as with the level of family representation in angiosperms; Hanson et al., 2003). Thus it will be important to monitor by 2009 whether the gaping chasms in the representation of African, South American and Chinese floras noted previously have yet resulted in a significant rise in first estimates for taxa from those regions. If not, then a major effort will be needed to correct this. The same applies to other groups of plants that have been identified as poorly represented in the Angiosperm DNA C-values database (e.g. halophytes, parasitic species and their hosts and tundra species). Whilst less certain, there is a good prospect that this vital process will occur in the next decade, driven by the Genome Size Initiative (GESI: see Bennett and Leitch, 2005).
Hitherto, when a question regarding C-value was framed (e.g. is genome size related to weediness?), it was often necessary to estimate C-values for many species before it could be addressed (Bennett et al., 1998). Clearly, the prime aim is to create a sample of C-values that is sufficiently representative for systematic, regional and life form variation as to allow most questions to be answered with confidence using the available dataset, without recourse to further C-value estimations. This goal is likely to be achieved in the next ten years. Thus, the next decade may be the last to see major efforts devoted to estimating first DNA C-values for taxa. Thereafter, new C-value research will probably concentrate on using and understanding such data, rather than acquiring them.
What important questions regarding genome size in angiosperms are likely to be answered in the next decade? Three closely interrelated issues concerning the possible significance of genome size for extinction, conservation and pollution are worth mentioning here.
The possibility that a large C-value might correlate with an increased risk and rate of extinction was suggested by Rejmanek (1996) and by Bennett et al. (2000). To test this, Vinogradov (2003) identified 3036 diploid species from the Plant DNA C-values database and compared each one against the United Nations Environmental Programme World Conservation Monitoring Centre (UNEP-WCMC) species database to determine its conservation status (i.e. global concern, local concern or no concern). He noted a striking relationship between genome size and conservation status; species with large genomes appeared to be at greater risk of extinction that those with smaller genomes.
Clearly, this was an important finding that now requires independent confirmation, drawn from further independent samples of species in different local regions and environments. Obtaining data for meaningfully large samples of species for such studies will probably be one main driver determining which taxa are targeted for C-value estimates in the future. If so, the next decade offers the prospect of a more definite and detailed understanding of any relationships between C-value and/or genome size and the risk of extinction. This, in turn, may have important practical and theoretical implications for conservation models and strategies. A key question is whether a large nuclear DNA amount gives an increased risk of extinction equally in diploid or polyploid taxa? Vinogradov (2003) tested whether ploidy played a role in increasing a species' risk of extinction, concluding that C-value per se was most important. Polyploidy is supposed to confer many advantages based on increased gene dosage and diversity, but do such advantages overcome the possible risks of a high C-value? If so, the proportion of polyploids should be higher for species with very high C-values than for those with lower C-values. In a test different from that of Vinogradov (2003), we compared the percentage of polyploids in 3400 extant species with known DNA amount and ploidy level in the Angiosperm DNA C-values database, ranked in order of increasing DNA amount and divided into five groups each containing 680 species. We found the percentage of polyploids for species in group 5 with the highest C-values (29·9 %) was actually lower than for species in group 4 (32·1 %) (Fig. 4). This confirms Vinogradov's finding that the prime factor determining increased risk of extinction is high C-value, and that polyploidy does not reduce this risk.
Fig. 4.

The percentage of diploids (open bars) and polyploids (closed bars) among 3400 species of known DNA amount and ploidy level ranked in order of increasing DNA amount and divided into five groups with 680 species per group. Data taken from the Angiosperm DNA C-values database (release 4.0, January 2003) and the present Appendix.
Other enquiries should test whether the risk of extinction in relation to high C-value or genome size varies for different threats and environments. This should compare variation in internal factors affecting the structure and ecology of the genome (e.g. increased ploidy level, and heterochromatin distribution), and in external factors (e.g. pollution and increased competition for space, minerals, light, and pollinators). Vilhar (pers. comm., and Vidic et al., 2003) investigated the effect of genome size on plant survival in lead-polluted soils. With increasing lead concentration in the soil the percentage of species with large genomes decreased significantly, suggesting that species with large genomes were at a selective disadvantage. Similar work on local floras in different areas with various threats is now needed to test whether their results are typical for other pollutants and environments. Understanding which species survive locally is always important, but especially as local loss equals global extinction if a species range is restricted to just that one locality. Such work will increasingly inform local environmental action plans and conservation strategies.
Holistic genomics
Early interest in plant genome size variation (c. 1950s and 1960s) ranged broadly across many fields including its genetic, developmental, ecological and evolutionary implications. However, after the molecular revolution the field fragmented somewhat as interest in DNA sequences was largely separated from more macro interests in C-values. However, given ‘complete sequences’ for genomes and homoeologous segments, and greater computing power, this post-genomic age is seeing a strong convergence of these interests. Thus, leading scientists who work at comparative sequence levels can also work on questions of genome size and evolution (e.g. Zhang and Wessler, 2004; Bennetzen et al., 2005). This is the age of holistic genomics in which knowledge of variation in genome size and C-value can be seamlessly joined up with information at all other levels to embrace information from sequences to ecology and from evolution to the environment. This powerful approach should permit or provoke quantum leaps in understanding the significance of extant variation in C-value and genome size, the processes that produce it, the rate at which it occurs, the factors that limit its extent and the advantages and disadvantages that it confers. Together, such understanding will link across biological fields to explain patterns of genome size variation in development, floras, ecological niches and evolution. The next ten years offer many exciting prospects for angiosperm genome size research. Work on DNA amount will remain a key core interest in biological research, but will increasingly become one integrated strand in holistic genomic studies and understanding, covering its origin(s), mechanisms of change, phenotypic and phenological effects, and its significance for ecological, developmental and environmental issues.
APPENDIX
Notes to the Appendix
The Appendix appears on pp. 59–88.
Named references in the following notes are given above in ‘Literature Cited’, while numbered references are given in ‘Original references for DNA values’ below.
(a) The original references for species DNA amounts in the Appendix are given in a numbered list following the Appendix table. Reference numbers follow on sequentially from those given in ‘Notes to Table 8’ by Bennett and Smith (1976, references 1–54) ‘Notes to Table 1’ by Bennett et al. (1982, references 55–107), Bennett and Smith (1991, references 108–163), ‘Notes to the Appendix’ by Bennett and Leitch (1995, references 164–269), ‘Notes to the Appendix’ by Bennett and Leitch (1997, references 270–306), and ‘Notes to the Appendix’ by Bennett et al. (2000, references 307–377).
(b1) Bennett and Smith (1991) gave absolute 4C DNA values for 11 angiosperm species recommended for use as calibration standards to estimate DNA amounts in other species. These species and their 4C DNA amounts are given in Table 4. If a species was calibrated in direct comparison with any one or more of the 11 standard species then the standard species used is identified in column 15 of the Appendix by the appropriate Key letter given above (e.g. F is Hordeum vulgare, etc.). If a species was first calibrated using a standard species listed above, then the original standard species is identified first and the intermediate standard species used to calibrate those species listed with it is also denoted by its number in column 1 of the Appendix. For instance, standard G (P. sativum) was used to calibrate Capsicum annuum ‘Doux Long des Landes’ (species 212 h in the Appendix), which was then used as an intermediate standard to estimate other Capsicum species given by Belletti et al. (1998, Ref. 434). The calibration standard for such Capsicum species is therefore given as G-212h.
Table 4.
The eleven angiosperm species recommended for use as calibration standards (see Notes to the Appendix, b1)
| Key |
Standard species |
4C DNA amount (pg) |
|---|---|---|
| A | Triticum aestivum ‘Chinese Spring’ | 69·27 |
| B | Allium cepa ‘Ailsa Craig’ | 67·00 |
| C | Vicia faba PBI, inbred line 6 | 53·31 |
| D | Anemone virginiana line AV 200 | 35·67 |
| E | Secale cereale ‘Petkus Spring’ | 33·14 |
| F | Hordeum vulgare ‘Sultan’ | 22·24 |
| G | Pisum sativum ‘Minerva Maple’ | 19·46 |
| H | Zea mays ‘W64A’ | 10·93 |
| I | Senecio vulgaris (PBI population) | 6·33 |
| J | Vigna radiata ‘Berken’ | 2·12 |
| K | Oryza sativa ‘IR36’ | 2·02 |
(b2) In Refs 444 (Wendel et al., 2002) and 447 (Lin et al., 2001) Pisum sativum ‘Minerva Maple’ was used as the calibration standard but they assumed a 4C DNA value of 19·12 pg (Johnston et al., 1999) instead of 19·46 pg, which is the value given in Bennett and Smith (1976) and listed in Table 4. The 4C-value of P. sativum ‘Minerva Maple’ used in Refs 444 and 447 was estimated using Hordeum vulgare ‘Sultan’ as the calibration standard with an assumed 4C DNA content of 22·24 pg (Johnston et al., 1999).
(c) In several references listed in ‘Original references for DNA values’ the authors used a cultivar of a standard species different from that listed in Table 4, these are listed in Table 5. In some cases the C-value of the cultivar used was assumed to be the same as that of the cultivar given in Table 4. Evidence of intraspecific variation in a number of species suggests that such assumptions may sometimes be incorrect. In other cases the C-value of the cultivar was determined by the authors and was different from that of the standard species listed in Table 4. For example Refs 386, 397, and 453 used the cultivar ‘Express Long’ of Pisum sativum with a 4C DNA value of 16·74 pg. This value is lower than the 4C DNA amount of the cultivar ‘Minerva Maple’ of 19·46 pg given in Table 4.
Table 5.
Cultivars of standard species used that differ from those listed in Table 4
| Original reference number |
Plant calibration standard used |
Assumed 4C DNA amount and reference (pg) |
|
|---|---|---|---|
| Allium cepa | |||
| 413 | ‘Frühstamm’ | 67·0 (reference not given) | |
| 409 | ‘Kantar topu’ | 67·0 (Van't Hof, 1965; Bennett and Smith, 1976) | |
| 435, 443, 449, 454, 455 | ‘Nasik Red’ | 67·0 (Van't Hof, 1965) | |
| 427 | ‘Stuttgarter Reisen’ | 67·0 (Bennett and Smith, 1976) | |
| 388, 411, 420, 422 | var. aggregata | – (amount and reference not given) | |
| Hordeum vulgare | |||
| 417 | ‘Stark’ | 21·36 (reference not given) | |
| 423 | strain NE 86954 | 20·48 (Lee et al., 1997) | |
| Oryza sativa | |||
| 424 | type japonica | 2·20 (Bennett and Smith, 1991) | |
| Pisum sativum | |||
| 386, 397, 453, 393, 394, 395, 396, 429, 458 | ‘Express Long’ | 16·74 (Marie and Brown, 1993) | |
| ‘Kleine Rheinländerin’ | 17·68 (Greilhuber and Ebert, 1994) | ||
| 434 | ‘Lincoln’ | 18·14 (Doležel et al., 1992) | |
| 418 | ssp. sativum convar. sativum var. ponderosum ‘Viktoria’ | 18·18 (Doležel et al., 1998) | |
| Secale cereale | |||
| 418 | ssp. cereale | 32·38 (Doležel et al., 1998) | |
| 394 | ‘Dankovske’ | 31·16 (Doležel et al., 1998) | |
| Vicia faba | |||
| 418 | ssp. minor var. minor subvar. rigida ‘Tinova’ | 54·00 (Doležel et al., 1998) | |
| 405 | ‘Aquadulce’ | 53·31 (pers. comm. 2002) | |
| 430 | ‘Superguadulce’ | 53·30 (Bennett and Smith, 1976) | |
| Zea mays | |||
| 391 | ‘CE-777’ | 10·86 (Lysák and Doležel, 1998) | |
| 437 | ‘C-777’ | 10·86 (Lysák and Doležel, 1998) | |
| 382 | Va35 | 10·93 (Bennett and Leitch, 1995) | |
(d) In References 407, 408 and 457 the cultivar of the calibration standard was not given. Refs 407 and 408 used Vicia faba as a calibration standard, whereas Ref. 457 used Allium cepa. In Ref. 408 Cremonini et al. (1992) assumed the same 4C-value for Vicia faba as for PBI line 6 (i.e. 53·3 pg) given in Table 4. If this species exhibits intraspecific variation then such assumptions may be incorrect.
(e) In a number of original references the authors used a plant species not listed in Table 4 as a calibration standard. These are listed in Table 6.
Table 6.
Plant species used as calibration standards but not listed in Table 4
| Original reference number |
Plant calibration standard used |
Assumed 4C DNA amount and reference (pg) |
Abbreviation used in column 15 of Appendix |
|---|---|---|---|
| 383 | Agave americana | 31·80 (Zonneveld and Van Iren, 2001) | Agave |
| Arabidopsis thaliana | |||
| 389 | ‘Columbia’ | – (amount and reference not given) | Arab. |
| 414 | ‘Columbia’ | 0·53 (Kaneko et al., 1998) | ” |
| 427 | Cerastium eriophorum | 5·20 (Boscaiu et al., 1999) | Cerastium |
| Glycine max | |||
| 393 | ‘Ceresia’ | 4·54 (Greilhuber and Obermayer, 1997) | Glycine |
| 429 | ‘Ceresia’ | 4·51 (Obermayer and Greilhuber, 1999) | ” |
| 421 | var. Palmetto | 5·00 (Doležel et al., 1994) | ” |
| 432 | ‘Burlison’ | 5·56 (Graham et al., 1994) | ” |
| 402 | ‘Polanka’ | 5·00 (Doležel et al., 1994) | ” |
| Lycopersicon esculentum | |||
| 380, 382, 465 | ‘Gardener's Delight’ | 4·00 (Obermayer et al., 2002) | Lycopers. |
| 437 | ‘Stukické’ | 3·92 (Doležel et al., 1992) | ” |
| 385 | ‘Montfavet’ | 4·02 (Marie and Brown, 1993) | ” |
| Nicotiana tabacum | |||
| 427 | ‘Petit Havana SR1’ | 18·00 (Bennett and Leitch, 1995) | Nicot. |
| 417 | ‘Samsun’ | 18·15 (reference not given) | ” |
| Petunia hybrida | |||
| 390, 410, 433, 452 | ‘PxPC6’ | 5·70 (Godelle et al., 1993; Marie and Brown, 1993) | Petunia |
| 390 | ‘Hit Parade Blau’ R | 5·70 (Marie and Brown, 1993) | ” |
| 401 | No cultivar given | 5·70 (Marie and Brown, 1993) | ” |
| Rhaphanus stativus | |||
| 404 | ‘Saxa’ | 2·20 (Doležel et al., 1992) | Rhaphanus |
| Sorghum bicolor | |||
| 439 | Line TX623 | 3·52 Price and Levin (pers. comm.) | Sorghum |
| 462 | No cultivar given | 3·20 (Bennett and Smith, 1991) | ” |
| 406 | Vicia narbonensis | 29·10 (Frediani et al., 1992) | Vicia narb. |
(f) Several papers listed in ‘Original references for DNA values’ used animal cells as the calibration standards. Thus Refs 387, 417, 426, 442, 456, 463 used chicken erythrocytes with an assumed 4C DNA value of 4·66 pg (Galbraith et al., 1983). The calibration standard is abbreviated to Gallus in column 15 of the Appendix. In Ref. 438 blood cells from the catfish Ictalurus punctatus were used as a standard with an assumed 4C-value of 4·00 pg (Tiersch et al., 1989), this is abbreviated to Ictal. in the Appendix. In Ref. 438 domestic swine (Sus scrofa) erythrocytes were used as a standard with an assumed 4C-value of 11·34 pg (Taliaferro et al., 1997), and is abbreviated to Sus in the Appendix. Human cells with an assumed 4C DNA amount of 14·00 pg (Tiersch et al., 1989) were used as calibration standards in Refs 384, 428 and 419 (leucocytes, Ref. 384, 428; lymphocytes, Ref. 419) and the abbreviation of Homo is used in the Appendix. Finally, Drosophila melanogaster with an assumed 1C DNA amount of 180 Mb (Adams et al., 2000) and Caenorhabditis elegans with a 1C DNA amount of 100·25 Mb, based on complete genome sequencing (see C. elegans Sequencing Consortium, 1998 and http://wormbase.org), were used as calibration standards for Ref. 461, and the abbreviation of Dros. and Caeno. respectively are used.
If a plant species was calibrated using an animal species and then subsequently used as the calibration species for other plants, then the animal species is identified first, and the intermediate plant species is identified by its entry number given in column 1 of the Appendix. Thus Mishiba et al. (2000, Ref. 387) used Gallus with an assumed 4C DNA amount of 4·66 pg (Galbraith et al., 1983) to calibrate Hordeum vulgare ‘New Golden’ (species 398p in Appendix), this was then used as the calibration standard to estimate DNA C-values of Petunia and Calibrachoa species given by Mishiba et al. (2000). The calibration standard for these Petunia and Calibrachoa species is given as Gallus-398p.
(g) When a new estimate (or estimates) is given for a species or subspecies already listed by Bennett and Smith (1976, 1991), Bennett et al. (1982, 2000) or Bennett and Leitch (1995, 1997), the estimate is given a number and a lower case letter in column 1 of the Appendix. An ‘a’ implies that the value is preferred to any estimate for that species listed previously by the first author. Where several estimates are available for the same species, the ‘a’ value would automatically be chosen in any arithmetical or statistical calculations. In this context, single estimates for species and ‘a’ values are referred to as ‘prime entries’.
(h) Intraspecific variation in nuclear DNA amount is claimed to occur in this species. Consequently the values given in the Appendix should not be assumed to be correct for all accessions of the species. Where several C-values are listed for a single species with the same ploidy level or chromosome number within a taxon, then only the minimum and maximum values reported from a single reference are listed in the Appendix.
(i) A range of DNA amounts was reported for this species in the reference cited in column 13 of the Appendix. Intraspecific variation was not claimed to occur, so the nature of this variation is unclear. Where estimates differed by more than 10 % the minimum and maximum values are given for the same ploidy level or chromosome number in the Appendix, otherwise only the highest value is given.
(j) According to the International Code of Botanical Nomenclature (Greuter et al., 1994), the names of plant families must end in -aceae. However, eight plant families are exceptions in that each has two alternative names, both of which are correct under the Botanical Code. One is a standard name, ending in -aceae, the other is an exception, sanctioned by long usage. These and their alternatives are the following: Palmae (Arecaceae), Gramineae (Poaceae), Cruciferae (Brassicaceae), Leguminosae (Fabaceae), Guttiferae (Clusiaceae), Umbelliferae (Apiaceae), Labiatae (Lamiaceae) and Compositae (Asteraceae). To be consistent with previous DNA lists (Bennett and Smith, 1976, 1991; Bennett et al., 1982, 2000; Bennett and Leitch, 1995, 1997) the ‘non-standard’ plant names are retained in the present work.
(k) Recent cladistic analysis using both molecular and non-molecular phylogenetic data has resulted in a revised classification of families by the Angiosperm Phylogeny Group (APG) (APG II, 2003). Familial names used in the APG classification are followed in the Appendix. Thus, although Zonneveld (2002, Ref. 440) placed Aloe in Aloeaceae, recent molecular and non-molecular phylogenetic data recognizes that this family is embedded within the newly circumscribed Xanthorrhoeaceae (APG II, 2003) so Xanthorrhoeaceae is given in the Appendix. Similarly, the APG II (2003) now recognizes that Hostaceae is embedded within the Asparagaceae, so Hosta, which was placed in Hostaceae in Ref. 384 (Zonneveld and Van Iren, 2000), is listed under Asparagaceae in the Appendix.
(l) The authority for this species is either unknown or unclear to the present authors.
(m) Whether or not voucher specimens exist for this species is unknown to the present authors.
(n) The chromosome number of this species is either unknown or unclear to the present authors.
(o) The chromosome count for this species was taken from the literature and not determined by the authors of the reference cited.
(p) The ploidy level of this species is either uncertain or unclear to the present authors.
(q) The life cycle type of this species is either unknown or unclear to the present authors.
(r) The method used to measure the DNA amount is unclear.
(s) The factor of 1 pg = 980 Mbp was used to convert picograms to Mbp (Cavalier-Smith, 1985; Bennett et al., 2000).
(t) As a rule, replicated diplophase nuclei contain a 4C DNA amount producing two unreplicated 2C nuclei by mitotic division and four 1C gametic nuclei after meiosis (irrespective of ploidy level). This convention applies well to polyploid taxa with diploidized meiotic chromosome pairing which produce functional balanced polyhaploid gametes with 1C DNA amounts at meiosis. Thus 4C estimates were automatically divided by 4 to generate 1C-values given for all taxa of even ploidy level listed in the Appendix. However the resulting ‘1C’ data are not biologically meaningful for taxa with odd ploidies. Consequently the Appendix gives only 2C- and 4C-values for such taxa.
(u) There is no obvious basic number for the genus Luzula due to the presence of holocentric chromosomes. It is therefore impossible to allocate Luzula species with high chromosome numbers to any ploidy level with certainty.
(v) Unal and Callow (1995, Ref. 412) obtained a regression of the nuclear fluorescence of Allium cepa (4C = 67·0 pg), Crepis capillaris (4C = 9·6 pg), Hordeum vulgare (4C = 22·2 pg), Pisum sativum (4C = 20·2 pg), Secale cereale (4C = 33·2 pg) and Vicia faba (4C = 47·9 pg) versus nuclear DNA content, and used this to estimate the DNA C-values of 13 Lathryus species. However, it is noted that the 4C-values for P. sativum, and Vicia faba are non-standard values compared with those given for these species in footnote (b1) above.
(w) The standard species used to convert arbitrary units into absolute DNA amounts is unclear to the present authors.
(x) The DNA value given for this species in the original reference differs considerably (i.e. >100 %) from that given in other original references cited in previous compiled lists of DNA amounts (i.e. Bennett and Smith, 1976, 1991; Bennett et al., 1982, 2000; Bennett and Leitch, 1995, 1997). The reason(s) for this is unknown. This C-value should therefore be used with caution until the question is resolved.
(y) The specific status of the material available for study is unclear. The data are included since information on DNA amounts for this genus is relatively sparse, so an indication of genome size in the genus may be useful.
(z) Zonneveld (2001, Ref. 383) gave C-values for 16 hybrid cultivars which fall within the range that he reported for Helleborus species listed in the Appendix. Our compiled lists have usually been restricted to C-values for species. Following this practice, C-values for Helleborus hybrids were not included in the present Appendix.
(aa) Zonneveld and Van Iren (2000, Ref. 384) gave DNA amounts for 94 accessions of Hosta which were recognised as 23 different species. Their table 1 gives a DNA amount for each accession together with a mean value for each recognised species. Only the later value is given in the present Appendix. They also included C-values for 16 Hosta cultivars (in their table 3). These were once recognized as species, but following pollen viability tests Zonneveld and Van Iren (2000) concluded they were hybrids. Our compiled lists have usually been restricted to C-values for species. Accordingly, C-values for Hosta hybrids were not included in the present Appendix.
(ab) Zonneveld and Van Iren (2000, Ref. 384) and Zonneveld (2002, Ref. 440) used male human leucocytes (2C = 7·0 pg; Tiersch et al., 1989) as their primary standard to estimate the DNA amount of three Agave species, namely: (i) Agave stricta (ii) A. americana and (iii) A. sisalana. A. americana was then used as internal calibration standard for most Hosta (Ref. 384) and Aloe (Ref. 440) taxa. However, in a few cases where the DNA content of Hosta or Aloe coincided with that of A. americana, one of the other two Agave species was used as the internal standard. As neither reference identified which Agave species was used, the calibration standard in column 15 of the present Appendix is given as Agave sp.
(ac) Thibault (1998, Ref. 385) claimed intraspecific variation ranging from 6 to 11 % in the Salix species he studied, but only a mean DNA C-value for each species was given in his table 3. It is these values that are listed in the present Appendix. Thibault (loc. cit.) also included C-values for five hybrids. Our compiled lists have usually been restricted to C-values for species, thus C-values for Salix hybrids are not included in the Appendix. In addition, Thibault (1998, Ref. 385) listed a C-value for ‘S. triandra?’ but concluded its identity was ‘hard to specify’. Consequently, this taxon was not included in the Appendix.
(ad) Thibault (1998, Ref. 385) used DNA C-values to predict the ploidy level of each Salix species given in his table 3, assuming direct proportionality. Moreover, their chromosome numbers were not counted by him, but derived by him assuming a constant basic chromosome number of n = 19 for the genus. These predictions are entered in columns 6 and 7 of the Appendix.
(ae) Some taxa once included in Petunia are now included in Calibrachoa. The taxonomy for most Petunia species listed in Mishiba et al. (2000, Ref. 387) follows that of Wijsman (1990) who split the genus Petunia sensu Jussieu (1803) into two; Petunia sensu Wijsman and Calibrachoa. However, five species listed in Mishiba et al. (2000) were not reclassified by Wijsman and so they were listed under the generic name of Petunia sensu Jussieu in Mishiba et al. (2000) although they ‘were regarded as Calibrachoa’ (see their table 3). By following the taxonomy of Petunia sensu Wijsman, several species originally listed under the genus Petunia, now belong to Calibrachoa (e.g. parviflora was listed in the genus Petunia by White and Rees, 1985, 1987) and this generic name was used in the list of Bennett and Leitch (1995). Yet Mishiba et al. (2000) assigned this species to Calibrachoa. To avoid confusion readers looking under Petunia are referred to Calibrachoa in the Appendix.
(af) Joachimiak et al. (2001, Ref. 391) reported chromosome numbers and C-values for six Bromus species. Chromosome numbers varied considerably in roots of three species, but variation in C-values was ‘virtually absent within leaf mesophyll cells’. The C-values given by Joachimiak et al. (2001) were obtained using leaf mesophyll cells and are listed in the Appendix.
(ag) The study by Rosato et al. (1998, Ref. 392) was primarily concerned with polymorphism in Zea mays ssp. mays races with B-chromosomes, but gave C-values only for plants lacking B-chromosomes. Thus, they listed DNA amounts for 17 populations which differed by 36 % (2C = 5·008–6·757 pg) in plants with 2n = 20. Similar intraspecific variation in this species was reported previously (Laurie and Bennett, 1985; Rayburn et al., 1985). Mean DNA amounts for only the populations with the largest and smallest C-values for A-chromosomes, are listed in the Appendix.
(ah) Dimitrova and Greilhuber (2000, Ref. 394) reported significant intraspecific variation in Crepis biennis (P < 0·05) and C. sancta (P < 0·01), some of which had variable numbers of B-chromosomes. As only means were given for material with 0–2 B-chromosomes, it was impossible to give values (presumably the largest) for the 2B complement. Consequently, the Appendix just lists the smallest and largest C-values for accessions without B-chromosomes.
(ai) Dimitrova and Greilhuber (2000, Ref. 394) reported significant (P < 0·001) intraspecific variation of 11 % for Crepis pulchra. They suggested that the two accessions with the higher C-values may belong to subspecies turkestanica. This is not recognized in the Bulgarian flora (where these accessions were collected), but was described by Babcock (1947). In the Appendix the higher C-values listed for this species (entry numbers 305c and e) may thus correspond to C. pulchra ssp. turkestanica.
(aj) Temsch and Greilhuber (2000, Ref. 395) estimated C-values in 11 accessions of Arachis hypogaea using both Feulgen microdensitometry and flow cytometry. C-values for different accessions showed great stability, so they calculated a mean C-value for each method in the ‘Results and Discussion’ of their paper. Only these mean values are listed in the Appendix.
(ak) Previous estimates for Vicia melanops (2n = 10) (e.g. Chooi, 1971; Raina and Rees, 1983; Raina and Bisht, 1988) all report a 4C-value of approx. 40 pg, which is much higher than the value of 27·6 pg given in Cremonini et al. (1992, Ref. 408, entry number 780d). Thus, this estimate should be viewed with caution until confirmed independently.
(al) Akpinar and Bilaloglu (1997, Ref. 409) gave a 2C-value of 13·1 pg for Vicia cracca ssp. cracca (with 2n = 2x = 14; their original count). However, six previous reports for V. cracca listed in the database (Bennett and Leitch, 2003) gave similar 2C-values (from 10 to 13 pg), but for 2n = 4x = 28. The cause of this discrepancy is unknown, thus the estimate by Akpinar and Bilaloglu (loc. cit.) should be viewed with caution until confirmed independently.
(am) Sakamoto et al. (1998, Ref. 414) estimated the C-value of Cannabis sativa using Arabidopsis thaliana ‘Columbia’ (1C = 130 Mb, Kaneko et al., 1998) as the calibration standard. However, the 1C-value assumed for A. thaliana was low compared with its recently confirmed estimate of 157 Mb (Bennett et al., 2003). If 157 Mb is assumed for A. thaliana, then the 1C-value for C. sativa would be 988 Mb = 1·01 pg (female) and 1016 Mb = 1·04 pg (male).
(an) Gammar et al. (1999, Ref. 416) gave DNA amounts for eight Lupinus species in arbitrary units (a.u.) listed as Mn(x) values in their Figs. 1–4. Bennett and Smith (1976) gave the 4C DNA amount of L. luteus as 4·0 pg (allowing for recalibration of Senecio vulgaris from 5·88 pg to 6·33 pg, see Bennett and Smith, 1991). Gammar et al. (loc. cit.) gave Mn(x) values for three L. luteus populations as 60·4, 58·4, and 63·8 in figure 1A, noting they were not statistically different. The mean of these three values was calculated to be 60·86 a.u. To convert the Mn(x) values for each Lupinus species into absolute DNA amounts, they were multiplied by a conversion factor of 0·07 (i.e. 4·0 pg ÷ 60·86 a.u.).
In some Lupinus species more than one population was studied, and several Mn(x) values were listed. If these did not differ significantly, the average Mn(x) value was calculated and converted into absolute DNA amounts. However, chromosome counts of 2n = 38, 42 and 44 were reported in L. angustifolius, so variation in Mn(x) may correspond to different cytotypes.
Some absolute DNA amounts calculated for Gammar et al. (loc. cit) do differ greatly from those previously reported for the same species (e.g. L. pilosus, 4C = 4·9 pg, is almost double the value of 2·5 pg given by Obermayer et al., 1999). Similarly, the 4C-value of 3·1 pg calculated for L. angustifolius with 2n = 38, 42, or 44 is similar to the estimate (4C = 3·7 pg) by Barlow (pers. comm., listed in Bennett et al., 1982), yet the latter was for material with 2n = 26. Data from Gammer et al. give a useful approximation of C-values in the five species not previously listed, but should be treated with caution unless confirmed independently.
(ao) Brandizzi and Grilli Caiola (1996, Ref. 419) gave 2n = 18 for Crocus biflorus in their table 1, but 2n = 8 in the first paragraph of their text. They also stated in their final paragraph: ‘However, C. biflorus and C. etruscus, having half the chromosome number with respect to C. thomasii and C. cartwrightianus…..’ As C. thomasii and C. cartwrightianus were both recorded with 2n = 16 by Brandizzi and Grilli Ciola (1996), we conclude that C. biflorus had 2n = 8, and so this number is entered in the Appendix.
(ap) The 4C DNA amounts reported by Mukerjee and Sharma (1993a, Ref. 420) for Luzula nivea and L. luzuloides are over 50 % larger than those reported by Barlow (pers. comm. 1976; reference 36 in Bennett and Smith, 1976). The chromosome numbers for each species were the same, so the cause of the discrepancy is unknown. However, Mukerjee and Sharma (1993a) used a single wavelength method, which may suffer from distributional error (Greilhuber, 2005, this volume). Thus, estimates for Luzula in Mukerjee and Sharma (1993a) should be viewed with caution until confirmed independently for these species.
(aq) Asif et al. (2001, Ref. 421) estimated DNA amounts in 14 genotypes of Musa acuminata. Genotype BC3 (belonging to the separate subspecies truncata) had the highest DNA amount and its C-value was shown to be significantly different (P < 0·01) from the other thirteen genotypes. Only the DNA amount of genotype BC3, corresponding to M. acuminata ssp. truncata, and the highest DNA amount out of the 13 other genotypes of M. acuminata are entered in the Appendix.
(ar) Chaudhuri and Sen (2001, Ref. 422) examined two Scilla indica cytotypes (entry numbers 710b and c) which differed considerably in both DNA amount and karyotype structure, although both had 2n = 30. The differences may reflect problems with taxonomy. Studies by Greilhuber and colleagues (Greilhuber, 1979; Greilhuber and Speta, 1985) have shown that large intraspecific differences in C-values in other Scilla species (e.g. S. bifolia) reduce to a level hardly more than methodological error following taxonomic splitting.
(as) Chung et al. (1998, Ref. 423) estimated C-values in 12 soybean (Glycine max) strains varying in seed size. They reported statistically significant differences of 4·6 % in the 2C-values between strains. Only the smallest and largest C-values are entered in the Appendix.
(at) Hartman et al. (2000, Ref. 425) estimated C-values in 22 Leucaena species using flow cytometry. Three species (Pisum sativum 4C = 17·6 pg, Oryza sativa 4C = 1·8 pg and Vicia faba 4C = 53·0 pg) were used as calibration standards at various times, but unfortunately the authors did not state which standard(s) was compared with which Leucaena species.
(au) Boscaiu et al. (1999, Ref. 427) referred to plants of Cerastium with 2n = 36 as diploids in contrast with various other authors who consider them as tetraploids. The assumption was based on Boscaiu et al.'s observations that, while the base chromosome number in Cerastium may be x = 9, no Cerastium species is known with 2n = 18.
(av) The C-values reported for Hedera helix by Obermayer and Greilhuber (1999, Ref. 429) agree well with previous reports by König et al. (1987) of 2C = 3·0 pg, but are only about one third the value reported by Marie and Brown (1993) of 2C = 8·2 pg, which is unsupported.
(aw) Blanco et al. (1996, Ref. 431) gave DNA amounts for Dasypyrum hordaceum and D. villosum in arbitrary units (a.u.), listed as mean values in their fig. 3. The value for D. hordaceum was converted into an absolute DNA amount by multiplying the mean value of 381·7 a.u. by a conversion factor of 0·11. This conversion factor was obtained as the ratio of the 4C estimate for Dasypyrum villosum (listed as the synonym Haynaldia villosa) reported by Bennett (1972) as 21·4 pg, and the estimate of 193·7 a.u. reported by Blanco et al. (1996).
(ax) Rayburn et al. (1997, Ref. 432) estimated C-values in 90 accessions of Glycine max. Accessions showed a 12 % variation in DNA amount and these differences were statistically significant. Only the smallest and largest C-values are listed in the Appendix.
(ay) Comparing C-values given by Belletti et al. (1998, Ref. 434) with those previously published showed DNA amounts for Capsicum baccatum, C. chinese, C. eximium, C. frutescens and C. pubescens were around one third greater than those of Owens (pers. comm.) listed in Bennett and Smith (1976). Belletti et al. (1998) suggested that the cause of the discrepancy could be that Owens used Allium cepa as the calibration standard, whose 2C-value of 33·5 pg differs considerably from those reported in Capsicum species studied.
(az) Široký et al. (2001, Ref. 437) investigated C-values in four Silene species, including S. latifolia, which has previously been listed by Bennett and Leitch (1995) under its synonym Melandrium album.
(ba) Taliaferro et al. (1997, Ref. 438) gave DNA C-values for 18 accessions of Cynodon corresponding to two species: C. transvaalensis (2n = 2x = 18), and C. dactylon var. dactylon (2n = 4x = 36 and 2n = 6x = 54). Only small differences in DNA amounts were noted between five diploid and five tetraploid accessions, and a mean 2C-value for each ploidy level was also given in table 2 of their paper. This mean value is listed in the Appendix. However, the three hexaploid accessions examined comprised one accession of C. dactylon var. dactylon and two hybrids. Thus, only the C-value estimate for hexaploid C. dactylon var. dactylon is entered in the Appendix, rather than the mean for the three hexaploid accessions given in table 2 of Taliaferro et al. (1997).
(bb) Blakesley et al. (2002, Ref. 441) examined seven populations of Acacia dealbata and four of A. mangium to determine ploidy and DNA amount. In A. dealbata, they identified naturally occurring diploid, triploid and tetraploid genotypes. Chromosome numbers were counted in only one diploid and one tetraploid genotype, and C-values for only these populations are given in the Appendix. In naturally occurring A. mangium only diploid populations were found, and the C-value for the only population whose chromosome number was determined is given in the Appendix. C-values for colchicine-induced tetraploid genotypes of A. mangium are not included.
The 2C-value for diploid A. dealbata (1·7 pg) is similar to that reported by Bukhari (1997) as 2C = 1·6 pg. In contrast the 2C-value (2·9 pg) reported by Mukherjee and Sharma (1993b) is nearly twice that of Blakesley et al. (2002). Perhaps this discrepancy reflects the use of Allium cepa (2C = 33·5 pg), whose genome size is over an order of magnitude greater than that of Acacia, as a calibration standard by Mukherjee and Sharma (1993b). Similar discrepancies were noted between DNA estimates for A. mangium reported by Blakesley et al. (loc. cit.) of 2C = 1·3 pg, and those by Mukherjee and Sharma (1995) of 2C = 2·3 pg.
(bc) Ohri and Singh (2002, Ref. 443) listed C-values for 20 wild relatives of cultivated pigeon pea (Cajanus cajan). However, C-values for 14 of these species had already been communicated to MD Bennett in 1996 and listed in the Appendix of Bennett and Leitch (1997) under Original reference number 303. To avoid duplication of data in the database, only C-values for six species not listed previously are included in the Appendix.
(bd) Wendel et al. (2002, Ref. 444) listed DNA amounts for 13 species in the tribe Gossypieae. However, C-values for three of these species had already been communicated to MD Bennett in 1999 and listed in the Appendix of Bennett et al. (2000) under Original reference number 349. To avoid duplication of data in the database, only C-values for ten species not listed previously are included in the Appendix.
(be) The C-value of Arabidopsis thaliana given by the Arabidopsis Genome Initiative (2000, Ref. 448) was based on DNA sequencing data for 115·4 Mb of the genome, plus a guestimate of 10 Mb for several unsequenced gaps in the genome. Recent work places its 1C-value around 157 Mb (Bennett et al., 2003).
(bf) Ohri (2002, Ref. 449) listed DNA amounts for 36 tropical hardwood species belonging to 13 families. However, C-values of 35 of these had already been communicated to MD Bennett in 1996 and listed in the Appendix of Bennett and Leitch (1997) under Original reference number 301. To avoid duplication of data in the database, a C-value for the only species not included in a previous compilation (Drypetes roxburghii) is listed in the present Appendix. A new C-value for Melaleuca leucadendra, double that given in Bennett and Leitch (1997), is also listed in the Appendix, to correct an error in communication which confused the 2C- and 4C-values for this species.
(bg) The C-value of 466 Mb for Oryza sativa ssp. indica given in Yu et al. (2002, Ref. 450) was based on DNA sequencing data for 362 Mb of sequenced scaffolds, and 104 Mb of ‘unassembled data’ subject to numerous assumptions (see Yu et al., loc. cit. – page 80).
(bh) The C-value of 420 Mb for Oryza sativa ssp. japonica given in Goff et al. (2002, Ref. 451) was derived from DNA sequencing data for 389·9 Mb, and their assumption that this equals 93 % of the genome, perhaps using some previously published 1C-value. The source of this assumption, as of any such DNA estimate, and the method by which it was obtained, was not clearly cited by Goff et al. (2002).
(bi) Redondo et al. (1996, Ref. 453) estimated C-values in four populations of Saxifraga granulata. In one population chromosome numbers ranged from 2n = 44 to 56, but 2n = 44 was predominant. They noted that the DNA amount was also variable but gave only one C-value, which is listed in the Appendix. However, intraspecific variation in DNA amount may occur in this species, so the C-value listed may not apply to all members of the population.
(bj) Redondo et al. (1996, Ref. 453) estimated DNA amounts in four populations of Saxifraga granulata. They reported DNA amounts for a population in which they could not obtain a chromosome count (entry number 706), but based on the DNA amount, they suggested this population may have 2n = 30.
(bk) Emshwiller (2002, Ref. 456) estimated C-values in 10 accessions of cultivated oca (2n = 8x = 64; Oxalis tuberosa), two tetraploid wild species, and 78 diploid accessions which were provisionally identified as 35 species. As variation in 2C DNA amounts was usually no more than 0·1 pg, and considered to be technical in nature, only the highest C-value was reported in table 3 of Emshwiller (2002) for most species and is entered in the Appendix. Variation in DNA amounts greater than 0·1 pg was considered real for O. spiralis (2C = 1·062–1·339 pg) and O. peduncularis (2C = 0·927–1·163 pg), so both the lowest and highest values are entered in the Appendix for these species. Emshwiller (2002) noted that this variation may reflect problems of taxonomy and species boundaries.
(bl) Emshwiller (2002, Ref. 456) estimated the DNA amounts in ten accessions of cultivated oca (2n = 8x = 64; Oxalis tuberosa). Variation was noted, even in measurements made for all accessions estimated on one day (see table 3 of Emshwiller, 2002), but she did not consider it to represent intraspecific variation and a mean 2C estimate calculated from all measurements made was given in her ‘Results’ section as 2C = 3·514 pg. It is this value that is entered in the Appendix.
(bm) Nagl et al. (1983, Ref. 457) included DNA amounts for 49 species. However, C-values for 20 of these had been published elsewhere, and already included in previous compilations by Bennett and colleagues (listed under original reference numbers 34, 36, 60, 61, 81, 82, 84, 85, 86). To avoid duplication only C-values for 29 species that had not been listed previously are included in the Appendix.
(bn) Values for Phaseolus coccineus and P. vulgaris given in Nagl et al. (1983, Ref. 457) are around twice those given in another paper by Nagl and Treviranus (1995, Ref. 390), listed in the present Appendix. 2C-values for both species given in Nagl et al. (loc. cit.) agree with those reported by Ayonoadu (1974), but are around twice that reported in Ingle et al. (1975) and Arumuganathan and Earle (1991). The basis of this discrepancy is unclear, so C-values for these Phaseolus taxa should be viewed with caution until confirmed independently.
(bo) The value for Sambucus nigra (2C = 30·5 pg ) given by Nagl et al. (1983, Ref. 457) is similar to the value of 2C = 21·8 pg for a related species, S. racemosa, reported by Nagl et al. (1979) and listed under Ref. 86 in Bennett et al. (1982). However, it is very different from 2C = 3·1 pg reported for S. nigra by Mowforth (1986) and listed under Ref. 158 in Bennett and Smith (1991). The cause for the discrepancy remains unclear, so C-values for S. nigra should be used with caution until confirmed independently.
(bp) Baranyi et al. (1996, Ref. 458) investigated C-values in 75 accessions of four wild Pisum species. Results were given as percentages relative to P. sativum ‘Kleine Rheinländerin’ ( = 100 %) which was used as the calibration standard. To convert these into absolute DNA amounts the 4C-values were multiplied by the value of Pisum sativum ‘Kleine Rheinländerin’ of 17·68 pg (Greilhuber and Ebert, 1994) and then divided by 100.
Pisum fulvum was homogeneous in DNA amount size (4C = approx. 19·3 pg), but wide variation was seen between accessions of the other species studied (P. abyssinicum, P. humile and P. elatius). This variation was interpreted to show that these taxa with variable genome sizes were genetically heterogeneous, suggesting that the current species delimitations did not reflect the true biological species groups adequately. Only the smallest and largest C-values for each of these species are listed in the Appendix.
(bq) Punina and Alexandrova (1992, Ref. 459) estimated DNA amounts in 11 Paeonia species but gave the results as percentage values relative to P. caucasica. Since Mulry and Hanson (pers. comm. 1999) had estimated the 4C DNA value of this species as 65·2 pg (see entry number 602 in Bennett et al., 2000), the relative percentage values given in Punina and Alexandrova (loc. cit.) were converted into absolute DNA amounts by multiplying by 0·652.
(br) A PhD thesis by Shi (1991) gave a 1C-value of 0·15 pg for two accessions of diploid Brachypodium distachyon (2n = 10), plus values for four other Brachypodium species. Later, Shi et al. (1993, Ref. 460) gave the 1C-value for diploid B. distachyon as 1C = 0·3 pg, but cited the PhD thesis (Shi, 1991) as the source for this figure. In order to confirm which was correct, Clive Stace kindly supplied seed of one original accession (B306) and RBG, Kew estimated its DNA amount as 0·36 pg using Oryza sativa ‘IR36’ (4C = 2·02 pg) as a calibration standard (see entry number 161a in the present Appendix). This was much closer to the value in Shi et al. (1993). As C-values for four other Brachypodium species in Shi (1991) may also be under-estimates, they are therefore not included in the present Appendix, and should be viewed with caution until confirmed independently.
Original references for DNA values
Named references in the ‘Notes to the Appendix’ are given in ‘Literature cited’. Only numbered references of original sources of species DNA values in the Appendix (column 13) are given in the Key below.
378. Hanson L, McMahon KA, Johnson MAT, Bennett MD. 2001. First nuclear DNA C-values for 25 angiosperm families. Annals of Botany 87: 251–258.
379. Hanson L, McMahon KA, Johnson MAT, Bennett MD. 2001. First nuclear DNA C-values for another 25 angiosperm families. Annals of Botany 88: 851–858.
380. Hanson L, Brown RL, Boyd A, Johnson MAT, Bennett MD. 2003. First nuclear DNA C-values for 28 angiosperm genera. Annals of Botany 91: 1–8.
381. Leitch IJ, Hanson L. 2002. DNA C-values in seven families fill phylogenetic gaps in the basal angiosperms. Botanical Journal of the Linnean Society 140: 175–179.
382. Obermayer R, Leitch IJ, Hanson L, Bennett MD. 2002. Nuclear DNA C-values in 30 species double the familial representation in pteridophytes. Annals of Botany 90: 209–217.
383. Zonneveld BJM. 2001. Nuclear DNA contents of all species of Helleborus (Ranunculaceae) discriminate between species and sectional divisions. Plant Systematics and Evolution 229: 125–130.
384. Zonneveld BJM, Van Iren F. 2001. Genome size and pollen viability as taxonomic criteria: Application to the genus Hosta Plant Biology 3: 176–185.
385. Thibault J. 1998. Nuclear DNA amount in pure species and hybrid willows (Salix): a flow cytometric investigation. Canadian Journal of Botany 76: 157–165.
386. Torrell M, Valles J. 2001. Genome size in 21 Artemisia L. species (Asteraceae, Anthemideae): Systematic, evolutionary, and ecological implications. Genome 44: 231–238.
387. Mishiba KI, Ando T, Mii M, Watanabe H, Kokubun H, Hashimoto G, Marchesi E. 2000. Nuclear DNA content as an index character discriminating taxa in the genus Petunia sensu Jussieu (Solanaceae). Annals of Botany 85: 665–673.
388. Sen S, Sharma AK. 1990. Chromosome complements, nuclear DNA and genetic distance as measures of interrelationship in Plantago The Nucleus 33: 4–10.
389. Ohmido N, Kijima K, Akiyama Y, de Jong JH, Fukui K. 2000. Quantification of total genomic DNA and selected repetitive sequences reveals concurrent changes in different DNA families in indica and japonica rice. Molecular and General Genetics 263: 388–394.
390. Nagl W, Treviranus A. 1995. A flow cytometric analysis of the nuclear 2C DNA content in 17 Phaseolus species (53 genotypes). Botanica Acta 108: 403–406.
391. Joachimiak A, Kula A, Sliwinska E, Sobieszczanska A. 2001. C-banding and nuclear DNA amount in six Bromus species. Acta Biologica Cracoviensia Series Botanica 43: 105–115.
392. Rosato M, Chiavarino AM, Naranjo CA, Hernandez JC, Poggio L. 1998. Genome size and numerical polymorphism for the B chromosome in races of maize (Zea mays ssp. mays, Poaceae). American Journal of Botany 85: 168–174.
393. Greilhuber J, Obermayer R. 1999. Cryptopolyploidy in Bunias (Brassicaceae) revisited – a flow-cytometric and densitometric study. Plant Systematics and Evolution 218: 1–4.
394. Dimitrova D, Greilhuber J. 2000. Karyotype and DNA-content evolution in ten species of Crepis (Asteraceae) distributed in Bulgaria. Botanical Journal of the Linnean Society 132: 281–297.
395. Temsch EM, Greilhuber J. 2000. Genome size variation in Arachis hypogaea and A. monticola re-evaluated. Genome 43: 449–451.
396. Temsch EM, Greilhuber J. 2001. Genome size in Arachis duranensis: a critical study. Genome 44: 826–830.
397. Cerbah M, Mortreau E, Brown S, Siljak-Yakovlev S, Bertrand H, Lambert C. 2001. Genome size variation and species relationships in the genus Hydrangea Theoretical and Applied Genetics 103: 45–51.
398. Hart H. 1991. Evolution and classification of the European Sedum species (Crassulaceae). Flora Mediterranea 1: 31–61.
399. Hart HT, Tomlik A, Alpinar K. 1993. Biosystematic studies in Sedum (Crassulaceae) fromTurkey. 4. The cytology of Sedum subsect. Spathulata Boriss. Acta Botanica Neerlandica 43: 289–298.
400. Geber G. 1989.Zur Karyosystematik der Lemnaceae. Ph.D. Thesis, University of Vienna, Vienna.
401. Noirot M, Barre P, Louarn J, Duperray C, Hamon S. 2002. Consequences of stoichiometric error on nuclear DNA content evaluation in Coffea liberica var. dewevrei using DAPI and propidium iodide. Annals of Botany 89: 385–389.
402. Lysák MA, Doleželova M, Horry JP, Swennen R, Doležel J. 1999. Flow cytometric analysis of nuclear DNA content in Musa Theoretical and Applied Genetics 98: 1344–1350.
403. Price HJ, Hodnett G, Johnston JS. 2000. Sunflower (Helianthus annuus) leaves contain compounds that reduce nuclear propidium iodide fluorescence. Annals of Botany 86: 929–934.
404. Ellul P, Boscaiu M, Vicente O, Moreno V, Rossello JA. 2002. Intra- and interspecific variation in DNA content in Cistus (Cistaceae). Annals of Botany 90: 345–351.
405. Cremonini R, Castiglione MR, Venora G, Blangiforti S, Losavio FP, Pignone D. 1998. Cytology of Vicia species. VI. Nuclear chromatin organization, karyomorphological analysis and DNA amount in Vicia serratifolia Jacq. Caryologia 51: 195–205.
406. Cremonini R, Miotto D, Ngu MA, Tota D, Pignone D, Blangiforti S, Venora G. 1998. Cytology of Vicia species. 5. Nuclear chromatin structure, karyomorphological analysis and DNA content in newly discovered relatives of Vicia faba L.: Vicia kalakhensis Khattab, Maxted et Bisby and Vicia eristalioides Maxted. Cytologia 63: 371–379.
407. Venora G, Blangiforti S, Castiglione MR, Black-Samuelsson S, Cremonini R. 1999. Cytology of Vicia species. VIII. Nuclear DNA contents, chromatin organization and karyotype evolution in Vicia pisiformis L. populations. Caryologia 52: 105–115.
408. Cremonini R, Funari S, Mazzuca S. 1992. Cytology of Vicia species: nuclear structure, karyological analysis and DNA content. Chromatin 1: 135–146.
409. Akpinar N, Bilaloglu R. 1997. Cytological investigations of certain species of Vicia L. Turkish Journal of Botany 21: 197–207.
410. Kamaté K, Brown S, Durand P, Bureau JM, De Nay D, Trinh TH. 2001. Nuclear DNA content and base composition in 28 taxa of Musa Genome 44: 622–627.
411. Ghosh P, Mukherjee S, Sharma AK. 2001. Cytophotometric estimation of in situ DNA content in several species of Araceae. Cytobios 105: 177–183.
412. Unal F, Callow RS. 1995. Estimation of genome size by Feulgen photometry. Turkish Journal of Botany 19: 523–529.
413. Greilhuber J, Ebert I, Lorenz A, Vyskot B. 2000. Origin of facultative heterochromatin in the endosperm of Gagea lutea (Liliaceae). Protoplasma 212: 217–226.
414. Sakamoto K, Akiyama Y, Fukui K, Kamada H, Satoh S. 1998. Characterization: genome sizes and morphology of sex chromosomes in Hemp (Cannabis sativa L.). Cytologia 63: 459–464.
415. Naranjo CA, Ferrari MR, Palermo AM, Poggio L. 1998. Karyotype, DNA content and meiotic behaviour in five South American species of Vicia (Fabaceae). Annals of Botany 82: 757–764.
416. Gammar ZG, Puech S, Zouaghi M. 1999. Flow cytometry DNA assay of Mediterranean lupins. Candollea 54: 45–56.
417. Arumuganathan K, Tallury SP, Fraser ML, Bruneau AH, Qu R. 1999. Nuclear DNA content of thirteen turfgrass species by flow cytometry. Crop Science 39: 1518–1521.
418. Ali HBM, Meister A, Schubert I. 2000. DNA content, rDNA loci, and DAPI bands reflect the phylogenetic distance between Lathyrus species. Genome 43: 1027–1032.
419. Brandizzi F, Grilli Caiola M. 1996. Quantitative DNA analysis in different Crocus species (Iridaceae) by means of flow cytometry. Giornale Botanico Italiano 130: 643–645.
420. Mukherjee S, Sharma AK. 1993. Cytophotometric DNA estimation in Luzula species. Current Science 65: 987–989.
421. Asif MJ, Mak C, Othman RY. 2001. Characterization of indigenous Musa species based on flow cytometric analysis of ploidy and nuclear DNA content. Caryologia 54: 161–168.
422. Chaudhuri D, Sen S. 2001. Genotypic diversity in species of Scilla Cytobios 104: 75–81.
423. Chung J, Lee JH, Arumuganathan K, Graef GL, Specht JE. 1998. Relationships between nuclear DNA content and seed and leaf size in soybean. Theoretical and Applied Genetics 96: 1064–1068.
424. Cros J, Combes MC, Chabrillange N, Duperray C, Desangles AM, Hamon S. 1995. Nuclear DNA content in the subgenus Coffea (Rubiaceae): inter- and intra-specific variation in African species. Canadian Journal of Botany 73: 14–20.
425. Hartman TPV, Jones J, Blackhall NW, Power JB, Cocking EC, Davey MR. 2000. Cytogenetics, molecular cytogenetics and genome size in Leucaena (Leguminosae, Mimosideae). In: Guttenberger H, Borzan Z, Schlarbaum SE, Hartman TPV, eds. Cytogenetic studies of forest trees and shrubs – review, present status, and outlook on the future. Zvolen, Slovakia: Arbora Publishers, 57–70.
426. Kayim M, Koç NK, Rokka VM. 1998. Variation of the nuclear DNA content of species of subtribe Citrinae (Rutaceae). Hortscience 33: 1247–1250.
427. Boscaiu M, Vicente O, Ehrendorfer F. 1999. Chromosome numbers, karyotypes and nuclear DNA contents from perennial polyploid groups of Cerastium (Caryophyllaceae). Plant Systematics and Evolution 218: 13–21.
428. Lysák MA, Rostková AR, Dixon JM, Rossi G, Doležel J. 2000. Limited genome size variation in Sesleria albicans Annals of Botany 86: 399–403.
429. Obermayer R, Greilhuber J. 2000. Genome size in Hedera helix L.—a clarification. Caryologia 53: 1–4.
430. Caceres ME, De Pace C, Mugnozza GTS, Kotsonis P, Ceccarelli M, Cionini PG. 1998. Genome size variations within Dasypyrum villosum: correlations with chromosomal traits, environmental factors and plant phenotypic characteristics and behaviour in reproduction. Theoretical and Applied Genetics 96: 559–567.
431. Blanco A, Simeone R, Resta P, DePace C, Delre V, Caccia R, Mugnozza GTS, Frediani M, Cremonini R, Cionini PG. 1996. Genomic relationships between Dasypyrum villosum (L) Candargy and D. hordeaceum (Cosson et Durieu) Candargy. Genome 39: 83–92.
432. Rayburn AL, Biradar DP, Bullock DG, Nelson RL, Gourmet C, Wetzel JB. 1997. Nuclear DNA content diversity in Chinese soybean introductions. Annals of Botany 80: 321–325.
433. Gallois A, Burrus M, Brown S. 1999. Evaluation of the nuclear DNA content and GC percent in four varieties of Fagus sylvatica L. Annals of Forest Science 56: 615–618.
434. Belletti P, Marzachi C, Lanteri S. 1998. Flow cytometric measurement of nuclear DNA content in Capsicum (Solanaceae). Plant Systematics and Evolution 209: 85–91.
435. Ohri D. 1999. Cytology of Cicer songaricum Steph ex DC, a wild relative of chickpea. Genetic Resources and Crop Evolution 46: 111–113.
436. Buitendijk JH, Peters A, Quené RJ, Ramanna MS. 1998. Genome size variation and C-band polymorphism in Alstroemeria aurea, A. ligtu and A. magnifica (Alstroemeriaceae). Plant Systematics and Evolution 212: 87–106.
437. Široký J, Lysák MA, Doležel J, Kejnovsky E, Vyskot B. 2001. Heterogeneity of rDNA distribution and genome size in Silene spp. Chromosome Research 9: 387–393.
438. Taliaferro CM, Hopkins AA, Henthorn JC, Murphy CD, Edwards RM. 1997. Use of flow cytometry to estimate ploidy level in Cynodon species. International Turfgrass Society Research Journal 8: 385–392.
439. Price HJ. 2002. (pers. comm. 2002).
440. Zonneveld BJM. 2002. Genome size analysis of selected species of Aloe (Aloaceae) reveals the most primitive species and results in some new combinations. Bradleya 20: 5–12.
441. Blakesley D, Allen A, Pellny TK, Roberts AV. 2002. Natural and induced polyploidy in Acacia dealbata Link. and Acacia mangium Willd. Annals of Botany 90: 391–398.
442. Nehra NS, Kartha KK, Stushnoff C. 1991. Nuclear DNA content and isozyme variation in relation to morphogenic potential of Strawberry (Fragaria × ananassa) callus cultures. Canadian Journal of Botany 69: 239–244.
443. Ohri D, Singh PS. 2002. Karyotypic and genome size variation in Cajanus cajan (L.) Millsp. (pigeonpea) and some wild relatives. Genetic Resources and Crop Evolution 49: 1–10.
444. Wendel JF, Cronn RC, Johnston JS, Price HJ. 2002. Feast and famine in plant genomes. Genetica 115: 37–47.
445. Hiremath SC, Nagasampige MH. (pers. comm. 2001).
446. Hiremath SC, Patil CG, Ksheerasagar AL. (pers. comm. 2001).
447. Lin S, Lee H-C, Chen W-H, Chen C-C, Kao Y-Y, Fu Y-M, Chen Y-H, Lin T-Y. 2001. Nuclear DNA contents of Phalaenopsis sp. and Doritis pulcherrima Journal of the American Society of Horticultural Science 126: 195–199.
448. Arabidopsis Genome Initiative. 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana Nature 408: 796–815.
449. Ohri D. 2002. Genome size variation in some tropical hardwoods. Biologia Plantarum 45: 455–457.
450. Yu J, Hu SN, Wang J, Wong GKS, Li SG et al. 2002. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 296: 79–92.
451. Goff SA, Ricke D, Lan TH, Presting G, Wang RL et al. 2002. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 296: 92–100.
452. Schwencke J, Bureau JM, Crosnier MT, Brown S. 1998. Cytometric determination of genome size and base composition of tree species of three genera of Casuarinaceae. Plant Cell Reports 18: 346–349.
453. Redondo N, Horjales M, Brown S, Villaverde C. 1996. Biometric and cytometric study of nuclear DNA within Saxifraga granulata L. Boletin da Sociedade Broteriana, Séries 2 67: 287–301.
454. Ohri D. (pers. comm. 2002).
455. Ohri D. (pers. comm. 2002).
456. Emshwiller E. 2002. Ploidy levels among species in the ‘Oxalis tuberosa Alliance’ as inferred by flow cytometry. Annals of Botany 89: 741–753.
457. Nagl W, Jeanjour M, Kling H, Kuhner S, Michels I, Muller T, Stein B. 1983. Genome and chromatin organization in higher plants. Biologisches Zentralblatt 102: 129–148.
458. Baranyi M, Greilhuber J, Swiecicki WK. 1996. Genome size in wild Pisum species. Theoretical and Applied Genetics 93: 717–721.
459. Punina EO, Alexandrova TV. 1992. The chromosome volume and relative DNA content in Caucasian representatives of the genus Paeonia Botanichesky Zhurnal 77: 16–23.
460. Shi Y, Draper D, Stace C. 1993. Ribosomal DNA variation and its phylogenetic implication in the genus Brachypodium (Poaceae). Plant Systematics and Evolution 188: 125–138.
461. Bennett MD, Leitch IJ, Price HJ, Johnston JS. 2003. Comparisons with Caenorhabditis (∼100 Mb) and Drosophila (∼175 Mb) using flow cytometry show genome size in Arabidopsis to be ∼157 Mb and thus 25 % larger than the Arabidopsis genome initiative estimate of ∼125 Mb. Annals of Botany 91: 547–557.
462. Bitonti MB, Cozza R, Chiappetta A, Contento A, Minelli S et al. 1999. Amount and organization of the heterochromatin in Olea europaea and related species. Heredity 83: 188–195.
463. Galbraith DW, Harkins KR, Knapp S. 1991. Systemic endopolyploidy in Arabidopsis thaliana Plant Physiology 96: 985–989.
464. Francis DM, Hulbert SH, Michelmore RW. 1990. Genome size and complexity of the obligate fungal pathogen, Bremia lactucae Experimental Mycology 14: 299–309.
465. Hanson L, Leitch IJ, Bennett MD. 2002. Unpublished values from the Jodrell Laboratory, Royal Botanic Gardens, Kew.
LITERATURE CITED
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD et al. 2000. The genome sequence of Drosophila melanogaster Science 287: 2185–2195. [DOI] [PubMed] [Google Scholar]
- Akpinar N, Bilaloglu R. 1997. Cytological investigations of certain species of Vicia L. Turkish Journal of Botany 21: 197–207. [Google Scholar]
- Antonius K, Ahokas H. 1996. Flow cytometric determination of polyploidy level in spontaneous clones of strawberries. Hereditas 124: 285. [Google Scholar]
- APG II. 2003. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants. Botanical Journal of the Linnean Society 141: 399–436. [Google Scholar]
- Arabidopsis Genome Initiative. 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana Nature 408: 796–815. [DOI] [PubMed] [Google Scholar]
- Arumuganathan K, Earle ED. 1991. Nuclear DNA content of some important plant species. Plant Molecular Biology Reporter 9: 208–218. [Google Scholar]
- Asif MJ, Mak C, Othman RY. 2001. Characterization of indigenous Musa species based on flow cytometric analysis of ploidy and nuclear DNA content. Caryologia 54: 161–168. [Google Scholar]
- Ayonoadu UW. 1974. Nuclear DNA variation in Phaseolus Chromosoma 48: 41–49. [Google Scholar]
- Babcock EB. 1947. The genus Crepis II. Systematic treatment. University of California Publications in Agricultural Science 22: 199–1030. [Google Scholar]
- Baranyi M, Greilhuber J, Swiecicki WK. 1996. Genome size in wild Pisum species. Theoretical and Applied Genetics 93: 717–721. [DOI] [PubMed] [Google Scholar]
- Beaton MJ. 1988.Genome size variation in the Cladocera. M.Sc. thesis, University of Windsor, Windsor, Ontario. [Google Scholar]
- Belletti P, Marzachì C, Lanteri S. 1998. Flow cytometric measurement of nuclear DNA content in Capsicum (Solanaceae). Plant Systematics and Evolution 209: 85–91. [Google Scholar]
- Bennett MD. 1972. Nuclear DNA content and minimum generation time in herbaceous plants. Proceedings of the Royal Society of London Series B, Biological Sciences 181: 109–135. [DOI] [PubMed] [Google Scholar]
- Bennett MD. 1998. Plant genome values: How much do we know? Proceedings of the National Academy of Sciences of the USA 95: 2011–2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett MD, Bhandol P, Leitch IJ. 2000. Nuclear DNA amounts in angiosperms and their modern uses – 807 new estimates. Annals of Botany 86: 859–909. [Google Scholar]
- Bennett MD, Leitch IJ. 1995. Nuclear DNA amounts in angiosperms. Annals of Botany 76: 113–176. [Google Scholar]
- Bennett MD, Leitch IJ. 1997. Nuclear DNA amounts in angiosperms—583 new estimates. Annals of Botany 80: 169–196. [Google Scholar]
- Bennett MD, Leitch IJ. 2003. Angiosperm DNA C-values database (release 4·0, Jan. 2003). http://www.rbgkew.org.uk/cval/homepage.html [Google Scholar]
- Bennett MD, Leitch IJ. 2005. Plant genome size research—a field in focus. Annals of Botany 95: 1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett MD, Leitch IJ, Hanson L. 1998. DNA amounts in two samples of angiosperm weeds. Annals of Botany 82: 121–134. [Google Scholar]
- Bennett MD, Leitch IJ, Price HJ, Johnston JS. 2003. Comparisons with Caenorhabditis (∼100 Mb) and Drosophila (∼175 Mb) using flow cytometry show genome size in Arabidopsis to be ∼157 Mb and thus 25 % larger than the Arabidopsis genome initiative estimate of ∼125 Mb. Annals of Botany 91: 547–557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett MD, Smith JB. 1976. Nuclear DNA amounts in angiosperms. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences 274: 227–274. [DOI] [PubMed] [Google Scholar]
- Bennett MD, Smith JB. 1991. Nuclear DNA amounts in angiosperms. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences 334: 309–345. [DOI] [PubMed] [Google Scholar]
- Bennett MD, Smith JB, Heslop-Harrison JS. 1982. Nuclear DNA amounts in angiosperms. Proceedings of the Royal Society of London Series B, Biological Sciences 216: 179–199. [DOI] [PubMed] [Google Scholar]
- Bennetzen J, Ma J, Devos KM. 2005. Mechanisms of recent genome size variation in flowering plants. Annals of Botany 95: 127–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blakesley D, Allen A, Pellny TK, Roberts AV. 2002. Natural and induced polyploidy in Acacia dealbata Link. and Acacia mangium Willd. Annals of Botany 90: 391–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanco A, Simeone R, Resta P, DePace C, Delre V, Caccia R, Mugnozza GTS, Frediani M, Cremonini R, Cionini PG. 1996. Genomic relationships between Dasypyrum villosum (L) Candargy and D. hordeaceum (Cosson et Durieu) Candargy. Genome 39: 83–92. [DOI] [PubMed] [Google Scholar]
- Bork P, Copley R. 2001. The draft sequences – filling in the gaps. Nature 409: 818–820. [DOI] [PubMed] [Google Scholar]
- Boscaiu M, Vicente O, Ehrendorfer F. 1999. Chromosome numbers, karyotypes and nuclear DNA contents from perennial polyploid groups of Cerastium (Caryophyllaceae). Plant Systematics and Evolution 218: 13–21. [Google Scholar]
- Bowers JE, Chapman BA, Rong JK, Paterson AH. 2003. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 422: 433–438. [DOI] [PubMed] [Google Scholar]
- Brandizzi F, Grilli Caiola M. 1996. Quantitative DNA analysis in different Crocus species (Iridaceae) by means of flow cytometry. Giornale Botanico Italiano 130: 643–645. [Google Scholar]
- Brenner S. 2000. The end of the beginning. Science 287: 2173–2174. [DOI] [PubMed] [Google Scholar]
- Bukhari YM. 1997. Nuclear DNA amounts in Acacia and Prosopis (Mimosaceae) and their evolutionary implications. Hereditas 126: 45–51. [Google Scholar]
- C. elegans Sequencing Consortium. 1998. Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 282: 2012–2018. [DOI] [PubMed] [Google Scholar]
- Cavalier-Smith T. 1985. Eukaryotic gene numbers, non-coding DNA and genome size. In: Cavalier-Smith T, ed. The evolution of genome size. Chichester, UK: John Wiley & Sons Ltd, 69–103. [Google Scholar]
- Chaudhuri D, Sen S. 2001. Genotypic diversity in species of Scilla Cytobios 104: 75–81. [PubMed] [Google Scholar]
- Check E. 2002. Venter aims for maximum impact with minimal genome. Nature 420: 350. [DOI] [PubMed] [Google Scholar]
- Chooi WY. 1971. Variation in nuclear DNA content in genus Vicia Genetics 68: 195–211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung J, Lee JH, Arumuganathan K, Graef GL, Specht JE. 1998. Relationships between nuclear DNA content and seed and leaf size in soybean. Theoretical and Applied Genetics 96: 1064–1068. [Google Scholar]
- Cremonini R, Funari S, Mazzuca S. 1992. Cytology of Vicia species: nuclear structure, karyological analysis and DNA content. Chromatin 1: 135–146. [Google Scholar]
- Devos KM, Gale MD. 1997. Comparative genetics in the grasses. Plant Molecular Biology 35: 3–15. [PubMed] [Google Scholar]
- Dimitrova D, Greilhuber J. 2000. Karyotype and DNA content evolution in ten species of Crepis (Asteraceae) distributed in Bulgaria. Botanical Journal of the Linnean Society 132: 281–297. [Google Scholar]
- Doležel J, Bartos J. 2005. Plant DNA flow cytometry and estimation of nuclear genome size. Annals of Botany 95: 99–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doležel J, Doleželova M, Novak FJ. 1994. Flow cytometric estimation of nuclear DNA amount in diploid bananas (Musa acuminata and M. balbisiana). Biologia Plantarum 36: 351–357. [Google Scholar]
- Doležel J, Greilhuber J, Lucretti S, Meister A, Lysak MA, Nardi L, Obermayer R. 1998. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Annals of Botany 82 (Suppl. A): 17–26. [Google Scholar]
- Doležel J, Sgorbati S, Lucretti S. 1992. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiologia Plantarum 85: 625–631. [Google Scholar]
- Emshwiller E. 2002. Ploidy levels among species in the ‘Oxalis tuberosa Alliance’ as inferred by flow cytometry. Annals of Botany 89: 741–753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis DM, Hulbert SH, Michelmore RW. 1990. Genome size and complexity of the obligate fungal pathogen, Bremia lactucae Experimental Mycology 14: 299–309. [Google Scholar]
- Frediani M, Sassoli O, Cremonini R. 1992. Nuclear DNA characterization of two species of Vicia – Vicia bithynica L. and Vicia narbonensis L. Biologia Plantarum 34: 335–344. [Google Scholar]
- Galbraith DW, Harkins KR, Knapp S. 1991. Systemic endopolyploidy in Arabidopsis thaliana Plant Physiology 96: 985–989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP, Firoozabady E. 1983. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 220: 1049–1051. [DOI] [PubMed] [Google Scholar]
- Gammar ZG, Puech S, Zouaghi M. 1999. Flow cytometry DNA assay of Mediterranean lupins. Candollea 54: 45–56. [Google Scholar]
- Godelle B, Cartier D, Marie D, Brown SC, Siljakyakovlev S. 1993. Heterochromatin study demonstrating the non-linearity of fluorometry useful for calculating genomic base composition. Cytometry 14: 618–626. [DOI] [PubMed] [Google Scholar]
- Goff SA, Ricke D, Lan TH, Presting G, Wang RL et al. 2002. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 296: 92–100. [DOI] [PubMed] [Google Scholar]
- Govaerts R. 2004. The Monocot Checklist Project. Taxon 53: 144–146. [Google Scholar]
- Graham MJ, Nickell CD, Rayburn AL. 1994. Relationship between genome size and maturity group in soybean. Theoretical and Applied Genetics 88: 429–432. [DOI] [PubMed] [Google Scholar]
- Gregory TR, Hebert PDN, Kolasa J. 2000. Evolutionary implications of the relationship between genome size and body size in flatworms and copepods. Heredity 84: 201–208. [DOI] [PubMed] [Google Scholar]
- Greilhuber J. 1979. Evolutionary changes of DNA and heterochromatin amounts in the Scilla bifolia group (Liliaceae). Plant Systematics and Evolution Suppl. 2: 263–280. [Google Scholar]
- Greilhuber J. 1998. Intraspecific variation in genome size: A critical reassessment. Annals of Botany 82 (Suppl. A): 27–35. [Google Scholar]
- Greilhuber J. 2005. Intraspecific variation in genome size in angiosperms— identifying its existence. Annals of Botany 95: 91–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greilhuber J, Doležel J, Lysak MA, Bennett MD. 2005. The origin, evolution and proposed stabilization of the terms ‘Genome Size’ and ‘C-Value’ to describe nuclear DNA contents. Annals of Botany 95: 255–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greilhuber J, Ebert I. 1994. Genome size variation in Pisum sativum Genome 37: 646–655. [DOI] [PubMed] [Google Scholar]
- Greilhuber J, Ebert I, Lorenz A, Vyskot B. 2000. Origin of facultative heterochromatin in the endosperm of Gagea lutea (Liliaceae). Protoplasma 212: 217–226. [Google Scholar]
- Greilhuber J, Obermayer R. 1997. Genome size and maturity group in Glycine max (soybean). Heredity 78: 547–551. [Google Scholar]
- Greilhuber J, Speta F. 1985. Geographical variation of genome size at low taxonomic levels in the Scilla bifolia alliance (Hyacinthaceae). Flora 176: 431–438. [Google Scholar]
- Greuter W, Barrie FR, Burdet HM, Chaloner WG, Demoulin V, Hawksworth DL, Jørgensen PM, Nicolson DH, Silva PC, PTrehane P, McNeil J. 1994. International code of botanical momenclature (Tokyo Code) adopten by the Fifteenth International Botanical Congress, Yokohama, August-September 1993. Regnum Vegetabile 131. [Google Scholar]
- Hanson L, Brown RL, Boyd A, Johnson MAT, Bennett MD. 2003. First nuclear DNA C-values for 28 angiosperm genera. Annals of Botany 91: 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson L, McMahon KA, Johnson MAT, Bennett MD. 2001. First nuclear DNA C-values for 25 angiosperm families. Annals of Botany 87: 251–258. [DOI] [PubMed] [Google Scholar]
- Hanson L, McMahon KA, Johnson MAT, Bennett MD. 2001. First nuclear DNA C-values for another 25 angiosperm families. Annals of Botany 88: 851–858. [DOI] [PubMed] [Google Scholar]
- Hardie DC, Gregory TR, Hebert PDN. 2002. From pixels to picograms: A beginners' guide to genome quantification by Feulgen image analysis densitometry. Journal of Histochemistry and Cytochemistry 50: 735–749. [DOI] [PubMed] [Google Scholar]
- Hart H. 1991. Evolution and classification of the European Sedum species (Crassulaceae). Flora Mediterranea 1: 31–61. [Google Scholar]
- Hartman TPV, Jones J, Blackhall NW, Power JB, Cocking EC, Davey MR. 2000. Cytogenetics, molecular cytogenetics and genome size in Leucaena (Leguminosae, Mimosideae). In: Guttenberger H, Borzan Z, Schlarbaum SE, Hartman TPV, eds. Cytogenetic studies of forest trees and shrubs—review, present status, and outlook on the future. Zvolen, Slovakia: Arbora Publishers, 57–70. [Google Scholar]
- Hosouchi T, Kumekawa N, Tsuruoka H, Kotani H. 2002. Physical map-based sizes of the centromeric regions of Arabidopsis thaliana chromosomes 1, 2, and 3. DNA Research 9: 117–121. [DOI] [PubMed] [Google Scholar]
- Ingle J, Timmis JN, Sinclair J. 1975. Relationship between satellite deoxyribonucleic acid, ribosomal ribonucleic acid gene redundancy, and genome size in plants. Plant Physiology 55: 496–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joachimiak A, Kula A, Sliwinska E, Sobieszczanska A. 2001. C-banding and nuclear DNA amount in six Bromus species. Acta Biologica Cracoviensia Series Botanica 43: 105–115. [Google Scholar]
- Johnston JS, Bennett MD, Rayburn AL, Galbraith DW, Price HJ. 1999. Reference standards for determination of DNA content of plant nuclei. American Journal of Botany 86: 609–613. [PubMed] [Google Scholar]
- Johnston JS, Pepper AE, Hall AE, Chen ZJ, Hodnett G, Drabek J, Lopez R, Price HJ. 2005. Evolution of genome size in Brassicaceae. Annals of Botany 95: 229–235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jussieu AL. 1803. Sur le Petunia, genre nouveau de la famille des plantes solanees. Annals du Museum National d'Histoire Naturelle 2: 214–216. [Google Scholar]
- Kaneko T, Kotani H, Nakamura Y, Sato S, Asamizu E, Miyajima N, Tabata S. 1998. Structural analysis of Arabidopsis thaliana chromosome 5. V. Sequence features of the regions of 1,381,565 bp covered by twenty one physically assigned P1 and TAC clones. DNA Research 5: 131–145. [DOI] [PubMed] [Google Scholar]
- Kapraun DF. 2005. Nuclear DNA content estimates in multicellular eukaryotic green, red and brown algae: phylogenetic considerations. Annals of Botany 95: 7–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knight CA, Ackerly DD. 2002. Variation in nuclear DNA content across environmental gradients: a quantile regression analysis. Ecology Letters 5: 66–76. [Google Scholar]
- Koce JD, Vilhar B, Bohanec B, Dermastia M. 2003. Genome size of Adriatic seagrasses. Aquatic Botany 77: 17–25. [Google Scholar]
- König C, Ebert I, Greilhuber J. 1987. A DNA cytophotometric and chromosome banding study in Hedera helix (Araliaceae), with reference to differential DNA replication associated with juvenile-adult phase change. Genome 29: 498–503. [Google Scholar]
- Krisai R, Greilhuber J. 1997.Cochlearia pyrenaica DC., das Löffelkraut, in Oberöstererreich (mit Anmerkungen zur Karyologie und zur Genomgrösse. Beiträge zur Naturkunde Oberösterreichs 5: 151–160. [Google Scholar]
- Laurie DA, Bennett MD. 1985. Nuclear DNA content in the genera Zea and Sorghum – Intergeneric, interspecific and intraspecific variation. Heredity 55: 307–313. [Google Scholar]
- Lee JH, Yen Y, Arumuganathan K, Baenziger PS. 1997. The absolute DNA content of wheat monosomics at interphase measured by laser-based flow cytometry. Theoretical and Applied Genetics 95: 1300–1304. [Google Scholar]
- Leitch IJ, Bennett MD. 2002. The genome: integrating genomic characters for a holistic approach to understanding plant genomes. In: Baijnath H, Singh Y eds. Rebirth of science in Africa – a shared vision for life and environmental sciences. Pretoria: Umdaus Press, 39–50. [Google Scholar]
- Leitch IJ, Bennett MD. 2002. New insights into patterns of nuclear genome size evolution in plants. Current Genomics 3: 551–562. [Google Scholar]
- Leitch IJ, Bennett MD. 2004. Genome downsizing in polyploid plants. Biological Journal of the Linnean Society 82: 651–663. [Google Scholar]
- Leitch IJ, Hanson L. 2002. DNA C-values in seven families fill phylogenetic gaps in the basal angiosperms. Botanical Journal of the Linnean Society 140: 175–179. [Google Scholar]
- Leitch IJ, Soltis DE, Soltis PS, Bennett MD. 2005. Evolution of DNA amounts across land plants (Embryophyta). Annals of Botany 95: 207–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leutwiler LS, Hough-Evans BR, Meyerowitz EM. 1984. The DNA of Arabidopsis thaliana Molecular & General Genetics 194: 15–23. [Google Scholar]
- Lin S, Lee H-C, Chen W-H, Chen C-C, Kao Y-Y, Fu Y-M, Chen Y-H, Lin T-Y. 2001. Nuclear DNA contents of Phalaenopsis sp. and Doritis pulcherrima Journal of the American Society of Horticultural Science 126: 195–199. [Google Scholar]
- Lysák MA, Doležel J. 1998. Estimation of nuclear DNA content in Sesleria (Poaceae). Caryologia 52: 123–132. [Google Scholar]
- Marie D, Brown SC. 1993. A cytometric exercise in plant DNA histograms, with 2C values for 70 species. Biology of the Cell 78: 41–51. [DOI] [PubMed] [Google Scholar]
- Meyerowitz EM. 1994. Structure and organization of the Arabidopsis thaliana nuclear genome. In: Meyerowitz EM, Somerville CR, eds. Arabidopsis. New York: Cold Spring Harbor Laboratory Press, 21–36. [Google Scholar]
- Mishiba KI, Ando T, Mii M, Watanabe H, Kokubun H, Hashimoto G, Marchesi E. 2000. Nuclear DNA content as an index character discriminating taxa in the genus Petunia sensu Jussieu (Solanaceae). Annals of Botany 85: 665–673. [Google Scholar]
- Mowforth MAG. 1986.Variation in nuclear DNA amounts in flowering plants: an ecological analysis. Ph.D. thesis, University of Sheffield, Sheffield, UK. [Google Scholar]
- Mukherjee S, Sharma AK. 1993. Cytophotometric DNA estimation in Luzula species. Current Science 65: 987–989. [Google Scholar]
- Mukherjee S, Sharma AK. 1993.In situ nuclear DNA content in perennial fast and slow growing acacias from arid zones. Cytobios 75: 33–36. [Google Scholar]
- Mukherjee S, Sharma AK. 1995.In situ nuclear DNA varation in Australian species of Acacia Cytobios 83: 59–64. [Google Scholar]
- Nagl W, Frisch B, Frolich E. 1979. Extra DNA during floral induction. Plant Systematics and Evolution Suppl. 2: 111–118. [Google Scholar]
- Nagl W, Jeanjour M, Kling H, Kuhner S, Michels I, Muller T, Stein B. 1983. Genome and chromatin organization in higher plants. Biologisches Zentralblatt 102: 129–148. [Google Scholar]
- Nagl W, Treviranus A. 1995. A flow cytometric analysis of the nuclear 2C DNA content in 17 Phaseolus species (53 genotypes). Botanica Acta 108: 403–406. [Google Scholar]
- Nishikawa K, Furuta Y, Ishitoba K. 1984. Chromosomal evolution in the genus Carex as viewed from nuclear DNA content, with special reference to its aneuploidy. Japanese Journal of Genetics 59: 465–472. [Google Scholar]
- Noirot M, Barre P, Duperray C, Hamon S, De Kochko A. 2005. Investigation on the origins of stoichiometric error in genome size estimation using heat experiments. Consequneces on data interpretation. Annals of Botany 95: 111–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noirot M, Barre P, Louarn J, Duperray C, Hamon S. 2000. Nucleus-cytosol interactions – A source of stoichiometric error in flow cytometric estimation of nuclear DNA content in plants. Annals of Botany 86: 309–316. [Google Scholar]
- NSF. 1990.Document 90–80. A long range plan for the multinational coordinated Arabidopsis thaliana genome research project. Washington, DC: National Science Foundation. [Google Scholar]
- Obermayer R, Greilhuber J. 1999. Genome size in Chinese soybean accessions – stable or variable? Annals of Botany 84: 259–262. [Google Scholar]
- Obermayer R, Leitch IJ, Hanson L, Bennett MD. 2002. Nuclear DNA C-values in 30 species double the familial representation in pteridophytes. Annals of Botany 90: 209–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obermayer R, Swiecicki WK, Greilhuber J. 1999. Flow cytometric determination of genome size in some old world Lupinus species (Fabaceae). Plant Biology 1: 403–407. [Google Scholar]
- Ohri D. 2002. Genome size variation in some tropical hardwoods. Biologia Plantarum 45: 455–457. [Google Scholar]
- Ohri D, Singh PS. 2002. Karyotypic and genome size variation in Cajanus cajan (L.) Millsp. (pigeonpea) and some wild relatives. Genetic Resources and Crop Evolution 49: 1–10. [Google Scholar]
- Pennisi E. 2000. Ideas fly at gene-finding jamboree. Science 287: 2182–2184. [DOI] [PubMed] [Google Scholar]
- Punina EO, Alexandrova TV. 1992. The chromosome volume and relative DNA content in Caucasian representatives of the genus Paeonia Botanichesky Zhurnal 77: 16–23. [Google Scholar]
- Raina SN, Bisht MS. 1988. DNA amounts and chromatin compactness in Vicia Genetica 77: 65–77. [Google Scholar]
- Raina SN, Rees H. 1983. DNA variation between and within chromosome complements of Vicia species. Heredity 51: 335–346. [Google Scholar]
- Rayburn AL, Biradar DP, Bullock DG, Nelson RL, Gourmet C, Wetzel JB. 1997. Nuclear DNA content diversity in Chinese soybean introductions. Annals of Botany 80: 321–325. [Google Scholar]
- Rayburn AL, Price HJ, Smith JD, Gold JR. 1985. C-band heterochromatin and DNA content in Zea mays American Journal of Botany 72: 1610–1617. [Google Scholar]
- Redi CA, Garagna S. 1987. Cytochemical evaluation of the nuclear DNA content as a tool for taxonomical studies in eutardigrades. In: Bertolani R, ed. Biology of Tardigrades. Mucchi: Modena, 73–80. [Google Scholar]
- Redondo N, Horjales M, Brown S, Villaverde C. 1996. Biometric and cytometric study of nuclear DNA within Saxifraga granulata L. Boletin da Sociedade Broteriana, Séries 2 67: 287–301. [Google Scholar]
- Rees H, Walters MR. 1965. Nuclear DNA and the evolution of wheat. Heredity 20: 73–82. [Google Scholar]
- Rejmanek M. 1996. A theory of seed plant invasiveness: the first sketch. Biological Conservation 78: 171–181. [Google Scholar]
- Rieger R, Michaelis A, Green MM. 1991.Glossary of genetics; classical and molecular, 5th edn. Berlin: Springer-Verlag. [Google Scholar]
- Robinson JG. 2004. The natural history of Madagascar. Science 304: 53–53. [Google Scholar]
- Rosato M, Chiavarino AM, Naranjo CA, Hernandez JC, Poggio L. 1998. Genome size and numerical polymorphism for the B chromosome in races of maize (Zea mays ssp. mays, Poaceae). American Journal of Botany 85: 168–174. [PubMed] [Google Scholar]
- Ruthmann A, Wenderoth H. 1975. Der DNA-Gehalt der Zellen bei dem primitiven Metazoon Trichoplax adhaerens F.E. Schulze. Cytobiologie 10: 421–431. [Google Scholar]
- Sakamoto K, Akiyama Y, Fukui K, Kamada H, Satoh S. 1998. Characterization: genome sizes and morphology of sex chromosomes in Hemp (Cannabis sativa L.). Cytologia 63: 459–464. [Google Scholar]
- Seo HC, Kube M, Edvardsen RB, Jensen MF, Beck A, Spriet E, Gorsky G, Thompson EM, Lehrach H, Reinhardt R, Chourrout D. 2001. Miniature genome in the marine chordate Oikopleura dioica Science 294: 2506–2506. [DOI] [PubMed] [Google Scholar]
- Shi Y. 1991.Molecular studies of the evolutionary relationships of Brachpodium (Poaceae). Ph.D., University of Leicester, UK. [Google Scholar]
- Shi Y, Draper D, Stace C. 1993. Ribosomal DNA variation and its phylogenetic implication in the genus Brachypodium (Poaceae). Plant Systematics and Evolution 188: 125–138. [Google Scholar]
- Siroký J, Lysák MA, Doležel J, Kejnovsky E, Vyskot B. 2001. Heterogeneity of rDNA distribution and genome size in Silene spp. Chromosome Research 9: 387–393. [DOI] [PubMed] [Google Scholar]
- Soldi R, Ramella L, Gambi MC, Sordino P, G. S. 1994. Genome size in polychaetes: relationship with body length and life habit. In: Dauvin J-C, Laubier L, Reish DJ, eds. Actes de la 4ième Conférence internationale des Polychètes, Museum national de Histoire naturelle, 129–135. [Google Scholar]
- Somerville C, Somerville S. 1999. Plant functional genomics. Science 285: 380–383. [DOI] [PubMed] [Google Scholar]
- Swift H. 1950. The constancy of desoxyribose nucleic acid in plant nuclei. Proceedings of the National Academy of Sciences of the USA 36: 643–654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swift H. 1953. The quantitative aspects of nuclear nucleoproteins. International Review of Cytology 2: 1–76. [Google Scholar]
- Taliaferro CM, Hopkins AA, Henthorn JC, Murphy CD, Edwards RM. 1997. Use of flow cytometry to estimate ploidy level in Cynodon species. International Turfgrass Society Research Journal 8: 385–392. [Google Scholar]
- Temsch EM, Greilhuber J. 2000. Genome size variation in Arachis hypogaea and A. monticola re-evaluated. Genome 43: 449–451. [PubMed] [Google Scholar]
- Temsch EM, Greilhuber J, Krisai R. 1998. Genome size in Sphagnum (peat moss). Botanica Acta 111: 325–330. [Google Scholar]
- Thibault J. 1998. Nuclear DNA amount in pure species and hybrid willows (Salix): a flow cytometric investigation. Canadian Journal of Botany 76: 157–165. [Google Scholar]
- Tiersch TR, Chandler RW, Wachtel SS, Elias S. 1989. Reference standards for flow cytometry and application in comparative studies of nuclear DNA content. Cytometry 10: 706–710. [DOI] [PubMed] [Google Scholar]
- Unal F, Callow RS. 1995. Estimation of genome size by Feulgen photometry. Turkish Journal of Botany 19: 523–529. [Google Scholar]
- United Nations Environment Programme. 1992.Convention on biological diversity. Nairobi, Kenya: United Nations Environment Programme. [Google Scholar]
- Van't Hof J. 1965. Relationships between mitotic cycle duration S period duration and average rate of DNA synthesis in root meristem cells of several plants. Experimental Cell Research 39: 48–58. [DOI] [PubMed] [Google Scholar]
- Vidic T, Greilhuber J, Vilhar B. 2003. Genome size is associated with differential survival of plant species. Abstracts of the Second Plant Genome Size Meeting held at The Royal Botanic Gardens, Kew, September 2003. [Google Scholar]
- Vilhar B, Greilhuber J, Koce JD, Temsch EM, Dermastia M. 2001. Plant genome size measurement with DNA image cytometry. Annals of Botany 87: 719–728. [Google Scholar]
- Vinogradov AE. 2003. Selfish DNA is maladaptive: evidence from the plant Red List. Trends in Genetics 19: 609–614. [DOI] [PubMed] [Google Scholar]
- Voglmayr H. 2000. Nuclear DNA amounts in mosses (Musci). Annals of Botany 85: 531–546. [Google Scholar]
- Wendel JF. 2000. Genome evolution in polyploids. Plant Molecular Biology 42: 225–249. [PubMed] [Google Scholar]
- Wendel JF, Cronn RC, Johnston JS, Price HJ. 2002. Feast and famine in plant genomes. Genetica 115: 37–47. [DOI] [PubMed] [Google Scholar]
- White J, Rees H. 1985. The chromosome cytology of a somatic hybrid Petunia Heredity 55: 53–59. [Google Scholar]
- White J, Rees H. 1987. Chromosome weights and measures in Petunia Heredity 58: 139–143. [Google Scholar]
- Wijsman HJW. 1990. On the interrelationships of certain species of Petunia 6. New names for the species of Calibrachoa formerly included into Petunia (Solanaceae). Acta Botanica Neerlandica 39: 101–102. [Google Scholar]
- Winkler H. 1920.Verbreitung und Ursache der Parthenogenese im Pflanzen- und Tierreiche. Jena: Fischer. [Google Scholar]
- Yamamoto M-T, Mitchelson A, Tudor M, Ohare K, Davies JA, Miklos GLG. 1990. Molecular and cytogenetic analysis of the heterochromatin-euchromatin junction region of the Drosophila melanogaster X-chromosome using cloned DNA sequences. Genetics 125: 821–832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokoya K, Roberts AV, Mottley J, Lewis R, Brandham PE. 2000. Nuclear DNA amounts in roses. Annals of Botany 85: 557–562. [Google Scholar]
- Yu J, Hu SN, Wang J, Wong GKS, Li SG et al. 2002. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 296: 79–92. [DOI] [PubMed] [Google Scholar]
- Zhang X, Wessler SR. 2004. Genome-wide comparative analysis of the transposable elements in the related species Arabidopsis thaliana and Brassica oleracea Proceedings of the National Academy of Sciences USA 101: 5589–5594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zonneveld BJM. 2001. Nuclear DNA contents of all species of Helleborus (Ranunculaceae) discriminate between species and sectional divisions. Plant Systematics and Evolution 229: 125–130. [Google Scholar]
- Zonneveld BJM. 2002. Genome size analysis of selected species of Aloe (Aloaceae) reveals the most primitive species and results in some new combinations. Bradleya 20: 5–12. [Google Scholar]
- Zonneveld BJM, Van Iren F. 2000. Flow cytometric analysis of DNA content in Hosta reveals ploidy chimeras. Euphytica 111: 105–110. [Google Scholar]
- Zonneveld BJM, Van Iren F. 2001. Genome size and pollen viability as taxonomic criteria: Application to the genus Hosta Plant Biology 3: 176–185. [Google Scholar]